Impact of Arginine Nutrition and Metabolism during Pregnancy on Offspring Outcomes



Abstract
1. Introduction
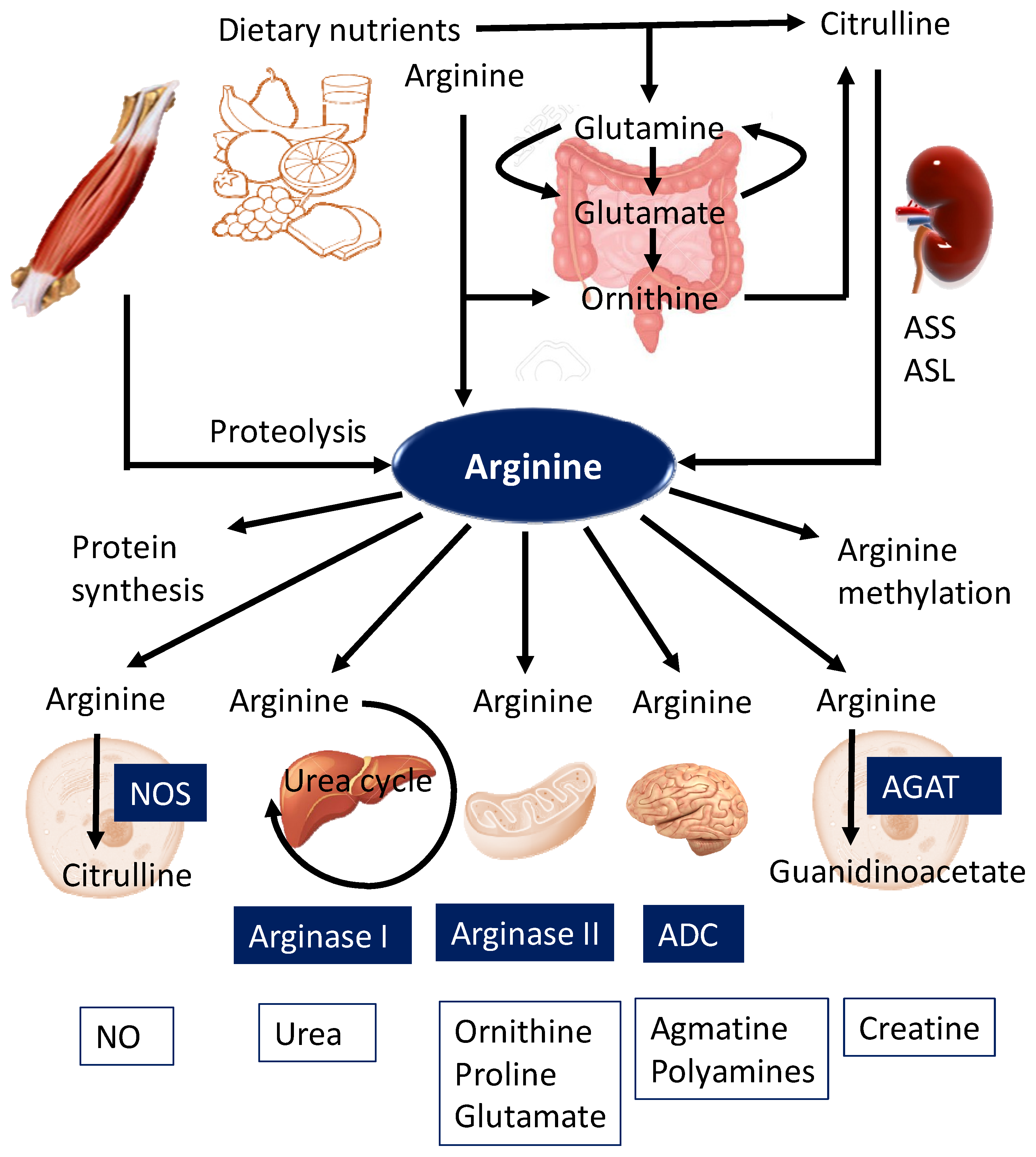
2. Biosynthesis and Metabolism of Arginine
2.1. De Novo Synthesis
2.2. Metabolism
2.3. Transport
2.4. Arginine Methylation
3. Arginine Nutrition in Pregnancy and Fetal Development
4. Impaired Arginine Metabolic Pathway in Compromised Pregnancy and Fetal Programming
4.1. Preeclampsia
4.2. Gestational Diabetes Mellitus
4.3. Intrauterine Growth Retardation
4.4. Prematurity and Low Birth Weight
4.5. Developmental Programming of Adult-Onset Diseases
5. Targeting the Arginine Metabolic Pathway to Prevent Adult-Onset Diseases
5.1. Arginine
5.2. Citrulline
5.3. Glutamate and Glutamine
5.4. Others
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Wu, G.; Bazer, F.W.; Davis, T.A.; Kim, S.W.; Li, P.; Marc Rhoads, J.; Carey Satterfield, M.; Smith, S.B.; Spencer, T.E.; Yin, Y. Arginine metabolism and nutrition in growth, health and disease. Amino Acids 2009, 37, 153–168. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Morris, S.M., Jr. Arginine metabolism: Nitric oxide and beyond. Biochem. J. 1998, 336, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Bazer, F.W.; Satterfield, M.C.; Li, X.; Wang, X.; Johnson, G.A.; Burghardt, R.C.; Dai, Z.; Wang, J.; Wu, Z. Impacts of arginine nutrition on embryonic and fetal development in mammals. Amino Acids 2013, 45, 241–256. [Google Scholar] [CrossRef] [PubMed]
- Hanson, M.; Gluckman, P. Developmental origins of noncommunicable disease: Population and public health implications. Am. J. Clin. Nutr. 2011, 94, 1754S–1758S. [Google Scholar] [CrossRef] [PubMed]
- Zarocostas, J. Need to increase focus on non-communicable diseases in global health, says WHO. Br. Med. J. 2010, 341, c7065. [Google Scholar] [CrossRef]
- Baird, J.; Jacob, C.; Barker, M.; Fall, C.H.; Hanson, M.; Harvey, N.C.; Inskip, H.M.; Kumaran, K.; Cooper, C. Developmental Origins of Health and Disease: A Lifecourse Approach to the Prevention of Non-Communicable Diseases. Healthcare 2017, 5, 14. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Joles, J.A. Reprogramming: A preventive strategy in hypertension focusing on the kidney. Int. J. Mol. Sci. 2016, 17, 23. [Google Scholar] [CrossRef]
- Paauw, N.D.; van Rijn, B.B.; Lely, A.T.; Joles, J.A. Pregnancy as a critical window for blood pressure regulation in mother and child: Programming and reprogramming. Acta Physiol. (Oxf). 2017, 219, 241–259. [Google Scholar] [CrossRef]
- McKnight, J.R.; Satterfield, M.C.; Jobgen, W.S.; Smith, S.B.; Spencer, T.E.; Meininger, C.J.; McNeal, C.J.; Wu, G. Beneficial effects of L-arginine on reducing obesity: Potential mechanisms and important implications for human health. Amino Acids 2010, 39, 349–357. [Google Scholar] [CrossRef]
- Rodrigues-Krause, J.; Krause, M.; Rocha, I.M.G.D.; Umpierre, D.; Fayh, A.P.T. Association of l-Arginine Supplementation with Markers of Endothelial Function in Patients with Cardiovascular or Metabolic Disorders: A Systematic Review and Meta-Analysis. Nutrients 2018, 11, 15. [Google Scholar] [CrossRef]
- Che, D.; Adams, S.; Zhao, B.; Qin, G.; Jiang, H. Effects of dietary L-arginine supplementation from conception to post-weaning in piglets. Curr. Protein Pept. Sci. 2019. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.N.; Tain, Y.L. The Double-Edged Sword Effects of Maternal Nutrition in the Developmental Programming of Hypertension. Nutrients 2018, 10, 1917. [Google Scholar] [CrossRef] [PubMed]
- Dhanakoti, S.N.; Brosnan, J.T.; Herzberg, G.R.; Brosnan, M.E. Renal arginine synthesis: Studies in vitro and in vivo. Am. J. Physiol. Endocrinol. Metab. 1990, 259, E437–E442. [Google Scholar] [CrossRef] [PubMed]
- Bratt, J.M.; Zeki, A.A.; Last, J.A.; Kenyon, N.J. Competitive metabolism of L-arginine: Arginase as a therapeutic target in asthma. J. Biomed. Res. 2011, 25, 299–308. [Google Scholar] [CrossRef]
- Chin-Dusting, J.P.; Willems, L.; Kaye, D.M. L-arginine transporters in cardiovascular disease: A novel therapeutic target. Pharmacol. Ther. 2007, 116, 428–436. [Google Scholar] [CrossRef] [PubMed]
- Castillo, L.; Beaumier, L.; Ajami, A.M.; Young, V.R. Whole body nitric oxide synthesis in healthy men determined from [15N] arginine-to-[15N] citrulline labeling. Proc. Natl. Acad. Sci. USA 1996, 93, 11460–11465. [Google Scholar] [CrossRef] [PubMed]
- Closs, E.I.; Simon, A.; Vékony, N.; Rotmann, A. Plasma membrane transporters for arginine. J. Nutr. 2004, 134, 2752S–2759S. [Google Scholar] [CrossRef] [PubMed]
- Bode-Böger, S.M.; Scalera, F.; Ignarro, L.J. The L-arginine paradox: Importance of the L-arginine/asymmetrical dimethylarginine ratio. Pharmacol. Ther. 2007, 114, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Blanc, R.S.; Richard, S. Arginine Methylation: The Coming of Age. Mol. Cell. 2017, 65, 8–24. [Google Scholar] [CrossRef]
- Morales, Y.; Cáceres, T.; May, K.; Hevel, J.M. Biochemistry and regulation of the protein arginine methyl transferases (PRMTs). Arch. Biochem. Biophys. 2016, 590, 138–152. [Google Scholar] [CrossRef]
- Tain, Y.L.; Hsu, C.N. Toxic Dimethylarginines: Asymmetric Dimethylarginine (ADMA) and Symmetric Dimethylarginine (SDMA). Toxins 2017, 9, 92. [Google Scholar] [CrossRef] [PubMed]
- Leiper, J.; Vallance, P. Biological significance of endogenous methylarginines that inhibit nitric oxide synthases. Cardiovasc. Res. 1999, 43, 542–548. [Google Scholar] [CrossRef]
- Bode-Boger, S.M.; Scalera, F.; Kielstein, J.T.; Martens-Lobenhoffer, J.; Breithardt, G.; Fobker, M.; Reinecke, H. Symmetrical dimethylarginine: A new combined parameter for renal function and extent of coronary artery disease. J. Am. Soc. Nephrol. 2006, 17, 1128–1134. [Google Scholar] [CrossRef] [PubMed]
- Bavoux, F.; Georges, P.; Bouy, M.; Leroy, B. Growth retardation and amino acids. Analysis of maternal plasma and amniotic fluid. J. Gynecol. Obstet. Biol. Reprod. 1977, 6, 931–940. [Google Scholar]
- Ban, H.; Shigemitsu, K.; Yamatsuji, T.; Haisa, M.; Nakajo, T.; Takaoka, M.; Nobuhisa, T.; Gunduz, M.; Tanaka, N.; Naomoto, Y. Arginine and Leucine regulate p70 S6 kinase and 4E-BP1 in intestinal epithelial cells. Int. J. Mol. Med. 2004, 13, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Herring, C.M.; Bazer, F.W.; Johnson, G.A.; Wu, G. Impacts of maternal dietary protein intake on fetal survival, growth, and development. Exp. Biol. Med. (Maywood) 2018, 243, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Rosselli, M.; Keller, P.J.; Dubey, R.K. Role of nitric oxide in the biology, physiology and pathophysiology of reproduction. Human Reprod. Update 1998, 4, 3–24. [Google Scholar] [CrossRef] [PubMed]
- Hussain, T.; Tan, B.; Ren, W.; Rahu, N.; Kalhoro, D.H.; Yin, Y. Exploring polyamines: Functions in embryo/fetal development. Anim. Nutr. 2017, 3, 7–10. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Lu, X.; Feng, Q. Deficiency in endothelial nitric oxide synthase impairs myocardial angiogenesis. Am. J. Physiol. Heart Circ. Physiol. 2002, 283, H2371–H2378. [Google Scholar] [CrossRef]
- Wang, X.; Frank, J.W.; Xu, J.; Dunlap, K.A.; Satterfield, M.C.; Burghardt, R.C.; Romero, J.J.; Hansen, T.R.; Wu, G.; Bazer, F.W. Functional role of arginine during the peri-implantation period of pregnancy. II. Consequences of loss of function of nitric oxide synthase NOS3 mRNA in ovine conceptus trophectoderm. Biol. Reprod. 2014, 91, 59. [Google Scholar] [CrossRef]
- Fickling, S.A.; Williams, D.; Vallance, P.; Nussey, S.S.; Whitley, G.S. Plasma of endogenous inhibitor of nitric oxide synthesis in normal pregnancy and pre-eclampsia. Lancet 1993, 342, 242–243. [Google Scholar] [CrossRef]
- Wu, G.; Bazer, F.W.; Johnson, G.A.; Herring, C.; Seo, H.; Dai, Z.; Wang, J.; Wu, Z.; Wang, X. Functional amino acids in the development of the pig placenta. Mol. Reprod. Dev. 2017, 84, 879–882. [Google Scholar] [CrossRef] [PubMed]
- Remacle, C.; Bieswal, F.; Bol, V.; Reusens, B. Developmental programming of adult obesity and cardiovascular disease in rodents by maternal nutrition imbalance. Am. J. Clin. Nutr. 2011, 94, 1846S–1852S. [Google Scholar] [CrossRef] [PubMed]
- Mathias, P.C.; Elmhiri, G.; de Oliveira, J.C.; Delayre-Orthez, C.; Barella, L.F.; Tófolo, L.P.; Fabricio, G.S.; Chango, A.; Abdennebi-Najar, L. Maternal diet, bioactive molecules, and exercising as reprogramming tools of metabolic programming. Eur. J. Nutr. 2014, 53, 711–722. [Google Scholar] [CrossRef] [PubMed]
- Roberts, J.M.; Mascalzoni, D.; Ness, R.B.; Poston, L. Global Pregnancy Collaboration. Collaboration to Understand Complex Diseases: Preeclampsia and Adverse Pregnancy Outcomes. Hypertension 2016, 67, 681–687. [Google Scholar] [CrossRef]
- Kim, Y.J.; Park, H.S.; Lee, H.Y.; Ha, E.H.; Suh, S.H.; Oh, S.K.; Yoo, H.S. Reduced L-arginine level and decreased placental eNOS activity in preeclampsia. Placenta 2006, 27, 438–444. [Google Scholar] [CrossRef] [PubMed]
- Noris, M.; Todeschini, M.; Cassis, P.; Pasta, F.; Cappellini, A.; Bonazzola, S.; Macconi, D.; Maucci, R.; Porrati, F.; Benigni, A.; et al. L-arginine depletion in preeclampsia orients nitric oxide synthase toward oxidant species. Hypertension 2004, 43, 614–622. [Google Scholar] [CrossRef] [PubMed]
- Pettersson, A.; Hedner, T.; Milsom, I. Increased circulating concentrations of asymmetric dimethyl arginine (ADMA), an endogenous inhibitor of nitric oxide synthesis, in preeclampsia. Acta Obstet. Gynecol. Scand. 1998, 77, 808–813. [Google Scholar] [CrossRef] [PubMed]
- Savvidou, M.D.; Hingorani, A.D.; Tsikas, D.; Frolich, J.C.; Vallance, P.; Nicolaides, K.H. Endothelial dysfunction and raised plasma concentrations of asymmetric dimethylarginine in pregnant women who subsequently develop pre-eclampsia. Lancet 2003, 361, 1511–1517. [Google Scholar] [CrossRef]
- Gui, S.; Jia, J.; Niu, X.; Bai, Y.; Zou, H.; Deng, J.; Zhou, R. Arginine supplementation for improving maternal and neonatal outcomes in hypertensive disorder of pregnancy: A systematic review. J. Renin Angiotensin Aldosterone Syst. 2014, 15, 88–96. [Google Scholar] [CrossRef]
- Helmbrecht, G.D.; Farhat, M.Y.; Lochbaum, L.; Brown, H.E.; Yadgarova, K.T.; Eglinton, G.S.; Ramwell, P.W. L-arginine reverses the adverse pregnancy changes induced by nitric oxide synthase inhibition in the rat. Am. J. Obstet. Gynecol. 1996, 175, 800–805. [Google Scholar] [CrossRef]
- Rytlewski, K.; Olszanecki, R.; Lauterbach, R.; Grzyb, A.; Basta, A. Effects of oral L-arginine on the foetal condition and neonatal outcome in preeclampsia: A preliminary report. Basic Clin. Pharmacol. Toxicol. 2006, 99, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Dolatkhah, N.; Hajifaraji, M.; Shakouri, S.K. Nutrition Therapy in Managing Pregnant Women with Gestational Diabetes Mellitus: A Literature Review. J. Family Reprod. Health 2018, 12, 57–72. [Google Scholar] [PubMed]
- Sobrevia, L.; Mann, G.E. Dysfunction of the endothelial nitric oxide signalling pathway in diabetes and hyperglycaemia. Exp. Physiol. 1997, 82, 423–452. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.S.; Sobrev, L. Gestational diabetes and the adenosine/L-arginine/nitric oxide (ALANO) pathway in human umbilical vein endothelium. Placenta 2006, 27, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, N.; Razi, F.; Nasli-Esfahani, E.; Qorbani, M.; Shirzad, N.; Larijani, B. Amino acid profiling in the gestational diabetes mellitus. J. Diabetes Metab. Disord. 2017, 16, 13. [Google Scholar] [CrossRef] [PubMed]
- Sertkaya, A.C.; Kafkasli, A.; Turkcuoglu, I.; Karabulut, A.B. Asymmetric dimethylarginine level in hyperglycemic gestation. Endocrine 2011, 40, 237–242. [Google Scholar] [CrossRef]
- Akturk, M.; Altinova, A.; Mert, I.; Dincel, A.; Sargin, A.; Buyukkagnici, U.; Arslan, M.; Danisman, N. Asymmetric dimethylarginine concentrations are elevated in women with gestational diabetes. Endocrine 2010, 38, 134–141. [Google Scholar] [CrossRef]
- Leiva, A.; Fuenzalida, B.; Barros, E.; Sobrevia, B.; Salsoso, R.; Sáez, T.; Villalobos, R.; Silva, L.; Chiarello, I.; Toledo, F.; et al. Nitric Oxide is a Central Common Metabolite in Vascular Dysfunction Associated with Diseases of Human Pregnancy. Curr. Vasc. Pharmacol. 2016, 14, 237–259. [Google Scholar] [CrossRef]
- Giri, H.; Chandel, S.; Dwarakanath, L.S.; Sreekumar, S.; Dixit, M. Increased endothelial inflammation, sTie-2 and arginase activity in umbilical cords obtained from gestational diabetic mothers. PLoS ONE 2013, 8, e84546. [Google Scholar] [CrossRef]
- Romo, A.; Carceller, R.; Tobajas, J. Intrauterine growth retardation (IUGR): Epidemiology and etiology. Pediatr. Endocrinol. Rev. 2009, 6, 332–336. [Google Scholar]
- Winterhager, E.; Gellhaus, A. Transplacental Nutrient Transport Mechanisms of Intrauterine Growth Restriction in Rodent Models and Humans. Front. Physiol. 2017, 8, 951. [Google Scholar] [CrossRef] [PubMed]
- Khalil, A.; Hardman, L.; O Brien, P. The role of arginine, homoarginine and nitric oxide in pregnancy. Amino Acids 2015, 47, 1715–1727. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Bazer, F.W.; Datta, S.; Johnson, G.A.; Li, P.; Satterfield, M.C.; Spencer, T.E. Proline metabolism in the conceptus: Implications for fetal growth and development. Amino Acids 2008, 35, 691–702. [Google Scholar] [CrossRef] [PubMed]
- Jansson, T.; Powell, T.L. Role of placental nutrient sensing in developmental programming. Clin. Obstet. Gynecol. 2013, 56, 591–601. [Google Scholar] [CrossRef] [PubMed]
- Malandro, M.S.; Beveridge, M.J.; Kilberg, M.S.; Novak, D.A. Effect of low-protein diet-induced intrauterine growth retardation on rat placental amino acid transport. Am. J. Physiol. 1996, 271, C295–C303. [Google Scholar] [CrossRef]
- Wu, G.; Bazer, F.W.; Wallace, J.M.; Spencer, T.E. BOARD-INVITED REVIEW: Intrauterine growth retardation: Implications for the animal sciences. J. Anim. Sci. 2006, 84, 2316–2337. [Google Scholar] [CrossRef]
- Vosatka, R.J.; Hassoun, P.M.; Harvey-Wilkes, K.B. Dietary L-arginine prevents fetal growth restriction in rats. Am. J. Obstet. Gynecol. 1998, 178, 242–246. [Google Scholar] [CrossRef]
- Xiao, X.M.; Li, L.P. L-Arginine treatment for asymmetric fetal growth restriction. Int. J. Gynaecol. Obstet. 2005, 88, 15–18. [Google Scholar] [CrossRef]
- Lawn, J.E.; Kinney, M. Preterm birth: Now the leading cause of child death worldwide. Sci. Transl. Med. 2014, 6, 263ed21. [Google Scholar] [CrossRef]
- Barker, D.J. Fetal origins of coronary heart disease. BMJ 1995, 311, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Ivorra, C.; Garcia-Vicent, C.; Chaves, F.J.; Monleón, D.; Morales, J.M.; Lurbe, E. Metabolomic profiling in blood from umbilical cords of low birth weight newborns. J. Transl. Med. 2012, 10, 142. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.; Liu, C.; Feng, C.; Fan, Z.; Dai, Z.; Lai, C.; Li, Z.; Wu, G.; Wang, J. Metabolomic analysis reveals differences in umbilical vein plasma metabolites between normal and growth-restricted fetal pigs during late gestation. J. Nutr. 2012, 142, 990–998. [Google Scholar] [CrossRef] [PubMed]
- Alexandre-Gouabau, M.C.; Courant, F.; Le Gall, G.; Moyon, T.; Darmaun, D.; Parnet, P.; Coupé, B.; Antignac, J.P. Offspring metabolomic response to maternal protein restriction in a rat model of intrauterine growth restriction (IUGR). J. Proteome Res. 2011, 10, 3292–3302. [Google Scholar] [CrossRef] [PubMed]
- Tiboni, G.M.; Giampietro, F. Inhibition of nitric oxide synthesis causes preterm delivery in the mouse. Hum. Reprod. 2000, 15, 1838–1842. [Google Scholar] [CrossRef]
- Facchinetti, F.; Neri, I.; Genazzani, A.R. L-arginine infusion reduces preterm uterine contractions. J. Perinat. Med. 1996, 24, 283–285. [Google Scholar] [CrossRef]
- Rytlewski, K.; Olszanecki, R.; Lauterbach, R.; Grzyb, A.; Kiec-Wilk, B.; Dembinska-Kiec, A.; Basta, A. Effects of oral L-arginine on the pulsatility indices of umbilical artery and middle cerebral artery in preterm labor. Eur. J. Obstet. Gynecol. Reprod. Biol. 2008, 138, 23–28. [Google Scholar] [CrossRef]
- Bai, S.Y.; Briggs, D.I.; Vickers, M.H. Increased systolic blood pressure in rat offspring following a maternal low-protein diet is normalized by maternal dietary choline supplementation. J. Dev. Orig. Health Dis. 2012, 3, 342–349. [Google Scholar] [CrossRef]
- McMillen, I.C.; Robinson, J.S. Developmental origins of the metabolic syndrome: Prediction, plasticity, and programming. Physiol. Rev. 2005, 85, 571–633. [Google Scholar] [CrossRef]
- Tain, Y.L.; Huang, L.T.; Lee, C.T.; Chan, J.Y.; Hsu, C.N. Maternal citrulline supplementation prevents prenatal NG-nitro-l-arginine-methyl ester (L-NAME)-induced programmed hypertension in rats. Biol. Reprod. 2015, 92, 7. [Google Scholar] [CrossRef]
- Tain, Y.L.; Hsu, C.N. Targeting on asymmetric dimethylarginine related nitric oxide-reactive oxygen species imbalance to reprogram the development of hypertension. Int. J. Mol. Sci. 2016, 17, 2020. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Sun, L.W.; Wang, Z.Y.; Deng, M.T.; Zhang, G.M.; Guo, R.H.; Ma, T.W.; Wang, F. Dietary-carbamylglutamate and rumen-protected -arginine supplementation ameliorate fetal growth restriction in undernourished ewes. J. Anim. Sci. 2016, 94, 2072–2085. [Google Scholar] [CrossRef] [PubMed]
- Lassala, A.; Bazer, F.W.; Cudd, T.A.; Datta, S.; Keisler, D.H.; Satterfield, M.C.; Spencer, T.E.; Wu, G. Parenteral administration of L-arginine prevents fetal growth restriction in undernourished ewes. J. Nutr. 2010, 140, 1242–1248. [Google Scholar] [CrossRef] [PubMed]
- Peine, J.L.; Jia, G.; Van Emon, M.L.; Neville, T.L.; Kirsch, J.D.; Hammer, C.J.; O’Rourke, S.T.; Reynolds, L.P.; Caton, J.S. Effects of maternal nutrition and rumen-protected arginine supplementation on ewe performance and postnatal lamb growth and internal organ mass. J. Anim. Sci. 2018, 96, 3471–3481. [Google Scholar] [CrossRef] [PubMed]
- Mateo, R.D.; Wu, G.; Bazer, F.W.; Park, J.C.; Shinzato, I.; Kim, S.W. Dietary L-arginine supplementation enhances the reproductive performance of gilts. J. Nutr. 2007, 137, 652–656. [Google Scholar] [CrossRef]
- Liu, X.D.; Wu, X.; Yin, Y.L.; Liu, Y.Q.; Geng, M.M.; Yang, H.S.; Blachier, F.; Wu, G.Y. Effects of dietary L-arginine or N-carbamylglutamate supplementation during late gestation of sows on the miR-15b/16, miR-221/222, VEGFA and eNOS expression in umbilical vein. Amino Acids 2012, 42, 2111–2119. [Google Scholar] [CrossRef] [PubMed]
- Tran, N.; Amarger, V.; Bourdon, A.; Misbert, E.; Grit, I.; Winer, N.; Darmaun, D. Maternal citrulline supplementation enhances placental function and fetal growth in a rat model of IUGR: Involvement of insulin-like growth factor 2 and angiogenic factors. J. Matern. Fetal Neonatal Med. 2017, 30, 1906–1911. [Google Scholar] [CrossRef]
- Bourdon, A.; Parnet, P.; Nowak, C.; Tran, N.; Winer, N.; Darmaun, D. L-Citrulline Supplementation Enhances Fetal Growth and Protein Synthesis in Rats with Intrauterine Growth Restriction. J. Nutr. 2016, 146, 532–541. [Google Scholar] [CrossRef]
- Tain, Y.L.; Hsieh, C.S.; Lin, I.C.; Chen, C.C.; Sheen, J.M.; Huang, L.T. Effects of maternal L-citrulline supplementation on renal function and blood pressure in offspring exposed to maternal caloric restriction: The impact of nitric oxide pathway. Nitric Oxide 2010, 23, 34–41. [Google Scholar] [CrossRef]
- Tain, Y.L.; Lee, W.C.; Hsu, C.N.; Lee, W.C.; Huang, L.T.; Lee, C.T.; Lin, C.Y. Asymmetric dimethylarginine is associated with developmental programming of adult kidney disease and hypertension in offspring of streptozotocin-treated mothers. PLoS ONE 2013, 8, e55420. [Google Scholar] [CrossRef]
- Tain, Y.L.; Sheen, J.M.; Chen, C.C.; Yu, H.R.; Tiao, M.M.; Kuo, H.C.; Huang, L.T. Maternal citrulline supplementation prevents prenatal dexamethasone-induced programmed hypertension. Free Radic. Res. 2014, 48, 580–586. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Lee, C.T.; Huang, L.T. Long-term effects of maternal citrulline supplementation on renal transcriptome prevention of nitric oxide depletion-related programmed hypertension: The impact of gene-nutrient interactions. Int. J. Mol. Sci. 2014, 15, 23255–23268. [Google Scholar] [CrossRef] [PubMed]
- Koeners, M.P.; van Faassen, E.E.; Wesseling, S.; Velden, M.S.; Koomans, H.A.; Braam, B.; Joles, J.A. Maternal Supplementation with Citrulline Increases Renal Nitric Oxide in Young Spontaneously Hypertensive Rats and Has Long-Term Antihypertensive Effects. Hypertension 2007, 50, 1077–1084. [Google Scholar] [CrossRef] [PubMed]
- Sawant, O.B.; Wu, G.; Washburn, S.E. Maternal L-glutamine supplementation prevents prenatal alcohol exposure-induced fetal growth restriction in an ovine model. Amino Acids 2015, 47, 1183–1192. [Google Scholar] [CrossRef] [PubMed]
- Luiking, Y.C.; Ten Have, G.A.M.; Wolfe, R.R.; Deutz, N.E.P. Arginine de novo and nitric oxide production in disease states. Am. J. Physiol. Endocrinol. Metab. 2012, 303, E1177–E1189. [Google Scholar] [CrossRef] [PubMed]
- Grimble, G.K. Adverse gastrointestinal effects of arginine and related amino acids. J. Nutr. 2007, 137, 1693S–1701S. [Google Scholar] [CrossRef] [PubMed]
- Alves, G.M.; Barão, M.A.; Odo, L.N.; Nascimento Gomes, G.; Franco Md Mdo, C.; Nigro, D.; Lucas, S.R.; Laurindo, F.R.; Brandizzi, L.I.; Zaladek Gil, F. L-Arginine effects on blood pressure and renal function of intrauterine restricted rats. Pediatr. Nephrol. 2002, 17, 856–862. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, D.S.; Diniz, M.M.; Haidar, A.A.; Cavanal, M.F.; da Silva Alves, E.; Carpinelli, A.R.; Gil, F.Z.; Hirata, A.E. L-Arginine supplementation improves insulin sensitivity and beta cell function in the offspring of diabetic rats through AKT and PDX-1 activation. Eur. J. Pharmacol. 2016, 791, 780–787. [Google Scholar] [CrossRef]
- Cynober, L.; Moinard, C.; De Bandt, J.P. The 2009 ESPEN Sir David Cuthbertson. Citrulline: A new major signaling molecule or just another player in the pharmaconutrition game? Clin. Nutr. 2010, 29, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Lassala, A.; Bazer, F.W.; Cudd, T.A.; Li, P.; Li, X.; Satterfield, M.C. Intravenous Administration of L-Citrulline to Pregnant Ewes Is More Effective Than L-Arginine for Increasing Arginine Availability in the Fetus. J. Nutr. 2009, 139, 660–665. [Google Scholar] [CrossRef] [PubMed]
- Schwedhelm, E.; Maas, R.; Freese, R.; Jung, D.; Lukacs, Z.; Jambrecina, A.; Spickler, W.; Schulze, F.; Boger, R.H. Pharmacokinetic and pharmacodynamic properties of oral L-citrulline and L-arginine: Impact on nitric oxide metabolism. Br. J. Clin. Pharmacol. 2008, 65, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Allerton, T.D.; Proctor, D.N.; Stephens, J.M.; Dugas, T.R.; Spielmann, G.; Irving, B.A. L-Citrulline Supplementation: Impact on Cardiometabolic Health. Nutrients 2018, 10, 921. [Google Scholar] [CrossRef] [PubMed]
- Tome, D. The roles of dietary glutamate in the intestine. Ann. Nutr. Metab. 2018, 73, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.Y.; Knabe, D.A.; Kim, S.W. Arginine nutrition in neonatal pigs. J. Nutr. 2004, 134, 2783–2790. [Google Scholar] [CrossRef] [PubMed]
- Chacher, B.; Wang, D.M.; Liu, H.Y.; Liu, J.X. Degradaton of L-arginine and N-carbamoyl glutamate and their effect on rumen fermentaton in vitro. Ital. J. Anim. Sci. 2012, 11, 4693–4696. [Google Scholar] [CrossRef]
- Caldwell, R.W.; Rodriguez, P.C.; Toque, H.A.; Narayanan, S.P.; Caldwell, R.B. Arginase: A Multifaceted Enzyme Important in Health and Disease. Physiol. Rev. 2018, 98, 641–665. [Google Scholar] [CrossRef] [PubMed]
- Krause, B.J.; Carrasco-Wong, I.; Caniuguir, A.; Carvajal, J.; Farías, M.; Casanello, P. Endothelial eNOS/arginase imbalance contributes to vascular dysfunction in IUGR umbilical and placental vessels. Placenta 2013, 34, 20–28. [Google Scholar] [CrossRef]
- Grandvuillemin, I.; Buffat, C.; Boubred, F.; Lamy, E.; Fromonot, J.; Charpiot, P.; Simoncini, S.; Sabatier, F.; Dignat-George, F.; Peyter, A.C.; et al. Arginase upregulation and eNOS uncoupling contribute to impaired endothelium-dependent vasodilation in a rat model of intrauterine growth restriction. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 315, R509–R520. [Google Scholar] [CrossRef]
- Tain, Y.L.; Huang, L.T. Asymmetric dimethylarginine: Clinical applications in pediatric medicine. J. Formos. Med. Assoc. 2011, 110, 70–77. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Interventions | Animal Models | Species/ Gender | Age at Measure | Reprogramming Effects | Ref. |
|---|---|---|---|---|---|
| Arginine | |||||
| 0.2 or 2% in water from gestational day 1 to 21 | Hypoxia exposure | Wistar rat/M and F | Gestational day 21 | Improved IUGR | [58] |
| 21 mg/kg daily from gestational day 16 to delivery | Maternal L-NAME exposure | SD rat/M and F | Birth | Improved IUGR | [41] |
| 10 g/day in diet from gestational day 35 to 110 | Maternal 50% caloric restriction | Ovine/M and F | Gestational day 110 | Improved IUGR | [72] |
| 155 μmol/kg i.v. 3 times daily from gestational day 60 to delivery | 50% caloric restriction | Ovine/M and F | Birth | Improved IUGR | [73] |
| 180 mg/kg once daily from gestational day 54 to delivery | 40% caloric restriction | Ovine/M and F | Birth | Improved IUGR | [74] |
| 1% in diet from gestational day 30 to 114 | Spontaneous IUGR | Swine/M and F | Birth | Improved IUGR | [75] |
| 1% in diet from gestational day 90 to delivery | Spontaneous IUGR | Swine/M and F | Birth | Improved IUGR | [76] |
| Citrulline | |||||
| 2 g/kg/day in water in pregnancy | Low protein diet | SD rat/M and F | Gestational day 21 | Prevented IUGR | [77] |
| 2 g/kg/day in water in pregnancy | Low protein diet | SD rat/M and F | Birth | Prevented IUGR | [78] |
| 2.5 g/L of water in pregnancy and lactation | Maternal 50% caloric restriction | SD rat/M | 12 weeks | Prevented low nephron number and renal dysfunction | [79] |
| 2.5 g/L of water in pregnancy and lactation | Streptozotocin-induced diabetes | SD rat/M | 12 weeks | Prevented hypertension and kidney injury | [80] |
| 2.5 g/L of water in pregnancy and lactation | Prenatal dexamethasone exposure | SD rat/M | 12 weeks | Prevented hypertension | [81] |
| 2.5 g/L of water in pregnancy and lactation | Maternal L-NAME exposure | SD rat/M | 12 weeks | Prevented hypertension | [70,82] |
| 2.5 g/L of water from gestational day 7 to postnatal week 6 | Genetic hypertension model | SHR/M and F | 50 weeks | Prevented hypertension | [83] |
| Glutamate (N-carbamoylglutamate) | |||||
| 2.5 g/day in diet from gestational day 35 to 110 | Maternal 50% caloric restriction | Ovine/M and F | Gestational day 110 | Improved IUGR | [72] |
| 0.1% in diet from gestational day 90 to delivery | Spontaneous IUGR | Swine/M and F | Birth | Improved IUGR | [76] |
| Glutamine | |||||
| 100 mg/kg i.v. 3 times daily from gestational day 109 to 132 | Maternal alcohol exposure | Ovine/M and F | Gestational day 132 | Improved IUGR | [84] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsu, C.-N.; Tain, Y.-L. Impact of Arginine Nutrition and Metabolism during Pregnancy on Offspring Outcomes. Nutrients 2019, 11, 1452. https://doi.org/10.3390/nu11071452
Hsu C-N, Tain Y-L. Impact of Arginine Nutrition and Metabolism during Pregnancy on Offspring Outcomes. Nutrients. 2019; 11(7):1452. https://doi.org/10.3390/nu11071452
Chicago/Turabian StyleHsu, Chien-Ning, and You-Lin Tain. 2019. "Impact of Arginine Nutrition and Metabolism during Pregnancy on Offspring Outcomes" Nutrients 11, no. 7: 1452. https://doi.org/10.3390/nu11071452
APA StyleHsu, C.-N., & Tain, Y.-L. (2019). Impact of Arginine Nutrition and Metabolism during Pregnancy on Offspring Outcomes. Nutrients, 11(7), 1452. https://doi.org/10.3390/nu11071452