Antenatal Influenza A-Specific IgA, IgM, and IgG Antibodies in Mother’s Own Breast Milk and Donor Breast Milk, and Gastric Contents and Stools from Preterm Infants



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants and Sample Collection
2.1.1. Participants and Enrollment
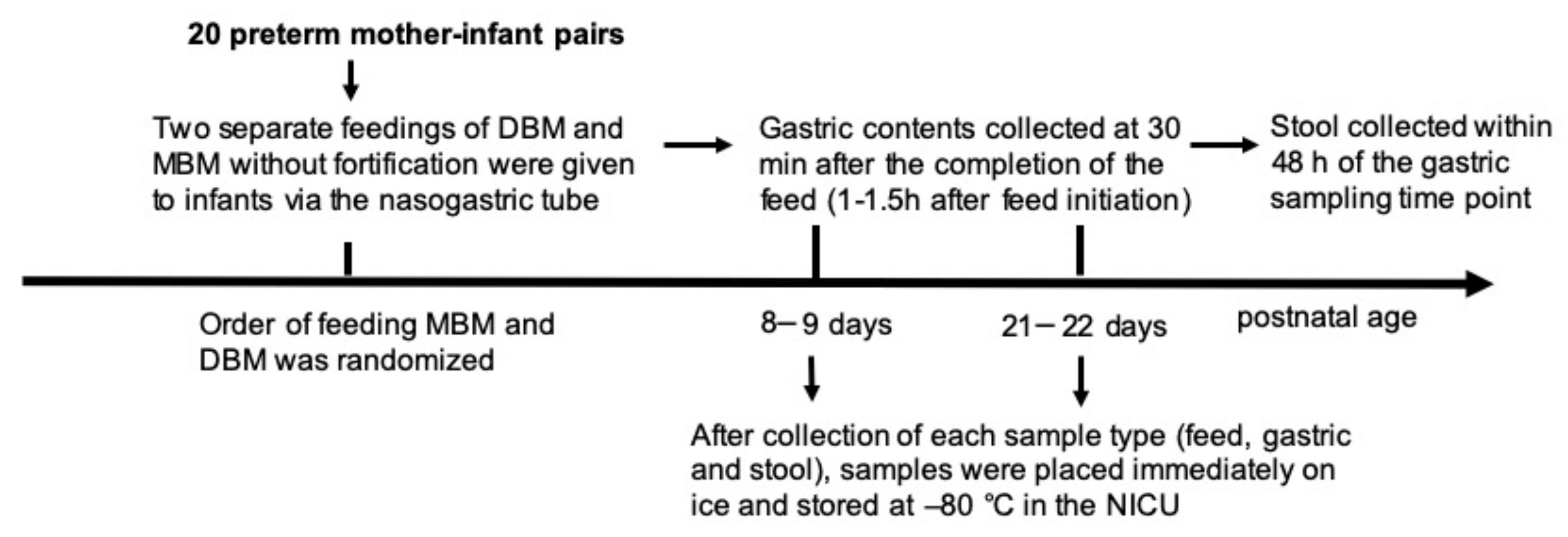
2.1.2. Feeding and Sampling
2.2. Sample Preparation and ELISAs
2.3. Pasteurization Effect on Antibodies Specific to Influenza A
2.4. Statistical Analyses
3. Results
3.1. Infant Demographics
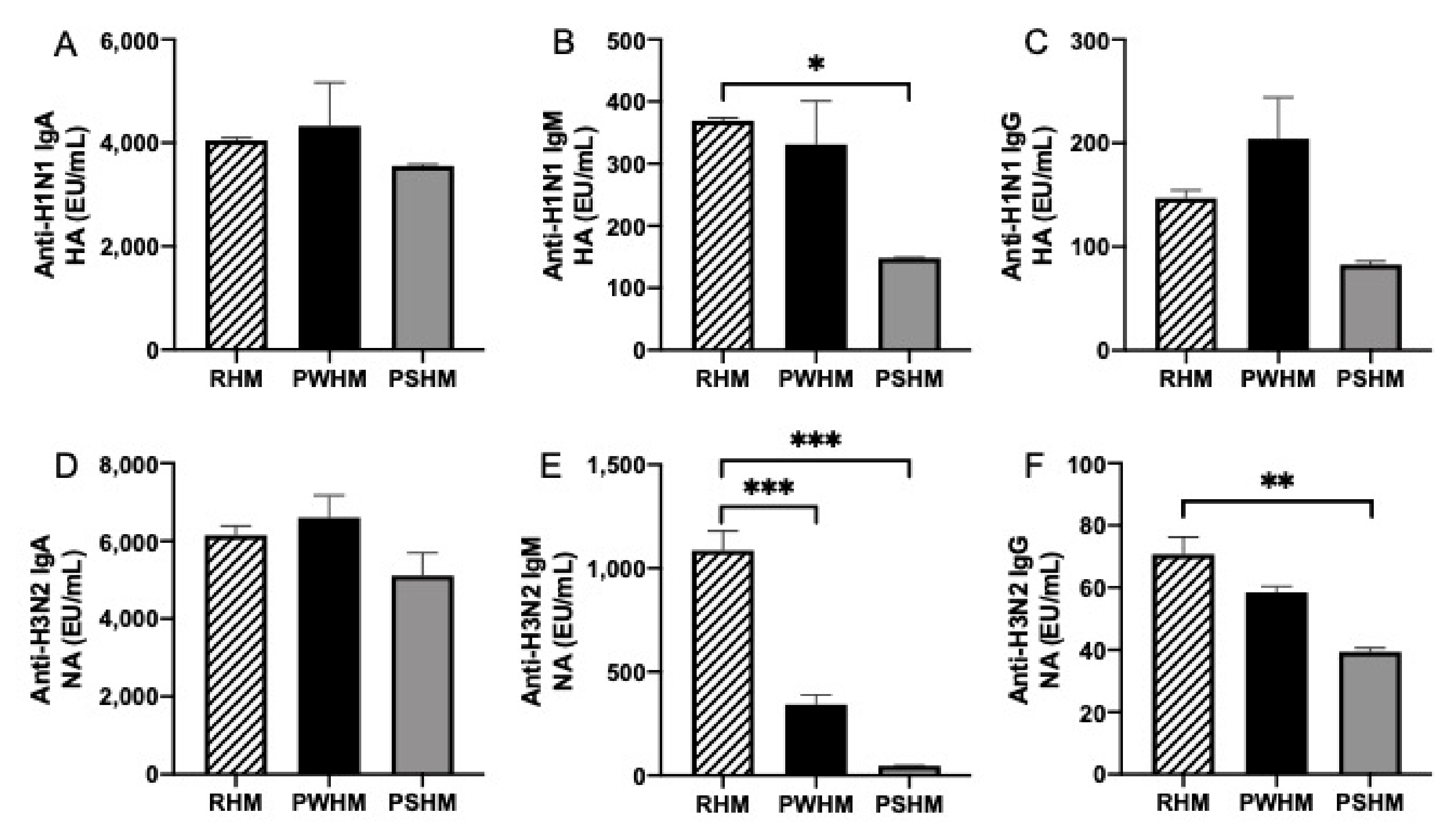
3.2. Anti-Influenza Antibody Relative Abundance in Feeds
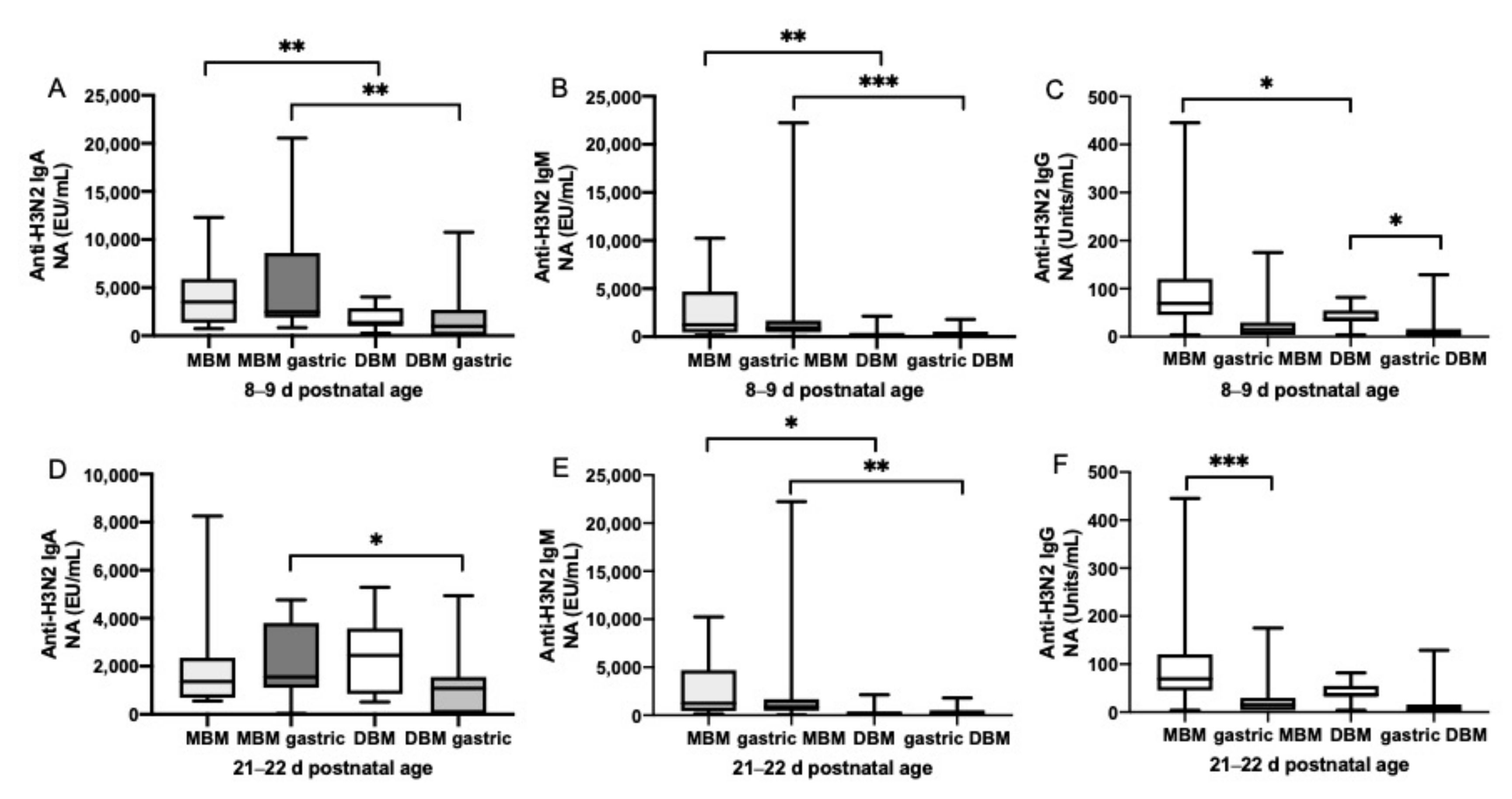
3.3. Anti-Influenza Antibody Relative Abundance in Gastric Contents
3.4. Gastric Digestion of Anti-Influenza Antibody from MBM and DBM
3.5. Postnatal Age Effects on Anti-Influenza A Antibodies in Preterm Infant Stools Derived from Mixed MBM/DBM
3.6. Pasteurization Effects on Human Milk Anti-Influenza A Antibodies
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Glezen, W.P.; Taber, L.H.; Frank, A.L.; Gruber, W.C.; Piedra, P.A. Influenza virus infections in infants. Pediatr. Infect. Dis. J. 1997, 16, 1065–1068. [Google Scholar] [CrossRef] [PubMed]
- Louie, J.K.; Schechter, R.; Honarmand, S.; Guevara, H.F.; Shoemaker, T.R.; Madrigal, N.Y.; Woodfill, C.J.; Backer, H.D.; Glaser, C.A. Severe pediatric influenza in California, 2003–2005: Implications for immunization recommendations. Pediatrics 2006, 2005, 1373. [Google Scholar] [CrossRef] [PubMed]
- Cox, R.J.; Brokstad, K.A.; Ogra, P.L. Influenza virus: Immunity and vaccination strategies. Comparison of the immune response to inactivated and live, attenuated influenza vaccines. Scand. J. Immunol. 2004, 59, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Salam, R.A.; Das, J.K.; Soeandy, C.D.; Lassi, Z.S.; Bhutta, Z.A. Impact of Haemophilus influenzae type B (Hib) and viral influenza vaccinations in pregnancy for improving maternal, neonatal and infant health outcomes. Cochrane Datab. Syst. Rev. 2015, 6, 1–41. [Google Scholar] [CrossRef] [PubMed]
- Omer, S.B.; Clark, D.R.; Aqil, A.R.; Tapia, M.D.; Nunes, M.C.; Kozuki, N.; Steinhoff, M.C.; Madhi, S.A.; Wairagkar, N. Maternal influenza immunization and prevention of severe clinical pneumonia in young infants. Pediatr. Infect. Dis. J. 2018, 37, 436–440. [Google Scholar] [CrossRef] [PubMed]
- Zaman, K.; Roy, E.; Arifeen, S.E.; Rahman, M.; Raqib, R.; Wilson, E.; Omer, S.B.; Shahid, N.S.; Breiman, R.F.; Steinhoff, M.C. Effectiveness of maternal influenza immunization in mothers and infants. N. Engl. J. Med. 2008, 359, 1555–1564. [Google Scholar] [CrossRef] [PubMed]
- Eick, A.A.; Uyeki, T.M.; Klimov, A.; Hall, H.; Reid, R.; Santosham, M.; O’Brien, K.L. Maternal influenza vaccination and effect on influenza virus infection in young infants. Arch. Pediatr. Adolesc. Med. 2011, 165, 104–111. [Google Scholar] [CrossRef]
- Niewiesk, S. Maternal Antibodies: Clinical Significance, Mechanism of interference with immune responses, and possible vaccination strategies. Front. Immunol. 2014, 5, 446. [Google Scholar] [CrossRef]
- Schlaudecker, E.P.; Steinhoff, M.C.; Omer, S.B.; McNeal, M.M.; Roy, E.; Arifeen, S.E.; Dodd, C.N.; Raqib, R.; Breiman, R.F.; Zaman, K. IgA and neutralizing antibodies to influenza A virus in human milk: A randomized trial of antenatal influenza immunization. PLoS ONE 2013, 8, e70867. [Google Scholar] [CrossRef]
- Finn, A. Clinical Trials of Influenza Vaccines: Special Challenges; Humana Press: New York, NY, USA, 2019; pp. 567–573. [Google Scholar]
- Sagrera, X.; Ginovart, G.; Raspall, F.; Rabella, N.; Sala, P.; Sierra, M.; Demestre, X.; Vila, C. Outbreaks of influenza A virus infection in neonatal intensive care units. Pediatr. Infect. Dis. J. 2002, 21, 196–200. [Google Scholar] [CrossRef]
- Carroll, K.; Herrmann, K.R. The cost of using donor human milk in the NICU to achieve exclusively human milk feeding through 32 weeks postmenstrual age. Breastfeed. Med. 2013, 8, 286–290. [Google Scholar] [CrossRef] [PubMed]
- Demers-Mathieu, V.; Huston, R.K.; Markell, A.M.; McCulley, E.A.; Martin, R.L.; Spooner, M.; Dallas, D.C. Differences in maternal immunoglobulins within mother’s own breast milk and donor breast milk and across digestion in preterm infants. Nutrients 2019, 11, 920. [Google Scholar] [CrossRef] [PubMed]
- Evans, T.J.; Ryley, H.C.; Neale, L.M.; Dodge, J.A.; Lewarne, V.M. Effect of storage and heat on antimicrobial proteins in human milk. Arch. Dis. Child. 1978, 53, 239–241. [Google Scholar] [CrossRef] [PubMed]
- Ford, J.E.; Law, B.A.; Marshall, V.M.; Reiter, B. Influence of the heat treatment of human milk on some of its protective constituents. J. Pediatr. 1977, 90, 29–35. [Google Scholar] [CrossRef]
- Demers-Mathieu, V.; Underwood, M.A.; Beverly, R.L.; Nielsen, S.D.; Dallas, D.C. Comparison of human milk immunoglobulin survival during gastric digestion between preterm and term infants. Nutrients 2018, 10, 631. [Google Scholar] [CrossRef] [PubMed]
- Chandra, R.K. Immunoglobulin and protein levels in breast milk produced by mothers of preterm infants. Nutr. Res. 1982, 2, 27–30. [Google Scholar] [CrossRef]
- Ballabio, C.; Bertino, E.; Coscia, A.; Fabris, C.; Fuggetta, D.; Molfino, S.; Testa, T.; Sgarrella, M.; Sabatino, G.; Restani, P. Immunoglobulin—A profile in breast milk from mothers delivering full term and preterm infants. Int. J. Immunopathol. Pharmacol. 2007, 20, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Montagne, P.; Cuillière, M.L.; Molé, C.; Béné, M.C.; Faure, G. Immunological and nutritional composition of human milk in relation to prematurity and mothers’ parity during the first 2 weeks of lactation. J. Pediatr. Gastroenterol. Nutr. 1999, 29, 75–80. [Google Scholar] [CrossRef]
- Gross, S.J.; Buckley, R.H.; Wakil, S.S.; McAllister, D.C.; David, R.J.; Faix, R.G. Elevated IgA concentration in milk produced by mothers delivered of preterm infants. J. Pediatr. 1981, 99, 389–393. [Google Scholar] [CrossRef]
- Koenig, Á.; Diniz, E.M.D.A.; Barbosa, S.F.C.; Vaz, F.A.C. Immunologic factors in human milk: The effects of gestational age and pasteurization. J. Hum. Lact. 2005, 21, 439–443. [Google Scholar] [CrossRef]
- Decarlo, J.O.; Tramer, A.; Startzman, H.N. Iodized oil aspiration in the newborn. Arch. Pediatr. Adolesc. Med. 1952, 84, 442–445. [Google Scholar] [CrossRef]
- Laegreid, A.; OTNÆSS, A.B.K.; Ørstavik, I.; Carlsen, K.H. Neutralizing activity in human milk fractions against respiratory syncytial virus. Acta Paediatr. 1986, 75, 696–701. [Google Scholar] [CrossRef]
- Mantis, N.J.; Cheung, M.C.; Chintalacharuvu, K.R.; Rey, J.; Corthésy, B.; Neutra, M.R. Selective adherence of IgA to murine Peyer’s patch M cells: Evidence for a novel IgA receptor. J. Immunol. 2002, 169, 1844–1851. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, A.; Kelsall, B.L. Unique functions of CD11b+, CD8α+, and double-negative Peyer’s patch dendritic cells. J. Immunol. 2001, 166, 4884–4890. [Google Scholar] [CrossRef] [PubMed]

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Demers-Mathieu, V.; Huston, R.K.; Markell, A.M.; McCulley, E.A.; Martin, R.L.; Dallas, D.C. Antenatal Influenza A-Specific IgA, IgM, and IgG Antibodies in Mother’s Own Breast Milk and Donor Breast Milk, and Gastric Contents and Stools from Preterm Infants. Nutrients 2019, 11, 1567. https://doi.org/10.3390/nu11071567
Demers-Mathieu V, Huston RK, Markell AM, McCulley EA, Martin RL, Dallas DC. Antenatal Influenza A-Specific IgA, IgM, and IgG Antibodies in Mother’s Own Breast Milk and Donor Breast Milk, and Gastric Contents and Stools from Preterm Infants. Nutrients. 2019; 11(7):1567. https://doi.org/10.3390/nu11071567
Chicago/Turabian StyleDemers-Mathieu, Veronique, Robert K. Huston, Andi M. Markell, Elizabeth A. McCulley, Rachel L. Martin, and David C. Dallas. 2019. "Antenatal Influenza A-Specific IgA, IgM, and IgG Antibodies in Mother’s Own Breast Milk and Donor Breast Milk, and Gastric Contents and Stools from Preterm Infants" Nutrients 11, no. 7: 1567. https://doi.org/10.3390/nu11071567
APA StyleDemers-Mathieu, V., Huston, R. K., Markell, A. M., McCulley, E. A., Martin, R. L., & Dallas, D. C. (2019). Antenatal Influenza A-Specific IgA, IgM, and IgG Antibodies in Mother’s Own Breast Milk and Donor Breast Milk, and Gastric Contents and Stools from Preterm Infants. Nutrients, 11(7), 1567. https://doi.org/10.3390/nu11071567