Network Analysis of the Potential Role of DNA Methylation in the Relationship between Plasma Carotenoids and Lipid Profile

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients and Design
2.2. Anthropometric and CM Measurements
2.3. DNA Extraction and Methylation Analysis
2.4. Carotenoid Measurements
2.5. Association between DNA Methylation Levels and Total Carotenoid Concentrations
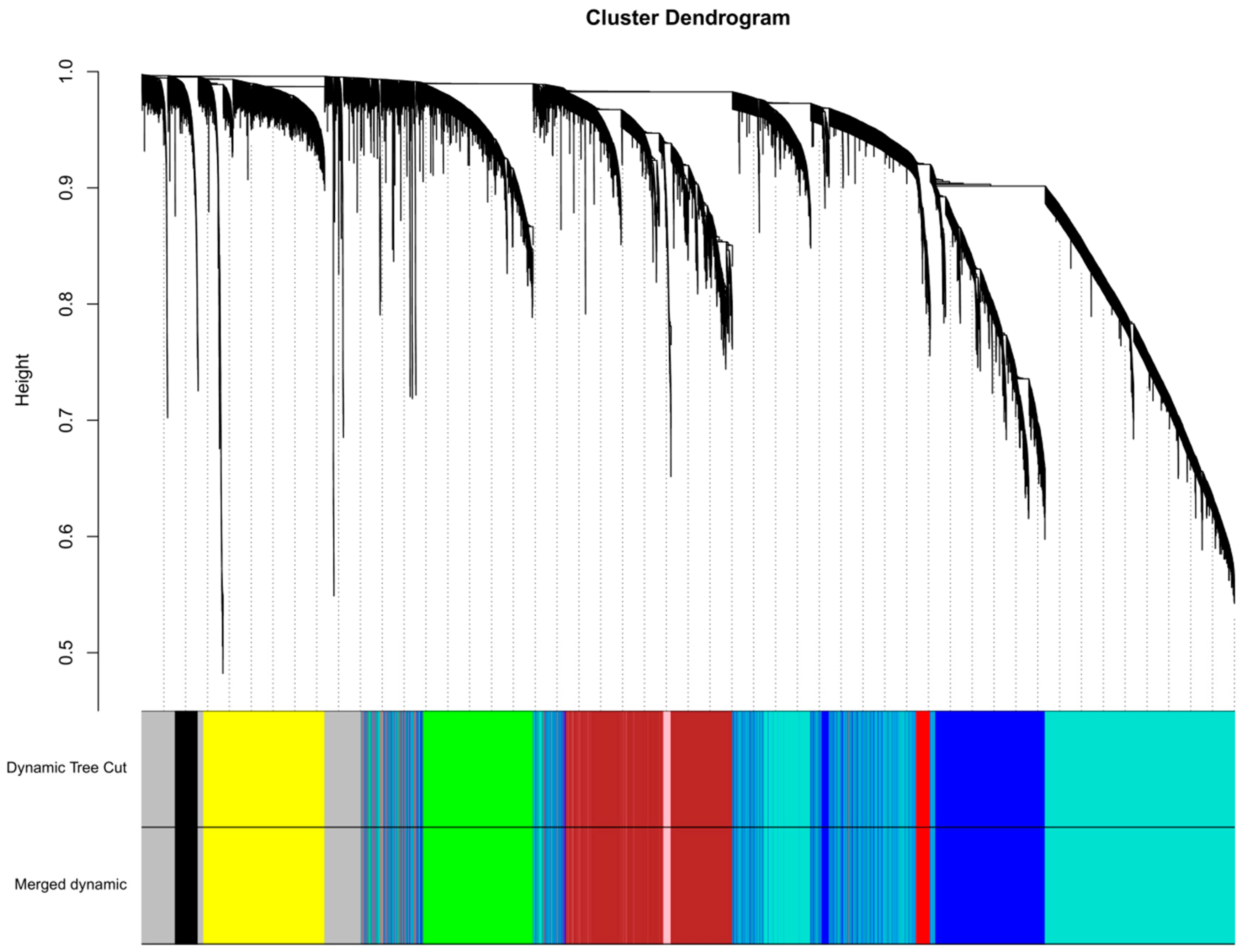
2.6. Weighted Gene Correlation Network Analysis (WGCNA)
3. Results
3.1. Characteristics and Biochemical Parameters
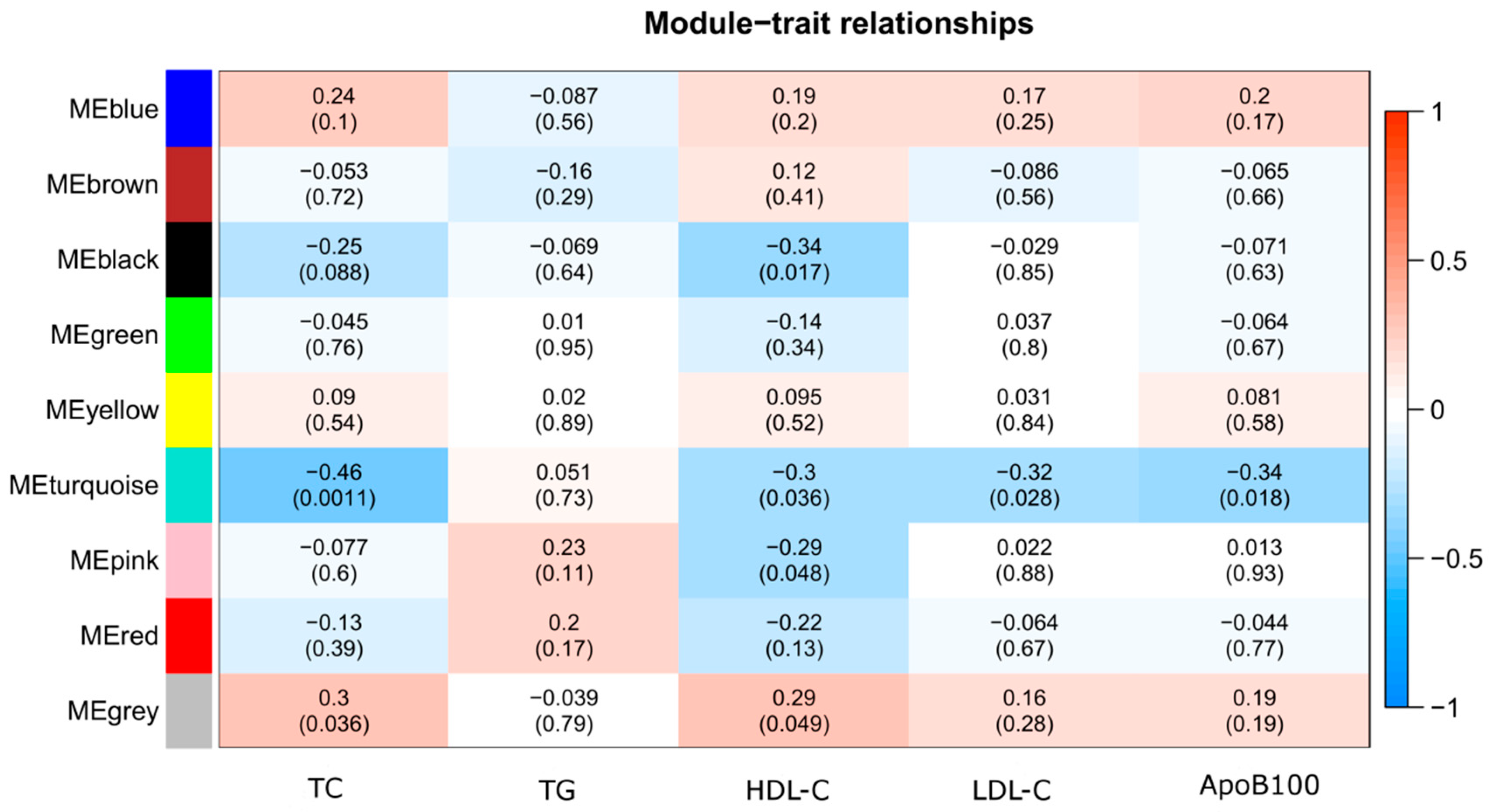
3.2. Weighted Gene Correlation Network Analysis (WGCNA)
3.3. Hub Gene Analysis
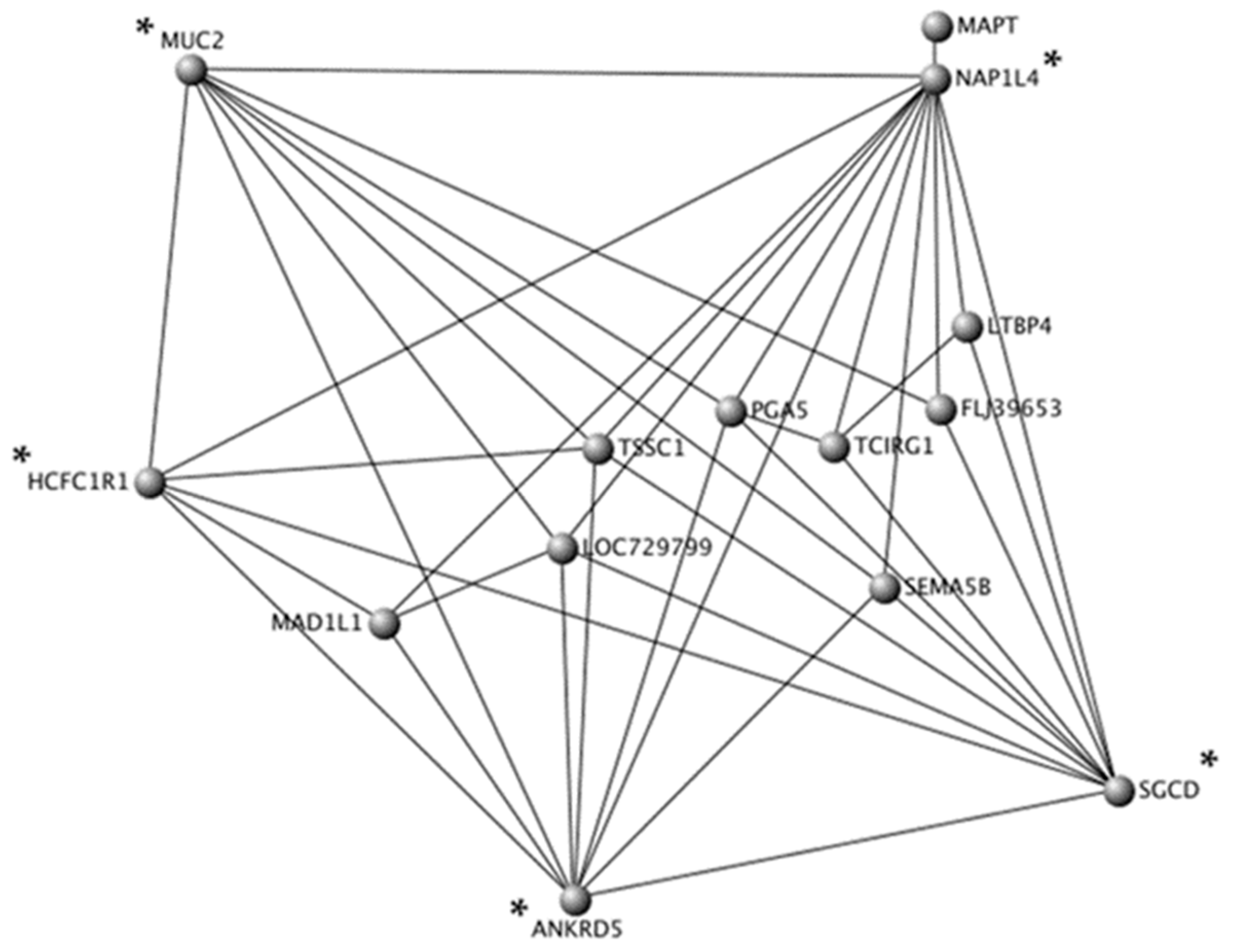
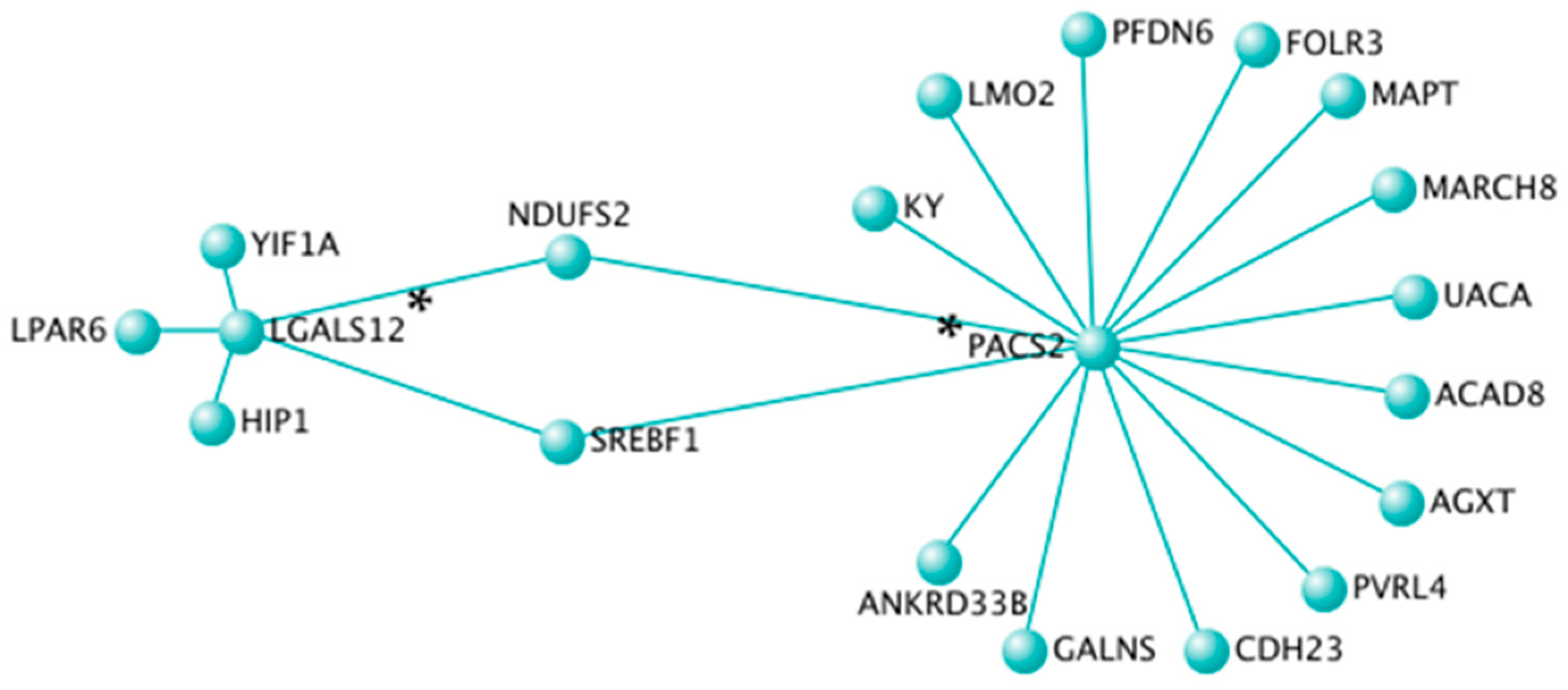
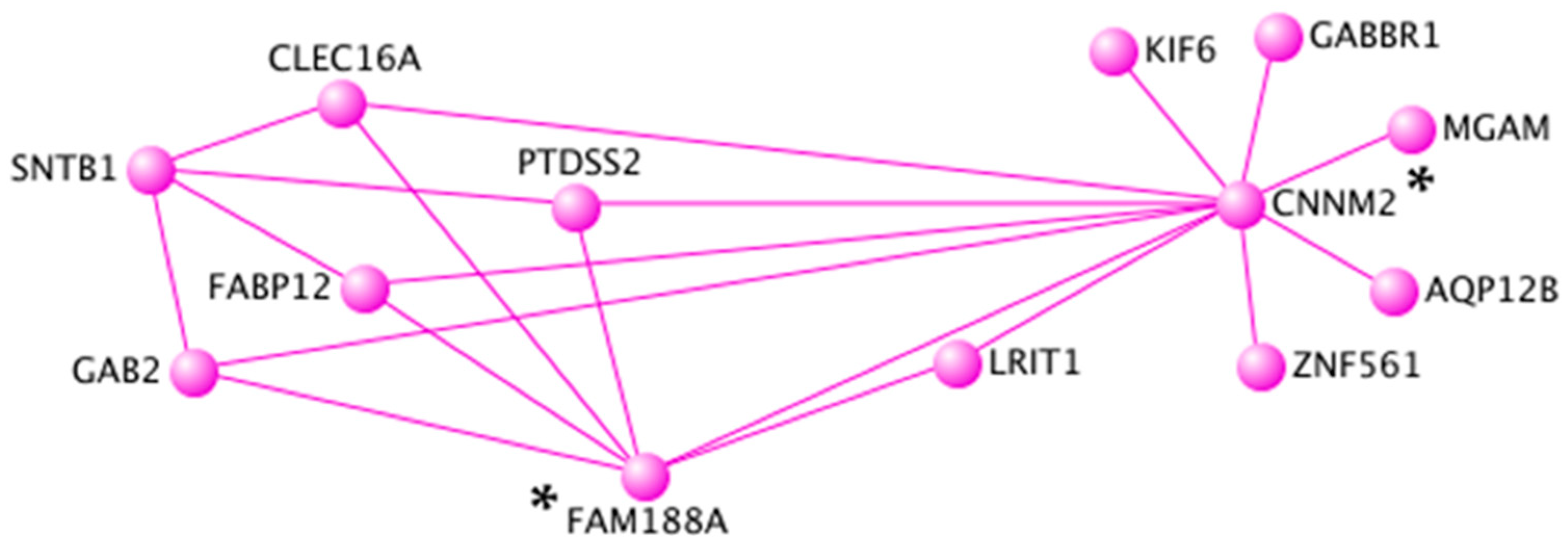
3.4. Topological Interaction Networks
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Guo, F.; Moellering, D.R.; Garvey, W.T. The progression of cardiometabolic disease: Validation of a new cardiometabolic disease staging system applicable to obesity. Obesity 2014, 22, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Gonzalez, M.A.; de la Fuente-Arrillaga, C.; Lopez-Del-Burgo, C.; Vazquez-Ruiz, Z.; Benito, S.; Ruiz-Canela, M. Low consumption of fruit and vegetables and risk of chronic disease: A review of the epidemiological evidence and temporal trends among spanish graduates. Public Health Nutr. 2011, 14, 2309–2315. [Google Scholar] [CrossRef] [PubMed]
- Couillard, C.; Lemieux, S.; Vohl, M.C.; Couture, P.; Lamarche, B. Carotenoids as biomarkers of fruit and vegetable intake in men and women. Br. J. Nutr. 2016, 116, 1206–1215. [Google Scholar] [CrossRef] [PubMed]
- Maiani, G.; Caston, M.J.; Catasta, G.; Toti, E.; Cambrodon, I.G.; Bysted, A.; Granado-Lorencio, F.; Olmedilla-Alonso, B.; Knuthsen, P.; Valoti, M.; et al. Carotenoids: Actual knowledge on food sources, intakes, stability and bioavailability and their protective role in humans. Mol. Nutr. Food Res. 2009, 53 (Suppl. 2), S194–S218. [Google Scholar] [CrossRef]
- Eroglu, A.; Harrison, E.H. Carotenoid metabolism in mammals, including man: Formation, occurrence, and function of apocarotenoids. J. Lipid Res. 2013, 54, 1719–1730. [Google Scholar] [CrossRef]
- Allore, T.; Lemieux, S.; Vohl, M.C.; Couture, P.; Lamarche, B.; Couillard, C. Correlates of the difference in plasma carotenoid concentrations between men and women. Br. J. Nutr. 2019, 121, 172–181. [Google Scholar] [CrossRef]
- Farook, V.S.; Reddivari, L.; Mummidi, S.; Puppala, S.; Arya, R.; Lopez-Alvarenga, J.C.; Fowler, S.P.; Chittoor, G.; Resendez, R.G.; Kumar, B.M.; et al. Genetics of serum carotenoid concentrations and their correlation with obesity-related traits in mexican american children. Am. J. Clin. Nutr. 2017, 106, 52–58. [Google Scholar] [CrossRef]
- Tremblay, B.L.; Guenard, F.; Lamarche, B.; Perusse, L.; Vohl, M.C. Genetic and common environmental contributions to familial resemblances in plasma carotenoid concentrations in healthy families. Nutrients 2018, 10, 1002. [Google Scholar] [CrossRef]
- Clevidence, B.A.; Bieri, J.G. Association of carotenoids with human plasma lipoproteins. Methods Enzymol. 1993, 214, 33–46. [Google Scholar]
- D’Adamo, C.R.; D’Urso, A.; Ryan, K.A.; Yerges-Armstrong, L.M.; Semba, R.D.; Steinle, N.I.; Mitchell, B.D.; Shuldiner, A.R.; McArdle, P.F. A common variant in the setd7 gene predicts serum lycopene concentrations. Nutrients 2016, 8, 82. [Google Scholar] [CrossRef]
- Beydoun, M.A.; Nalls, M.A.; Canas, J.A.; Evans, M.K.; Zonderman, A.B. Gene polymorphisms and gene scores linked to low serum carotenoid status and their associations with metabolic disturbance and depressive symptoms in african-american adults. Br. J. Nutr. 2014, 112, 992–1003. [Google Scholar] [CrossRef] [PubMed]
- Ferrucci, L.; Perry, J.R.; Matteini, A.; Perola, M.; Tanaka, T.; Silander, K.; Rice, N.; Melzer, D.; Murray, A.; Cluett, C.; et al. Common variation in the beta-carotene 15,15’-monooxygenase 1 gene affects circulating levels of carotenoids: A genome-wide association study. Am. J. Hum. Genet. 2009, 84, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Zubair, N.; Kooperberg, C.; Liu, J.; Di, C.; Peters, U.; Neuhouser, M.L. Genetic variation predicts serum lycopene concentrations in a multiethnic population of postmenopausal women. J. Nutr. 2015, 145, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Borel, P.; de Edelenyi, F.S.; Vincent-Baudry, S.; Malezet-Desmoulin, C.; Margotat, A.; Lyan, B.; Gorrand, J.M.; Meunier, N.; Drouault-Holowacz, S.; Bieuvelet, S. Genetic variants in bcmo1 and cd36 are associated with plasma lutein concentrations and macular pigment optical density in humans. Ann. Med. 2011, 43, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Bohn, T.; Desmarchelier, C.; Dragsted, L.O.; Nielsen, C.S.; Stahl, W.; Ruhl, R.; Keijer, J.; Borel, P. Host-related factors explaining interindividual variability of carotenoid bioavailability and tissue concentrations in humans. Mol. Nutr. Food Res. 2017, 61, 1600685. [Google Scholar] [CrossRef] [PubMed]
- Sharoni, Y.; Danilenko, M.; Dubi, N.; Ben-Dor, A.; Levy, J. Carotenoids and transcription. Arch. Biochem. Biophys. 2004, 430, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.G.; Erdman, J.W., Jr. Lycopene and apo-10’-lycopenal do not alter DNA methylation of gstp1 in lncap cells. Biochem. Biophys. Res. Commun. 2011, 412, 479–482. [Google Scholar] [CrossRef] [PubMed]
- King-Batoon, A.; Leszczynska, J.M.; Klein, C.B. Modulation of gene methylation by genistein or lycopene in breast cancer cells. Environ. Mol. Mutagenes. 2008, 49, 36–45. [Google Scholar] [CrossRef]
- Bollati, V.; Favero, C.; Albetti, B.; Tarantini, L.; Moroni, A.; Byun, H.M.; Motta, V.; Conti, D.M.; Tirelli, A.S.; Vigna, L.; et al. Nutrients intake is associated with DNA methylation of candidate inflammatory genes in a population of obese subjects. Nutrients 2014, 6, 4625–4639. [Google Scholar] [CrossRef]
- Quach, A.; Levine, M.E.; Tanaka, T.; Lu, A.T.; Chen, B.H.; Ferrucci, L.; Ritz, B.; Bandinelli, S.; Neuhouser, M.L.; Beasley, J.M.; et al. Epigenetic clock analysis of diet, exercise, education, and lifestyle factors. Aging 2017, 9, 419–446. [Google Scholar] [CrossRef]
- Mittelstrass, K.; Waldenberger, M. DNA methylation in human lipid metabolism and related diseases. Curr. Opin. Lipidol. 2018, 29, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Langfelder, P.; Horvath, S. Wgcna: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, B.L.; Guenard, F.; Lamarche, B.; Perusse, L.; Vohl, M.C. Familial resemblances in blood leukocyte DNA methylation levels. Epigenetics 2016, 11, 831–838. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, B.L.; Guenard, F.; Lamarche, B.; Perusse, L.; Vohl, M.C. Familial resemblances in human whole blood transcriptome. BMC Genom. 2018, 19, 300. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, B.L.; Guenard, F.; Lamarche, B.; Perusse, L.; Vohl, M.C. Familial resemblances in human plasma metabolites are attributable to both genetic and common environmental effects. Nutr. Res. 2019, 61, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Callaway, C.; Chumlea, W.; Bouchard, C.; Himes, J.; Lohman, T.; Martin, A.; Mitchell, C.; Mueller, W.; Roche, A.; Seefeldt, V. Standardization of Anthropomeric Measurements: The Airlie (va) Consensus Conference; Human Kinetics: Champaign, IL, USA, 1988; pp. 39–80. [Google Scholar]
- McNamara, J.R.; Schaefer, E.J. Automated enzymatic standardized lipid analyses for plasma and lipoprotein fractions. Clin. Chim. Acta 1987, 166, 1–8. [Google Scholar] [CrossRef]
- Burstein, M.; Samaille, J. On a rapid determination of the cholesterol bound to the serum alpha- and beta-lipoproteins. Clin. Chim. Acta 1960, 5, 609. [Google Scholar] [CrossRef]
- Albers, J.J.; Warnick, G.R.; Wiebe, D.; King, P.; Steiner, P.; Smith, L.; Breckenridge, C.; Chow, A.; Kuba, K.; Weidman, S.; et al. Multi-laboratory comparison of three heparin-mn2+ precipitation procedures for estimating cholesterol in high-density lipoprotein. Clin. Chem. 1978, 24, 853–856. [Google Scholar]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar] [PubMed]
- Laurell, C.B. Quantitative estimation of proteins by electrophoresis in agarose gel containing antibodies. Anal. Biochem. 1966, 15, 45–52. [Google Scholar] [CrossRef]
- Pirro, M.; Bergeron, J.; Dagenais, G.R.; Bernard, P.M.; Cantin, B.; Despres, J.P.; Lamarche, B. Age and duration of follow-up as modulators of the risk for ischemic heart disease associated with high plasma c-reactive protein levels in men. Arch. Intern. Med. 2001, 161, 2474–2480. [Google Scholar] [CrossRef] [PubMed]
- Richterich, R.; Dauwalder, H. Determination of plasma glucose by hexokinase-glucose-6-phosphate dehydrogenase method. Schweizerische Medizinische Wochenschrift 1971, 101, 615–618. [Google Scholar] [PubMed]
- Desbuquois, B.; Aurbach, G.D. Use of polyethylene glycol to separate free and antibody-bound peptide hormones in radioimmunoassays. J. Clin. Endocrinol. Metab. 1971, 33, 732–738. [Google Scholar] [CrossRef] [PubMed]
- Price, M.E.; Cotton, A.M.; Lam, L.L.; Farre, P.; Emberly, E.; Brown, C.J.; Robinson, W.P.; Kobor, M.S. Additional annotation enhances potential for biologically-relevant analysis of the illumina infinium humanmethylation450 beadchip array. Epigenet. Chromatin 2013, 6, 4. [Google Scholar] [CrossRef] [PubMed]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Tremblay, B.L.; Guenard, F.; Lamarche, B.; Perusse, L.; Vohl, M.C. Weighted gene co-expression network analysis to explain the relationship between plasma total carotenoids and lipid profile. Genes Nutr. 2019, 14, 16. [Google Scholar] [CrossRef] [PubMed]
- Langfelder, P.; Horvath, S. Fast R functions for robust correlations and hierarchical clustering. J. Stat. Softw. 2012, 46, 1–17. [Google Scholar] [CrossRef]
- Zhang, B.; Horvath, S. A general framework for weighted gene co-expression network analysis. Stat. Appl. Genet. Mol. Biol. 2005, 4, 17. [Google Scholar] [CrossRef] [PubMed]
- Langfelder, P.; Horvath, S. Eigengene networks for studying the relationships between co-expression modules. BMC Syst. Biol. 2007, 1, 54. [Google Scholar] [CrossRef] [PubMed]
- Horvath, S.; Zhang, B.; Carlson, M.; Lu, K.V.; Zhu, S.; Felciano, R.M.; Laurance, M.F.; Zhao, W.; Qi, S.; Chen, Z.; et al. Analysis of oncogenic signaling networks in glioblastoma identifies aspm as a molecular target. Proc. Natl. Acad. Sci. USA 2006, 103, 17402–17407. [Google Scholar] [CrossRef] [PubMed]
- Visant 5.0. Visual Analyses of Metabolic Networks in Cells and Ecosystems. Available online: http://visant.bu.edu (accessed on 18 December 2018).
- Hu, Z.; Snitkin, E.S.; DeLisi, C. Visant: An integrative framework for networks in systems biology. Brief. Bioinform. 2008, 9, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [PubMed]
- Borel, P.; Desmarchelier, C.; Nowicki, M.; Bott, R. A combination of single-nucleotide polymorphisms is associated with interindividual variability in dietary beta-carotene bioavailability in healthy men. J. Nutr. 2015, 145, 1740–1747. [Google Scholar] [CrossRef] [PubMed]
- Borel, P.; Desmarchelier, C.; Nowicki, M.; Bott, R. Lycopene bioavailability is associated with a combination of genetic variants. Free Radic. Biol. Med. 2015, 83, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Borel, P.; Desmarchelier, C.; Nowicki, M.; Bott, R.; Morange, S.; Lesavre, N. Interindividual variability of lutein bioavailability in healthy men: Characterization, genetic variants involved, and relation with fasting plasma lutein concentration. Am. J. Clin. Nutr. 2014, 100, 168–175. [Google Scholar] [CrossRef]
- Fu, L.J.; Ding, Y.B.; Wu, L.X.; Wen, C.J.; Qu, Q.; Zhang, X.; Zhou, H.H. The effects of lycopene on the methylation of the gstp1 promoter and global methylation in prostatic cancer cell lines pc3 and lncap. Int. J. Endocrinol. 2014, 2014, 620165. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Fuentes, F.; Shu, L.; Wang, C.; Pung, D.; Li, W.; Zhang, C.; Guo, Y.; Kong, A.N. Epigenetic cpg methylation of the promoter and reactivation of the expression of gstp1 by astaxanthin in human prostate lncap cells. AAPS J. 2017, 19, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Willer, C.J.; Schmidt, E.M.; Sengupta, S.; Peloso, G.M.; Gustafsson, S.; Kanoni, S.; Ganna, A.; Chen, J.; Buchkovich, M.L.; Mora, S.; et al. Discovery and refinement of loci associated with lipid levels. Nat. Genet. 2013, 45, 1274–1283. [Google Scholar] [PubMed]
- Ligthart, S.; Vaez, A.; Hsu, Y.H.; Inflammation Working Group of the CHARGE Consortium; PMI-WG-XCP; LifeLines Cohort Study; Stolk, R.; Uitterlinden, A.G.; Hofman, A.; Alizadeh, B.Z.; et al. Bivariate genome-wide association study identifies novel pleiotropic loci for lipids and inflammation. BMC Genom. 2016, 17, 443. [Google Scholar]
- Huan, T.; Rong, J.; Liu, C.; Zhang, X.; Tanriverdi, K.; Joehanes, R.; Chen, B.H.; Murabito, J.M.; Yao, C.; Courchesne, P.; et al. Genome-wide identification of microrna expression quantitative trait loci. Nat. Commun. 2015, 6, 6601. [Google Scholar] [CrossRef] [PubMed]
- Willer, C.J.; Sanna, S.; Jackson, A.U.; Scuteri, A.; Bonnycastle, L.L.; Clarke, R.; Heath, S.C.; Timpson, N.J.; Najjar, S.S.; Stringham, H.M.; et al. Newly identified loci that influence lipid concentrations and risk of coronary artery disease. Nat. Genet. 2008, 40, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Comuzzie, A.G.; Cole, S.A.; Laston, S.L.; Voruganti, V.S.; Haack, K.; Gibbs, R.A.; Butte, N.F. Novel genetic loci identified for the pathophysiology of childhood obesity in the hispanic population. PLoS ONE 2012, 7, e51954. [Google Scholar] [CrossRef] [PubMed]
- Altunrende, M.E.; Gezen-Ak, D.; Atasoy, I.L.; Candas, E.; Dursun, E. The role of astaxanthin on transcriptional regulation of nmda receptors voltage sensitive calcium channels and calcium binding proteins in primary cortical neurons. Noro Psikiyatr. Ars. 2018, 55, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Notarangelo, L.D.; Kim, M.S.; Walter, J.E.; Lee, Y.N. Human rag mutations: Biochemistry and clinical implications. Nat. Rev. Immunol. 2016, 16, 234–246. [Google Scholar] [CrossRef] [PubMed]
- Reardon, C.A.; Blachowicz, L.; White, T.; Cabana, V.; Wang, Y.; Lukens, J.; Bluestone, J.; Getz, G.S. Effect of immune deficiency on lipoproteins and atherosclerosis in male apolipoprotein e-deficient mice. Arterioscler. Thromb. Vasc. Biol. 2001, 21, 1011–1016. [Google Scholar] [CrossRef] [PubMed]
- Hall, A.M.; Krishnamoorthy, L.; Orlow, S.J. 25-hydroxycholesterol acts in the golgi compartment to induce degradation of tyrosinase. Pigment Cell Res. 2004, 17, 396–406. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, T.J.; Theusch, E.; Haldar, T.; Ranatunga, D.K.; Jorgenson, E.; Medina, M.W.; Kvale, M.N.; Kwok, P.Y.; Schaefer, C.; Krauss, R.M.; et al. A large electronic-health-record-based genome-wide study of serum lipids. Nat. Genet. 2018, 50, 401–413. [Google Scholar] [CrossRef] [PubMed]
- Kaess, B.M.; Tomaszewski, M.; Braund, P.S.; Stark, K.; Rafelt, S.; Fischer, M.; Hardwick, R.; Nelson, C.P.; Debiec, R.; Huber, F.; et al. Large-scale candidate gene analysis of hdl particle features. PLoS ONE 2011, 6, e14529. [Google Scholar] [CrossRef]
- Simmen, T.; Aslan, J.E.; Blagoveshchenskaya, A.D.; Thomas, L.; Wan, L.; Xiang, Y.; Feliciangeli, S.F.; Hung, C.H.; Crump, C.M.; Thomas, G. Pacs-2 controls endoplasmic reticulum-mitochondria communication and bid-mediated apoptosis. EMBO J. 2005, 24, 717–729. [Google Scholar] [CrossRef]
- Yang, R.Y.; Hsu, D.K.; Yu, L.; Chen, H.Y.; Liu, F.T. Galectin-12 is required for adipogenic signaling and adipocyte differentiation. J. Biol. Chem. 2004, 279, 29761–29766. [Google Scholar] [CrossRef]
- The Coronary Artery Disease (C4D) Genetics Consortium. A genome-wide association study in europeans and south asians identifies five new loci for coronary artery disease. Nat. Genet. 2011, 43, 339–344. [Google Scholar] [CrossRef]
- Dichgans, M.; Malik, R.; Konig, I.R.; Rosand, J.; Clarke, R.; Gretarsdottir, S.; Thorleifsson, G.; Mitchell, B.D.; Assimes, T.L.; Levi, C.; et al. Shared genetic susceptibility to ischemic stroke and coronary artery disease: A genome-wide analysis of common variants. Stroke 2014, 45, 24–36. [Google Scholar] [CrossRef] [PubMed]
- Wain, L.V.; Verwoert, G.C.; O’Reilly, P.F.; Shi, G.; Johnson, T.; Johnson, A.D.; Bochud, M.; Rice, K.M.; Henneman, P.; Smith, A.V.; et al. Genome-wide association study identifies six new loci influencing pulse pressure and mean arterial pressure. Nat. Genet. 2011, 43, 1005–1011. [Google Scholar] [CrossRef] [PubMed]
- Guénard, F.; Deshaies, Y.; Hould, F.S.; Lebel, S.; Tchernof, A.; Marceau, P.; Vohl, M.C. Use of blood as a surrogate model for the assessment of visceral adipose tissue methylation profiles associated with the metabolic syndrome in men. J. Mol. Genet. Med. 2016, 10. [Google Scholar] [CrossRef]
- Gruber, M.; Chappell, R.; Millen, A.; LaRowe, T.; Moeller, S.M.; Iannaccone, A.; Kritchevsky, S.B.; Mares, J. Correlates of serum lutein + zeaxanthin: Findings from the third national health and nutrition examination survey. J. Nutr. 2004, 134, 2387–2394. [Google Scholar] [CrossRef] [PubMed]
- Al-Delaimy, W.K.; van Kappel, A.L.; Ferrari, P.; Slimani, N.; Steghens, J.P.; Bingham, S.; Johansson, I.; Wallstrom, P.; Overvad, K.; Tjonneland, A.; et al. Plasma levels of six carotenoids in nine European countries: Report from the European prospective investigation into cancer and nutrition (epic). Public Health Nutr. 2004, 7, 713–722. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biochemical Parameters | Parents (n = 22) | Children (n = 26) |
|---|---|---|
| Age (years) | 42.3 ± 5.3 | 11.3 ± 3.4 |
| BMI (kg/m2) | 23.9 ± 3.0 | - |
| BMI percentile | - | 50 ± 31.1 |
| TC (mmol/L) | 4.68 ± 0.55 | 4.28 ± 0.51 |
| HDL-C (mmol/L) | 1.63 ± 0.38 | 1.55 ± 0.29 |
| LDL-C (mmol/L) | 2.61 ± 0.55 | 2.31 ± 0.45 |
| TC/HDL-C | 3.02 ± 0.78 | 2.83 ± 0.47 |
| ApoB100 (g/L) | 0.80 ± 0.15 | 0.70 ± 0.13 |
| TG (mmol/L) | 0.95 ± 0.35 | 0.93 ± 0.39 |
| Glucose (mmol/L) | 5.20 ± 0.34 | 4.86 ± 0.23 |
| Insulin (pmol/L) | 66.59 ± 33.54 | 73.00 ± 28.96 |
| CRP (mg/L) | 0.77 ± 0.84 | 0.43 ± 0.66 |
| Total carotenoids (μmol/L) | 6.35 ± 2.39 | 5.70 ± 2.05 |
| Module-Lipid | CpG Site | Gene (Chr, Accession Number) | GS | p-Value GS | MM | p-Value MM |
|---|---|---|---|---|---|---|
| Black HDL-C | cg01999908 | PAX4 (Chr7, NM_006193) | −0.37 | 0.0092 | 0.85 | 2.10 × 10−14 |
| cg08731068 | TBC1D16 (Chr17, NM_019020) | −0.37 | 0.011 | 0.89 | 1.70 × 10−17 | |
| cg04010296 | PGA5 (Chr11, NM_014224) | −0.36 | 0.011 | 0.83 | 4.08 × 10−13 | |
| Turquoise TC | cg00000029 | RBL2 (Chr16, NM_005611) | −0.59 | 8.49 × 10−6 | 0.83 | 3.45 × 10−13 |
| cg02500883 | GRIN3A (Chr9, NM_133445) | −0.57 | 1.98 × 10−5 | 0.81 | 2.52 × 10−12 | |
| cg00864012 | TEX2 (Chr17, NM_018469) | −0.56 | 3.39 × 10−5 | 0.85 | 2.39 × 10−14 | |
| Turquoise HDL-C | cg27616007 | LST1 (Chr6, NR_029461) | −0.42 | 0.0027 | 0.81 | 3.82 × 10−12 |
| cg24240870 | GOSR1 (Chr17, NM_004871) | −0.42 | 0.0030 | 0.81 | 4.55 × 10−12 | |
| cg18089000 | GBGT1 (Chr9, NM_021996) | −0.42 | 0.0031 | 0.86 | 3.88 × 10−15 | |
| Turquoise LDL-C | cg07665923 | C7orf50 (Chr7, NM_001134396) | −0.45 | 0.0012 | 0.83 | 2.22 × 10−13 |
| cg02119755 | PDPK1 (Chr16, NM_031268) | −0.44 | 0.0017 | 0.87 | 1.62 × 10−15 | |
| cg06142108 | SYT17 (Chr16, NM_016524) | −0.43 | 0.0022 | 0.88 | 1.75 × 10−16 | |
| Turquoise ApoB100 | cg17053201 | GABBR1 (Chr6, NM_021904) | −0.46 | 0.00096 | 0.82 | 8.40 × 10−13 |
| cg21671607 | RAG2 (Chr11, NM_000536) | −0.46 | 0.0011 | 0.84 | 7.35 × 10−14 | |
| cg07665923 | C7orf50 (Chr7, NM_001134396) | −0.46 | 0.0011 | 0.83 | 2.22 × 10−13 | |
| Pink HDL-C | cg15505294 | RNASE11 (Chr14, NM_145250) | 0.36 | 0.012 | −0.95 | 1.87 × 10−24 |
| cg01719157 | TRIM68 (Chr11, NM_018073) | −0.35 | 0.013 | 0.81 | 4.10 × 10−12 | |
| cg09113768 | DEPDC1 (Chr1, NM_017779) | −0.35 | 0.015 | 0.83 | 2.22 × 10−13 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tremblay, B.L.; Guénard, F.; Lamarche, B.; Pérusse, L.; Vohl, M.-C. Network Analysis of the Potential Role of DNA Methylation in the Relationship between Plasma Carotenoids and Lipid Profile. Nutrients 2019, 11, 1265. https://doi.org/10.3390/nu11061265
Tremblay BL, Guénard F, Lamarche B, Pérusse L, Vohl M-C. Network Analysis of the Potential Role of DNA Methylation in the Relationship between Plasma Carotenoids and Lipid Profile. Nutrients. 2019; 11(6):1265. https://doi.org/10.3390/nu11061265
Chicago/Turabian StyleTremblay, Bénédicte L., Frédéric Guénard, Benoît Lamarche, Louis Pérusse, and Marie-Claude Vohl. 2019. "Network Analysis of the Potential Role of DNA Methylation in the Relationship between Plasma Carotenoids and Lipid Profile" Nutrients 11, no. 6: 1265. https://doi.org/10.3390/nu11061265
APA StyleTremblay, B. L., Guénard, F., Lamarche, B., Pérusse, L., & Vohl, M.-C. (2019). Network Analysis of the Potential Role of DNA Methylation in the Relationship between Plasma Carotenoids and Lipid Profile. Nutrients, 11(6), 1265. https://doi.org/10.3390/nu11061265