Lack of a Synergistic Effect on Cardiometabolic and Redox Markers in a Dietary Supplementation with Anthocyanins and Xanthophylls in Postmenopausal Women


 , ,
, ,  ,
,  and
and 
Abstract
1. Introduction
2. Material and Methods
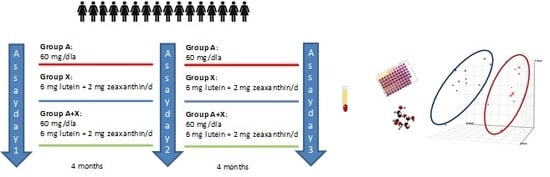
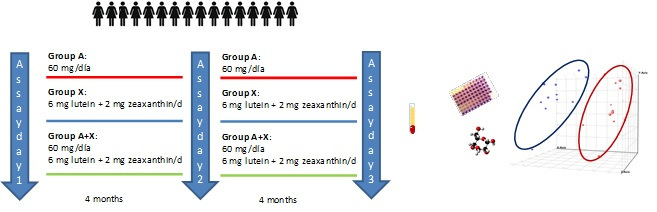
2.1. Subjects and Study Design
2.2. Preparation of Blood Samples
2.3. Cytokines and Metaloproteinases Analysis
2.4. Total Polyphenols and Ferric Reducing Antioxidant Power (FRAP) Analysis
2.5. Untargeted Serum Metabolomics by HPLC-QTOF
2.6. Statistics
3. Results
3.1. Study Outcomes: Biochemical and Cardiometabolomic Parameters
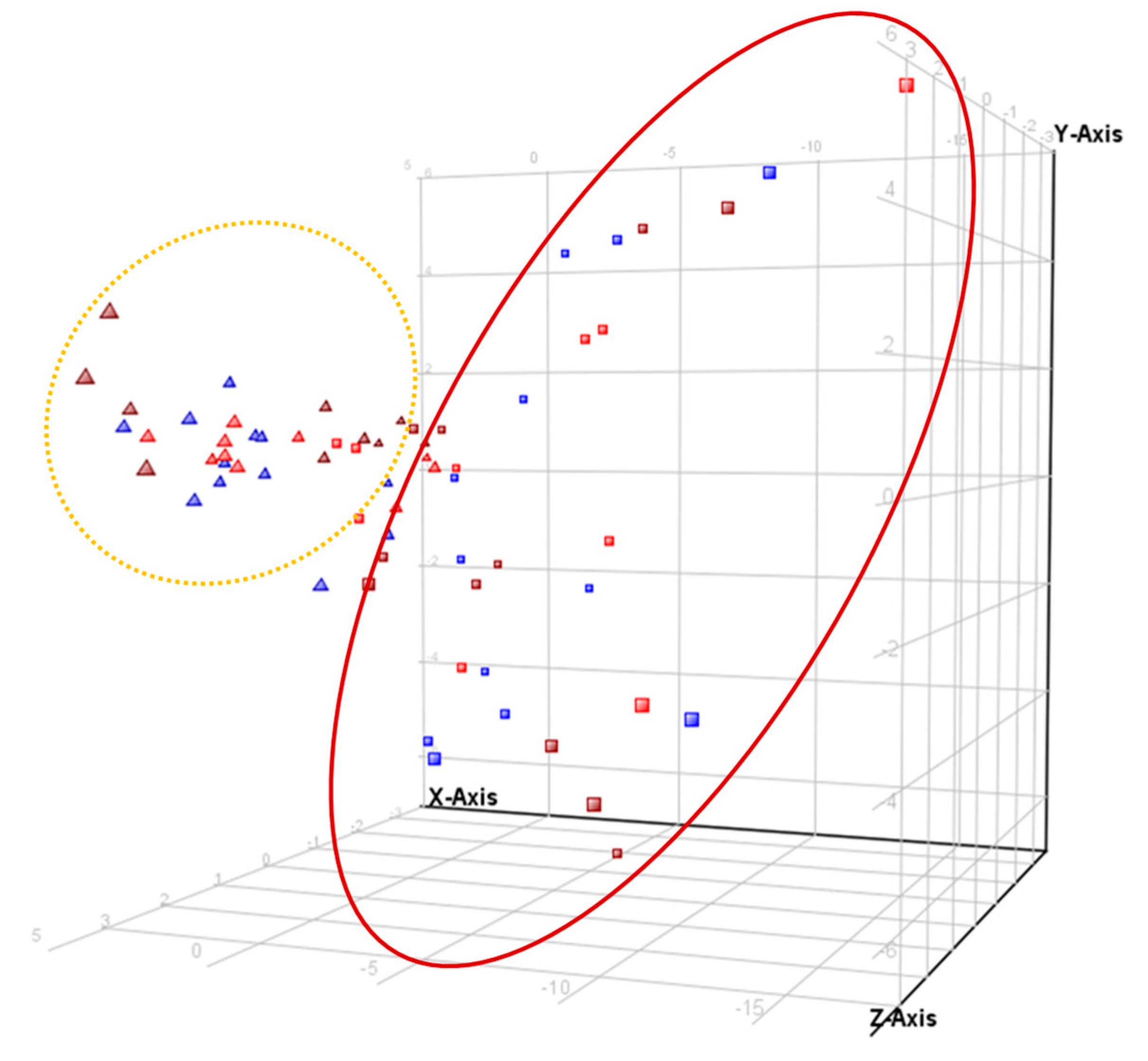
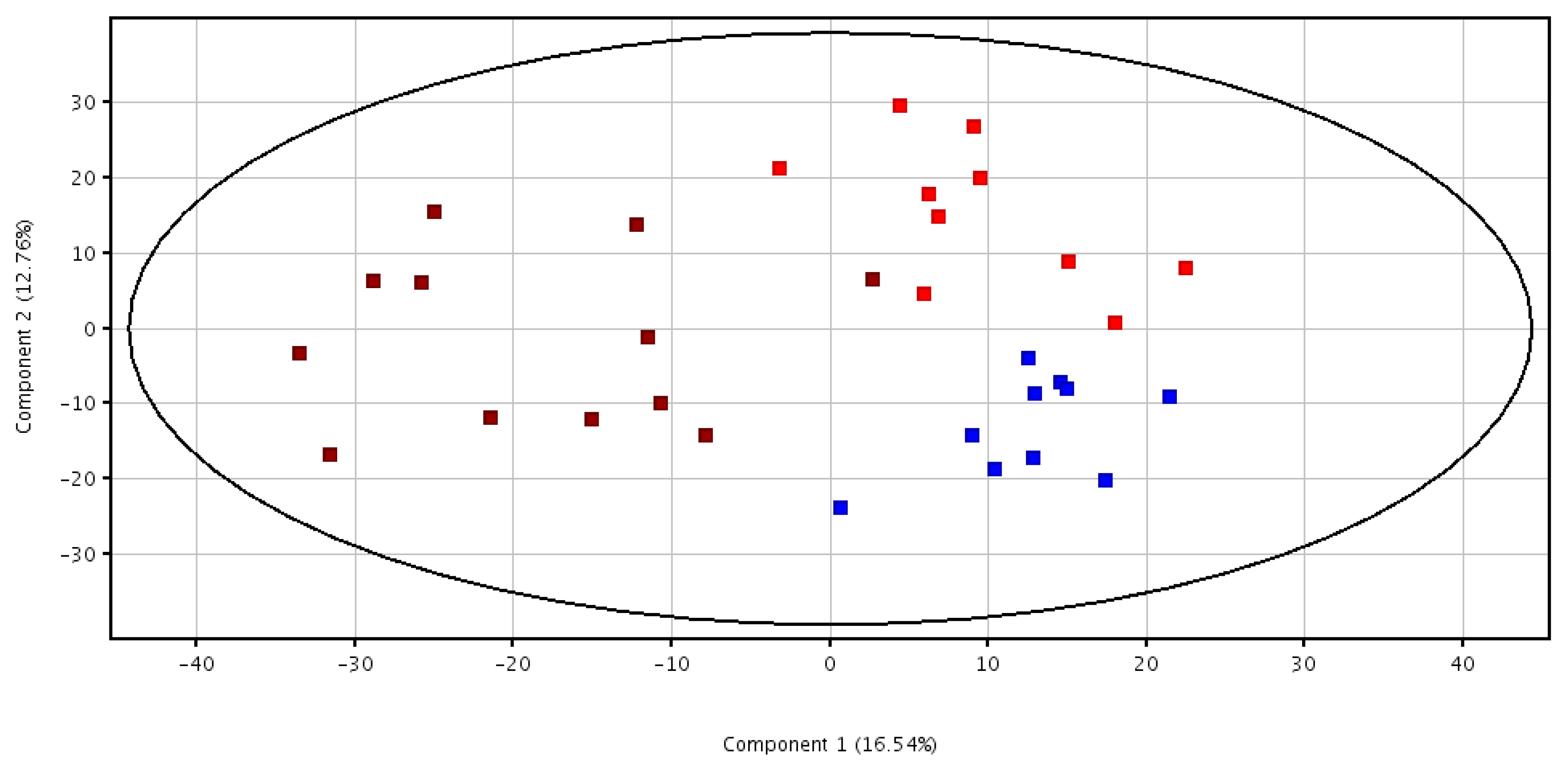
3.2. Metabolomic Analysis
4. Discussion
Metabolomic Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Aune, D.; Keum, N.; Giovannucci, E.; Fadnes, L.T.; Boffetta, P.; Greenwood, D.C.; Tonstad, S.; Vatten, L.J.; Riboli, E.; Norat, T. Dietary intake and blood concentrations of antioxidants and the risk of cardiovascular disease. total cancer. and all-cause mortality: A systematic review and dose-response meta-analysis of prospective studies. Am. J. Clin. Nutr. 2018, 108, 1069–1091. [Google Scholar] [CrossRef] [PubMed]
- García-Conesa, M.T.; Chambers, K.; Combet, E.; Pinto, P.; Garcia-Aloy, M.; Andrés-Lacueva, C.; de Pascual-Teresa, S.; Mena, P.; Konic Ristic, A.; Hollands, W.J.; et al. Meta-Analysis of the Effects of Foods and Derived Products Containing Ellagitannins and Anthocyanins on Cardiometabolic Biomarkers: Analysis of Factors Influencing Variability of the Individual Responses. Int. J. Mol. Sci. 2018, 19, 694. [Google Scholar] [CrossRef] [PubMed]
- García-Alonso, M.; Rimbach, G.; Rivas-Gonzalo, J.C.; De Pascual-Teresa, S. Antioxidant and cellular activities of anthocyanins and their corresponding vitisins A-studies in platelets, monocytes, and human endothelial cells. J. Agric. Food Chem. 2004, 52, 3378–3384. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, M.; Martin-Santamaria, S.; Recio, I.; Sanchez-Moreno, C.; de Pascual-Teresa, B.; Rimbach, G.; de Pascual-Teresa, S. Potential anti-inflammatory, anti-adhesive, anti/estrogenic, and angiotensin-converting enzyme inhibitory activities of anthocyanins and their gut metabolites. Genes Nutr. 2012, 7, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Edwards, M.; Czank, C.; Woodward, G.M.; Cassidy, A.; Kay, C.D. Phenolic metabolites of anthocyanins modulate mechanisms of endothelial function. J. Agric. Food Chem. 2015, 63, 2423–2431. [Google Scholar] [CrossRef] [PubMed]
- Chalopin, M.; Tesse, A.; Martínez, M.C.; Rognan, D.; Arnal, J.F.; Andriantsitohaina, R. Estrogen receptor alpha as a key target of red wine polyphenols action on the endothelium. PLoS ONE 2010, 5, e8554. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.X.; Jiao, J.H.; Li, Z.Y.; Liu, R.R.; Shi, Q.; Ma, L. Lutein supplementation reduces plasma lipid peroxidation and C-reactive protein in healthy nonsmokers. Atherosclerosis 2013, 227, 380–385. [Google Scholar] [CrossRef]
- Verma, S.; Wang, C.H.; Li, S.H.; Dumont, A.S.; Fedak, P.W.M.; Badiwala, M.V.; Dhillon, B.; Weisel, R.D.; Li, R.K.; Mickle, D.A.; et al. A self-fulfilling prophecy: C-reactive protein attenuates nitric oxide production and inhibits angiogenesis. Circulation 2002, 106, 913–919. [Google Scholar] [CrossRef]
- Belwal, T.; Nabavi, S.F.; Nabavi, S.M.; Habtemariam, S. Dietary Anthocyanins and Insulin Resistance: When Food Becomes a Medicine. Nutrients 2017, 9, 1111. [Google Scholar] [CrossRef]
- de Pascual-Teresa, S. Molecular mechanisms involved in the cardiovascular and neuroprotective effects of anthocyanins. Arch. Biochem. Biophys. 2014, 559, 68–74. [Google Scholar] [CrossRef]
- Silvan, J.M.; Mingo, E.; Hidalgo, M.; de Pascual-Teresa, S.; Carrascosa, A.V.; Martinez-Rodriguez, A.J. Antibacterial activity of grape seed extract against Campylobacter spp. Food Control 2013, 29, 25–31. [Google Scholar] [CrossRef]
- Lester, G.E.; Lewers, K.S.; Medina, M.B.; Saftner, R.A. Comparative analysis of strawberry total phenolics via Fast Blue BB vs. Folin-Ciocalteu Assay interference by ascorbic acid. J. Food Compos. Anal. 2012, 27, 102–107. [Google Scholar] [CrossRef]
- Soriano-Maldonado, A.; Hidalgo, M.; Arteaga, P.; de Pascual-Teresa, S.; Nova, E. Effects of regular consumption of vitamin C-rich or polyphenol-rich apple juice on cardiometabolic markers in healthy adults: A randomized crossover trial. Eur. J. Nutr. 2014, 53, 1645–1657. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S.; Feunang, Y.D.; Marcu, A.; Guo, A.C.; Liang, K.; Vázquez-Fresno, R.; Sajed, T.; Johnson, D.; Li, C.; Karu, N.; et al. HMDB 4.0, the human metabolome database for 2018. Nucleic Acids Res. 2018, 46, D608–D617. [Google Scholar] [CrossRef] [PubMed]
- Olmedilla-Alonso, B.; Estévez-Santiago, R.; Silván, J.M.; Sánchez-Prieto, M.; de Pascual-Teresa, S. Effect of long-term xanthophyll and anthocyanin supplementation on lutein and zeaxanthin serum concentrations and macular pigment optical density in postmenopausal women. Nutrients 2018, 10, 959. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo, R.; Libuy, M.; Feliú, F.; Hasson, D. Oxidative Stress-Related Biomarkers in Essential Hypertension and Ischemia-Reperfusion Myocardial Damage. Dis. Markers. 2013, 35, 773–790. [Google Scholar] [CrossRef]
- Rodrigo, R.; Bächler, J.P.; Araya, J.; Prat, H.; Passalacqua, W. Relationship between (Na + K)-ATPase activity, lipid peroxidation and fatty acid profile in erythrocytes of hypertensive and normotensive subjects. Mol. Cell. Biochem. 2007, 303, 73–81. [Google Scholar] [CrossRef]
- Parenica, J.; Nemec, P.; Tomandl, J.; Ondrasek, J.; Pavkova-Goldbergova, M.; Tretina, M.; Jarkovsky, J.; Littnerova, S.; Poloczek, M.; Pokorny, P.; et al. Prognostic utility of biomarkers in predicting of one-year outcomes in patients with aortic stenosis treated with transcatheter or surgical aortic valve implantation. PLoS ONE 2012, 7, e48851. [Google Scholar] [CrossRef][Green Version]
- Martin, M.A.; Goya, L.; Ramos, S. Antidiabetic actions of cocoa flavanols. Mol. Nutr. Food Res. 2016, 60, 1756–1769. [Google Scholar] [CrossRef]
- de Mello, V.D.; Lankinen, M.A.; Lindström, J.; Puupponen-Pimiä, R.; Laaksonen, D.E.; Pihlajamäki, J.; Lehtonen, M.; Uusitupa, M.; Tuomilehto, J.; Kolehmainen, M.; et al. Fasting serum hippuric acid is elevated after bilberry (Vaccinium myrtillus) consumption and associates with improvement of fasting glucose levels and insulin secretion in persons at high risk of developing type 2 diabetes. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef]
- van Oostrom, A.J.; van Wijk, J.; Cabezas, M.C. Lipaemia, inflammation and atherosclerosis: Novel opportunities in the understanding and treatment of atherosclerosis. Drugs 2004, 64, 19–41. [Google Scholar] [CrossRef] [PubMed]
- Hinojosa-Nogueira, D.; Muros, J.; Rufián-Henares, J.A.; Pastoriza, S. New Method to estimate total polyphenol excretion: Comparison of Fast Blue BB versus Folin-Ciocalteu performance in urine. J. Agric. Food Chem. 2017, 65, 4216–4222. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.J.; Beno, D.W.; Zhang, X.; Shapiro, R.; Mason, M.; Mason-Bright, T.; Surber, B.; Edens, N.K. A (14) C-leucine absorption, distribution, metabolism and excretion (ADME) study in adult Sprague-Dawley rat reveals β-hydroxy-β-methylbutyrate as a metabolite. Amino Acids 2015, 47, 917–924. [Google Scholar] [CrossRef] [PubMed]
- Neis, E.P.J.G.; Dejong, C.H.C.; Rensen, S.S. The Role of Microbial Amino Acid Metabolism in Host Metabolism. Nutrients 2015, 7, 2930–2946. [Google Scholar] [CrossRef] [PubMed]
- Loke, W.M.; Jenner, A.M.; Proudfoot, J.M.; McKinley, A.J.; Hodgson, J.M.; Halliwell, B.; Croft, K.D. A metabolite profiling approach to identify biomarkers of flavonoid intake in humans. J. Nutr. 2009, 139, 2309–2314. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.B.; Sun, L.Y.; Gong, Z.D.; Du, Y. Veratric Acid Inhibits LPS-Induced IL-6 and IL-8 Production in Human Gingival Fibroblasts. Inflammation 2016, 39, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Rechner, A.R.; Kuhnle, G.; Hu, H.; Roedig-Penman, A.; van den Braak, M.H.; Moore, K.P.; Rice-Evans, C.A. The metabolism of dietary polyphenols and the relevance to circulating levels of conjugated metabolites. Free Radic. Res. 2002, 36, 1229–1241. [Google Scholar] [CrossRef]
- Jacobs, D.M.; Spiesser, L.; Garnier, M.; de Roo, N.; van Dorsten, F.; Hollebrands, B.; van Velzen, E.; Draijer, R.; van Duynhoven, J. SPE-NMR metabolite sub-profiling of urine. Anal. Bioanal. Chem. 2012, 404, 2349–2361. [Google Scholar] [CrossRef]
- Gonthier, M.P.; Rios, L.Y.; Verny, M.; Rémésy, C.; Scalbert, A. Novel liquid chromatography-electrospray ionization mass spectrometry method for the quantification in human urine of microbial aromatic acid metabolites derived from dietary polyphenols. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2003, 89, 247–255. [Google Scholar] [CrossRef]
- Jaglin, M.; Rhimi, M.; Philippe, C.; Pons, N.; Bruneau, A.; Goustard, B.; Daugé, V.; Maguin, E.; Naudon, L.; Rabot, S. Indole, a signaling molecule produced by the gut microbiota, negatively impacts emotional behaviors in rats. Front. Neurosci. 2018, 12, 216. [Google Scholar] [CrossRef]
- Hidalgo, M.; Oruna-Concha, M.J.; Kolida, S.; Walton, G.E.; Kallithraka, S.; Spencer, J.P.; de Pascual-Teresa, S. Metabolism of anthocyanins by human gut microflora and their influence on gut bacterial growth. J. Agric. Food Chem. 2012, 60, 3882–3890. [Google Scholar] [CrossRef] [PubMed]
- Lear, R.; O’Leary, M.; O’Brien Andersen, L.; Holt, C.C.; Stensvold, C.R.; van der Giezen, M.; Bowtell, J.L. Tart Cherry Concentrate Does Not Alter the Gut Microbiome, Glycaemic Control or Systemic Inflammation in a Middle-Aged Population. Nutrients 2019, 11, 1063. [Google Scholar] [CrossRef] [PubMed]
- Boto-Ordóñez, M.; Urpi-Sarda, M.; Queipo-Ortuño, M.; Corella, D.; Tinahones, F.J.; Estruch, R.; Andres-Lacueva, C. Microbial metabolomic fingerprinting in urine after regular dealcoholized red wine consumption in humans. J. Agric. Food Chem. 2013, 61, 9166–9175. [Google Scholar] [CrossRef] [PubMed]
- Boto-Ordóñez, M.; Urpi-Sarda, M.; Queipo-Ortuño, M.; Tulipani, S.; Tinahones, F.J.; Andres-Lacueva, C. High levels of Bifidobacteria are associated with increased levels of anthocyanin microbial metabolites: A randomized clinical trial. Food Funct. 2014, 5, 1932–1938. [Google Scholar] [CrossRef] [PubMed]
- Mayta-Apaza, A.C.; Pottgen, E.; De Bodt, J.; Papp, N.; Marasini, D.; Howard, L.; Abranko, L.; Van de Wiele, T.; Lee, S.O.; Carbonero, F. Impact of tart cherries polyphenols on the human gut microbiota and phenolic metabolites in vitro and in vivo. J. Nutr. Biochem. 2018, 59, 160–172. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Variable | Group A (n = 23) | Group X (n = 26) | Group A+X (n = 23) | Total (n = 72) |
|---|---|---|---|---|
| Age (years) | 58 ± 6 | 60 ± 6 | 60 ± 5 | 59 ± 6 |
| BMI (kg/m2) | 24.9 ± 3.3 | 24.8 ± 2.8 | 24.5 ± 2.7 | 24.8 ± 2.9 |
| Cholesterol (mmol/L) | 5.41 ± 1.00 | 5.45 ± 0.69 | 5.75 ± 0.58 | 5.53 ± 0.78 |
| HDL- cholesterol (mmol/L) | 1.64 ± 0.33 | 1.72 ± 0.33 | 1.85 ± 0.35 | 1.73 ± 0.34 |
| LDL-cholesterol (mmol/L) | 3.77 ± 1.10 | 3.73 ± 0.77 | 3.91 ± 0.61 | 3.80 ± 0.84 |
| TG (mmol/L) | 1.04 ± 0.37 | 0.95 ± 0.38 | 0.92 ± 0.30 | 0.97 ± 0.35 |
| Systolic blood pressure, (mm Hg) | 118 ± 17 | 115 ± 14 | 121 ± 12 | 118 ± 15 |
| Diastolic blood pressure, (mm Hg) | 76 ± 11 | 74 ± 10 | 79 ± 9 | 76 ± 10 |
| Glucose (mmol/L) | 4.98 ± 0.47 | 5.13 ± 0.41 | 4.89 ± 0.38 | 5.01 ± 0.43 |
| Group A | Group X | Group A+X | |
|---|---|---|---|
| Mean ± SD | |||
| Total Polyphenols (FB as g/L GAeq) – baseline | 0.142 ± 0.028 a | 0.138 ± 0.029 a | 0.146 ± 0.018 a |
| 4 m | 0.169 ± 0.026 b * | 0.162 ± 0.032 b | 0.155 ± 0.036 a |
| 8 m | 0.159 ± 0.029 ab | 0.153 ± 0.024 ab | 0.151 ± 0.034 a |
| Total Polyphenols (F g/L GAeq) – baseline | 0.249 ± 0.045 a | 0.272 ± 0.066 a | 0.247 ± 0.037 a |
| 4 m | 0.217 ± 0.077 a | 0.221 ± 0.084 b | 0.224 ± 0.053 a |
| 8 m | 0.209 ± 0.031 a | 0.237 ± 0.072 ab | 0.220 ± 0.060 a |
| Vitamin C (µmol/L) – baseline | 52.79 ± 13.40 a | 50.42 ± 8.4 a | 49.83 ± 12.47 a |
| 4 m | 48.44 ± 16.12 a | 50.90 ± 13.31 a | 54.66 ± 19.14 a |
| 8 m | 60.94 ± 14.90 b | 58.41 ± 17.83 a | 55.78 ± 19.15 a |
| FRAP (Trolox eq) – baseline | 588 ± 78 a | 568 ± 75 a | 569 ± 75 a |
| 4 m | 704 ± 117 b * | 677 ± 95 b | 650 ± 116 b * |
| 8 m | 687 ± 136 b | 661 ± 72 b | 642 ± 115 b |
| Group A | Group X | Group A+X | |
|---|---|---|---|
| Mean ± SD | |||
| SBP (mm Hg) – baseline | 118 ± 17 | 115 ± 14 | 121 ± 12 |
| 4 m | 118 ± 17 | 115 ± 17 | 118 ± 15 |
| 8 m | 120 ± 16 | 113 ± 13 | 120 ± 12 |
| DBP (mm Hg) – baseline | 76 ± 11 | 74 ± 10 | 79 ± 9 |
| 4 m | 75 ± 9 | 74 ± 10 | 78 ± 9 |
| 8 m | 76 ± 11 | 74 ± 9 | 76 ± 10 |
| Glucose (mmol/L) – baseline | 4.98 ± 0.47 | 5.13 ± 0.41 | 4.88 ± 0.38 |
| 4 m | 5.12 ± 0.50 | 5.12 ± 0.37 | 4.98 ± 0.37 |
| 8 m | 5.12 ± 0.37 | 5.07 ± 0.56 | 4.75 ± 0.33 * |
| Insulin (pmol/L) – baseline | 49.3 ± 21.5 | 50.7 ± 29.9 | 41.0 ± 14.0 |
| 4 m | 50.7 ± 26.4 | 52.8 ± 27.1 | 47.2 ± 23.6 |
| 8 m | 55.6 ± 23.6 | 50.7 ± 25.0 | 45.8 ± 22.2 |
| C3 (g/L) – baseline | 1.02 ± 0.02 | 1.06 ± 0.01 | 1.00 ± 0.01 |
| 4 m | 1.09 ± 0.02 | 1.14 ± 0.02 | 1.04 ± 0.01 |
| 8 m | 1.09 ± 0.02 | 1.06 ± 0.02 | 0.98 ± 0.01 * |
| C4 (g/L) – baseline | 0.29 ± 0.01 | 0.29 ± 0.01 | 0.30 ± 0.01 |
| 4 m | 0.31 ± 0.01 | 0.30 ± 0.01 | 0.29 ± 0.01 |
| 8 m | 0.30 ± 0.01 | 0.29 ± 0.01 | 0.26 ± 0.01 |
| CRP (mg/L) – baseline | 1.23 ± 0.89 | 1.50 ± 1.78 | 1.38 ± 1.32 |
| 4 m | 1.78 ± 1.90 | 1.77 ± 2.18 | 1.53 ± 1.99 |
| 8 m | 1.26 ± 1.15 | 1.40 ± 1.23 | 1.22 ± 1.48 |
| IL-6 (pg/mL) – baseline | 0.8 ± 0.2 | 1.0 ± 0.6 | 1.1 ± 0.5 |
| 8 m | 1.4 ± 1.9 | 1.1 ± 0.8 | 1.2 ± 0.9 |
| ICAM-1 (ng/mL) – baseline | 381 ± 208 | 344 ± 179 | 346 ± 192 |
| 8 m | 351 ± 122 | 320 ± 132 | 345 ± 106 |
| VCAM-1 (ng/mL) – baseline | 185 ± 76 | 174 ± 62 | 152 ± 52 |
| 8 m | 166 ± 54 | 160 ± 66 | 149 ± 83 |
| MCP-1 (pg/mL) – baseline | 126 ± 29 | 127 ± 30 | 137 ± 40 |
| 8 m | 135 ± 65 | 137 ± 67 | 137 ± 48 |
| MMP2 (µg/mL) – baseline | 215 ± 63 | 257 ± 110 | 275 ± 161 |
| 8 m | 222 ± 79 | 279 ± 115 | 275 ± 120 |
| MMP9 (µg/mL) – baseline | 93 ± 30 | 98 ± 42 | 85 ± 39 |
| 8 m | 117 ± 43 | 104 ± 33 | 113 ± 38 |
| Theoretical Mass (m/z) | RT | pA | pX | pA+X | A | X | A+X |
|---|---|---|---|---|---|---|---|
| 112.017 | 4.4 | 0.0251 | |||||
| 112.026 | 3.3 | 0.0044 | |||||
| 126.032 | 5.4 | 0.0206 | |||||
| 136.038 | 3.1 | 0.0394 | |||||
| 138.043 | 3.1 | 0.0271 | 0.0337 | ||||
| 166.050 | 3.0 | 0.0428 | 0.0479 | ||||
| 173.104 | 12.2 | 0.0024 | 0.0200 | 0.0000 | |||
| 174.018 | 3.3 | 0.0473 | 0.0244 | ||||
| 174.008 | 21.6 | 0.0099 | |||||
| 182.058 | 11.1 | 0.0353 | |||||
| 187.120 | 18.7 | 0.0223 | 0.0066 | ||||
| 188.104 | 29.3 | 0.0087 | 0.0123 | ||||
| 195.050 | 10.2 | 0.0325 | |||||
| 201.136 | 25.9 | 0.0016 | 0.0055 | 0.0000 | |||
| 215.152 | 33.0 | 0.0105 | |||||
| 219.057 | 5.4 | 0.0484 | |||||
| 236.934 | 2.1 | 0.0213 | |||||
| 244.071 | 3.3 | 0.0254 | |||||
| 306.059 | 4.4 | 0.0224 | |||||
| 308.924 | 5.1 | 0.0486 | |||||
| 318.154 | 2.7 | 0.0212 | |||||
| 340.234 | 37.7 | 0.0003 | 0.0073 | ||||
| 375.228 | 14.4 | 0.0024 | |||||
| 403.925 | 2.1 | 0.0272 | |||||
| 582.398 | 21.5 | 0.0382 | |||||
| 711.357 | 19.7 | 0.0373 | |||||
| 785.427 | 9.5 | 0.0493 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Estévez-Santiago, R.; Silván, J.M.; Can-Cauich, C.A.; Veses, A.M.; Alvarez-Acero, I.; Martinez-Bartolome, M.A.; San-Román, R.; Cámara, M.; Olmedilla-Alonso, B.; de Pascual-Teresa, S. Lack of a Synergistic Effect on Cardiometabolic and Redox Markers in a Dietary Supplementation with Anthocyanins and Xanthophylls in Postmenopausal Women. Nutrients 2019, 11, 1533. https://doi.org/10.3390/nu11071533
Estévez-Santiago R, Silván JM, Can-Cauich CA, Veses AM, Alvarez-Acero I, Martinez-Bartolome MA, San-Román R, Cámara M, Olmedilla-Alonso B, de Pascual-Teresa S. Lack of a Synergistic Effect on Cardiometabolic and Redox Markers in a Dietary Supplementation with Anthocyanins and Xanthophylls in Postmenopausal Women. Nutrients. 2019; 11(7):1533. https://doi.org/10.3390/nu11071533
Chicago/Turabian StyleEstévez-Santiago, Rocío, José Manuel Silván, Cesar Abraham Can-Cauich, Ana Maria Veses, Inma Alvarez-Acero, Miguel Angel Martinez-Bartolome, Ricardo San-Román, Montaña Cámara, Begoña Olmedilla-Alonso, and Sonia de Pascual-Teresa. 2019. "Lack of a Synergistic Effect on Cardiometabolic and Redox Markers in a Dietary Supplementation with Anthocyanins and Xanthophylls in Postmenopausal Women" Nutrients 11, no. 7: 1533. https://doi.org/10.3390/nu11071533
APA StyleEstévez-Santiago, R., Silván, J. M., Can-Cauich, C. A., Veses, A. M., Alvarez-Acero, I., Martinez-Bartolome, M. A., San-Román, R., Cámara, M., Olmedilla-Alonso, B., & de Pascual-Teresa, S. (2019). Lack of a Synergistic Effect on Cardiometabolic and Redox Markers in a Dietary Supplementation with Anthocyanins and Xanthophylls in Postmenopausal Women. Nutrients, 11(7), 1533. https://doi.org/10.3390/nu11071533