Protective Effects of Omega-3 Fatty Acids in Cancer-Related Complications

Abstract
1. Introduction
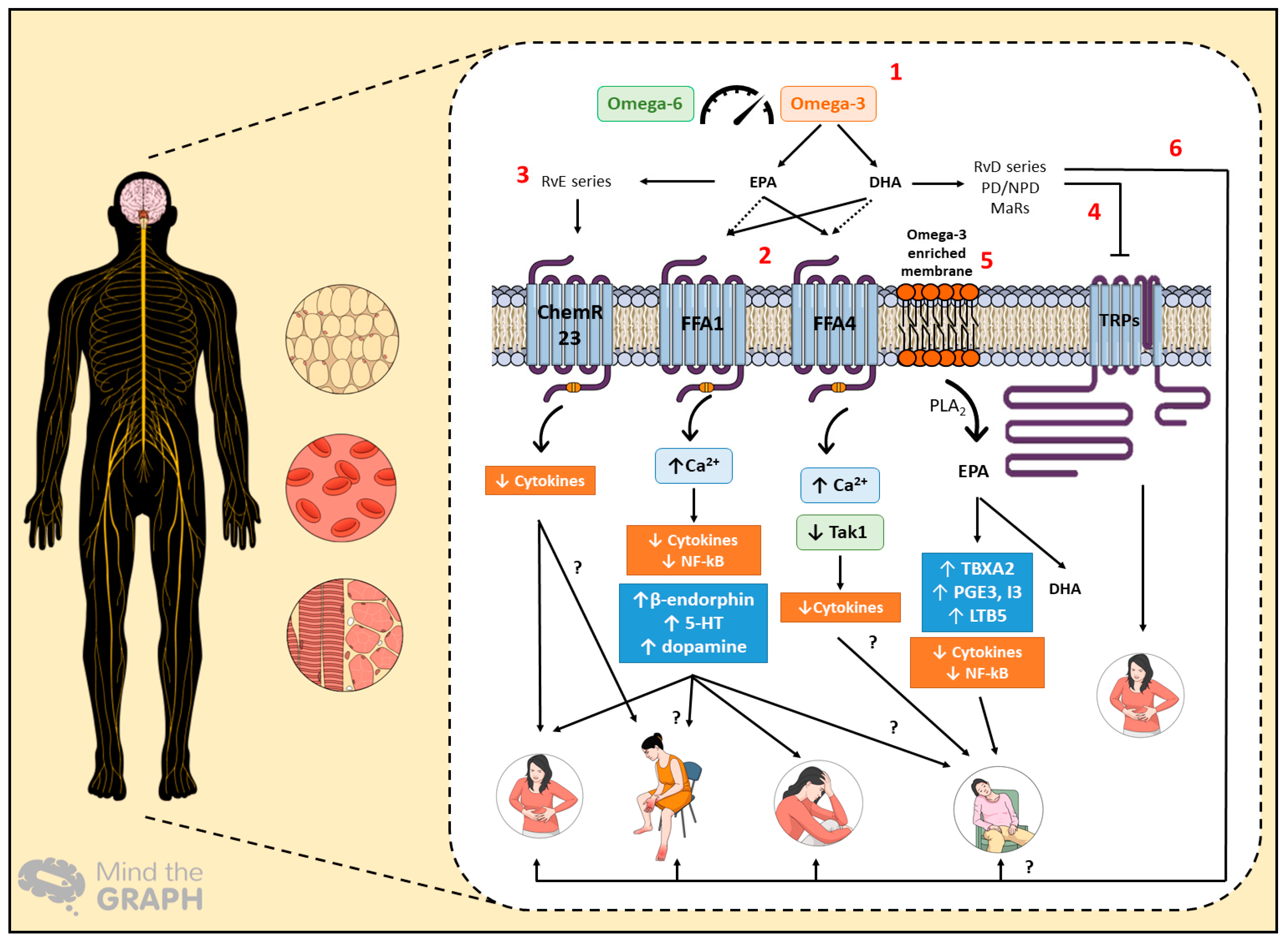
2. Omega-3 PUFAs and the Possible Mechanisms of Action in Cancer Complications
3. Omega-3 PUFAs as Part of Pharmaconutrition in Cancer Patients
4. Cancer-Related Pain
5. Anorexia-Cachexia Syndrome
6. Major Depression Disorder (MDD)
7. Paraneoplastic Syndromes
8. Literature Trends Regarding Omega-3 PUFAs and Cancer Complications
9. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dyerberg, J.; Bang, H.O.; Hjorne, N. Fatty Acid composition of the plasma lipids in Greenland Eskimos. Am. J. Clin. Nutr. 1975, 28, 958–966. [Google Scholar] [CrossRef]
- Bang, H.O.; Dyerberg, M.D.; Sinclair, H.M. The composition of the Eskimo western food in north. Am. J. Clin. Nutr. 1980, 33, 2657–2661. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, Y.; Tatsuno, I. Omega-3 polyunsaturated fatty acids for cardiovascular diseases: Present, past and future. Expert Rev. Clin. Pharm. 2017, 10, 865–873. [Google Scholar] [CrossRef]
- Simopoulos, A.P. An Increase in the Omega-6/Omega-3 Fatty Acid Ratio Increases the Risk for Obesity. Nutrients 2016, 8, 128. [Google Scholar] [CrossRef]
- Fang, X.; Ge, K.; Song, C.; Ge, Y.; Zhang, J. Biochemical and Biophysical Research Communications Effects of n-3PUFAs on autophagy and inflammation of hypothalamus and body weight in mice. Biochem. Biophys. Res. Commun. 2018, 501, 927–932. [Google Scholar] [CrossRef] [PubMed]
- Spencer, L.; Mann, C.; Metcalfe, M.; Webb, M.B.; Pollard, C.; Spencer, D. The effect of omega-3 FAs on tumour angiogenesis and their therapeutic potential. Eur. J. Cancer 2009, 45, 2077–2086. [Google Scholar] [CrossRef]
- Briscoe, C.P.; Tadayyon, M.; Andrews, J.L.; Benson, W.G.; Chambers, J.K.; Eilert, M.M. The orphan G protein-coupled receptor GPR40 is activated by medium and long chain fatty acids. J. Biol. Chem. 2003, 278, 11303–11311. [Google Scholar] [CrossRef] [PubMed]
- Hirasawa, A.; Tsumaya, K.; Awaji, T.; Katsuma, S.; Adachi, T.; Yamada, M. Free fatty acids regulate gut incretin glucagon-like peptide-1 secretion through GPR120. Nat. Med. 2005, 11, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Quesada-López, T.; Cereijo, R.; Turatsinze, J.V.; Planavila, A.; Cairó, M.; Gavaldà-Navarro, A. The lipid sensor GPR120 promotes brown fat activation and FGF21 release from adipocytes. Nat. Commun. 2016, 7, 13479. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, V.; Marinho, R.; Vitorino, D.; Santos, G.A.; Moraes, J.C.; Dragano, N. Diets containing alpha-linolenic (n-3) or oleic (n-9) fatty acids rescues obese mice from insulin resistance. Endocrinology 2015, 156, 4033–4046. [Google Scholar] [CrossRef]
- Freitas, R.D.S.; Costa, K.M.; Nicoletti, N.F.; Kist, L.W.; Bogo, M.R.; Campos, M.M. Omega-3 fatty acids are able to modulate the painful symptoms associated to cyclophosphamide-induced-hemorrhagic cystitis in mice. J. Nutr. Biochem. 2016, 27, 219–232. [Google Scholar] [CrossRef]
- Nakamoto, K.; Nishinaka, T.; Ambo, A.; Mankura, M.; Kasuya, F.; Tokuyama, S. Possible involvement of β-endorphin in docosahexaenoic acid-induced antinociception. Eur. J. Pharmacol. 2011, 666, 100–104. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global Cancer Statistics 2018: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancer in 185 Countries. Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef]
- Neufeld, N.J.; Elnahal, S.M.; Alvarez, R.H. Cancer pain: A review of epidemiology, clinical quality and value impact. Future Oncol. 2017, 13, 833–841. [Google Scholar] [CrossRef]
- Goyal, A.; Bhatnagar, S. Neuropathic pain in cancer. Ann. Palliat. Med. 2014, 3, 1–3. [Google Scholar]
- Bennet, M.I.; Kaasa, S.; Barke, A.; Korwisi, B.; Riek, W.; Treede, R.-D. The IASP classification of chronic pain for ICD-11: Chronic secondary headache and orofacial pain. Pain 2019, 160, 38–44. [Google Scholar] [CrossRef]
- Candido, K.D.; Kusper, T.M.; Knezevic, N.N. New Cancer Pain Treatment Options. Curr. Pain Headache Rep. 2017, 21, 1–12. [Google Scholar] [CrossRef]
- Argilés, J.M.; Stemmler, B.; López-Soriano, F.J.; Busquets, S. Inter-tissue communication in cancer cachexia. Nat. Rev. Endocrinol. 2018. [Google Scholar] [CrossRef]
- Fearon, K.; Strasser, F.; Anker, S.D.; Bosaeus, I.; Bruera, E.; Fainsinger, R.L. Definition and classification of cancer cachexia: An international consensus. Lancet Oncol. 2011, 12, 489–495. [Google Scholar] [CrossRef]
- Argilés, J.M.; Busquets, S.; Stemmler, B.; López-Soriano, F.J. Cachexia and sarcopenia: Mechanisms and potential targets for intervention. Curr. Opin. Pharm. 2015, 22, 100–106. [Google Scholar] [CrossRef]
- Seelaender, M.; Laviano, A.; Busquets, S.; Püschel, G.P.; Margaria, T.; Batista, M.L., Jr. Inflammation in Cachexia. Mediators Inflamm. 2015, 2015, 2–4. [Google Scholar] [CrossRef] [PubMed]
- Argilés, J.M.; Busquets, S.; Stemmler, B.; López-Soriano, F.J. Cancer cachexia: Understanding the molecular basis. Nat. Rev. Cancer 2014, 14, 754–762. [Google Scholar] [CrossRef] [PubMed]
- Bortolato, B.; Hyphantis, T.N.; Valpione, S.; Perini, G.; Maes, M.; Morris, G. Depression in cancer: The many biobehavioral pathways driving tumor progression. Cancer Treat Rev. 2017, 52, 58–70. [Google Scholar] [CrossRef] [PubMed]
- Sotelo, J.L.; Musselman, D.; Nemeroff, C. The biology of depression in cancer and the relationship between depression and cancer progression. Int. Rev. Psychiatry 2014, 26, 16–30. [Google Scholar] [CrossRef]
- Watts, S.; Prescott, P.; Mason, J.; McLeod, N.; Lewith, G. Depression and anxiety in ovarian cancer: A systematic review and meta-analysis of prevalence rates. BMJ Open 2015, 5, e007618. [Google Scholar] [CrossRef] [PubMed]
- Pelosof, L.C.; Gerber, D.E. Paraneoplastic Syndromes: An Approach to Diagnosis and Treatment. Mayo Clin. Proc. 2010, 85, 838–854. [Google Scholar] [CrossRef] [PubMed]
- Efthymiou, C.; Spyratos, D.; Kontakiotis, T. Endocrine paraneoplastic syndromes in lung cancer. Hormones 2018, 17, 351–358. [Google Scholar] [CrossRef]
- Viau, M.; Renaud, M.C.; Grégoire, J.; Sebastianelli, A.; Plante, M. Paraneoplastic syndromes associated with gynecological cancers: A systematic review. Gynecol. Oncol. 2017, 146, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Marian, M.J. Dietary Supplements Commonly Used by Cancer Survivors: Are There Any Benefits? Nutr. Clin. Pract. 2017, 32, 607–627. [Google Scholar] [CrossRef]
- Saini, R.K.; Keum, Y. Omega-3 and omega-6 polyunsaturated fatty acids: Dietary sources, metabolism, and significance—A review. Life Sci. 2018, 203, 255–267. [Google Scholar] [CrossRef]
- Simopoulos, A.P.; DiNicolantonio, J.J. The importance of a balanced ω-6 to ω-3 ratio in the prevention and management of obesity. Open Hear 2016, 3, e000385. [Google Scholar] [CrossRef]
- Simopoulos, A.P. Evolutionary aspects of diet, the omega-6/omega-3 ratio and genetic variation: Nutritional implications for chronic diseases. Biomed. Pharmacother. 2006, 60, 502–507. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P. The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomed. Pharmacother. 2002, 56, 365–379. [Google Scholar] [CrossRef]
- Jiang, J.; Li, K.; Wang, F.; Yang, B.; Fu, Y. Effect of Marine-Derived n-3 Polyunsaturated Fatty Acids on Major Eicosanoids: A Systematic Review and Meta-Analysis from 18 Randomized Controlled Trials. PLoS ONE 2016, 11, e0147351. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Fuentes, N.R.; Hou, T.Y.; Barhoumi, R.; Li, X.C.; Deutz, N.E.P. Remodelling of primary human CD4+ T cell plasma membrane order by n-3 PUFA. Br. J. Nutr. 2018, 119, 163–175. [Google Scholar] [CrossRef]
- Fuentes, N.R.; Kim, E.; Fan, Y.; Chapkin, R.S. Molecular Aspects of Medicine Omega-3 fatty acids, membrane remodeling and cancer prevention. Mol. Asp. Med. 2018, 2017, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Amézaga, J.; Arranz, S.; Urruticoechea, A.; Ugartemendia, G.; Larraioz, A.; Louka, M. Altered Red Blood Cell Membrane Fatty Acid Profile. Nutrients 2018, 10, 1853. [Google Scholar] [CrossRef] [PubMed]
- Gilroy, D.W.; Bailey, D.B. Lipid mediators in immune regulation and resolution. Br. J. Pharmacol. 2019, 176, 1009–1023. [Google Scholar] [CrossRef]
- Serhan, C.N.; Savill, J. Resolution of inflammation: The beginning programs the end. Nat. Immunol. 2005, 6, 1191–1197. [Google Scholar] [CrossRef]
- Maddox, B.J.E.; Serhan, C.N. Lipoxin A4 and B4 Are Potent Stimuli for Human Monocyte Migration and Adhesion: Selective Inactivation by Dehydrogenation and Reduction. J. Exp. Med. 1996, 183, 137–146. [Google Scholar] [CrossRef]
- Chandrasekharan, J.; Sharma-Walia, N. Lipoxins: Nature’s way to resolve inflammation. J. Inflamm. Res. 2015, 8, 181–192. [Google Scholar] [PubMed]
- Serhan, C.N.; Yacoubian, S. Anti-Inflammatory and Proresolving Lipid Mediators. Annu. Rev. Pathol. 2008, 3, 279–312. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.J.; Spite, M. Resolvins: Anti-inflammatory and proresolving mediators derived from omega-3 polyunsaturated fatty acids. Annu. Rev. Nutr. 2012, 32, 203–227. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Chiang, N.; Dalli, J.; Levy, B.D. Lipid Mediators in the Resolution of Inflammation. Cold Spring Harb. Perspect. Biol. 2015, 7, a016311. [Google Scholar] [CrossRef] [PubMed]
- Sulciner, M.L.; Serhan, C.N.; Gilligan, M.M.; Mudge, D.K.; Chang, J.; Gartung, A. Resolvins suppress tumor growth and enhance cancer therapy. J. Exp. Med. 2018, 215, 115–140. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Terrando, N.; Xu, Z.; Bang, S.; Jordt, S.; Baker, O.J. Distinct Analgesic Actions of DHA and DHA-Derived Specialized Pro-Resolving Mediators on Post-operative Pain After Bone Fracture in Mice. Front. Pharmacol. 2018, 9, 412. [Google Scholar] [CrossRef] [PubMed]
- Deyama, S.; Shimoda, K.; Suzuki, H.; Ishikawa, Y. Resolvin E1/E2 ameliorate lipopolysaccharide-induced depression-like behaviors via ChemR23. Psychopharmacology 2018, 235, 329–336. [Google Scholar] [CrossRef]
- Klein, C.P.; Sperotto, N.D.M.; Maciel, I.S.; Leite, C.E.; Souza, A.H.; Campos, M.M. Effects of D-series resolvins on behavioral and neurochemical changes in a fibromyalgia-like model in mice. Neuropharmacology 2014, 86, 57–66. [Google Scholar] [CrossRef]
- Seelaender, M.; Batista, M.; Lira, F.; Silverio, R.; Rossi-Fanelli, F. Inflammation in cancer cachexia: To resolve or not to resolve (is that the question?). Clin. Nutr. 2012, 31, 562–566. [Google Scholar] [CrossRef]
- Ma, D.; Tao, B.; Warashina, S.; Kotani, S.; Lu, L.; Kaplamadzhiev, D.B. Expression of free fatty acid receptor GPR40 in the central nervous system of adult monkeys. Neurosci. Res. 2007, 58, 394–401. [Google Scholar] [CrossRef]
- Edfalk, S.; Steneberg, P.; Edlund, H. Gpr40 is expressed in enteroendocrine cells and mediates free fatty acid stimulation of incretin secretion. Diabetes 2008, 57, 2280–2287. [Google Scholar] [CrossRef]
- Itoh, Y.; Kawamata, Y.; Harada, M.; Kobayashi, M.; Fujii, R.; Fukusumi, S. Free fatty acids regulate insulin secretion from pancreatic beta cells through GPR40. Nature 2003, 422, 173–176. [Google Scholar] [CrossRef] [PubMed]
- Gorski, J.N.; Pachanski, M.J.; Mane, J.; Plummer, C.W.; Souza, S.; Thomas-Fowlkes, B.S. GPR40 reduces food intake and body weight through GLP-1. Am. J. Physiol. Endocrinol. Metab. 2017, 313, E37–E47. [Google Scholar] [CrossRef]
- Liou, A.; Lu, X.; Sei, Y.; Zhao, X.; Pechhold, S.; Carrero, R. The G-Protein Coupled Receptor GPR40 Directly Mediates Long-Chain Fatty Acid Induced Secretion of Cholecystokinin. Gastroenterology 2011, 140, 903–912. [Google Scholar] [CrossRef]
- Dragano, N.R.V.; Solon, C.; Ramalho, A.F.; Moura RFDe Razolli, D.S.; Christiansen, E. Polyunsaturated fatty acid receptors, GPR40 and GPR120, are expressed in the hypothalamus and control energy homeostasis and inflammation. J. Neuroinflamm. 2017, 14, 91. [Google Scholar] [CrossRef]
- Li, G.; Robles, S.; Lu, Z.; Li, Y.; Krayer, J.W.; Leite, R.S. Upregulation of free fatty acid receptors in periodontal tissues of patients with metabolic syndrome and periodontitis. J. Periodontal Res. 2018. [Google Scholar] [CrossRef]
- Gravena, C.; Mathias, P.; Ashcroft, S.J. Acute effects of fatty acids on insulin secretion from rat and human islets of Langerhans. J. Endocrinol. 2002, 173, 73–80. [Google Scholar] [CrossRef][Green Version]
- Ichimura, A.; Hasegawa, S.; Kasubuchi, M.; Kimura, I. Free fatty acid receptors as therapeutic targets for the treatment of diabetes. Front. Pharmacol. 2014, 5, 236. [Google Scholar] [CrossRef]
- Yashiro, H.; Tsujihata, Y.; Takeuchi, K.; Hazama, M.; Johnson, P.R.V.; Rorsman, P. The Effects of TAK-875, a Selective G Protein-Coupled Receptor 40/Free Fatty Acid 1 Agonist, on Insulin and Glucagon Secretion in Isolated Rat and Human Islets. J. Pharmacol. Exp. Ther. 2012, 340, 483–489. [Google Scholar] [CrossRef]
- Burant, C.; Viswanathan, P.; Marcinak, J.; Cao, C.; Vakilynejad, M. TAK-875 versus placebo or glimepiride in type 2 diabetes mellitus: A phase 2, randomised, double-blind, placebo-controlled trial. Lancet 2012, 379, 1403–1411. [Google Scholar] [CrossRef]
- Harada, S.; Haruna, Y.; Aizawa, F.; Matsuura, W.; Nakamoto, K.; Yamashita, T. Involvement of GPR40, a long-chain free fatty acid receptor, in the production of central post-stroke pain after global cerebral ischemia. Eur. J. Pharmacol. 2014, 744, 115–123. [Google Scholar] [CrossRef]
- Nakamoto, K.; Nishinaka, T.; Sato, N.; Aizawa, F.; Yamashita, T.; Mankura, M. The activation of supraspinal GPR40/FFA1 receptor signalling regulates the descending pain control system. Br. J. Pharmacol. 2015, 172, 1250–1262. [Google Scholar] [CrossRef]
- Liotti, A.; Cosimato, V.; Mirra, P.; Calì, G.; Conza, D.; Secondo, A.; Luongo, G.; Terracciano, D.; Formisano, P.; Beguinot, F.; et al. Oleic acid promotes prostate cancer malignant phenotype via the G protein-coupled receptor FFA1/GPR40. J. Cell. Physiol. 2018, 233, 7367–7378. [Google Scholar] [CrossRef]
- Alexander, S.P.H.; Christopoulos, A.; Davenport, A.P.; Kelly, E.; Marrion, N.V.; Peters, J.A. The Concise Guide TO Pharmacology 2017/18:G protein-coupled receptors. Br. J. Pharmacol. 2017, 174, 17–129. [Google Scholar]
- Ulven, T.; Christiansen, E. Dietary Fatty Acids and Their Potential for Controlling Metabolic Diseases Through Activation of FFA4/GPR120. Annu. Rev. Nutr. 2015, 35, 239–263. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, R.; Veiga, C.; Bessi, M.; Dátilo, M.; Sant’Ana, M.; Rodrigues, P. Unsaturated fatty acids from flaxseed oil and exercise modulate GPR120 but not GPR40 in the liver of obese mice: A new anti-inflammatory approach. J. Nutr. Biochem. 2019, 66, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Dátilo, M.N.; Ramos, M.; Formigari, G.; Rodrigues, P.; de Moura, L.; da Silva, A. Omega-3 from Flaxseed Oil Protects Obese Mice Against Diabetic Retinopathy Through GPR120 Receptor. Sci. Rep. 2018, 8, 14318. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.H.; Park, S.; Baek, J.; Lee, S.; Baek, W.; Lee, S. Formation and Suppresses Bone Resorption in the Presence of Elevated n-3 Fatty Acid Levels. Endocrinology 2016, 4, 2621–2635. [Google Scholar] [CrossRef] [PubMed]
- Im, D.S. Functions of omega-3 fatty acids and FFA4 (GPR120) in macrophages. Eur. J. Pharmacol. 2016, 785, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Cintra, D.; Ropelle, E.; Moraes, J.; Pauli, J.; Morari, J.; de Souza, C. Unsaturated Fatty Acids Revert Diet-Induced Hypothalamic Inflammation in Obesity. PLoS ONE 2012, 7, e30571. [Google Scholar] [CrossRef] [PubMed]
- Auguste, S.; Fisette, A.; Fernandes, M.F.; Hryhorczuk, C.; Poitout, V.; Alquier, T. Central Agonism of GPR120 Acutely Inhibits Food Intake and Food Reward and Chronically Suppresses Anxiety-Like Behavior in Mice. Int. J. Neuropsychopharmacol. 2016, 19, pyw014. [Google Scholar] [CrossRef] [PubMed]
- Schilperoort, M.; van Dam, A.D.; Hoeke, G.; Shabalina, I.G.; Okolo, A.; Hanyaloglu, A.C. The GPR120 agonist TUG-891 promotes metabolic health by stimulating mitochondrial respiration in brown fat. EMBO Mol. Med. 2018, 10, e8047. [Google Scholar] [CrossRef]
- Jones, N.E.; Heyland, D.K. Pharmaconutrition: A new emerging paradigm. Curr. Opin. Gastroenterol. 2008, 24, 215–222. [Google Scholar] [CrossRef]
- Virizuela, J.A.; Camblor-Álvarez, M.; Luengo-Pérez, L.M.; Grande, E.; Álvarez-Hernández, J.; Sendrós-Madroño, M.J.; Jiménez-Fonseca, P.; Cervera-Peris, M.; Ocón-Bretón, M.J. Nutritional support and parenteral nutrition in cancer patients: An expert consensus report. Clin. Transl. Oncol. 2018, 20, 619–629. [Google Scholar] [CrossRef] [PubMed]
- Ryan, A.M.; Power, D.G.; Daly, L.; Cushen, S.J.; Ní, E. Cancer-associated malnutrition, cachexia and sarcopenia: The skeleton in the hospital closet 40 years later. Proc. Nutr. Soc. 2016, 75, 199–211. [Google Scholar] [CrossRef]
- Gangadharan, A.; Choi, S.E.; Hassan, A.; Ayoub, N.M.; Balwani, S.; Kim, Y.H.; Pecora, A.; Goy, A.; Suh, K.S. Protein calorie malnutrition, nutritional intervention and personalized cancer care. Oncotarget 2017, 8, 24009–24030. [Google Scholar] [CrossRef]
- Mcmillan, D.C. The systemic inflammation-based Glasgow Prognostic Score: A decade of experience in patients with cancer. Cancer Treat. Rev. 2013, 39, 534–540. [Google Scholar] [CrossRef] [PubMed]
- Diakos, C.I.; Charles, K.A.; Mcmillan, D.C.; Clarke, S.J. Cancer-related inflammation and treatment effectiveness. Lancet Oncol. 2014, 15, 493–503. [Google Scholar] [CrossRef]
- Talvas, J.; Garrait, G.; Goncalves-mendes, N.; Rouanet, J. Immunonutrition stimulates immune functions and antioxidant defense capacities of leukocytes in radiochemotherapy-treated head & neck and esophageal cancer patients: A double-blind randomized clinical trial. Clin. Nutr. 2015, 34, 810–817. [Google Scholar] [CrossRef]
- Hamza, N.; Darwish, A.; Reilly, D.A.O.; Denton, J.; Sheen, A.J.; Chang, D. Perioperative Enteral Immunonutrition Modulates Systemic and Mucosal Immunity and the Inflammatory Response in Patients with Periampullary Cancer Scheduled for Pancreaticoduodenectomy A Randomized Clinical Trial. Pancreas 2015, 44, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Takahashi, T.; Yamaguchi, K.; Osada, S. Elemental diet plus glutamine for the prevention of mucositis in esophageal cancer patients receiving chemotherapy: A feasibility study. Support Care Cancer 2016, 24, 933–941. [Google Scholar] [CrossRef] [PubMed]
- Mudge, L.A.; Watson, D.I.; Smithers, B.M.; Isenring, E.A.; Smith, L.; Jamieson, G.G. Multicentre factorial randomized clinical trial of perioperative immunonutrition versus standard nutrition for patients undergoing surgical resection of oesophageal cancer. Br. J. Surg. 2018, 105, 1262–1272. [Google Scholar] [CrossRef] [PubMed]
- Klek, S.; Scislo, L.; Walewska, E.; Choruz, R.; Galas, A. Enriched enteral nutrition may improve short-term survival in stage IV gastric cancer patients: A randomized, controlled trial. Nutrition 2017, 36, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Martins, E.; Oliveira, A.C.D.M.; Pizato, N.; Muniz-junqueira, M.I.; Magalhães, K.G.; Nakano, E.Y. The effects of EPA and DHA enriched fish oil on nutritional and immunological markers of treatment naïve breast cancer patients: A randomized double-blind controlled trial. Nutr. J. 2017, 16, 71. [Google Scholar] [CrossRef]
- Chagas, T.R.; Borges, D.S.; Oliveira PFDe Mocellin, M.C.; Barbosa, A.M.; Camargo, C.Q. Oral fish oil positively influences nutritional-inflammatory risk in patients with haematological malignancies during chemotherapy with an impact on long-term survival: A randomised clinical trial. J. Hum. Nutr. Diet. 2017, 30, 681–692. [Google Scholar] [CrossRef] [PubMed]
- Bougnoux, P.; Hajjaji, N.; Ferrasson, M.N.; Giraudeau, B.; Couet, C.; Floch, O.L. Improving outcome of chemotherapy of metastatic breast cancer by docosahexaenoic acid: A phase II trial. Br. J. Cancer 2009, 101, 1978–1985. [Google Scholar] [CrossRef] [PubMed]
- Ouldamer, L.; Goupille, C.; Vildé, A.; Arbion, F.; Body, G. N-3 Polyunsaturated Fatty Acids of Marine Origin and Multifocality in Human Breast Cancer. PLoS ONE 2016, 11, e0147148. [Google Scholar] [CrossRef]
- Caraceni, A.; Portenoy, R. An international survey of cancer pain characteristics and syndromes. Pain 1999, 82, 263–274. [Google Scholar] [CrossRef]
- Lara-Solares, A.; Ahumada Olea, M.; Basantes Pinos A de los, Á.; Bistre Cohén, S.; Bonilla Sierra, P.; Duarte Juárez, E.R. Latin-American guidelines for cancer pain management. Pain Manag. 2017, 7, 287–298. [Google Scholar] [CrossRef]
- Falk, S.; Dickenson, A.H. Pain and Nociception: Mechanisms of Cancer-Induced Bone Pain. J. Clin. Oncol. 2014, 32, 1647–1654. [Google Scholar] [CrossRef] [PubMed]
- Elinav, E.; Nowarski, R.; Thaiss, C.A.; Hu, B.; Jin, C.; Flavell, R.A. Inflammation-induced cancer: Crosstalk between tumours, immune cells and microorganisms. Nat. Rev. Cancer 2013, 13, 759–771. [Google Scholar] [CrossRef] [PubMed]
- Shinko, D.; Diakos, C.I.; Clarke, S.J.; Charles, K.A. Cancer-Related Systemic Inflammation: The Challenges and Therapeutic Opportunities for Personalized Medicine. Clin. Pharmacol. Ther. 2017, 102, 599–610. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.D.A.P.; de Souza Fabre, M.E.; Waitzberg, D.L. Omega-3 supplements for patients in chemotherapy and/or radiotherapy: A systematic review. Clin. Nutr. 2015, 34, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Trabal, J.; Leyes, P.; Forga, M.; Maurel, J. Potential usefulness of an EPA-enriched nutritional supplement on chemotherapy tolerability in cancer patients without overt malnutrition. Nutr. Hosp. 2010, 25, 736–740. [Google Scholar] [PubMed]
- Ye, Y.; Scheff, N.; Bernabé, D.; Salvo, E.; Ono, K.; Cheng, L. Anti-cancer and analgesic effects of resolvin D2 in oral squamous cell carcinoma. Neuropharmacology 2018, 139, 182–193. [Google Scholar] [CrossRef] [PubMed]
- Hershman, D.L.; Unger, J.M.; Crew, K.D.; Awad, D.; Dakhil, S.R.; Gralow, J. Randomized Multicenter Placebo-Controlled Trial of Omega-3 Fatty Acids for the Control of Aromatase Inhibitor—Induced Musculoskeletal Pain: SWOG S0927. J. Clin. Oncol. 2015, 33, 1910–1917. [Google Scholar] [CrossRef]
- Shen, S.; Unger, J.M.; Crew, K.D.; Till, C.; Greenlee, H.; Gralow, J. Omega-3 fatty acid use for obese breast cancer patients with aromatase inhibitor-related arthralgia (SWOG S0927). Breast Cancer Res. Treat. 2018, 172, 603–610. [Google Scholar] [CrossRef]
- Martínez, N.; Herrera, M.; Frías, L.; Provencio, M.; Pérez-Carrión, R.; Díaz, V. A combination of hydroxytyrosol, omega-3 fatty acids and curcumin improves pain and inflammation among early stage breast cancer patients receiving adjuvant hormonal therapy: Results of a pilot study. Clin. Transl. Oncol. 2019, 21, 489–498. [Google Scholar] [CrossRef]
- Ghoreishi, Z.; Esfahani, A.; Djazayeri, A.; Djalali, M.; Golestan, B.; Ayromlou, H. Omega-3 fatty acids are protective against paclitaxel-induced peripheral neuropathy: A randomized double-blind placebo controlled trial. BMC Cancer 2012, 12, 355. [Google Scholar] [CrossRef]
- Maschio, M.; Zarabla, A.; Maialetti, A.; Marchesi, F.; Giannarelli, D.; Gumenyuk, S. Prevention of Bortezomib-Related Peripheral Neuropathy with Docosahexaenoic Acid and α-Lipoic Acid in Patients with Multiple Myeloma: Preliminary Data. Integr. Cancer Ther. 2018, 17, 1115–1124. [Google Scholar] [CrossRef]
- Lavriv, D.S.; Neves, P.M.; Ravasco, P. Should omega-3 fatty acids be used for adjuvant treatment of cancer cachexia? Clin. Nutr. ESPEN 2018, 25, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Mattox, T.W. Cancer Cachexia: Cause, Diagnosis, and Treatment Causes of CC. Nutr. Clin. Pract. 2017, 5, 599–606. [Google Scholar] [CrossRef]
- Hanai, N.; Terada, H.; Hirakawa, H.; Suzuki, H.; Nishikawa, D.; Beppu, S. Prospective randomized investigation implementing immunonutritional therapy using a nutritional supplement with a high blend ratio of ω-3 fatty acids during the perioperative period for head and neck carcinomas. Jpn. J. Clin. Oncol. 2018, 48, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Persson, C.; Glimelius, B.; Rönnelid, J.; Nygren, P. Impact of fish oil and melatonin on cachexia in patients with advanced gastrointestinal cancer: A randomized pilot study. Nutrition 2005, 21, 170–178. [Google Scholar] [CrossRef]
- Shirai, Y.; Okugawa, Y.; Hishida, A.; Ogawa, A.; Okamoto, K. Fish oil-enriched nutrition combined with systemic chemotherapy for gastrointestinal cancer patients with cancer cachexia. Sci. Rep. 2017, 7, 4826. [Google Scholar] [CrossRef] [PubMed]
- Ries, A.; Trottenber, P.; Elsner, F.; Stiel, S.; Haugen, D.; Kaasa, S. A systematic review on the role of fish oil for the treatment of cachexia in advanced cancer: An EPCRC cachexia guidelines project. Palliat. Med. 2011, 26, 294–304. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, M.; Haji, S.; Amagai, T. Elevated Serum AA/EPA Ratio as a Predictor of Skeletal Muscle Depletion in Cachexic Patients with Advanced Gastro-intestinal Cancers. In Vivo 2017, 31, 1003–1009. [Google Scholar]
- Murphy, R.A.; Mourtzakis, M.; Chu, Q.S.; Reiman, T.; Mazurak, V.C. Skeletal Muscle Depletion Is Associated with Reduced Plasma (n-3) Fatty Acids in Non-Small. J. Nutr. 2010, 140, 1602–1606. [Google Scholar] [CrossRef] [PubMed]
- Gorjao, R.; Miguel, C.; Duarte, T.; Serdan, A.; Leonardo, V.; Diniz, S. New insights on the regulation of cancer cachexia by N-3 polyunsaturated fatty acids. Pharmacol. Ther. 2019, 196, 117–134. [Google Scholar] [CrossRef]
- Solís-martínez, O.; Plasa-carvalho, V.; Phillips-sixtos, G. Effect of Eicosapentaenoic Acid on Body Composition and Inflammation Markers in Patients with Head and Neck Squamous Cell Cancer from a Public Hospital in Mexico. Nutr. Cancer 2018, 70, 663–670. [Google Scholar] [CrossRef]
- Pappalardo, G.; Almeida, A.; Ravasco, P. Eicosapentaenoic acid in cancer improves body composition and modulates metabolism. Nutrition 2015, 31, 549–555. [Google Scholar] [CrossRef]
- Colomer, R.; Moreno-Nogueira, J.; García-Luna, P.; García-Peris, P.; García-de-Lorenzo, A.; Zarazaga, A. Systematic Review n-3 Fatty acids, cancer and cachexia: A systematic review of the literature. Br. J. Nutr. 2007, 97, 823–831. [Google Scholar] [CrossRef] [PubMed]
- Mazzotta, P.; Jeney, C.M. Anorexia-Cachexia Syndrome: A Systematic Review of the Role of Dietary Polyunsaturated Fatty Acids in the Management of Symptoms, Survival, and Quality of Life. J. Pain Symptom Manag. 2008, 37, 1069–1077. [Google Scholar] [CrossRef] [PubMed]
- Russell, S.T.; Tisdale, M.J.Ã. Effect of eicosapentaenoic acid (EPA) on expression of a lipid mobilizing factor in adipose tissue in cancer cachexia. Prostaglandins Leukot. Essent Fat. Acids 2005, 72, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Yang, Y.; Wang, Y.; Xue, C. EPA-enriched phospholipids ameliorate cancer-associated cachexia mainly via inhibiting lipolysis. Food Funct. 2015, 6, 3652–3662. [Google Scholar] [CrossRef] [PubMed]
- Penna, F.; Busquets, S.; Pin, F.; Toledo, M.; Baccino, F.M.; López-Soriano, F.J. Combined approach to counteract experimental cancer cachexia: Eicosapentaenoic acid and training exercise. J. Cachexia Sarcopenia Muscle 2011, 2, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Hajjaji, N.; Couet, C.; Besson, P.; Bougnoux, P. DHA Effect on Chemotherapy-Induced Body Weight Loss: An Exploratory Study in a Rodent Model of Mammary Tumors. Nutr. Cancer 2012, 61, 37–41. [Google Scholar] [CrossRef]
- Schiessel, D.L.; Yamazaki, R.K.; Kryczyk, M.; Coelho, I.; Yamaguchi, A.A.; Pequito, D.C.T. α-Linolenic Fatty Acid Supplementation Decreases Tumor Growth and Cachexia Parameters in Walker 256 Tumor-Bearing Rats. Nutr. Cancer 2015, 67, 839–846. [Google Scholar] [CrossRef] [PubMed]
- Werner, K.; Küllenberg de Gaudry, D.; Taylor, L.A.; Keck, T.; Unger, C.; Hopt, U.T. Dietary supplementation with n-3-fatty acids in patients with pancreatic cancer and cachexia: Marine phospholipids versus fish oil—A randomized controlled double-blind trial. Lipids Health Dis. 2017, 16, 104. [Google Scholar] [CrossRef] [PubMed]
- Muzio, G.; Ricci, M.; Traverso, N.; Monacelli, F.; Oraldi, M.; Maggiora, M. 4-Hydroxyhexenal and 4-hydroxynonenal are mediators of the anti-cachectic effect of n-3 and n-6 polyunsaturated fatty acids on human lung cancer cells. Free Radic. Biol. Med. 2016, 99, 63–70. [Google Scholar] [CrossRef]
- Caruso, R.; Nanni, M.; Riba, M.; Sabato, S.; Mitchell, A.; Croce, E. Depressive spectrum disorders in cancer: Prevalence, risk factors and screening for depression: A critical review. Acta Oncol. 2017, 56, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Young, K.; Singh, G. Biological Mechanisms of Cancer-Induced Depression. Front. Psychiatry 2018, 9, 299. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.R. Depression in cancer patients: Pathogenesis, implications and treatment (Review). Oncol. Lett. 2015, 9, 1509–1514. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Zhou, F.; He, C.; Hui, L.; Huang, T.; Wei, Y. Leptin-LepRb Expressed in Gastric Cancer Patients and Related to Cancer-Related Depression. Biomed Res. Int. 2017, 2017, 1–7. [Google Scholar] [CrossRef]
- Suzuki, S.; Akechi, T.; Kobayashi, M.; Taniguchi, K.; Goto, K.; Sasaki, S. Daily omega-3 fatty acid intake and depression in Japanese patients with newly diagnosed lung cancer. Br. J. Cancer 2004, 90, 787–793. [Google Scholar] [CrossRef]
- Kobayakawa, M.; Yamawaki, S.; Hamazaki, K.; Akechi, T.; Inagaki, M.; Uchitomi, Y. Levels of omega-3 fatty acid in serum phospholipids and depression in patients with lung cancer. Br. J. Cancer 2005, 93, 1329–1333. [Google Scholar] [CrossRef] [PubMed]
- Bigornia, S.J.; Harris, W.S.; Falc, L.M.; Tucker, K.L. The Omega-3 Index Is Inversely Associated with Depressive Symptoms among Individuals with Elevated Oxidative Stress Biomarkers 1–3. J. Nutr. 2016, 146, 758–766. [Google Scholar] [CrossRef] [PubMed]
- Müller, C.P.; Reichel, M.; Mühle, C.; Rhein, C.; Gulbins, E.; Kornhuber, J. Brain membrane lipids in major depression and anxiety disorders. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2015, 1851, 1052–1065. [Google Scholar] [CrossRef]
- Smith, D.J.; Sarris, J.; Dowling, N.; Connor, M.O.; Chee, H.; Smith, D.J. Adjunctive low-dose docosahexaenoic acid (DHA) for major depression: An open-label pilot trial. Nutr. Neurosci. 2017, 21, 224–228. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Villegas, A.; Álvarez-Pérez, J.; Toledo, E.; Salas-Salvadó, J.; Ortega-Azorín, C.; Zomeño, M. Seafood Consumption, Omega-3 Fatty Acids Intake, and Life-time Prevalence of Depression in the PREDIMED-Plus Trial. Nutrients 2018, 10, 2000. [Google Scholar] [CrossRef] [PubMed]
- Parletta, N.; Zarnowiecki, D.; Cho, J.; Wilson, A.; Procter, N.; Gordon, A. People with schizophrenia and depression have a low omega-3 index. Prostaglandins Leukot. Essent. Fat Acids 2016, 110, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Orchard, T.S.; Kellie, M.M.G.; Devries, A.C. Clearing the fog: A review of the effects of dietary omega-3 fatty acids and added sugars on chemotherapy-induced cognitive deficits. Breast Cancer Res. Treat. 2017, 161, 391–398. [Google Scholar] [CrossRef]
- Dang, R.; Zhou, X.; Tang, M.; Xu, P.; Gong, X. Fish oil supplementation attenuates neuroinflammation and alleviates depressive-like behavior in rats submitted to repeated lipopolysaccharide. Eur. J. Nutr. 2018, 57, 893–906. [Google Scholar] [CrossRef]
- Kang, J.X. Fat-1 transgenic mice: A new model for omega-3 research. Prostaglandins Leukot. Essent. Fat Acids 2007, 77, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Gu, M.; Li, Y.; Tang, H.; Zhang, C.; Li, W.; Zhang, Y. Endogenous Omega (n)-3 Fatty Acids in Fat-1 Mice Attenuated Depression-Like Behavior, Imbalance between Microglial M1 and M2 Phenotypes, and Dysfunction of Neurotrophins Induced by Lipopolysaccharide Administration. Nutrients 2018, 10, 1351. [Google Scholar] [CrossRef]
- Larrieu, T.; Layé, S. Food for mood: Relevance of nutritional omega-3 fatty acids for depression and anxiety. Front. Physiol. 2018, 9, 1047. [Google Scholar] [PubMed]
- Nishinaka, T.; Yamashita, T.; Nakamoto, K.; Kasuya, F.; Tokuyama, S. Involvement of the Long-Chain Fatty Acid Receptor GPR40 in Depression-Related Behavior. J. Pharmacol. Sci. 2014, 115, 112–115. [Google Scholar] [CrossRef]
- Aizawa, F.; Nishinaka, T.; Yamashita, T.; Nakamoto, K.; Kurihara, T.; Hirasawa, A. GPR40/FFAR1 deficient mice increase noradrenaline levels in the brain and exhibit abnormal behavior. J. Pharmacol. Sci. 2016, 132, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Aizawa, F.; Ogaki, Y.; Kyoya, N.; Nishinaka, T.; Nakamoto, K.; Kurihara, T. The Deletion of GPR40/FFAR1 Signaling Damages Maternal Care and Emotional Function in Female Mice. Biol. Pharm. Bull. 2017, 40, 1255–1259. [Google Scholar] [CrossRef][Green Version]
- Deyama, S.; Ishikawa, Y.; Yoshikawa, K.; Ide, S.; Satoh, M. Resolvin D1 and D2 Reverse Lipopolysaccharide-Induced Depression-Like Behaviors Through the mTORC1 Signaling Pathway. Int. J. Neuropsychopharmacol. 2017, 20, 575–584. [Google Scholar] [CrossRef]
- Deyama, S.; Shimoda, K.; Ikeda, H.; Fukuda, H.; Shuto, S. Resolvin E3 attenuates lipopolysaccharide-induced depression-like behavior in mice. J. Pharmacol. Sci. 2018, 138, 86–88. [Google Scholar] [CrossRef]
- Ishikawa, Y.; Deyama, S.; Shimoda, K.; Yoshikawa, K.; Ide, S. Rapid and sustained antidepressant effects of resolvin D1 and D2 in a chronic unpredictable stress model. Behav. Brain Res. 2017, 332, 233–236. [Google Scholar] [CrossRef]
- Chhetry, B.; Hezghia, A.; Miller, J.; Lee, S.; Rubin-Falcone, H.; Cooper, T. Omega-3 polyunsaturated fatty acid supplementation and white matter changes in major depression. J. Psychiatr. Res. 2016, 75, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.-Q.; Dang, R.-L.; Tag, M.-M.; Cai, H.-L.; Li, H.-D.; Liao, D.-H. Long Chain Omega-3 Polyunsaturated Fatty Acid Supplementation Alleviates Doxorubicin-Induced Depressive-Like Behaviors and Neurotoxicity in Rats: Involvement of Oxidative Stress and Neuroinflammation. Nutrients 2016, 8, 243. [Google Scholar] [CrossRef]
- Dimitriadis, G.K.; Angelousi, A.; Weickert, M.O.; Randeva, H.S. Paraneoplastic endocrine syndromes. Endocr. Relat Cancer 2017, 24, 173–190. [Google Scholar] [CrossRef]
- Kir, S.; White, J.P.; Kleiner, S.; Kazak, L.; Cohen, P.; Baracos, V.E. Tumour-derived PTH-related protein triggers adipose tissue browning and cancer cachexia. Nature 2014, 513, 100–104. [Google Scholar] [CrossRef]
- Azuma, M.; Gomes-Filho, J.; Ervolino, E.; Pipa, C.; Cardoso, C.; Andrada, A.C. Omega 3 Fatty Acids Reduce Bone Resorption While Promoting Bone Generation in Rat. J. Endod. 2017, 43, 970–976. [Google Scholar] [CrossRef]
- Tartibian, B.; Maleki, B.H.; Abbasi, A. The calciotropic hormone response to omega-3 supplementation during long-term weight-bearing exercise training in post menopausal women. J. Sport Sci. Med. 2010, 9, 245–252. [Google Scholar]
- Graus, F.; Dalmau, J. Paraneoplastic neurological syndromes. Curr. Opin. Neurol. 2012, 25, 795–801. [Google Scholar] [CrossRef]
- Sioka, C.; Fotopoulos, A. Paraneoplastic immune-mediated neurological effects of systemic cancers. Expert Rev. Clin. Immunol. 2014, 10, 621–630. [Google Scholar] [CrossRef]
- Chen, X.; Wu, S.; Chen, C.; Xie, B.; Fang, Z.; Hu, W. Omega-3 polyunsaturated fatty acid supplementation attenuates microglial-induced inflammation by inhibiting the HMGB1/TLR4/NF-κB pathway following experimental traumatic brain injury. J. Neuroinflamm. 2017, 14, 143. [Google Scholar] [CrossRef] [PubMed]
- Obrosov, A.; Coppey, L.J.; Shevalye, H.; Yorek, M.A. Effect of Fish Oil vs. Resolvin D1, E1, Methyl Esters of Resolvins D1 or D2 on Diabetic Peripheral Neuropathy. J. Neurol. Neurophysiol. 2018, 8, 453. [Google Scholar] [CrossRef] [PubMed]
- Yorek, M.S.; Coppey, L.J.; Shevalye, H.; Obrosov, A.; Kardon, R.H.; Yorek, M.A. Effect of Treatment with Salsalate, Menhaden Oil, Combination of Salsalate and Menhaden Oil, or Resolvin D1 of C57Bl/6J Type 1 Diabetic Mouse on Neuropathic Endpoints. J. Nutr. Metab. 2016, 2016, 5905891. [Google Scholar] [CrossRef] [PubMed]
- Azar, L.; Khasnis, A. Paraneoplastic rheumatologic syndromes. Curr. Opin. Rheumatol. 2013, 25, 44–49. [Google Scholar] [CrossRef]
- Manger, B.; Schett, G. Paraneoplastic syndromes in rheumatology. Nat. Rev. Rheumatol. 2014, 10, 662–670. [Google Scholar] [CrossRef] [PubMed]
- Barrouin-melo, S.M.; Anturaniemi, J.; Sankari, S.; Griinari, M.; Atroshi, F. Evaluating oxidative stress, serological-and haematological status of dogs suffering from osteoarthritis, after supplementing their diet with fish or corn oil. Lipids Health Dis. 2016, 15, 139. [Google Scholar] [CrossRef]
- Dawczynski, C.; Dittrich, M.; Neumann, T.; Goetze, K.; Welzel, A.; Oelzner, P. Docosahexaenoic acid in the treatment of rheumatoid arthritis: A double-blind, placebo-controlled, randomized cross-over study with microalgae vs. sunflower oil. Clin. Nutr. 2018, 37, 494–504. [Google Scholar] [CrossRef]
- Capel, F.; Acquaviva, C.; Pitois, E.; Laillet, B.; Rigaudière, J.; Jouve, C. DHA at nutritional doses restores insulin sensitivity in skeletal muscle by preventing lipotoxicity and inflammation. J. Nutr. Biochem. 2015, 26, 949–959. [Google Scholar] [CrossRef]
- Minisola, S.; Peacock, M.; Fukumoto, S.; Cipriani, C.; Pepe, J.; Tella, S.H. Tumour-induced osteomalacia. Nat. Rev. Dis. Prim. 2017, 3, 17044. [Google Scholar] [CrossRef]
- Baia, L.C.; Van den Berg, E.; Vervloet, M.G.; Heilberg, I.P.; Navis, G.; Bakker, S.J.L.; Geleijnse, J.M.; Kromhout, D.; Soedamah-Muthu, S.S.; De Borst, M.H. Fish and omega-3 fatty acid intake in relation to circulating fibroblast growth factor 23 levels in renal transplant recipient. Nutr. Metab. Cardiovasc. Dis. 2014, 24, 1310–1316. [Google Scholar] [CrossRef]
- Buggiani, G.; Krysenka, A.; Grazzini, M.; Vasku, V.; Hercogova, J.; Lotti, T. Paraneoplastic vasculitis and paraneoplastic vascular syndromes. Dermatol. Ther. 2010, 23, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Wang, P.; Yang, S. Cosmetic and Therapeutic Applications of Fish Oil’s Fatty Acids on the Skin. Mar. Drugs 2018, 16, 256. [Google Scholar] [CrossRef]
- McDaniel, J.C.; Belury, M.; Ahijevych, K.; Blakely, W. Omega-3 fatty acids effect on wound healing. Wound Repair Regen. 2008, 16, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Miyata, J.; Arita, M. Role of omega-3 fatty acids and their metabolites in asthma and allergic diseases. Allergol. Int. 2015, 64, 27–34. [Google Scholar] [CrossRef]
- Mullard, A. 2018 FDA drug approvals. Nat. Rev. Drug Discov. 2019, 18, 85–89. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Authors | Cancer-Related Complication | Species | Cancer Type | Treatment Scheme | Major Outcome |
|---|---|---|---|---|---|
| Hershmann et al., 2015 [96] | Aromatase-inhibitor associated arthralgia | Human | Breast cancer | 3.3 g 1 FO (560 mg EPA + DHA; 40:20) | Decreased pain, evaluated by the 2 BPI between the baseline and week 24 (p < 0.01) |
| Shen et al., 2018 [97] | Aromatase-inhibitor associated arthralgia | Human | Breast cancer (obese) | 3.3 g FO (560 mg EPA + DHA; 40:20) | Pain reduction in 3 BMI > 30 kg/m² patients (p = 0.02) |
| Martínez et al., 2018 [98] | Aromatase-inhibitor musculoskeletal symptoms (AIMSS) | Human | Breast cancer | 460 mg EPA + DHA 12.5 mg hydroxytyrosol 50 g curcumin | Decrease of the BPI total score after 30 days (p = 0.011) |
| Ghroreishi et al., 2012 [99] | Paclitaxel-induced neuropathy | Human | Breast cancer | 640 mg FO (54% DHA + 10% EPA) | 70% did not develop neuropathy no pain score assessed |
| Maschio et al., 2018 [100] | Bortezomib-related neuropathy | Human | Multiple myeloma | Neuronorm® (400 mg DHA + 600 mg ALA) | Pain failed to increase significantly (p = 0.33) |
| Freitas et al., 2016 [11] | Cyclophosphamide-induced hemorrhagic cystitis | Mice | - | 20% FO-enriched diet or 1 µmol/kg i.p. | Decrease in spontaneous pain behavior and abdominal allodynia (p < 0.01) |
| Ye et al., 2018 [95] | Oral and paw cancer pain | Mice | Oral squamous cell carcinoma | RvD1 (100 ng or 200 ng) or RvD2 (100 ng or 200 ng) i.p. | RvD2 inhibited thermal and mechanical pain; RvD1 inhibited thermal pain |
| Authors | Cancer-Related Complication | Species | Cancer Type | Treatment Scheme | Major Outcome |
|---|---|---|---|---|---|
| Hanai et al., 2018 [103] | Cachexia-anorexia syndrome | Human | Head and neck squamous cell carcinoma | Prosure® (1056 mg EPA) | No significant difference among experimental groups |
| Persson et al., 2005 [104] | Cachexia-anorexia syndrome | Human | Advanced gastrointestinal cancer | 30 mL/d 1 FO (4.9g EPA + 3.2 g DHA) | FO stabilized weight in 27% patients |
| Shirai et al., 2017 [105] | Cachexia-anorexia syndrome | Human | Advanced gastrointestinal cancer | Prosure® (1.1 g EPA + 0.5 g DHA) | Increase of body weight and lean body mass (p = 0.002/p < 0.001) |
| Werner et al., 2017 [119] | Cachexia-anorexia syndrome | Human | Pancreatic cancer | 6.9 g EPA/13.6 g DHA in 100 g or 8.5 g EPA/ 12.3 g DHA in 100g | No significant differences between omega-3 PUFA treatments |
| Solis-Martínez et al., 2018 [110] | Cachexia-anorexia syndrome | Human | Head and neck squamous cell carcinoma | 2 g EPA | Weight and 2 LBM maintenance |
| Hajjaji et al., 2012 [117] | Chemotherapy-induced cachexia | Rat | Chemically-induced tumor + doxorubicin treatment | DHA-enriched diet (80 g/kg diet) | DHA diet avoided weight loss |
| Schissel et al., 2015 [118] | Cancer-associated cachexia | Rat | Breast carcinoma (Walker 256 cell line) | 53.6% EPA + DHA or 54.4% ALA | ALA and EPA improved weight gain (cachectic vs. cachectic + omega-3 p < 0.05) |
| Du et al., 2015 [115] | Cancer-related cachexia | Mice | Sarcoma (S180 cell line) | 42% EPA + 6.8% DHA | Decreased lipolysis and increased body weight (p < 0.001) |
| Penna et al., 2011 [116] | Cancer-related cachexia | Mice | Lewis lung carcinoma | EPA (0.5 g/kg) or EPA (0.5 g/kg) + exercise | EPA + exercise significantly improved muscle weight (p < 0.05) |
| Muzio et al., 2016 [120] | Cachexia in vitro model | Human | Lung adenocarcinoma | 50 µM EPA + DHA | Myoblast formation |
| Authors | Clinical or Experimental Condition | Species | Treatment Scheme | Major Outcome |
|---|---|---|---|---|
| Chhetry et al, 2016 [143] | MDD | Human | 4 g 1 FO (1.6 g EPA + 0.8 g DHA) | Improved MDD-related white matter deficiency |
| Smith et al., 2017 [129] | MDD | Human | 260 mg or 520 mg DHA | 54% of patients showed a reduction of depression severity ≥ 50% |
| Wu et al., 2018 [144] | Chemotherapy-induced depression | Rat | 1.5 g/kg omega-3 PUFAs (34% EPA + 24% DHA) | PUFAs inhibited depressive-like behaviors (p < 0.001) |
| Dang et al., 2018 [133] | LPS-induced depression | Rat | 1.5 g/kg omega-3 PUFAs (34% EPA + 24% DHA) | Omega-3 PUFAs decreased depressive behavior (p < 0.001) |
| Nishinaka et al., 2014 [137] | Behavioral despair paradigm | Mice | GW9508 (1.0, 10 or 25 µg/mouse) i.c.v | FFA1 activation decreased immobility in a tail suspension test (p < 0.05) |
| Deyama et al., 2017 [140] | LPS-induced depression | Mice | RvD1 (10 ng i.c.v.) or RvD2 (10 ng i.c.v.) | Both treatments inhibited depressive-like behaviors (p < 0.005) |
| Deyama et al., 2018 [141] | LPS-induced depression | Mice | RvE3 (10 and 100 ng i.c.v.) | Inhibition of depressive behavior (p < 0.005) |
| Ishikawa et al., 2017 [142] | Chronic unpredictable stress-related depression | Mice | RvD1 (10 ng i.c.v.) or RvD2 (10 ng i.c.v.) | Both treatments inhibited depressive behavior for 24 h (p < 0.05) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Freitas, R.D.S.; Campos, M.M. Protective Effects of Omega-3 Fatty Acids in Cancer-Related Complications. Nutrients 2019, 11, 945. https://doi.org/10.3390/nu11050945
Freitas RDS, Campos MM. Protective Effects of Omega-3 Fatty Acids in Cancer-Related Complications. Nutrients. 2019; 11(5):945. https://doi.org/10.3390/nu11050945
Chicago/Turabian StyleFreitas, Raquel D. S., and Maria M. Campos. 2019. "Protective Effects of Omega-3 Fatty Acids in Cancer-Related Complications" Nutrients 11, no. 5: 945. https://doi.org/10.3390/nu11050945
APA StyleFreitas, R. D. S., & Campos, M. M. (2019). Protective Effects of Omega-3 Fatty Acids in Cancer-Related Complications. Nutrients, 11(5), 945. https://doi.org/10.3390/nu11050945