Vitamin D Modulates the Response of Bronchial Epithelial Cells Exposed to Cigarette Smoke Extract

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.1.1. The 16HBE Cell Line
2.1.2. Primary Bronchial Epithelial Cells
2.2. Preparation Cigarette Smoke Extract
2.3. CSE and Vitamin D Exposure
2.4. IL-8 ELISA
2.5. qPCR
2.6. Wound Healing Assay
2.7. Statistics
3. Results
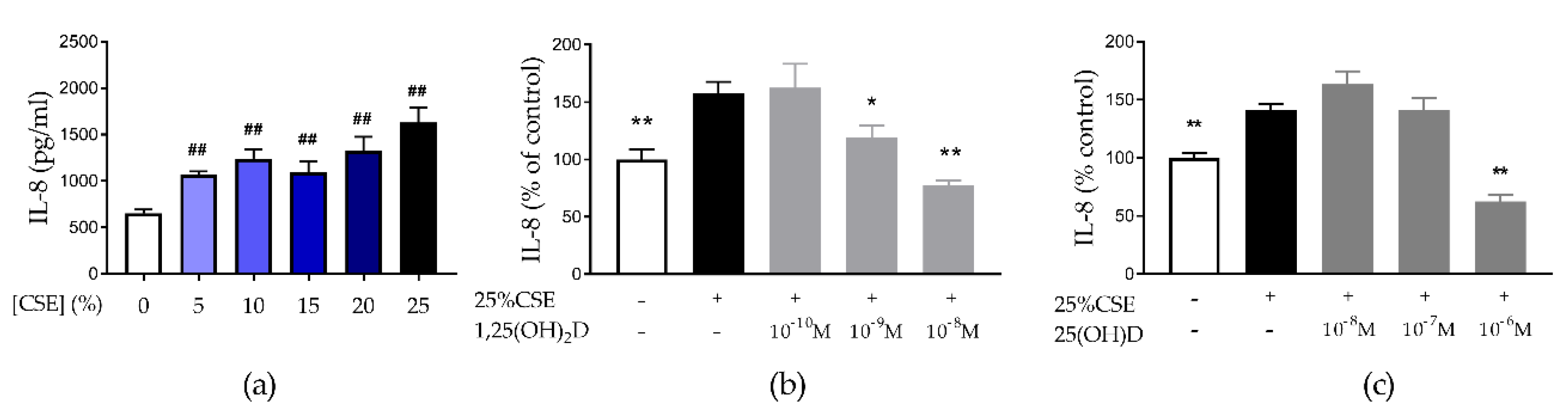
3.1. Both 1,25(OH)2D and 25(OH)D Reduce CSE-Induced IL-8 by 16HBE Cells
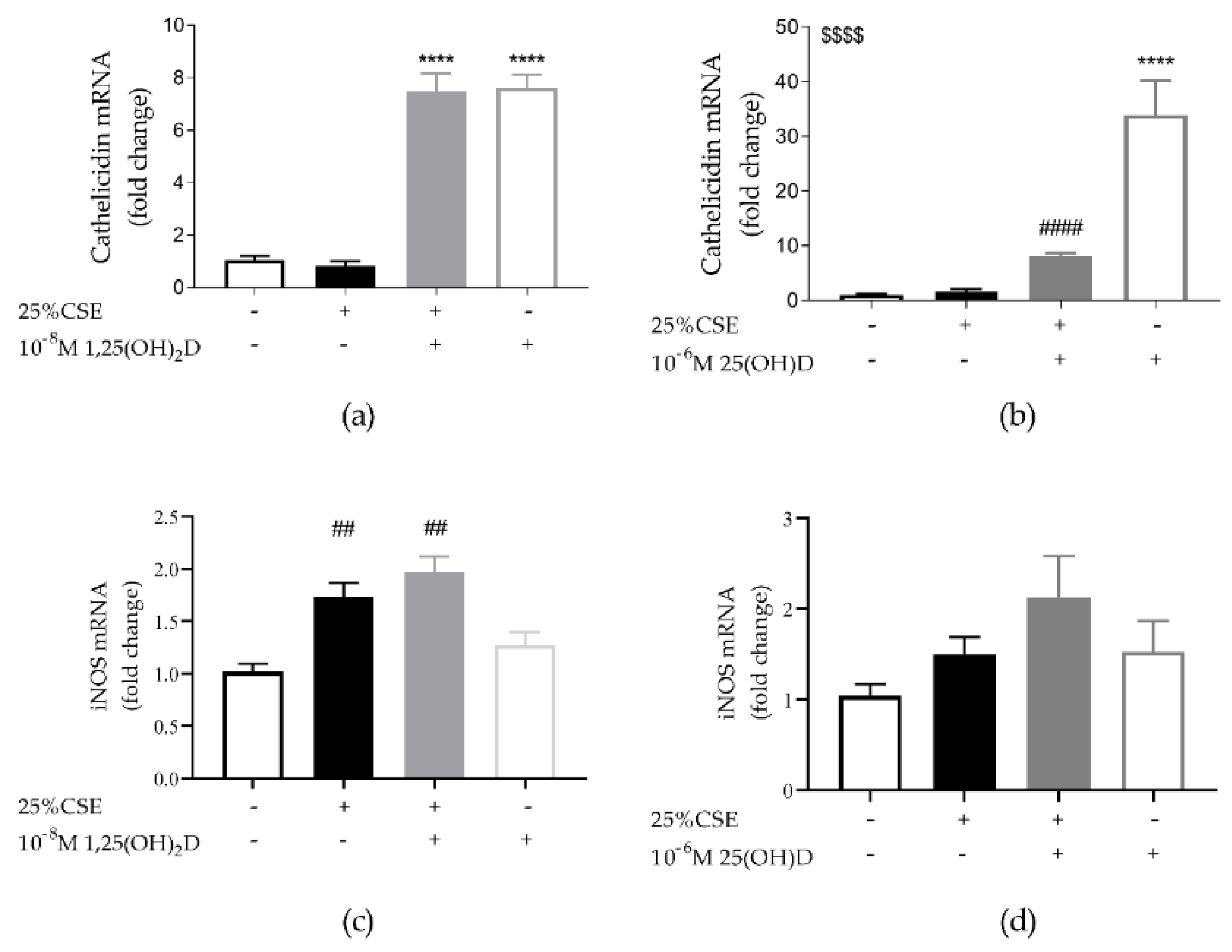
3.2. Both 1,25(OH)2D and 25(OH)D are Able to Induce Cathelicidin Expression in 16HBE Cells
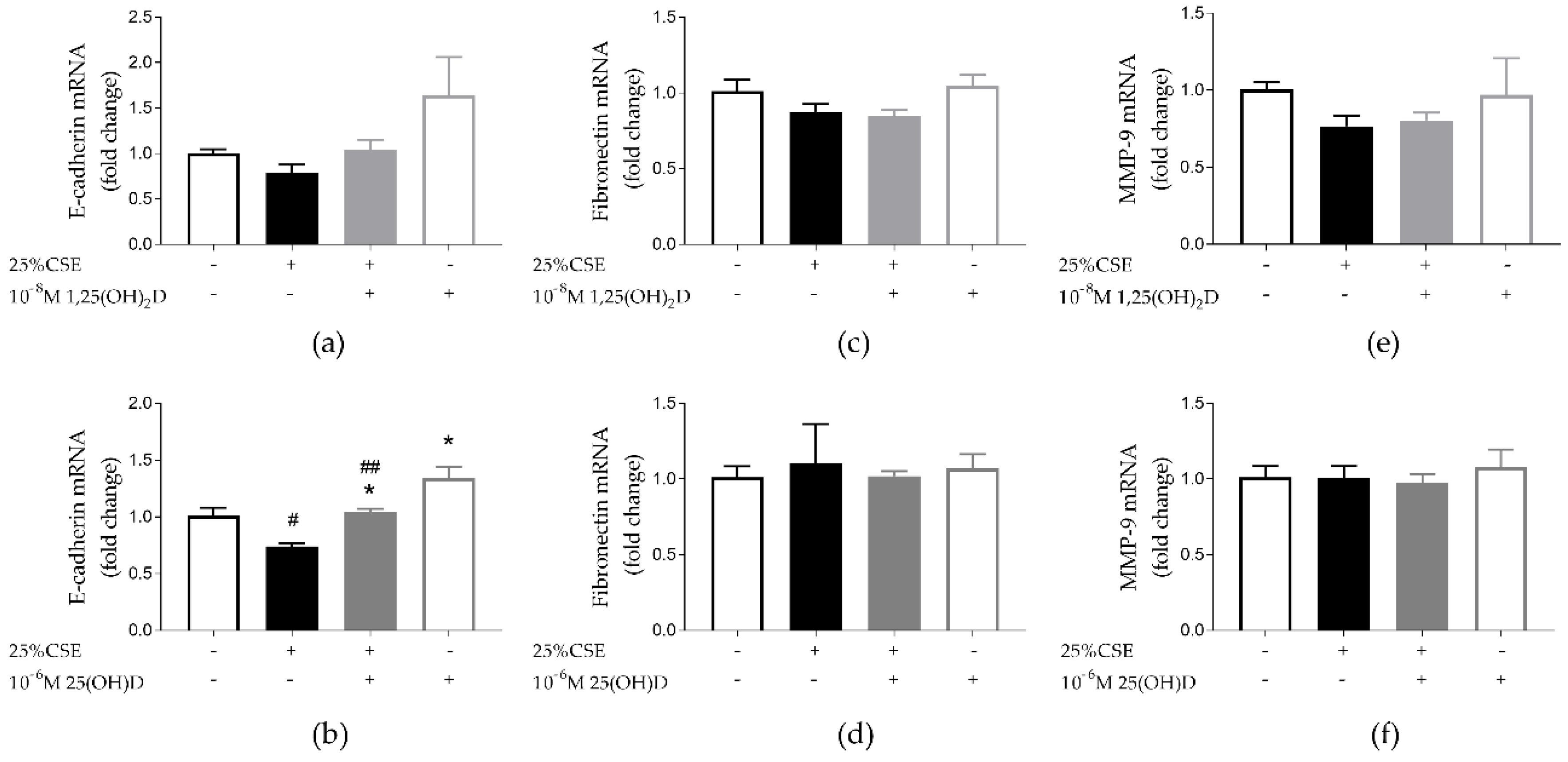
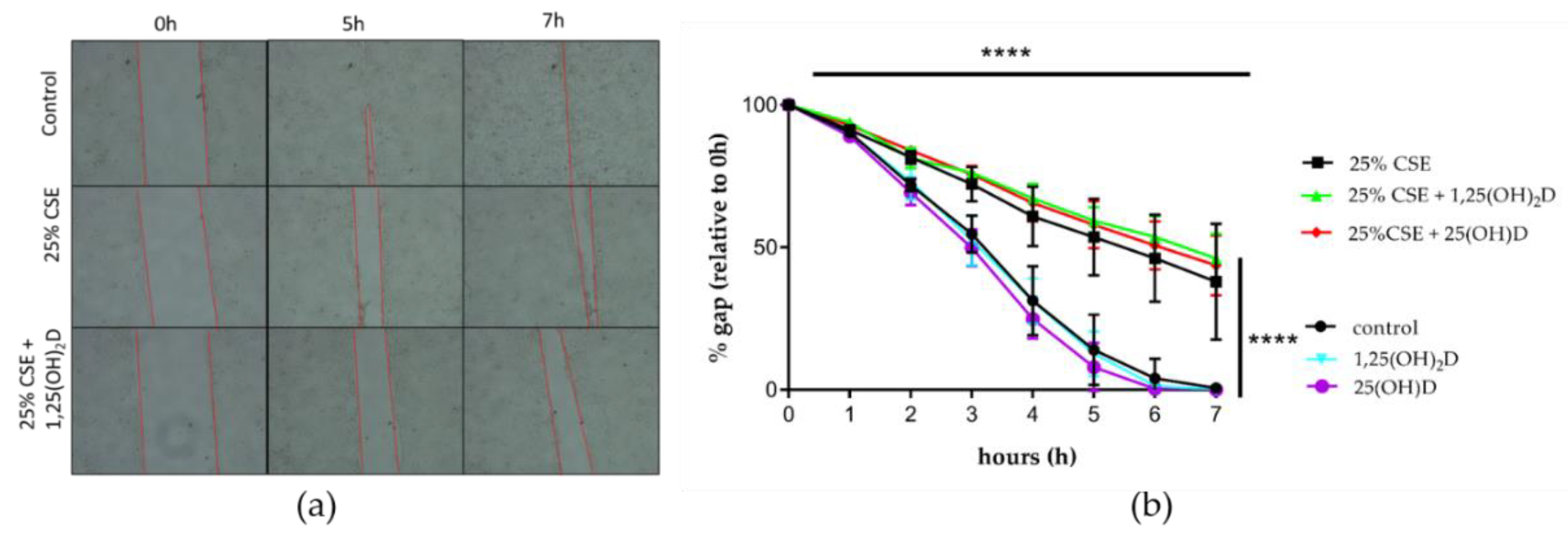
3.3. Effect of CSE and Vitamin D on Airway Remodeling and Wound Healing in 16HBE Cells
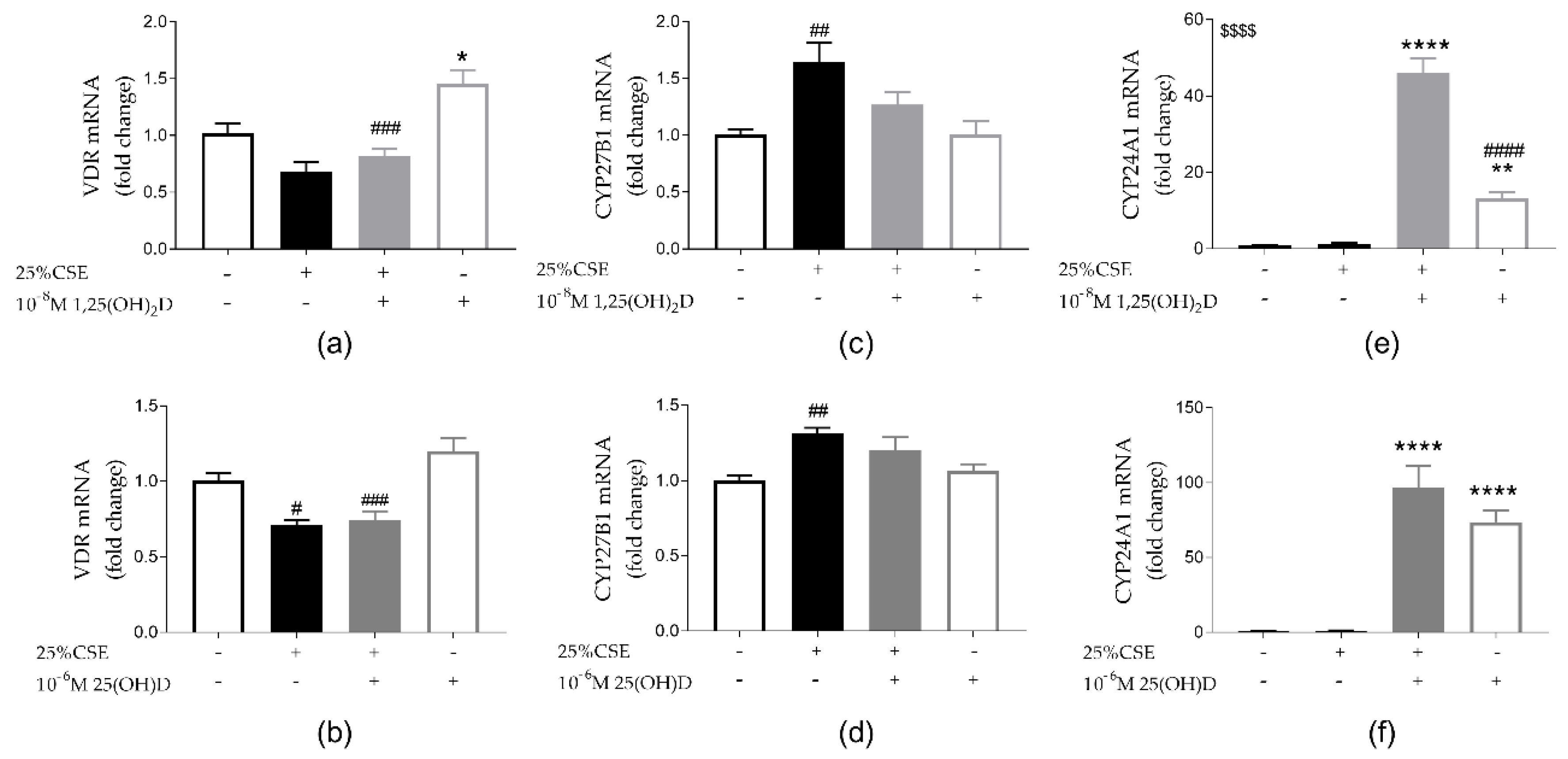
3.4. Effect of CSE and Vitamin D on Vitamin D Metabolism
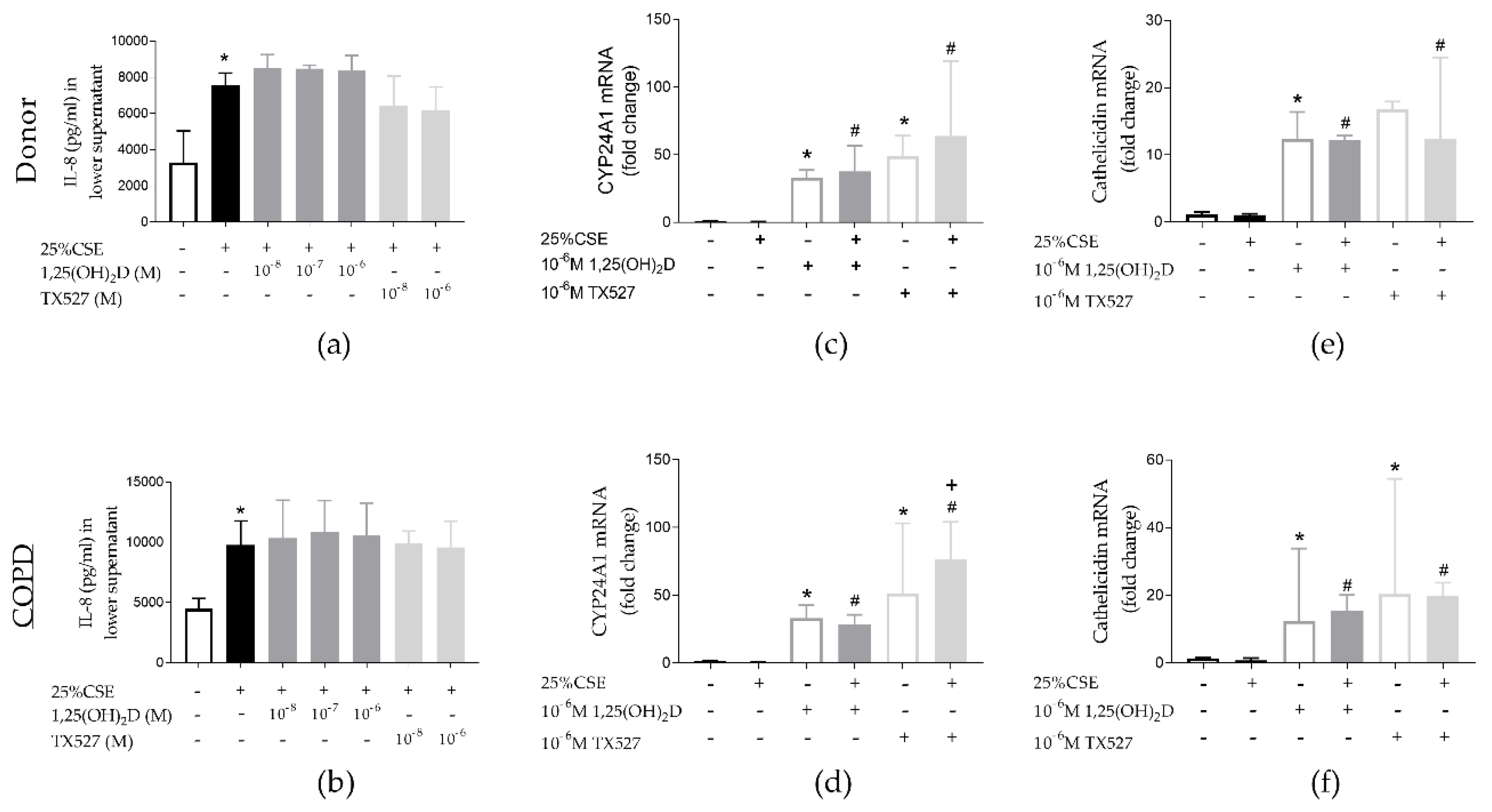
3.5. Effect of CSE and Vitamin D on PBEC from Unused Donor Lungs and COPD Explant Lungs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Global Initiative for Chronic Obstructive Lung Disease; Gold Report. Fontana, CA, USA, 2019; Volume 2019, pp. 1–155. Available online: https://goldcopd.org/ (accessed on 15 July 2019).
- Gao, W.; Li, L.; Wang, Y.; Zhang, S.; Adcock, I.M.; Barnes, P.J.; Huang, M.; Yao, X. Bronchial epithelial cells: The key effector cells in the pathogenesis of chronic obstructive pulmonary disease? Respirology 2015, 20, 722–729. [Google Scholar] [CrossRef]
- Phillips, J.; Kluss, B.; Richter, A.; Massey, E.D. Exposure of bronchial epithelial cells to whole cigarette smoke: Assessment of cellular responses. ATLA Altern. Lab. Anim. 2005, 33, 239–248. [Google Scholar] [CrossRef]
- Heulens, N.; Korf, H.; Janssens, W. Innate immune modulation in chronic obstructive pulmonary disease: Moving closer toward vitamin D therapy. J. Pharmacol. Exp. Ther. 2015, 353, 360–368. [Google Scholar] [CrossRef]
- Grzela, K.; Litwiniuk, M.; Zagorska, W.; Grzela, T. Airway Remodeling in Chronic Obstructive Pulmonary Disease and Asthma: The Role of Matrix Metalloproteinase-9. Arch. Immunol. Ther. Exp. 2016, 64, 47–55. [Google Scholar] [CrossRef]
- Amatngalim, G.D.; Broekman, W.; Daniel, N.M.; van der Vlugt, L.; van Schadewijk, A.; Taube, C.; Hiemstra, P. Cigarette smoke modulates repair and innate immunity following injury to airway epithelial cells. PLoS ONE 2016, 11, e0166255. [Google Scholar] [CrossRef]
- Baeke, F.; Takiishi, T.; Korf, H.; Gyseman, C.; Mathieu, C. Vitamin D: Modulator of the immune system. Curr. Opin. Pharmacol. 2010, 10, 482–496. [Google Scholar] [CrossRef]
- Janssens, W.; Lehouck, A.; Carremans, C.; Bouillon, R.; Mathieu, C.; Decramer, M. Pulmonary Perspective Vitamin D Beyond Bones in Chronic Obstructive Pulmonary Disease Time to Act. Am. J. Respir. Crit. Care Med. 2009, 179, 630–636. [Google Scholar] [CrossRef]
- Hansdottir, S.; Monick, M.M.; Lovan, N.; Powers, L.; Gerke, A.; Hunninghake, G.W. Vitamin D Decreases Respiratory Syncytial Virus Induction of NF-κB-Linked Chemokines and Cytokines in Airway Epithelium While Maintaining the Antiviral State. J. Immunol. 2010, 184, 965–974. [Google Scholar] [CrossRef]
- Heulens, N.; Korf, H.; Mathyssen, C.; Everaerts, S.; De Smidt, E.; Dooms, C.; Yserbyt, J.; Gyseman, C.; Gayan-Ramirez, G.; Mathieu, C.; et al. 1,25-Dihydroxyvitamin D Modulates Antibacterial and Inflammatory Response in Human Cigarette Smoke-Exposed Macrophages. PLoS ONE 2016, 11, e0160482. [Google Scholar] [CrossRef]
- Hansdottir, S.; Monick, M.M.; Hinde, S.L.; Lovan, N.; Look, D.C.; Hunninghake, G.W. Respiratory Epithelial Cells Convert Inactive Vitamin D to Its Active Form: Potential Effects on Host Defense. J. Immunol. 2008, 181, 7090–7099. [Google Scholar] [CrossRef]
- Menezes, R.J.; Cheney, R.T.; Husain, A.; Tretiakova, M.; Loewen, G.; Johnson, C.S.; Jayaprakash, V.; Moysich, K.B.; Salgia, R.; Reid, M.E. Vitamin D receptor expression in normal, premalignant, and malignant human lung tissue. Cancer Epidemiol. Biomark. Prev. 2008, 17, 1104–1110. [Google Scholar] [CrossRef]
- Jolliffe, D.A.; Greenberg, L.; Hooper, R.L.; Mathyssen, C.; Rafiq, R.; de Jongh, R.T.; Camargo, C.A.; Griffiths, C.J.; Janssens, W.; Martineau, A.R. Vitamin D to prevent exacerbations of COPD: Systematic review and meta-analysis of individual participant data from randomised controlled trials. Thorax 2019, 74, 337–345. [Google Scholar] [CrossRef]
- Tebben, P.J.; Singh, R.J.; Kumar, R. Vitamin D-mediated hypercalcemia: Mechanisms, diagnosis, and treatment. Endocr. Rev. 2016, 37, 521–547. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of Image Analysis HHS Public Access. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Butler, A.; Walton, G.M.; Sapey, E. Neutrophilic Inflammation in the Pathogenesis of Chronic Obstructive Pulmonary Disease. COPD J. Chronic Obstr. Pulm. Dis. 2018, 15, 392–404. [Google Scholar] [CrossRef]
- Beeh, K.M.; Kornmann, O.; Buhl, R.; Culpitt, S.V.; Giembycz, M.A.; Barnes, P.J. Neutrophil chemotactic activity of sputum from patients with COPD: Role of interleukin 8 and leukotriene B4. Chest 2003, 123, 1240–1247. [Google Scholar] [CrossRef]
- Comer, D.M.; Elborn, J.S.; Ennis, M. Cigarette Smoke, Airway Epithelial Cells and Host Defence. Inflamm Cell Signal 2014, 1, e203. [Google Scholar]
- Mortaz, E.; Henricks, P.A.J.; Kraneveld, A.D.; Givi, M.E.; Folkerts, G. Cigarette smoke induces the release of CXCL-8 from human bronchial epithelial cells via TLRs and induction of the inflammasome. Biochim. Biophys. Acta Mol. Basis Dis. 2011, 1812, 1104–1110. [Google Scholar] [CrossRef]
- Kode, A.; Yang, S.R.; Rahman, I. Differential effects of cigarette smoke on oxidative stress and proinflammatory cytokine release in primary human airway epithelial cells and in a variety of transformed alveolar epithelial cells. Respir. Res. 2006, 7, 132. [Google Scholar] [CrossRef]
- Glader, P.; Möller, S.; Lilja, J.; Wieslander, E.; Löfdahl, C.-G.; von Wachenfeldt, K. Cigarette smoke extract modulates respiratory defence mechanisms through effects on T-cells and airway epithelial cells. Respir. Med. 2006, 100, 818–827. [Google Scholar] [CrossRef]
- Mulligan, J.K.; Nagel, W.; O’Connell, B.P.; Wentzel, J.; Atkinson, C.; Schlosser, R.J. Cigarette smoke exposure is associated with vitamin D3 deficiencies in patients with chronic rhinosinusitis. J. Allergy Clin. Immunol. 2014, 134, 342–349. [Google Scholar] [CrossRef]
- Takano, Y.; Mitsuhashi, H.; Ueno, K. 1α,25-Dihydroxyvitamin D₃ inhibits neutrophil recruitment in hamster model of acute lung injury. Steroids 2011, 76, 1305–1309. [Google Scholar] [CrossRef]
- Hiemstra, P.S.; Amatngalim, G.D.; Van Der Does, A.M.; Taube, C. Antimicrobial peptides and innate lung defenses: Role in infectious and noninfectious lung diseases and therapeutic applications. Chest 2016, 149, 545–551. [Google Scholar] [CrossRef]
- Amatngalim, G.D.; Schrumpf, J.A.; Henic, A.; Dronkers, E.; Verhoosel, R.M.; Soledad, S.R.; Haagsam, H.P.; Funetes, M.E.; Sridhar, S.; Aarbiou, J.; et al. Antibacterial Defense of Human Airway Epithelial Cells from Chronic Obstructive Pulmonary Disease Patients Induced by Acute Exposure to Nontypeable Haemophilus influenzae: Modulation by Cigarette Smoke. J. Innate Immun. 2017, 9, 359–374. [Google Scholar] [CrossRef]
- Wang, T.-T.; Nestel, F.P.; Bourdeau, V.; Nagai, Y.; Wang, Q.; Liao, J.; Tavera-Mendoza, L.; Lin, R.; Hanrahan, J.H.; Mader, S.; et al. Cutting Edge: 1,25-Dihydroxyvitamin D3 is a direct inducer of antimicrobial peptide gene expression. J. Immunol. 2004, 173, 2909–2912. [Google Scholar] [CrossRef]
- Liu, P.T.; Stenger, S.; Tang, D.H.; Modlin, R.L. Cutting edge: Vitamin D-mediated human antimicrobial activity against Mycobacterium tuberculosis is dependent on the induction of cathelicidin. J. Immunol. 2007, 179, 2060–2063. [Google Scholar] [CrossRef]
- Telcian, A.G.; Zdrenghea, M.T.; Edwards, M.R.; Laza-Stanca, V.; Mallia, P.; Johnston, S.L.; Stanciu, L.A. Vitamin D increases the antiviral activity of bronchial epithelial cells in vitro. Antivir. Res. 2017, 137, 93–101. [Google Scholar] [CrossRef]
- Rockett, K.A.; Brookes, R.; Udalova, I.; Vidal, V.; Hill, A.V.S.; Kwiatkowski, D. 1, 25-Dihydroxyvitamin D3 inducesnitric oxide synthase and surpresses growth of Mycobacterium tuberculosis in a human macrophage-like cell line. Infect. Immun. 1998, 66, 5314–5321. [Google Scholar]
- Serré, J.; Mathyssen, C.; Ajime, T.T.; Korf, H.; Maes, K.; Heulens, N.; Gysemans, C.; Mathieu, C.; Vanaudenaerde, B.; Janssens, W.; et al. Airway Infection with Nontypeable Haemophilus influenza is more rapidly eradicated in vitamin D deficient mice. J. Steroid Biochem. Mol. Biol. 2018. [Google Scholar]
- Hogg, J.; Chu, F.; Utokaparch, S.; Woods, R.; Elliott, W.M.; Buzatu, L.; Cherniack, R.M.; Rogers, R.M.; Sciurba, F.C.; Coxson, H.O.; et al. The Nature of Small-Airway Obstruction in Chronic Obsructive Pulmonary Disease. N. Engl. J. Med. 2004, 350, 2645–2653. [Google Scholar] [CrossRef]
- Thorley, A.J.; Tetley, T.D. Pulmonary epithelium, cigarette smoke, and chronic obstructive pulmonary disease. Int. J. Chron. Obstruct. Pulmon. Dis. 2007, 2, 409–428. [Google Scholar]
- Chung, K.F. The Role of Airway Smooth Muscle in the Pathogenesis of Airway Wall Remodeling in Chronic Obstructive Pulmonary Disease. Proc. Am. Thorac. Soc. 2005, 2, 347–354. [Google Scholar] [CrossRef]
- Schamberger, A.C.; Staab-Weijnitz, C.A.; Mise-Racek, N.; Eickelberg, O. Cigarette smoke alters primary human bronchial epithelial cell differentiation at the air-liquid interface. Sci. Rep. 2015, 5, 8163. [Google Scholar] [CrossRef]
- Eurlings, I.M.J.; Reynaert, N.L.; Van Den Beucken, T.; Gosker, H.R.; de Theije, C.C.; Verhamme, F.M.; Bracke, K.R.; Wouters, E.F.M.; Dentner, M.A. Cigarette smoke extract induces a phenotypic shift in epithelial cells; involvement of HIF1alpha in mesenchymal transition. PLoS ONE 2014, 9, e107757. [Google Scholar] [CrossRef]
- Milara, J.; Peiró, T.; Serrano, A.; Cortijo, J. Epithelial to mesenchymal transition is increased in patients with COPD and induced by cigarette smoke. Thorax 2013, 68, 410–420. [Google Scholar] [CrossRef]
- Russell, R.E.; Culpitt, S.V.; DeMatos, C.; Donnelly, L.; Smith, M.; Wiggins, J.; Barnes, P.J. Release and activity of matrix metalloproteinase-9 and tissue inhibitor of metalloproteinase-1 by alveolar macrophages from patients with chronic obstructive pulmonary disease. Am. J. Respir. Cell Mol. Biol. 2002, 26, 602–609. [Google Scholar] [CrossRef]
- Perotin, J.-M.; Adam, D.; Vella-Boucaud, J.; Delepine, G.; Sandu, S.; Jonvel, A.-C.; Prevost, A.; Berthiot, G.; Pison, C.; Lebargy, F.; et al. Delay of airway epithelial wound repair in COPD is associated with airflow obstruction severity. Respir. Res. 2014, 15, 151. [Google Scholar] [CrossRef]
- Silverstein, P. Smoking and wound healing. Am. J. Med. 1992, 93 (Suppl. 1), 22–24. [Google Scholar] [CrossRef]
- Heilborn, J.D.; Frohm Nilsson, M.; Kratz, G.; Weber, G.; Sørensen, O.; Borregaard, N.; Ståhle-Bäckdahl, M. The Cathelicidin Anti-Microbial Peptide LL-37 is Involved in Re-Epithelialization of Human Skin Wounds and is Lacking in Chronic Ulcer Epithelium. J. Investig. Dermatol. 2003, 120, 379–389. [Google Scholar] [CrossRef]
- Fisher, K.; Agrawal, D. Vitamin D regulating TGF-β induced epithelial-mesenchymal transition. Respir. Res. 2014, 15, 146. [Google Scholar] [CrossRef]
- Liu, P.T.; Stenger, S.; Li, H.; Wenzel, L.; Tan, B.H.; Krutzik, S.R.; Ochoa, M.T.; Schauber, J.; Wu, K.; Meinken, C.; et al. Toll-Like Receptor Triggering of a Vitamin-D-Mediated Human Antimbacterial Response. Science 2006, 311, 23–26. [Google Scholar] [CrossRef]
- Ishii, M.; Yamaguchi, Y.; Nakamura, T.; Akishita, M. The Vitamin D Receptors May Function as Antiinflammatory Effects in Patients with COPD. Chest 2015, 148, 690A. [Google Scholar] [CrossRef]
- Buonfiglio, L.G.V.; Cano, M.; Pezzulo, A.A.; Vangas Calderon, D.G.; Zabner, J.; Gerke, A.K.; Comellas, A.P. Effect of vitamin D 3 on the antimicrobial activity of human airway surface liquid: Preliminary results of a randomised placebo-controlled double- blind trial. Open Respir. Res. 2017, 4, e000211. [Google Scholar] [CrossRef]
- Pohl, C.; Hermanns, M.I.; Uboldi, C.; Bock, M.; Fuchs, S.; Dei-Anang, J.; Mayer, E.; Kehe, K.; Kummer, W.; Kirkpatrick, C.J. Barrier functions and paracellular integrity in human cell culture models of the proximal respiratory unit. Eur. J. Pharm. Biopharm. 2009, 72, 339–349. [Google Scholar] [CrossRef]
- Cozens, A.L.; Yezzi, M.J.; Kunzelmann, K.; Ohrui, T.; Chin, L.; Eng, K.; Finkbeiner, W.E.; Widdicombe, J.H.; Gruenert, D.C. CFfR Expression and Chloride Secretion in Polarized Immortal Human Bronchial Epithelial Cells. Am. J. Respir. Cell Mol. Bio. 1994, 10, 38–47. [Google Scholar] [CrossRef]
- Banerjee, B.; Kicic, A.; Musk, M.; Sutanto, E.N.; Stick, S.M.; Chambers, D.C. Successful establishment of primary small airway cell cultures in human lung transplantation.pdf. Respir. Res. 2009, 10, 99. [Google Scholar] [CrossRef][Green Version]
- Forbes, B. Human airway epithelial cell lines for in vitro drug transport and metabolism studies. Pharm. Sci. Technol. Today 2000, 3, 18–27. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| GAPDH | TGGTATCGTGGAAGGACTCA | CCAGTAGAGGCAGGGATGAT |
| VDR | GATTGGAGAAGCTGGACGAG | GTTCGTGTGAATGATGGTGGA |
| CYP27B1 | CGCACTGTCCCAAAGCTG | CGGAGCTTGGCAGACATC |
| CYP24A1 | GTGACCATCATCCTCCCAAA | AGTATCTGCCTCGTGTTGTATG |
| Cathelicidin | GGGCTCCTTTGACATCAGTT | AGCAGGGCAAATCTCTTGTT |
| iNOS | TTCAGTATCACAACCTCAGCAAG | TGGACCTGCAAGTTAAAATCCC |
| Fibronectin | AAACCAATTCTTGGAGGAGG | CCATAAAGGGCAACCAAGAG |
| E-cadherin | GAAGGTGACAGAGCCTCTGGAT | GATCGGTTACCGTGATCAAAATC |
| MMP-9 | GCACGACGTCTTCCAGTACC | CAGGATGTCATAGGTCACGTAGC |
| Donor | COPD | p-Value | |
|---|---|---|---|
| Age (years) | 57 (55–58) | 57 (53–60) | 0.873 |
| gender (M/F) | 1/3 | 1/3 | >0.999 |
| FEV1% | NA | 24.5 (21.0–25.8) | NA |
| FEV1%/FVC% | NA | 34 (24.75–30.25) | NA |
| FVC% | NA | 62.5 (56.5–64.0) | NA |
| DLco% | NA | 36 (27–39) | NA |
| Smoking history (Y/N/U) | 1/2/1 | 4/0/0 | |
| VitD supplementation (Y/N/U) | 0/0/4 | 3/1/0 | NA |
| Serum 25(OH)D pretransplant µg/L | NA | 27 (21–43) | NA |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mathyssen, C.; Serré, J.; Sacreas, A.; Everaerts, S.; Maes, K.; Verleden, S.; Verlinden, L.; Verstuyf, A.; Pilette, C.; Gayan-Ramirez, G.; et al. Vitamin D Modulates the Response of Bronchial Epithelial Cells Exposed to Cigarette Smoke Extract. Nutrients 2019, 11, 2138. https://doi.org/10.3390/nu11092138
Mathyssen C, Serré J, Sacreas A, Everaerts S, Maes K, Verleden S, Verlinden L, Verstuyf A, Pilette C, Gayan-Ramirez G, et al. Vitamin D Modulates the Response of Bronchial Epithelial Cells Exposed to Cigarette Smoke Extract. Nutrients. 2019; 11(9):2138. https://doi.org/10.3390/nu11092138
Chicago/Turabian StyleMathyssen, Carolien, Jef Serré, Annelore Sacreas, Stephanie Everaerts, Karen Maes, Stijn Verleden, Lieve Verlinden, Annemieke Verstuyf, Charles Pilette, Ghislaine Gayan-Ramirez, and et al. 2019. "Vitamin D Modulates the Response of Bronchial Epithelial Cells Exposed to Cigarette Smoke Extract" Nutrients 11, no. 9: 2138. https://doi.org/10.3390/nu11092138
APA StyleMathyssen, C., Serré, J., Sacreas, A., Everaerts, S., Maes, K., Verleden, S., Verlinden, L., Verstuyf, A., Pilette, C., Gayan-Ramirez, G., Vanaudenaerde, B., & Janssens, W. (2019). Vitamin D Modulates the Response of Bronchial Epithelial Cells Exposed to Cigarette Smoke Extract. Nutrients, 11(9), 2138. https://doi.org/10.3390/nu11092138