Purple Sweet Potato Polyphenols Differentially Influence the Microbial Composition Depending on the Fermentability of Dietary Fiber in a Mixed Culture of Swine Fecal Bacteria

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation and Determination of Sweet Potato Polyphenols
2.2. Feces and In Vitro Fermentation
2.3. Bacterial Analysis
2.4. DNA Extraction and 16S Ribosomal RNA (16S rRNA) Gene Sequences
2.5. Short-Chain Fatty Acid (SCFA) Analysis
2.6. Measurement of Putrefactive Products
2.7. Statistical Analysis
3. Results
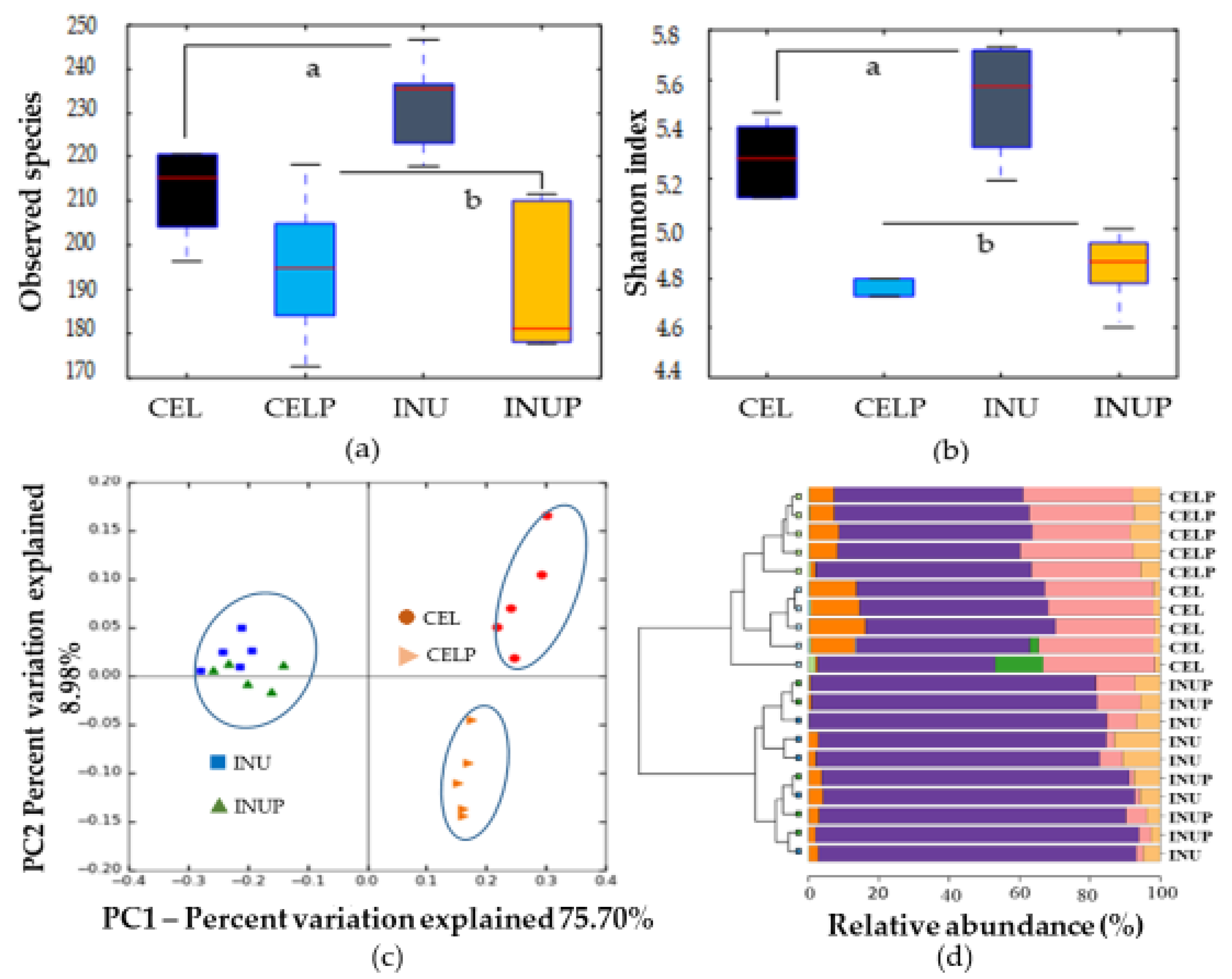
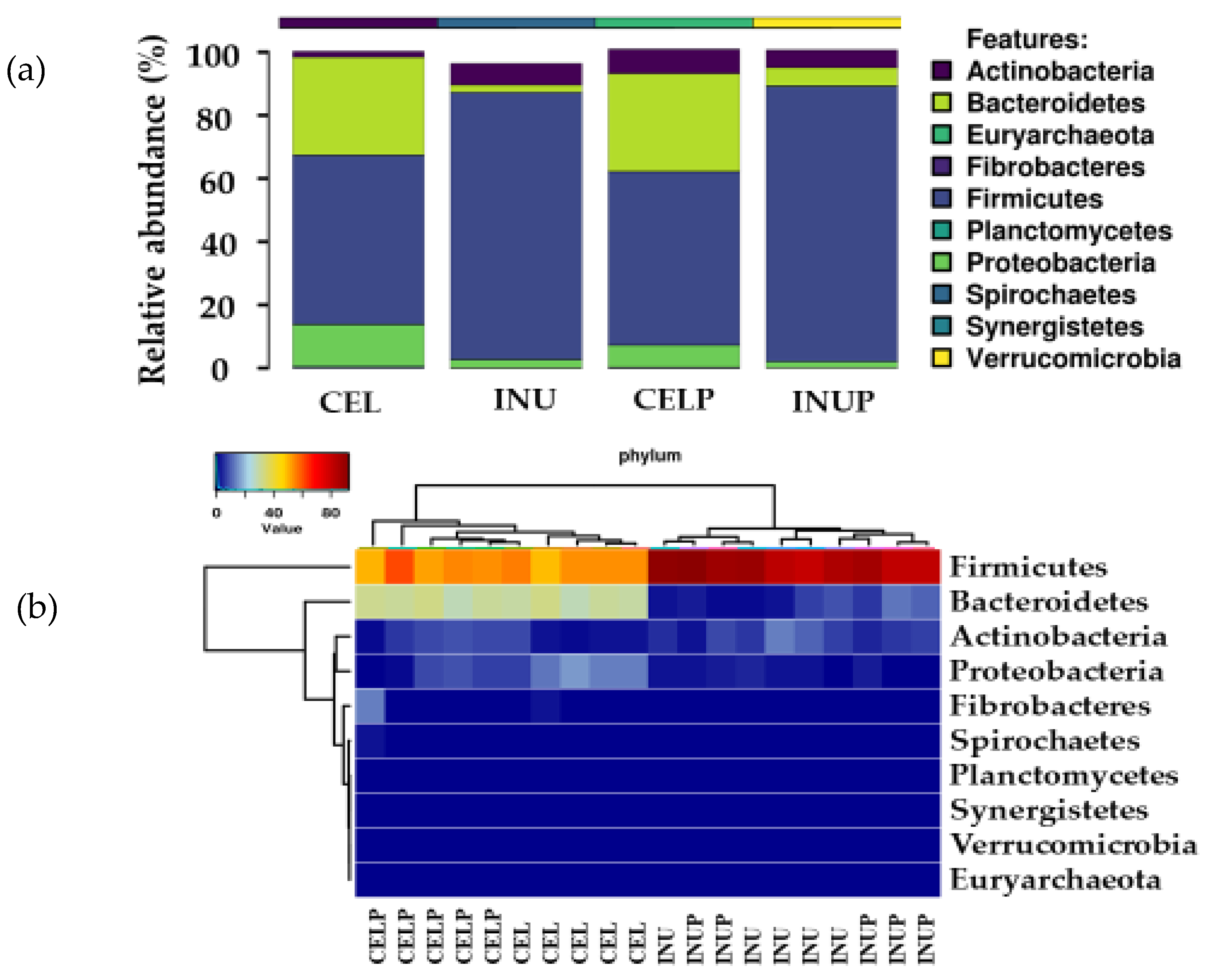
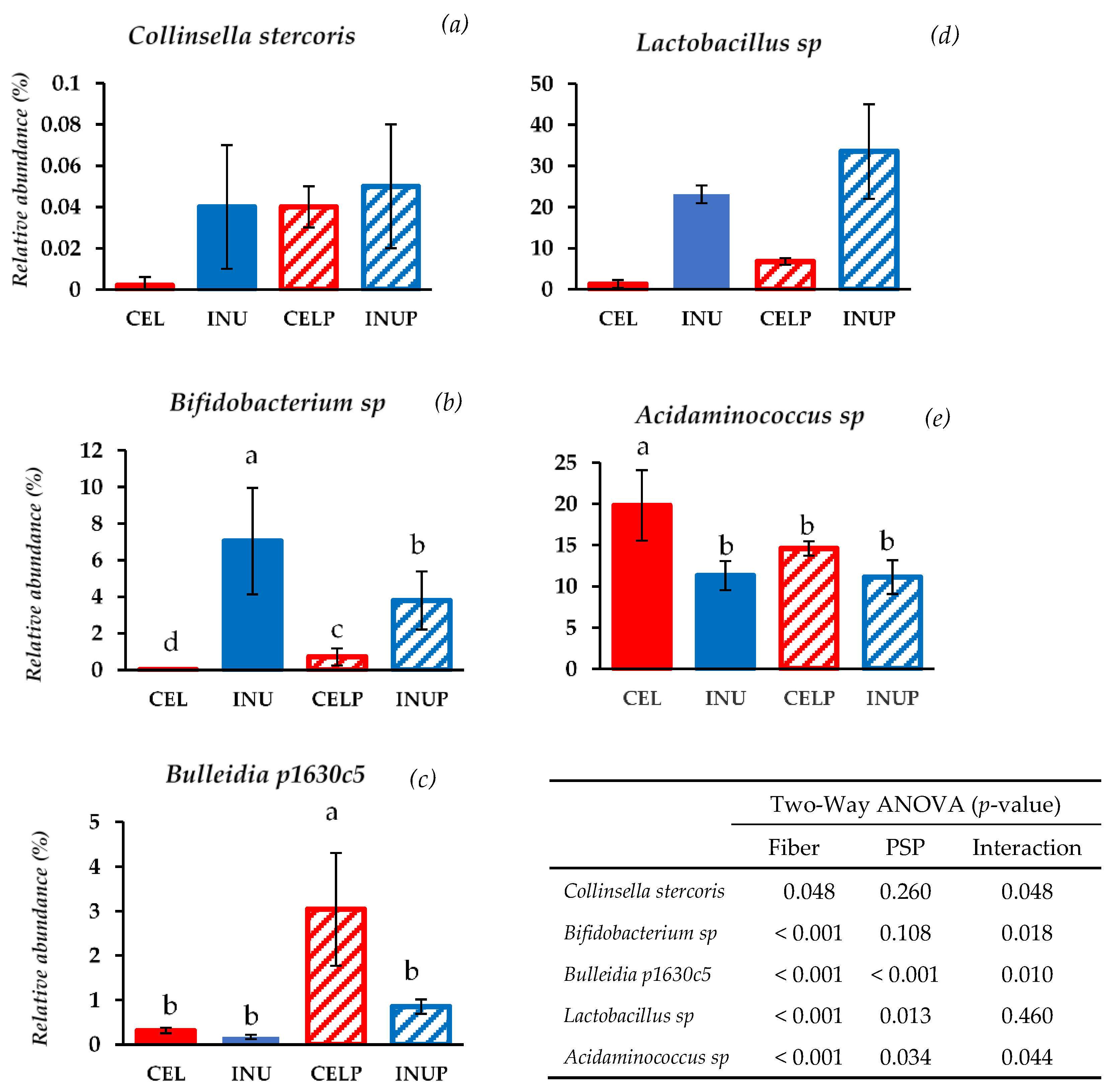
3.1. Gut Microbial Taxonomic Analysis
3.2. SCFA Concentration in Fermenters
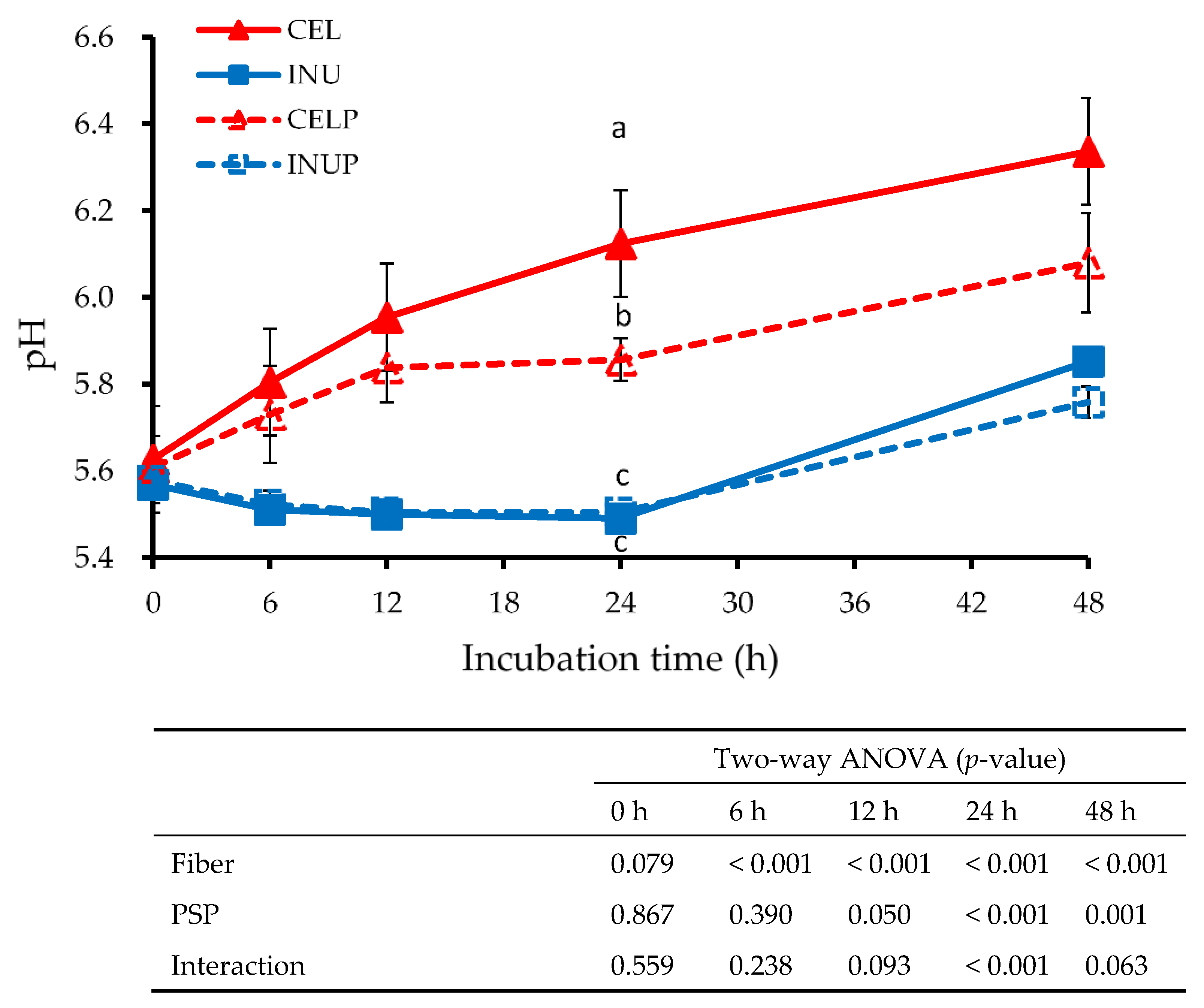
3.3. pH in Fermenters
3.4. Putrefactive Products in Fermenters
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Drewnowski, A.; Popkin, B.M. The nutrition transition: New trends in the global diet. Nutr. Rev. 1997, 55, 31–43. [Google Scholar] [CrossRef] [PubMed]
- Ferro-Luzzi, A.; Martino, L. Obesity and physical activity. Ciba Found. Symp. 1996, 201, 207–221. [Google Scholar] [PubMed]
- Van Dam, R.M.; Rimm, E.B.; Willett, W.C.; Stampfer, M.J.; Hu, F.B. Dietary patterns and risk for type 2 diabetes mellitus in U.S. men. Ann. Intern. Med. 2002, 136, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Kerver, J.M.; Yang, E.J.; Bianchi, L.; Song, W.O. Dietary patterns associated with risk factors for cardiovascular disease in healthy US adults. Am. J. Clin. Nutr. 2003, 78, 1103–1110. [Google Scholar] [CrossRef] [PubMed]
- Kobyliak, N.; Conte, C.; Cammarota, G.; Haley, A.P.; Styriak, I.; Gaspar, L.; Fusek, J.; Rodrigo, L.; Kruzliak, P. Probiotics in prevention and treatment of obesity: A critical view. Nutr. Metab. 2016, 13, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Xenoulis, P.G.; Palculict, B.; Allenspach, K.; Steiner, J.M.; Van House, A.M.; Suchodolski, J.S. Molecular-phylogenetic characterization of microbial communities’ imbalances in the small intestine of dogs with inflammatory bowel disease. FEMS Microbiol. Ecol. 2008, 66, 579–589. [Google Scholar] [CrossRef]
- Eckburg, P.B.; Bik, E.M.; Bernstein, C.N.; Purdom, E.; Dethlefsen, L.; Sargent, M.; Gill, S.R.; Nelson, K.E.; Relman, D.A. Diversity of the human intestinal microbial flora. Science 2005, 308, 1635–1638. [Google Scholar] [CrossRef]
- Graf, D.; Di Cagno, R.; Fåk, F.; Flint, H.J.; Nyman, M.; Saarela, M.; Watzl, B. Contribution of diet to the composition of the human gut microbiota. Microb. Ecol. Health Dis. 2015, 26, 26146. [Google Scholar] [CrossRef]
- Dicksved, J.; Halfvarson, J.; Rosenquist, M.; Jarnerot, G.; Tysk, C.; Apajalahti, J.; Engstrand, L.; Jansson, J.K. Molecular analysis of the gut microbiota of identical twins with Crohn’s disease. ISME J. 2008, 2, 716–727. [Google Scholar] [CrossRef]
- Frank, D.N.; St Amand, A.L.; Feldman, R.A.; Boedeker, E.C.; Harpaz, N.; Pace, N.R. Molecular-phylogenetic characterization of microbial community imbalances in human inflammatory bowel diseases. Proc. Natl. Acad. Sci. USA 2007, 104, 13780–13785. [Google Scholar] [CrossRef]
- Gibson, G.R.; Probert, H.M.; Loo, J.V.; Rastall, R.A.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota: Updating the concept of prebiotics. Nutr. Res. Rev. 2004, 17, 259–275. [Google Scholar] [CrossRef] [PubMed]
- Cherbut, C. Inulin and oligofructose in the dietary fibre concept. Br. J. Nutr. 2002, 87, 159–162. [Google Scholar] [CrossRef] [PubMed]
- McRorie, J.W., Jr. Psyllium is not fermented in the human gut. Neurogastroenterol. Motil. 2015, 27, 1681–1682. [Google Scholar] [CrossRef] [PubMed]
- Puupponen-Pimiä, R.; Aura, A.M.; Oksman-Caldentey, K.M.; Myllärinen, P.; Saarela, M.; Mattila-Sandholm, T.; Poutenen, K. Development of functional ingredients for gut health. Trends Food Sci. Technol. 2002, 13, 3–11. [Google Scholar] [CrossRef]
- Weng, C.J.; Yen, G.C. Chemopreventive effects of dietary phytochemicals against cancer invasion and metastasis: Phenolic acids, monophenol, polyphenol and their derivatives. Cancer Treat. Rev. 2012, 38, 76–87. [Google Scholar] [CrossRef] [PubMed]
- Mursu, J.; Voutilainen, S.; Nurmi, T.; Tuomainen, T.P.; Kurt, S.; Salonen, J.T. Flavonoid intake and the risk of ischaemic stroke and CVD mortality in middle-aged Finnish men: The kuopio ischemic heart disease risk factor study. Br. J. Nutr. 2008, 100, 890–895. [Google Scholar] [CrossRef] [PubMed]
- Xia, D.; Wu, X.; Shi, J.; Yang, Q.; Zhang, Y. Phenolic compounds from the edible seeds extract of Chinese Mei (Prunusmume Sieb. Et Zucc) and their antimicrobial activity. Food Sci. Technol. 2011, 44, 347–349. [Google Scholar]
- Lim, S.; Xu, J.; Kim, J.; Chen, T.Y.; Su, X.; Standard, J.; Carey, E.; Griffin, J.; Herndon, B.; Katz, B.; et al. Role of anthocyanin-enriched purple-fleshed sweet potato p40 in colorectal cancer prevention. Mol. Nutr. Food Res. 2013, 57, 1908–1917. [Google Scholar] [CrossRef]
- Han, K.H.; Matsumoto, A.; Shimada, K.; Sekikawa, M.; Fukushima, M. Effects of anthocyanin-rich purple potato flakes on antioxidant status in F344 rats fed a cholesterol-rich diet. Br. J. Nutr. 2007, 98, 914–921. [Google Scholar] [CrossRef]
- Jawi, M.; Wita, W.; Suprapta, D.N. Extract of purple sweet potato tuber increases Sod and decreases VCAM-1 Expression by increasing Nrf2 expression in the aortic endothelia of hypercholesterolemic rabbits. J. Biol. Agric. Healthcare 2014, 4, 76–84. [Google Scholar]
- Kano, M.; Takayanagi, T.; Harada, K.; Makino, K.; Ishikawa, F. Antioxidative activity of anthocyanins from purple sweet potato, Ipomea batatas cultivar Ayamurasaki. Biosci. Biotechnol. Biochem. 2005, 69, 979–988. [Google Scholar] [CrossRef] [PubMed]
- Han, K.H.; Sekikawa, M.; Shimada, K.; Hashimoto, M.; Hashimoto, N.; Noda, T.; Tanaka, H.; Fukushima, M. Anthocyanin-rich purple potato flake extract has antioxidant capacity and improves antioxidant potential in rats. Br. J. Nutr. 2006, 96, 1125–1133. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total polyphenol and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Nagata, R.; Echizen, M.; Yamaguchi, Y.; Han, K.H.; Shimada, K.; Ohba, K.; Kitano-Okada, T.; Nagura, T.; Uchino, H.; Fukushima, M. Effect of a combination of inulin and polyphenol containing adzuki bean extract on intestinal fermentation In Vitro and In Vivo. Biosci. Biotechnol. Biochem. 2018, 82, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Morrison, M. Improved extraction of PCR-quality community DNA from digesta and fecal samples. Biotechniques 2004, 36, 808–812. [Google Scholar] [CrossRef] [PubMed]
- Warren, F.J.; Fukuma, N.M.; Mikkelsen, D.; Flanagan, B.M.; Williams, B.A.; Lisle, A.T.; Cuív, P.Ó.; Morrison, M.; Gidley, M.J. Food starch structure impacts gut microbiome composition. Msphere 2018, 3, 86–118. [Google Scholar] [CrossRef]
- Lozupone, C.; Knight, R. UniFrac: A new phylogenetic method for comparing microbial communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef] [PubMed]
- Zakrzewski, M.; Proietti, C.; Ellis, J.J.; Hasan, S.; Brion, M.J.; Berger, B.; Krause, L. Calypso: A user-friendly web-server for mining and visualizing microbiome-environment interactions. Bioinformatics 2017, 33, 782–783. [Google Scholar] [CrossRef]
- Ikeda, T.; Tanaka, Y.; Yamamoto, K.; Morii, H.; Kamisako, T.; Ogawa, H. Geranium Dielsianum extract powder (Miskamiska) improves the intestinal environment through alteration of microbiota and microbial metabolites in rats. J. Funct. Foods 2014, 11, 12–19. [Google Scholar] [CrossRef]
- Heinritz, S.N.; Rainer Mosenthin, R.; Weiss, E. Use of pigs as a potential model for research into dietary modulation of the human gut microbiota. Nutr. Res. Rev. 2013, 26, 191–209. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed]
- Frolinger, T.; Sims, S.; Smith, C.; Wang, J.; Cheng, H.; Faith, J.; Ho, L.; Hao, K.; Pasinetti, G.M. The gut microbiota composition affects dietary polyphenols-mediated cognitive resilience in mice by modulating the bioavailability of phenolic acids. Sci. Rep. 2019, 9, 3546. [Google Scholar] [CrossRef] [PubMed]
- Espley, R.V.; Butts, C.A.; Laing, W.A.; Martell, S.; Smith, H.; McGhie, T.K.; Zhang, J.; Paturi, G.; Hedderley, D.; Bovy, A.; et al. Dietary flavonoids from modified apple reduce inflammation markers and modulate gut microbiota in mice. J. Nutr. 2014, 144, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Kovatcheva-Datchary, P.; Nilsson, A.; Akrami, R.; Lee, Y.S.; De Vadder, F.; Arora, T.; Hallen, A.; Martens, E.; Björck, I.; Bäckhed, F. Dietary fiber-induced improvement in glucose metabolism is associated with increased abundance of Prevotella. Cell Metab. 2015, 22, 971–982. [Google Scholar] [CrossRef] [PubMed]
- Canard, B.; Cole, S.T. Genome organization of the anaerobic pathogen Clostridium perfringens. Proc. Natl. Acad. Sci. USA. 1989, 86, 6676–6680. [Google Scholar] [CrossRef] [PubMed]
- Koropatkin, N.M.; Cameron, E.A.; Martens, E.C. How glycan metabolism shapes the human gut microbiota. Nat. Rev. Microbiol. 2012, 10, 323–335. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yang, Y.; Wu, Z.; Weng, P. The modulatory effect of anthocyanins from purple sweet potato on human intestinal microbiota in vitro. J. Agric. Food Chem. 2016, 64, 2582–2590. [Google Scholar] [CrossRef]
- Molan, A.L.; Liu, Z.; Kruger, M. The ability of blackcurrant extracts to positively modulate key markers of gastrointestinal function in rats. World, J. Microbiol. Biotechnol. 2010, 26, 1735–1743. [Google Scholar] [CrossRef]
- Scharlau, D.; Borowicki, A.; Habermann, N.; Hofmann, T.; Klenow, S.; Miene, C.; Munjal, U.; Stein, K.; Glei, M. Mechanisms of primary cancer prevention by butyrate and other products formed during gut flora-mediated fermentation of dietary fibre. Mutat. Res. 2009, 682, 39–53. [Google Scholar] [CrossRef]
- Crouse, J.R.; Gerson, C.D.; DeCarli, L.M.; Lieber, C.S. Role of acetate in the reduction of plasma free fatty acids produced by ethanol in man. J. Lipid Res. 1968, 9, 509–512. [Google Scholar]
- Nastasi, C.; Candela, M.; Bonefeld, C.M.; Geisler, C.; Hansen, M.; Krejsgaard, T.; Biagi, E.; Andersen, M.H.; Brigidi, P.; θdum, N.; et al. The effect of short-chain fatty acids on human monocyte-derived dendritic cells. Sci. Rep. 2015, 5, 16148. [Google Scholar] [CrossRef] [PubMed]
- Maeda, T.; Towatari, M.; Kosugi, H.; Saito, H. Up-regulation of costimulatory/adhesion molecules by histone deacetylase inhibitors in acute myeloid leukemia cells. Blood 2000, 96, 3847–3856. [Google Scholar] [PubMed]
- Kim, H.J.; White, P.J. In vitro fermentation of oat flours from typical and high β-glucan oat lines. J. Agric. Food Chem. 2009, 57, 7529–7536. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.L.; Zhang, X.; Sun, Y.K.; Su, D.; Sun, Y.; Hu, B.; Zeng, X.X. Purification and fermentation in vitro of sesaminol triglucoside from sesame cake by human intestinal microbiota. J. Agric. Food Chem. 2013, 61, 1868–1877. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota: Introducing the concept of prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [CrossRef] [PubMed]
- Davila, A.M.; Blachier, F.; Gotteland, M.; Andriamihaja, M.; Benetti, P.H.; Sanz, Y.; Tomé, D. Intestinal luminal nitrogen metabolism: Role of the gut microbiota and consequences for the host. Parmacol. Res. 2013, 68, 95–107. [Google Scholar] [CrossRef]
- Macfarlane, G.T.; Macfarlane, S. Human colonic microbiota: Ecology, physiology and metabolic potential of intestinal bacteria. Scand. J. Gastroenterol. Suppl. 1997, 222, 3–9. [Google Scholar] [CrossRef]
- Vanholder, R.; Schepers, E.; Pletinck, A.; Nagler, E.V.; Glorieux, G. The uremic toxicity of indoxyl sulfate and p-cresyl sulfate: A systematic review. J. Am. Soc. Nephrol. 2014, 25, 1897–1907. [Google Scholar] [CrossRef]
- Brocca, A.; Virzì, G.M.; de Cal, M.; Cantaluppi, V.; Ronco, C. Cytotoxic effects of p-cresol in renal epithelial tubular cells. Blood Purif. 2013, 36, 219–225. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
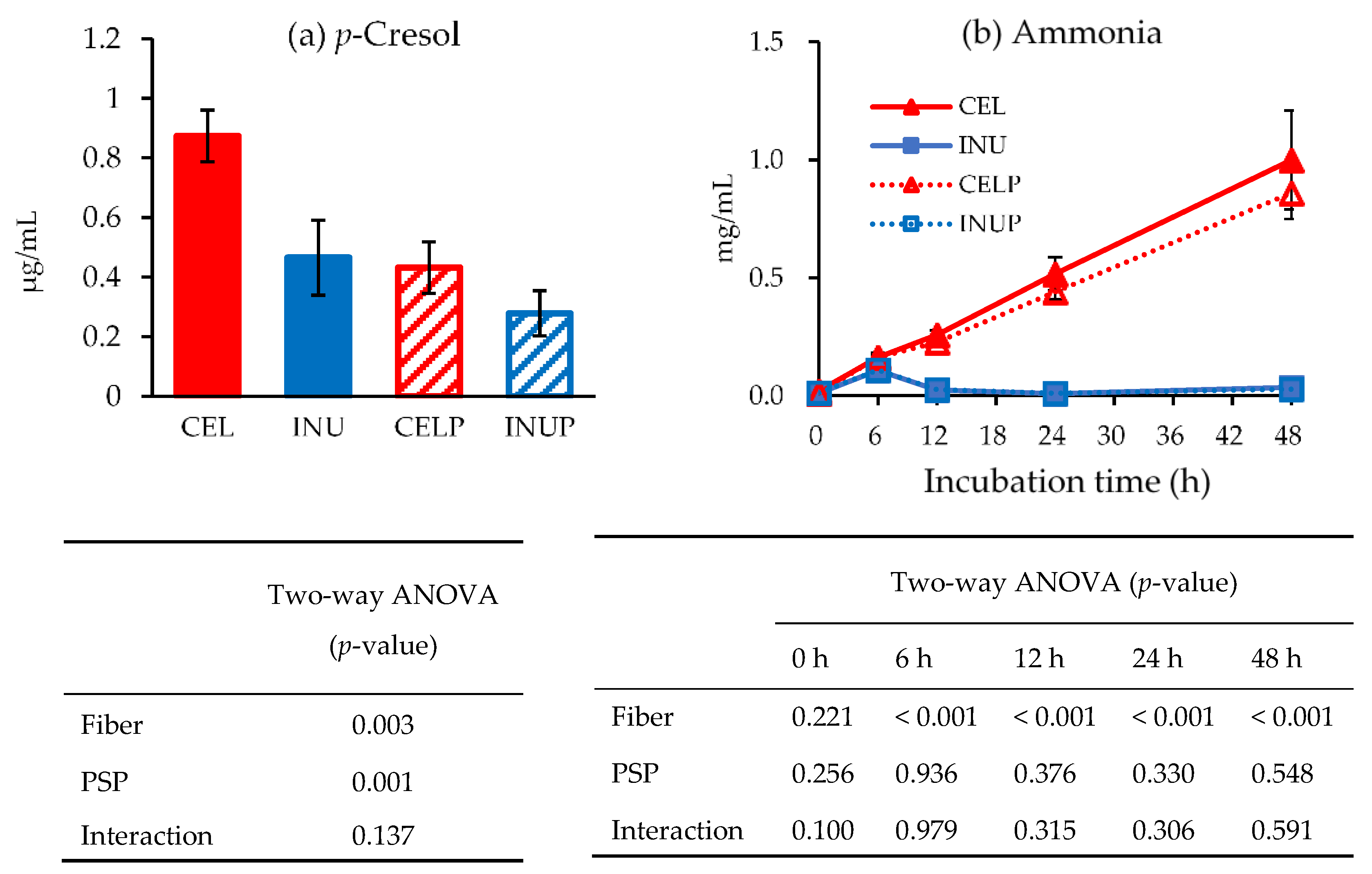{kind=link}
| Groups | Two-Way ANOVA (p-Value) | ||||||
|---|---|---|---|---|---|---|---|
| CEL | INU | CELP | INUP | Fiber | PSP | Interaction | |
| Anaerobes (log10 CFU mL−1) | 8.20 ± 0.10 | 8.75 ± 0.10 | 8.70 ± 0.10 | 8.90 ± 0.10 | 0.001 | 0.003 | 0.080 |
| Lactobacillus (log10 CFU mL−1) | 6.57 ± 0.30 b | 7.25 ± 0.20 ab | 7.17 ± 0.10 ab | 7.86 ± 0.10 a | 0.002 | 0.005 | 0.048 |
| Genus (Relative abundance, %) | |||||||
| Bacteroides | 1.74 ± 0.30 | 20.2 ± 1.6 | 3.79 ± 1.20 | 16.9 ± 6.5 | <0.001 | 0.409 | 0.302 |
| Prevotella | 20.0 ± 2.4 | 3.64 ± 2.90 | 20.0 ± 2.4 | 6.39 ± 4.70 | <0.001 | 0.008 | 0.367 |
| Bifidobacterium | 0.01 ± 0.01 | 7.03 ± 2.90 | 0.71 ± 0.50 | 3.78 ± 1.60 | 0.002 | 0.072 | 0.071 |
| Clostridium | 2.18 ± 1.20 a | 0.29 ± 0.10 b | 1.61 ± 0.70 a | 0.33 ± 0.20 b | <0.001 | 0.108 | 0.018 |
| Lactobacillus | 1.27 ± 0.90 | 23.1 ± 2.1 | 6.76 ± 0.80 | 33.5 ± 11.0 | <0.001 | 0.039 | 0.334 |
| Sharpea | 2.01 ± 1.50 | 0.02 ± 0.01 | 0.62 ± 0.60 | 0.02 ± 0.03 | <0.001 | 0.354 | 0.356 |
| Coprococcus | 0.01 ± 0.01 | 0.02 ± 0.01 | 0.01 ± 0.01 | 0.04 ± 0.03 | 0.021 | 0.035 | 0.219 |
| Bulleidia | 0.82 ± 0.50 b | 0.33 ± 0.10 b | 3.42 ± 1.30 a | 1.06 ± 0.10 a | <0.001 | <0.001 | 0.010 |
| Acidaminococcus | 19.8 ± 4.3 a | 11.3 ± 1.8 b | 14.6 ± 0.9 b | 11.1 ± 2.1 b | <0.001 | 0.034 | 0.044 |
| Incubation Time (h) | CEL | INU | CELP | INUP | Two-Way ANOVA (p-Value) | |||
|---|---|---|---|---|---|---|---|---|
| µmol mL−1 | Fiber | PSP | Interaction | |||||
| Acetate | 0 | 1.92 ± 2.00 | 2.00 ± 2.00 | 1.94 ± 0.22 | 1.94 ± 0.19 | 0.840 | 0.935 | 0.865 |
| 6 | 4.54 ± 0.70 | 9.00 ± 1.50 | 5.68 ± 0.60 | 9.36 ± 1.50 | 0.003 | 0.529 | 0.744 | |
| 12 | 8.30 ± 0.70 | 34.2 ± 10.9 | 10.7 ± 0.4 | 31.6 ± 10.5 | 0.007 | 0.992 | 0.748 | |
| 24 | 12.0 ± 1.6 | 126 ± 20 | 16.6 ± 1.0 | 107 ± 25 | <0.001 | 0.672 | 0.482 | |
| 48 | 32.8 ± 9.6 | 200 ± 18 | 32.9 ± 4.4 | 182 ± 27 | <0.001 | 0.591 | 0.588 | |
| Propionate | 0 | 0.57 ± 0.06 | 0.63 ± 0.07 | 0.54 ± 0.11 | 0.68 ± 0.11 | 0.255 | 0.935 | 0.664 |
| 6 | 1.97 ± 0.92 | 2.55 ± 1.86 | 1.90 ± 1.01 | 2.60 ± 1.77 | 0.660 | 0.992 | 0.967 | |
| 12 | 3.97 ± 0.76 | 21.2 ± 12.5 | 5.10 ± 0.30 | 20.0 ± 13.3 | 0.097 | 0.995 | 0.898 | |
| 24 | 5.56 ± 0.85 | 83.8 ± 26.2 | 6.50 ± 0.60 | 65.5 ± 29.1 | 0.003 | 0.663 | 0.629 | |
| 48 | 13.5 ± 4.1 | 178 ± 12 | 12.6 ± 1.4 | 150 ± 23 | <0.001 | 0.290 | 0.319 | |
| n-Butyrate | 0 | 0.11 ± 0.05 | 0.08 ± 0.03 | 0.11 ± 0.05 | 0.07 ± 0.04 | 0.396 | 0.902 | 0.975 |
| 6 | 0.57 ± 0.13 | 1.06 ± 0.19 | 0.70 ± 0.13 | 1.00 ± 0.20 | 0.017 | 0.846 | 0.789 | |
| 12 | 1.68 ± 0.27 | 2.25 ± 0.22 | 2.00 ± 0.18 | 1.88 ± 0.09 | 0.276 | 0.903 | 0.111 | |
| 24 | 3.07 ± 0.30 | 3.81 ± 0.34 | 3.29 ± 0.11 | 3.43 ± 0.28 | 0.127 | 0.754 | 0.288 | |
| 48 | 5.33 ± 0.92 | 10.2 ± 1.3 | 5.16 ± 0.33 | 9.55 ± 2.39 | 0.005 | 0.770 | 0.861 | |
| Total SCFA | 0 | 2.60 ± 0.22 | 2.71 ± 0.30 | 2.58 ± 0.36 | 2.70 ± 0.31 | 0.722 | 0.960 | 0.994 |
| 6 | 7.10 ± 1.70 | 12.6 ± 3.2 | 8.30 ± 1.70 | 13.0 ± 3.2 | 0.062 | 0.760 | 0.887 | |
| 12 | 14.0 ± 1.7 | 57.6 ± 22.8 | 17.8 ± 0.8 | 53.5 ± 24.1 | 0.030 | 0.992 | 0.812 | |
| 24 | 20.7 ± 2.5 | 213 ± 46 | 26.4 ± 1.4 | 176 ± 53 | <0.001 | 0.661 | 0.549 | |
| 48 | 51.6 ± 14.6 | 388 ± 31 | 50.6 ± 6.1 | 341 ± 52 | <0.001 | 0.447 | 0.465 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kilua, A.; Nomata, R.; Nagata, R.; Fukuma, N.; Shimada, K.; Han, K.-H.; Fukushima, M. Purple Sweet Potato Polyphenols Differentially Influence the Microbial Composition Depending on the Fermentability of Dietary Fiber in a Mixed Culture of Swine Fecal Bacteria. Nutrients 2019, 11, 1495. https://doi.org/10.3390/nu11071495
Kilua A, Nomata R, Nagata R, Fukuma N, Shimada K, Han K-H, Fukushima M. Purple Sweet Potato Polyphenols Differentially Influence the Microbial Composition Depending on the Fermentability of Dietary Fiber in a Mixed Culture of Swine Fecal Bacteria. Nutrients. 2019; 11(7):1495. https://doi.org/10.3390/nu11071495
Chicago/Turabian StyleKilua, Aldrine, Riri Nomata, Ryuji Nagata, Naoki Fukuma, Kenichiro Shimada, Kyu-Ho Han, and Michihiro Fukushima. 2019. "Purple Sweet Potato Polyphenols Differentially Influence the Microbial Composition Depending on the Fermentability of Dietary Fiber in a Mixed Culture of Swine Fecal Bacteria" Nutrients 11, no. 7: 1495. https://doi.org/10.3390/nu11071495
APA StyleKilua, A., Nomata, R., Nagata, R., Fukuma, N., Shimada, K., Han, K.-H., & Fukushima, M. (2019). Purple Sweet Potato Polyphenols Differentially Influence the Microbial Composition Depending on the Fermentability of Dietary Fiber in a Mixed Culture of Swine Fecal Bacteria. Nutrients, 11(7), 1495. https://doi.org/10.3390/nu11071495