Phaseolus vulgaris L. Extract: Alpha-Amylase Inhibition against Metabolic Syndrome in Mice

 ,
,  ,
,  and
and 
Abstract

1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Induction of Metabolic Syndrome
2.3. Extract Preparation
2.4. Treatments
2.5. Body Weight and Food Intake
2.6. Collection of Blood and Analytical Methods
2.7. Glucose and Insulin Tolerance Tests
2.8. Behavioral Tests
2.9. Tissues Explant
2.10. Liver and Kidney Histopathology
2.11. Analysis of Lipid Deposition in the Arterial Wall
2.12. Lipid Peroxidation (Thiobarbituric Acid-Reactive Substances Assay)
2.13. Carbonylated Proteins
2.14. Western Blot Analysis
2.15. Statistical Analysis
3. Results
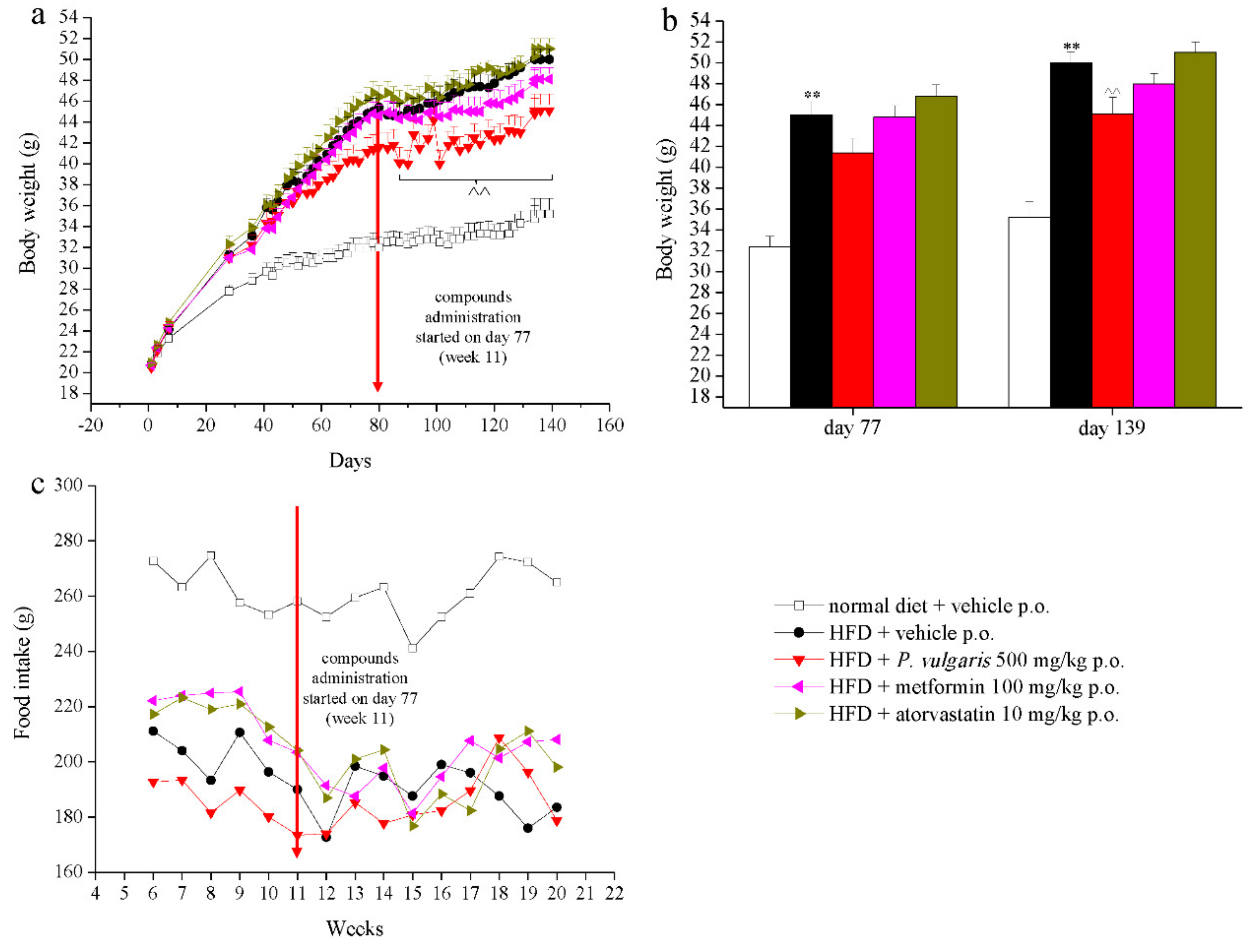
3.1. Treatments, Body Weight, and Food Intake
3.2. Hematic Metabolic Parameters
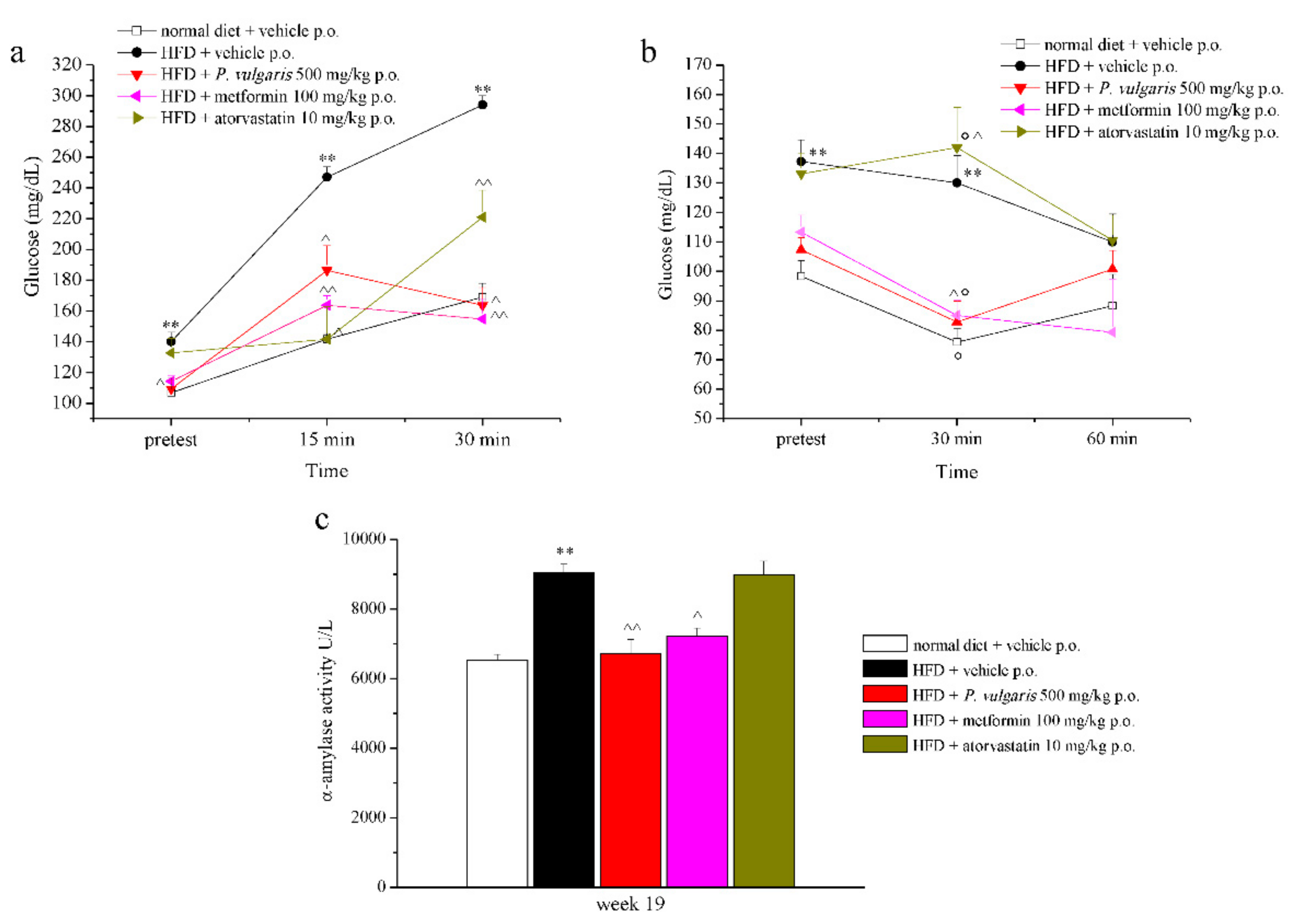
3.3. Glucose and Insulin Tolerance Test
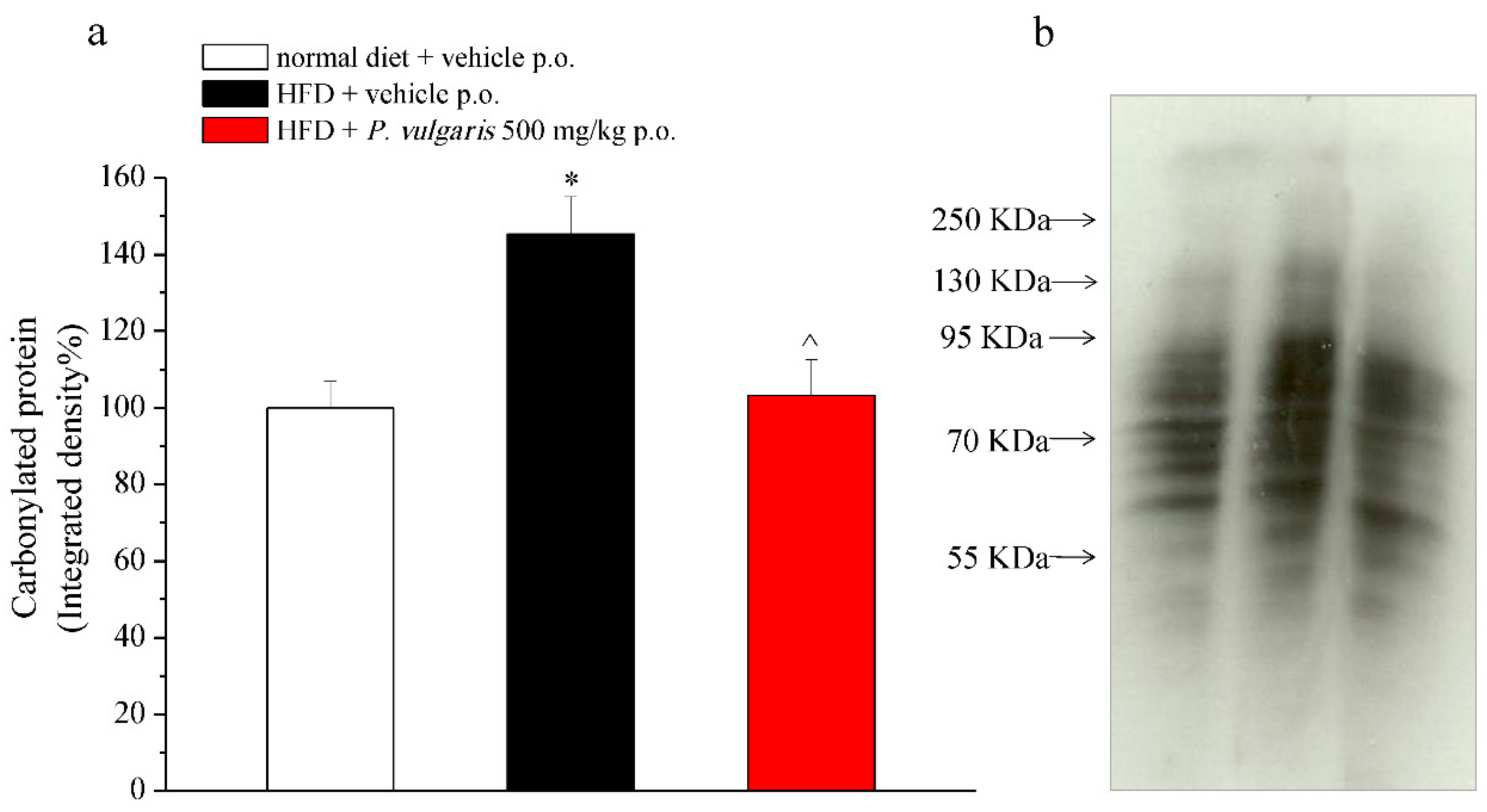
3.4. Plasma Enzymatic and Oxidative Parameters
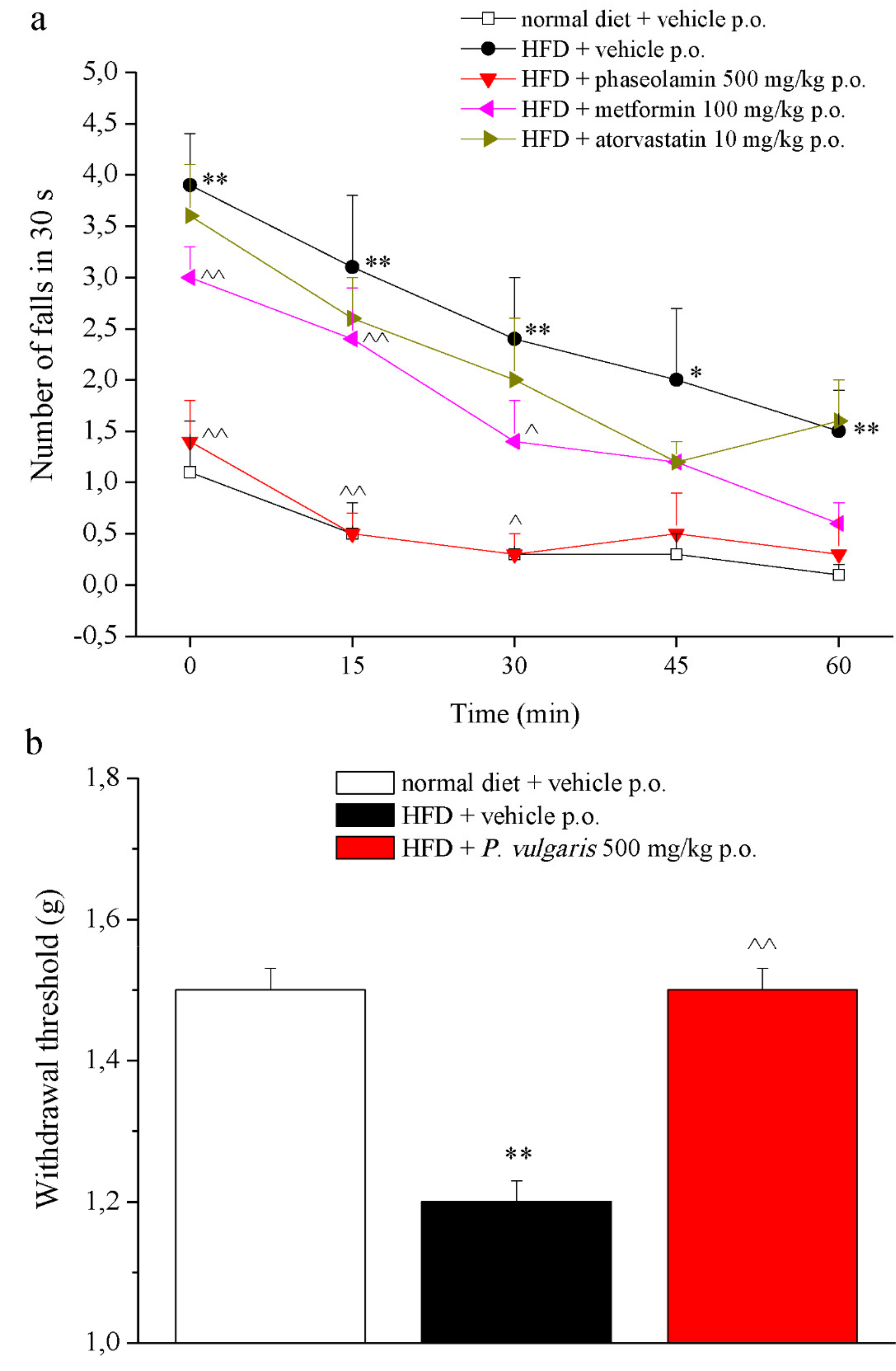
3.5. Behavioral Measurements
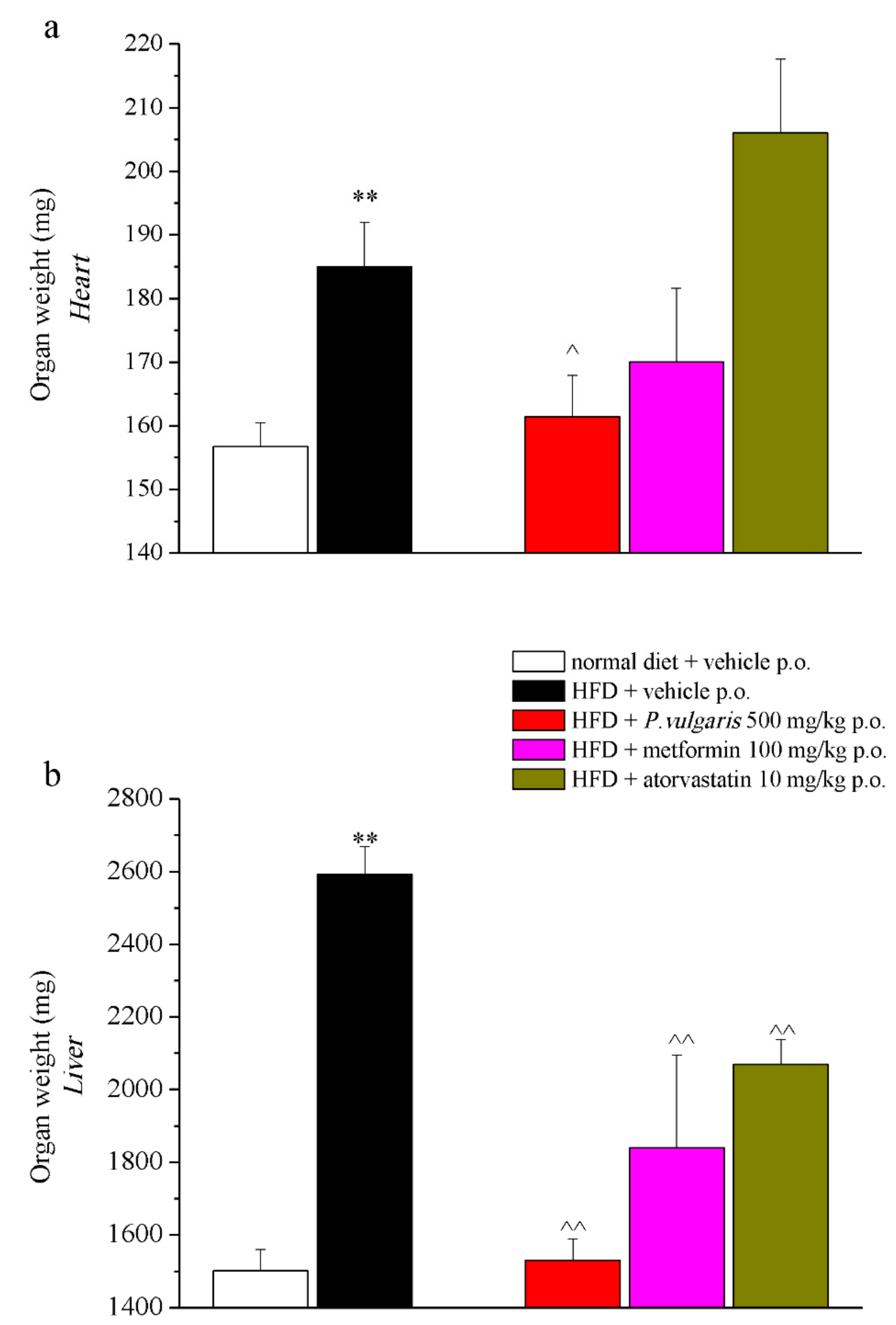
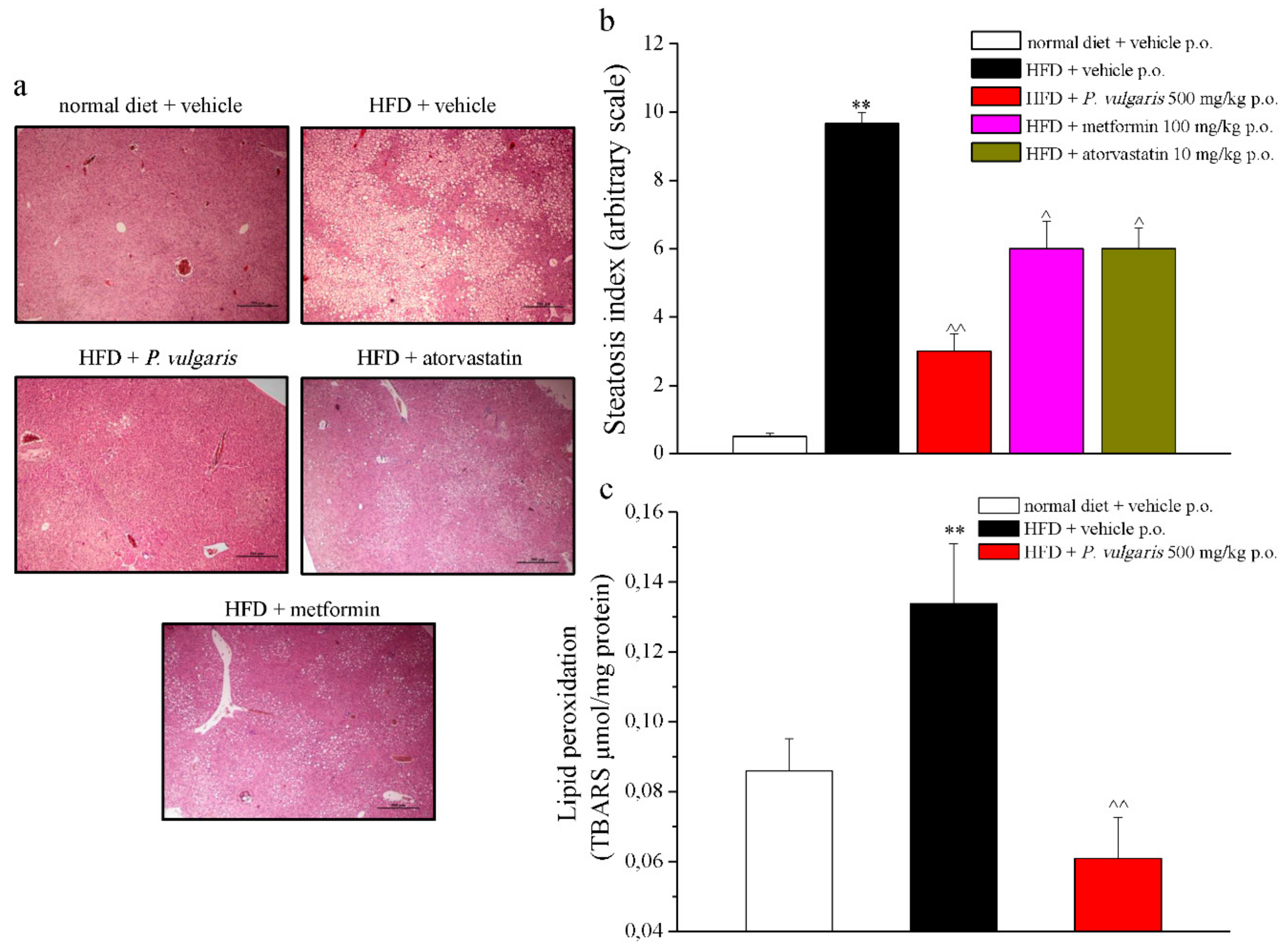
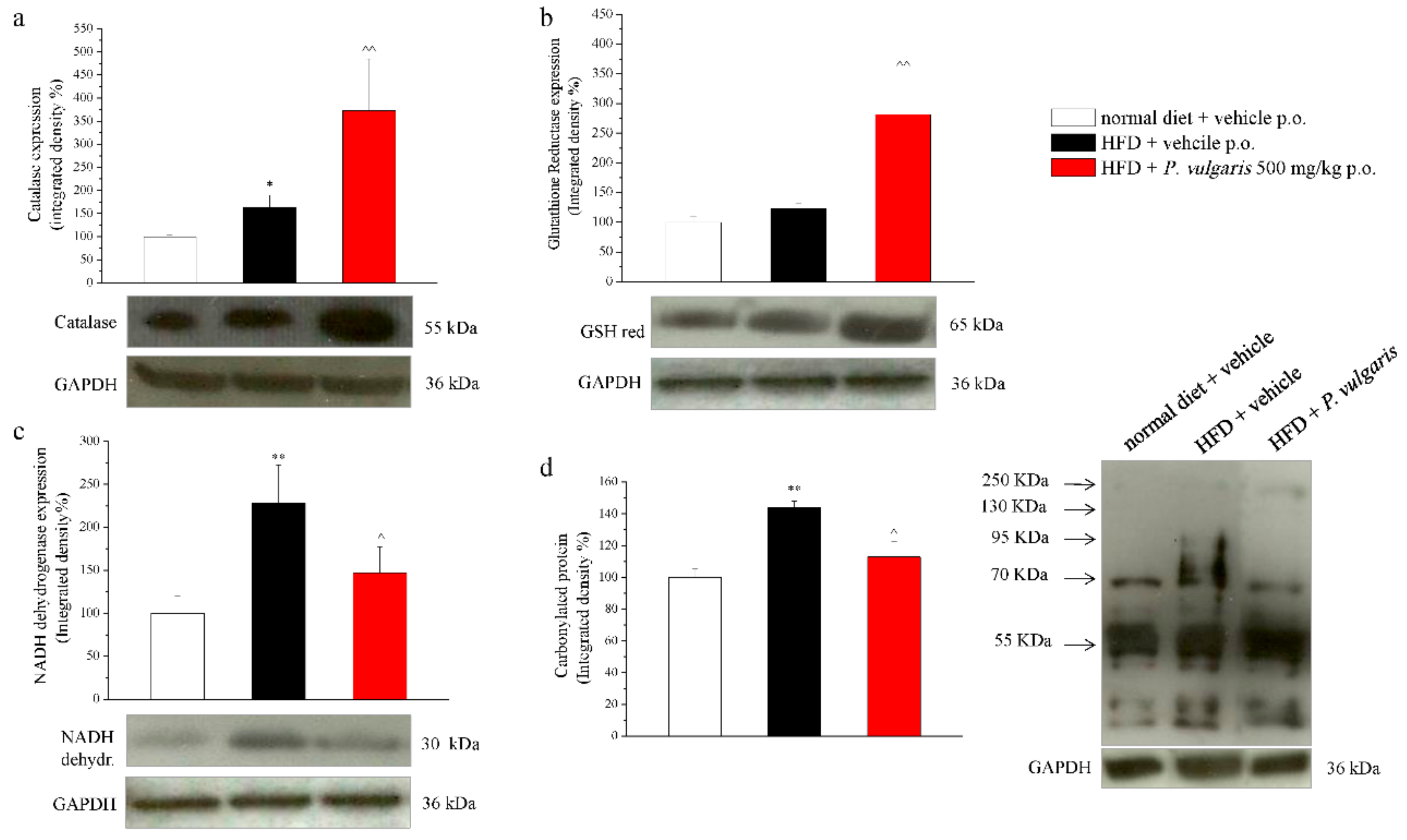
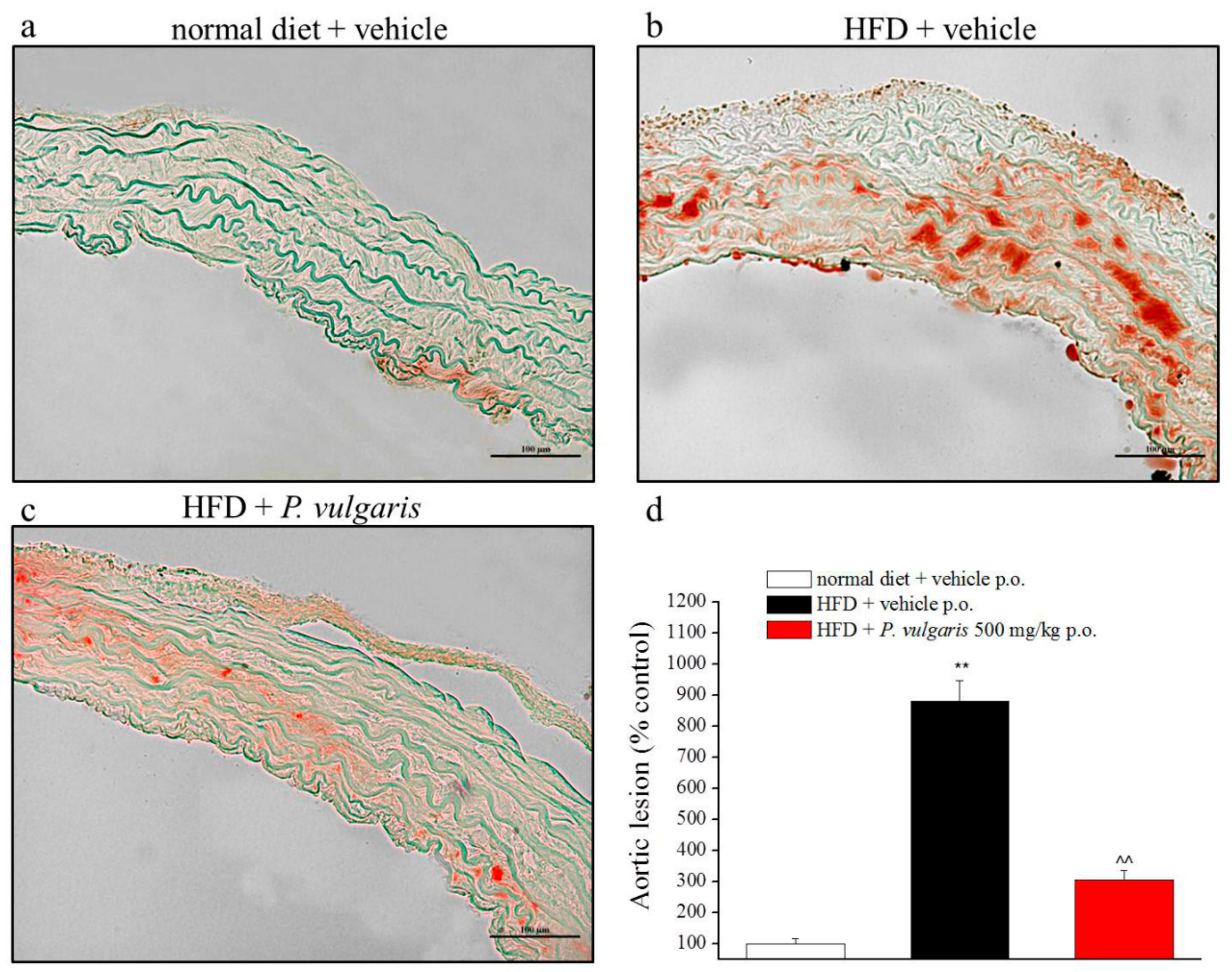
3.6. Ex Vivo Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Reaven, G.M. Resistance to insulin-stimulated glucose-uptake and hyperinsulinemia: Role in non-insulin-dependent diabetes, high blood-pressure, dyslipidemia and coronary heart disease. Diabetes Metab. 1991, 17, 78–86. [Google Scholar]
- Grundy, S.M.; Cleeman, J.I.; Daniels, S.R.; Donato, K.A.; Eckel, R.H.; Franklin, B.A.; Gordon, D.J.; Krauss, R.M.; Savage, P.J.; Smith, S.C.; et al. Diagnosis and management of the metabolic syndrome—An American Heart Association/National Heart, Lung, and Blood Institute Scientific Statement. Circulation 2005, 112, 2735–2752. [Google Scholar] [CrossRef] [PubMed]
- Mariotti, F.; Hermier, D.; Sarrat, C.; Magne, J.; Fenart, E.; Evrard, J.; Tome, D.; Huneau, J.F. Rapeseed protein inhibits the initiation of insulin resistance by a high-saturated fat, high-sucrose diet in rats. Br. J. Nutr. 2008, 100, 984–991. [Google Scholar] [CrossRef] [PubMed]
- Vijay-Kumar, M.; Aitken, J.D.; Carvalho, F.A.; Cullender, T.C.; Mwangi, S.; Srinivasan, S.; Sitaraman, S.V.; Knight, R.; Ley, R.E.; Gewirtz, A.T. Metabolic Syndrome and Altered Gut Microbiota in Mice Lacking Toll-Like Receptor 5. Science 2010, 328, 228–231. [Google Scholar] [CrossRef] [PubMed]
- Cicero, A.F.; Colletti, A. Role of phytochemicals in the management of metabolic syndrome. Phytomedicine 2016, 23, 1134–1144. [Google Scholar] [CrossRef] [PubMed]
- Patti, A.M.; Al-Rasadi, K.; Giglio, R.V.; Nikolic, D.; Mannina, C.; Castellino, G.; Chianetta, R.; Banach, M.; Cicero, A.F.; Lippi, G.; et al. Natural approaches in metabolic syndrome management. Arch. Med. Sci. 2018, 14, 422–441. [Google Scholar] [CrossRef] [PubMed]
- Obiro, W.C.; Zhang, T.; Jiang, B. The nutraceutical role of the Phaseolus vulgaris alpha-amylase inhibitor. Br. J. Nutr. 2008, 100, 1–12. [Google Scholar] [CrossRef]
- Brain-Isasi, S.; Alvarez-Lueje, A.; Higgins, T.J.V. Heterologous expression of an alpha-amylase inhibitor from common bean (Phaseolus vulgaris) in Kluyveromyces lactis and Saccharomyces cerevisiae. Microb. Cell. Fact. 2017, 16, 110. [Google Scholar] [CrossRef]
- Bowman, D.E. Amylase inhibitor of navy beans. Science 1945, 102, 358–359. [Google Scholar] [CrossRef]
- Marshall, J.J.; Lauda, C.M. Purification and properties of phaseolamin, an inhibitor of alpha-amylase, from the kidney bean, Phaseolus vulgaris. J. Biol. Chem. 1975, 250, 8030–8037. [Google Scholar]
- Oliveira, R.J.S.; De Oliveira, V.N.; Deconte, S.R.; Calabria, L.K.; Moraes, A.d.S.; Espindola, F.S. Phaseolamin treatment prevents oxidative stress and collagen deposition in the hearts of streptozotocin-induced diabetic rats. Diab. Vasc. Dis. Res. 2014, 11, 110–117. [Google Scholar] [CrossRef] [PubMed]
- McGrath, J.C.; Lilley, E. Implementing guidelines on reporting research using animals (ARRIVE etc.): New requirements for publication in BJP. Br. J. Pharmacol. 2015, 172, 3189–3193. [Google Scholar] [CrossRef] [PubMed]
- Mali, V.R.; Ning, R.; Chen, J.; Yang, X.P.; Xu, J.; Palaniyandi, S.S. Impairment of aldehyde dehydrogenase-2 by 4-hydroxy-2-nonenal adduct formation and cardiomyocyte hypertrophy in mice fed a high-fat diet and injected with low-dose streptozotocin. Exp. Biol. Med. 2014, 239, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Reed, M.J.; Meszaros, K.; Entes, L.J.; Claypool, M.D.; Pinkett, J.G.; Gadbois, T.M.; Reaven, G.M. A new model of type 2 diabetes: The fat-fed streptozotocin-treated rat. Metabolism 2000, 49, 1390–1394. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, K.; Viswanad, B.; Asrat, L.; Kaul, C.L.; Ramarao, P. Combination of high-fat diet-fed and low-dose streptozotocin-treated rat: A model for type 2 diabetes and pharmacological screening. Pharmacol. Res. 2005, 52, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, Y.; Nakamura, T.; Ishikawa, S.; Fujisaka, S.; Usui, I.; Tsuneyama, K.; Ichihara, Y.; Wada, T.; Hirata, Y.; Suganami, T.; et al. The radioprotective 105/MD-1 complex contributes to diet-induced obesity and adipose tissue inflammation. Diabetes 2012, 61, 1199–1209. [Google Scholar] [CrossRef]
- Fantini, N.; Cabras, C.; Lobina, C.; Colombo, G.; Gessa, G.L.; Riva, A.; Donzelli, F.; Morazzoni, P.; Bombardelli, E.; Carai, M.A.M. Reducing effect of a Phaseolus vulgaris dry extract on food intake, body weight, and glycemia in rats. J. Agric. Food. Chem. 2009, 57, 9316–9323. [Google Scholar] [CrossRef]
- Loi, B.; Fantini, N.; Colombo, G.; Gessa, G.L.; Riva, A.; Bombardelli, E.; Morazzoni, P.; Carai, M.A.M. Reducing effect of an extract of Phaseolus vulgaris on food intake in mice–focus on highly palatable foods. Fitoterapia 2013, 85, 14–19. [Google Scholar] [CrossRef]
- Gao, X.; Nan, Y.; Zhao, Y.; Yaun, Y.; Ren, B.; Sun, C.; Cao, K.; Yu, M.; Feng, X.; Ye, J. Atorvastatin reduces lipid accumulation in the liver by activating protein kinase A-mediated phosphorylation of perilipin 5. Biochim. Biophys. Acta. Mol. Cell. Biol. Lipids 2017, 1862, 1512–1519. [Google Scholar] [CrossRef]
- Wang, J.H.; Bose, S.; Shin, N.R.; Chin, Y.W.; Choi, Y.H.; Kim, H. Pharmaceutical impact of Houttuynia Cordata and metformin combination on high-fat-diet-induced metabolic disorders: Link to intestinal microbiota and metabolic endotoxemia. Front. Endocrinol. (Lousanne) 2018, 9, 620. [Google Scholar] [CrossRef]
- Vaught, J.; Pelley, K.; Costa, L.G.; Sether, P.; Enna, S.J. A comparison of the antinociceptive responses to GABA-receptor agonists THIP and baclofen. Neuropharmacology 1985, 24, 211–216. [Google Scholar] [CrossRef]
- Baptista-de-Souza, D.; Di Cesare Mannelli, L.; Zanardelli, M.; Micheli, L.; Nunes-de-Souza, R.L.; Canto-de-Souza, A.; Ghelardini, C. Serotonergic modulation in neuropathy induced by oxaliplatin: Effect on the 5HT2C receptor. Eur. J. Pharmacol. 2014, 735, 141–149. [Google Scholar] [CrossRef]
- Hargreaves, K.M.; Dubner, R.; Brown, F.; Flores, C.; Joris, J. A New and Sensitive Method for Measuring Thermal Nociception in Cutaneous Hyperalgesia. Pain 1988, 32, 77–88. [Google Scholar] [CrossRef]
- Komada, M.; Takao, K.; Miyakawa, T. Elevated plus maze for mice. J. Vis. Exp. 2008. [Google Scholar] [CrossRef]
- Shoji, H.; Takao, K.; Hattori, S.; Miyakawa, T. Age-related changes in behaviour in C57BL/6J mice from young adulthood to middle age. Mol. Brain 2016, 9, 11. [Google Scholar] [CrossRef]
- Zhang, Y.; Si, Y.; Yao, S.; Yang, N.; Song, G.; Sang, H.; Zu, D.; Xu, X.; Wang, J.; Qin, S. Celastrus orbiculatus Thunb. decreases athero-susceptibility in lipoproteins and the aorta of guinea pigs fed high fat diet. Lipids 2013, 48, 619–631. [Google Scholar] [CrossRef]
- Cinci, L.; Corti, F.; Di Cesare Mannelli, L.; Micheli, L.; Zanardelli, M.; Ghelardini, C. Oxidative, metabolic, and apoptotic responses of Schwann cells to high glucose levels. J. Biochem. Mol. Toxicol. 2015, 29, 274–279. [Google Scholar] [CrossRef]
- McGuinness, P.O.; Ayala, J.E.; Laughlin, M.R.; Wasserman, D.H. NIH experiment in centralized mouse phenotyping: The Vanderbilt experience and recommendations for evaluating glucose homeostasis in the mouse. Am. J. Physiol. Endocrinol. Metab. 2009, 297, E849–E855. [Google Scholar] [CrossRef]
- Fellmann, L.; Nascimento, A.R.; Tibirica, E.; Bousquet, P. Murine models for pharmacological studies of the metabolic syndrome. Pharmacol. Ther. 2013, 137, 331–340. [Google Scholar] [CrossRef]
- Carai, M.A.; Fantini, N.; Loi, B.; Colombo, G.; Gessa, G.L.; Riva, A.; Bombardelli, E.; Morazzoni, P. Multiple cycles of repeated treatments with a Phaseolus vulgaris dry extract reduce food intake and body weight in obese rats. Br. J. Nutr. 2011, 106, 762–768. [Google Scholar] [CrossRef]
- Lorrai, I.; Piga, V.; Carai, M.A.; Riva, A.; Morazzoni, P.; Gessa, G.L.; Colombo, G.; Maccioni, P. A Phaseolus vulgaris Extract Reduces Cue-Induced Reinstatement of Chocolate Seeking in Rats. Front. Pharmacol. 2016, 7, 109. [Google Scholar] [CrossRef]
- Reverri, E.J.; Randolph, J.M.; Kappagoda, C.T.; Park, E.; Edirisinghe, I.; Burton-Freeman, B.M. Assessing beans as a source of intrinsic fiber on satiety in men and women with metabolic syndrome. Appetite 2017, 118, 75–81. [Google Scholar] [CrossRef]
- Tormo, M.A.; Gil-Exojo, I.; De Tejada, A.R.; Campillo, J.E. Hypoglycaemic and anorexigenic activities of an alpha-amylase inhibitor from white kidney beans (Phaseolus vulgaris) in Wistar rats. Br. J. Nutr. 2004, 92, 785–790. [Google Scholar] [CrossRef]
- Nuñez-Aragón, P.N.; Segura-Campos, M.; Negrete-León, E.; Acevedo-Fernández, J.J.; Betancur-Ancona, D.; Chel-Guerrero, L.; Castañeda-Corral, G. Protein hydrolysates and ultrafiltered <1 KDa fractions from Phaseolus lunatus, Phaseolus vulgaris and Mucuna pruriens exhibit antihyperglycemic activity, intestinal glucose absorption and α-glucosidase inhibition with no acute toxicity in rodents. J. Sci Food Agric. 2019, 99, 587–595. [Google Scholar]
- Tormo, M.A.; Gil-Exojo, I.; de Tejada, A.R.; Campillo, J.E. White bean amylase inhibitor administered orally reduces glycaemia in type 2 diabetic rats. Br. J. Nutr. 2006, 96, 539–544. [Google Scholar]
- Barrett, M.L.; Udani, J.K. A proprietary alpha-amylase inhibitor from white bean (Phaseolus vulgaris): A review of clinical studies on weight loss and glycemic control. Nutr. J. 2011, 10, 24. [Google Scholar] [CrossRef]
- Spadafranca, A.; Rinelli, S.; Riva, A.; Morazzoni, P.; Magni, P.; Bertoli, S.; Battezzati, A. Phaseolus vulgaris extract affects glycometabolic and appetite control in healthy human subjects. Br. J. Nutr. 2013, 109, 1789–1795. [Google Scholar] [CrossRef]
- Dali-Youcef, N.; Mecili, M.; Ricci, R.; Andres, E. Metabolic inflammation: Connecting obesity and insulin resistance. Ann. Med. 2013, 45, 242–253. [Google Scholar] [CrossRef]
- Guarino, D.; Nannipieri, M.; Iervasi, G.; Taddei, S.; Bruno, R.M. The role of the autonomic nervous system in the pathophysiology of obesity. Front. Physiol. 2017, 8, 665. [Google Scholar] [CrossRef]
- Chokshi, D. Subchronic oral toxicity of a standardized white kidney bean (Phaseolus vulgaris) extract in rats. Food Chem. Toxicol. 2007, 45, 32–40. [Google Scholar] [CrossRef]
- Stino, A.M.; Smith, A.G. Peripheral neuropathy in prediabetes and the metabolic syndrome. J. Diabetes Investig. 2017, 8, 646–655. [Google Scholar] [CrossRef]
- Takase, K.; Tsuneoka, Y.; Oda, S.; Kuroda, M.; Funato, H. High-fat diet feeding alters olfactory-, social-, and reward-related behaviors of mice independent of obesity. Obesity 2016, 24, 886–894. [Google Scholar] [CrossRef]
- Bechmann, L.P.; Hannivoort, R.A.; Gerken, G.; Hotamisligil, G.S.; Trauner, M.; Canbay, A. The interaction of hepatic lipid and glucose metabolism in liver diseases. J. Hepatol. 2012, 56, 952–964. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
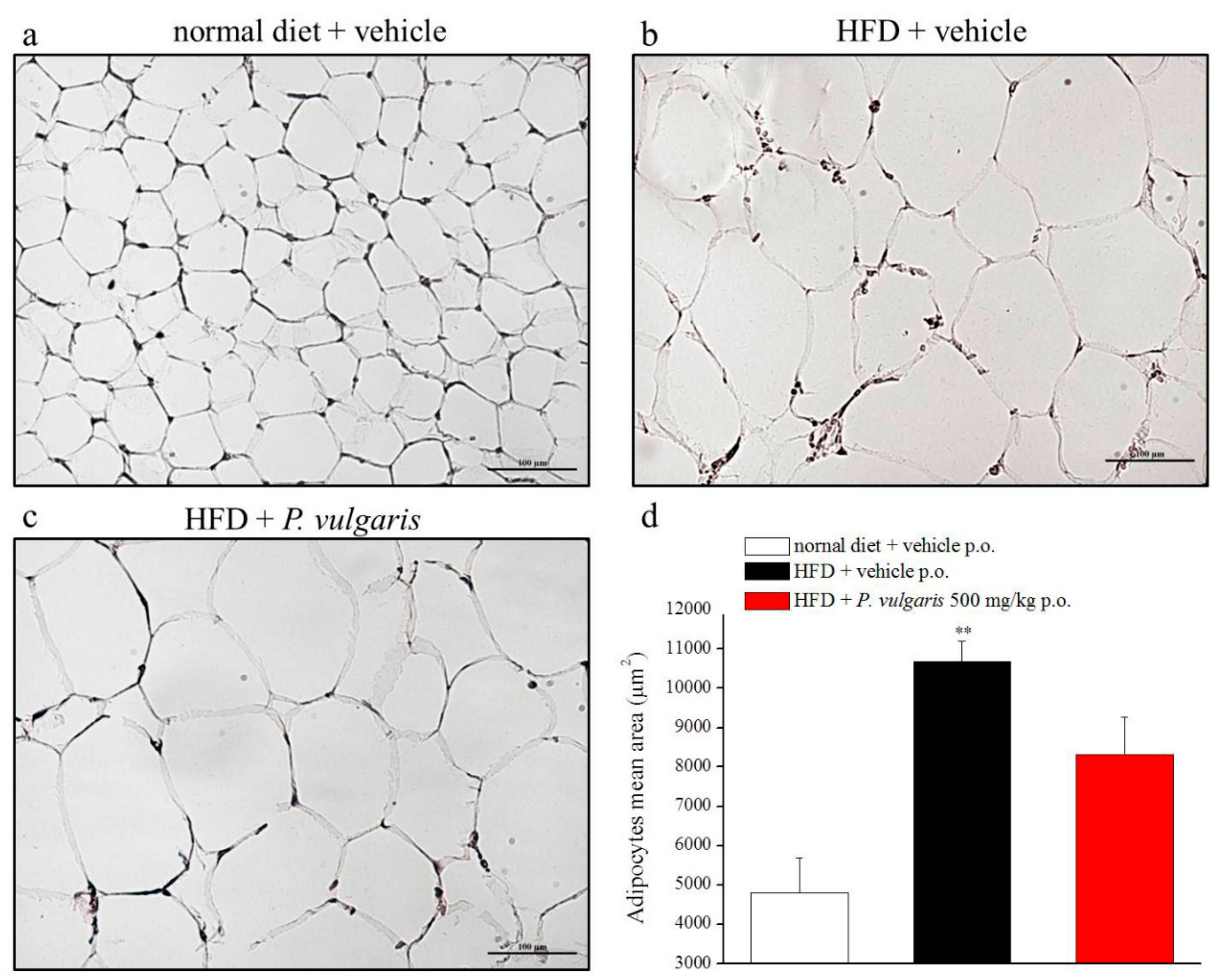{kind=link}
| Group | ||||||
|---|---|---|---|---|---|---|
| Week | Normal Diet + Vehicle | HFD + Vehicle | HFD + P. vulgaris | HFD + Metformin | HFD + Atorvastatin | |
| Glucose (mg/dL) | 13 | 101.0 ± 3.4 | 139.3 ± 6.9 * | 103.0 ± 5.1^ | 132.0 ± 9.9 | 133.0 ± 6.2 |
| 15 | 99.0 ± 10.1 | 149.5 ± 3.9 ** | 103.0 ± 8.0^^ | 113.3 ± 7.4 ^ | 112.8 ± 7.1 ^ | |
| 17 | 103.3 ± 6.4 | 138.5 ± 9.1 * | 127.8 ± 2.0 ^^ | 121.0 ± 5.9 ^ | 146.3 ± 5.5 | |
| 19 | 99.3 ± 4.6 | 137.3 ± 9.4 ** | 112.0 ± 4.4 ^ | 116.3 ± 5.7 ^ | 137.3 ± 14.0 | |
| Triglycerides (mg/dL) | 13 | 62.0 ± 6.9 | 132.3 ± 11.3 * | 123.0 ± 18.0 | 105.0 ± 14.6 | 102.0 ± 9.0 ^ |
| 15 | 72.3 ± 13.6 | 135.3 ± 6.0 ** | 86.5 ± 10.7 ^ | 93.3 ± 10.4 ^ | 103.3 ± 9.4 ^ | |
| 17 | 78.4 ± 2.7 | 136.8 ± 2.3 ** | 105.3 ± 7.8 ^^ | 113.8 ± 5.8 ^ | 108.8 ± 7.2 ^^ | |
| 19 | 75.8 ± 11.0 | 133.5 ± 7.7 ** | 107.5 ± 9.3 ^^ | 118.3 ± 3.7 | 104.0 ± 3.0 ^^ | |
| Total cholesterol (mg/dL) | 13 | <100 | 133.7 ± 4.4 ** | 104.3 ± 3.5 ^^ | 128.6 ± 8.2 | 105.0 ± 6.0 ^^ |
| 15 | 105.0 ± 5.0 | 135.8 ± 1.3 ** | 103.0 ± 2.3 ^^ | 118.3 ± 7.2 | 105.0 ± 4.3 ^^ | |
| 17 | 107.0 ± 1.2 | 147.0 ± 2.8 ** | 123.0 ± 6.4 ^^ | 139.3 ± 12.5 | 109.5 ± 5.0 ^^ | |
| 19 | 103.8 ± 4.1 | 147.3 ± 4.1 ** | 100.5 ± 7.6 ^^ | 103.9 ± 9.2 ^^ | 76.4 ± 1.4 ^^ | |
| HDL (mg/dL) | 19 | 51.8 ± 8.5 | 62.3 ± 7.0 | 41.7 ± 10.2 | 59.1 ± 10.5 | 46.8 ± 9.5 |
| LDL (mg/dL) | 19 | 43.8 ± 6.7 | 62.3 ± 7.0 | 38.4 ± 6.3 ^^ | 46.0 ± 9.4 ^ | 50.2 ± 3.0 ^^ |
| Group | |||||
|---|---|---|---|---|---|
| Normal Diet + Vehicle | HFD + Vehicle | HFD + P. vulgaris | HFD + Metformin | HFD + Atorvastatin | |
| Insulin (ng/mL) | 4.2 ± 0.5 | 46.1 ± 7.3 ** | 8.9 ± 8.2 ^^ | 10.2 ± 7.6 ^^ | 34.9 ± 10.5 |
| Ghrelin (ng/mL) | 38.6 ± 5.8 | 10.5 ± 3.4 * | 17.0 ± 2.8 | 13.8 ± 4.3 | 15.8 ± 2.4 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Micheli, L.; Lucarini, E.; Trallori, E.; Avagliano, C.; De Caro, C.; Russo, R.; Calignano, A.; Ghelardini, C.; Pacini, A.; Di Cesare Mannelli, L. Phaseolus vulgaris L. Extract: Alpha-Amylase Inhibition against Metabolic Syndrome in Mice. Nutrients 2019, 11, 1778. https://doi.org/10.3390/nu11081778
Micheli L, Lucarini E, Trallori E, Avagliano C, De Caro C, Russo R, Calignano A, Ghelardini C, Pacini A, Di Cesare Mannelli L. Phaseolus vulgaris L. Extract: Alpha-Amylase Inhibition against Metabolic Syndrome in Mice. Nutrients. 2019; 11(8):1778. https://doi.org/10.3390/nu11081778
Chicago/Turabian StyleMicheli, Laura, Elena Lucarini, Elena Trallori, Carmen Avagliano, Carmen De Caro, Roberto Russo, Antonio Calignano, Carla Ghelardini, Alessandra Pacini, and Lorenzo Di Cesare Mannelli. 2019. "Phaseolus vulgaris L. Extract: Alpha-Amylase Inhibition against Metabolic Syndrome in Mice" Nutrients 11, no. 8: 1778. https://doi.org/10.3390/nu11081778
APA StyleMicheli, L., Lucarini, E., Trallori, E., Avagliano, C., De Caro, C., Russo, R., Calignano, A., Ghelardini, C., Pacini, A., & Di Cesare Mannelli, L. (2019). Phaseolus vulgaris L. Extract: Alpha-Amylase Inhibition against Metabolic Syndrome in Mice. Nutrients, 11(8), 1778. https://doi.org/10.3390/nu11081778