Dietary Docosahexaenoic Acid (DHA) and Eicosapentaenoic Acid (EPA) Operate by Different Mechanisms to Modulate Hepatic Steatosis and Hyperinsulemia in fa/fa Zucker Rats


 ,
, 

Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Serum Biochemistry
2.3. Hepatic Lipid and Fatty Acid Composition
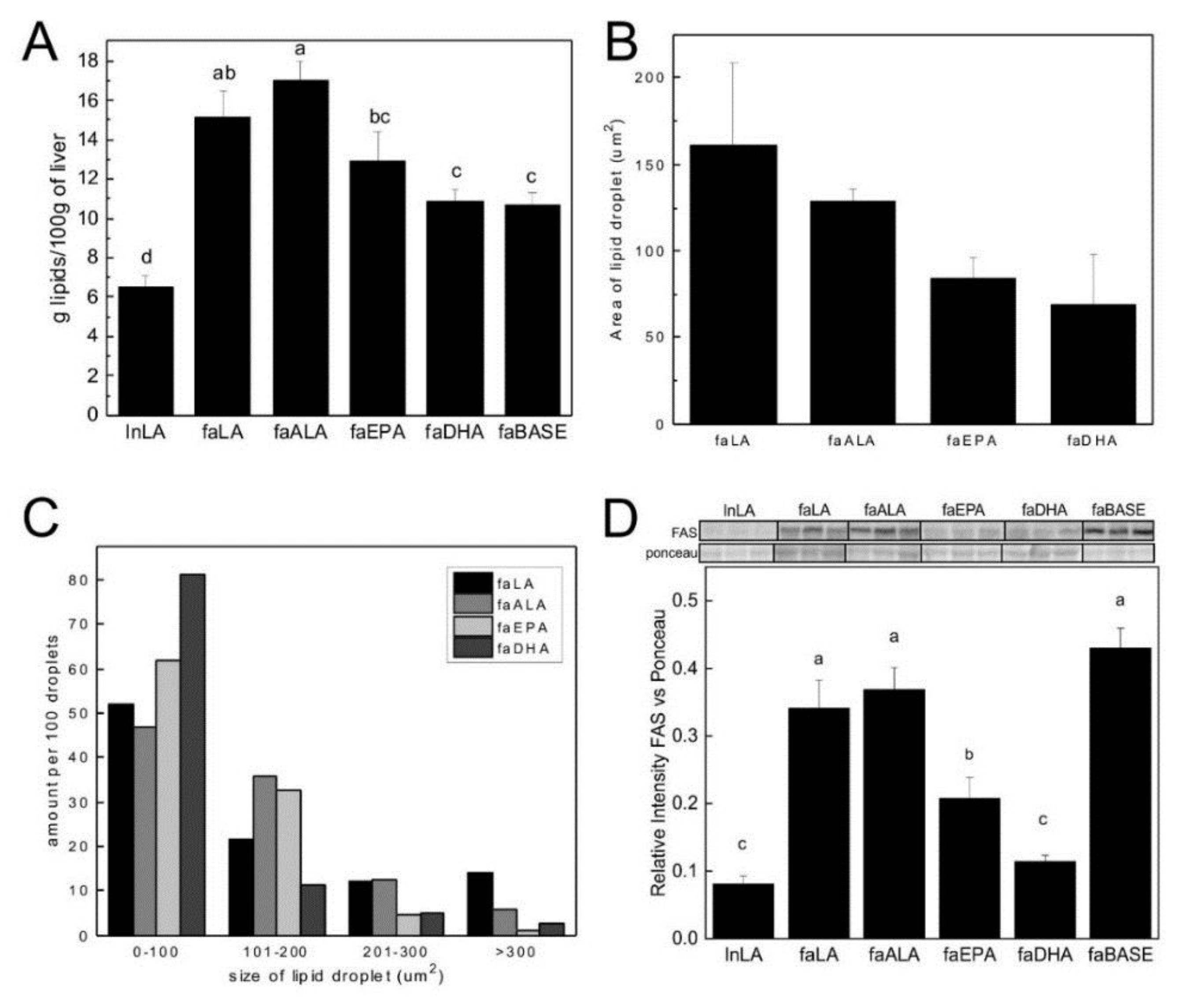
2.4. Hepatic Lipid Droplets
2.5. Western Immunoblotting
2.6. Statistical Methods
3. Results
3.1. Feed Intake and Body Weight
3.2. Hepatic Steatosis
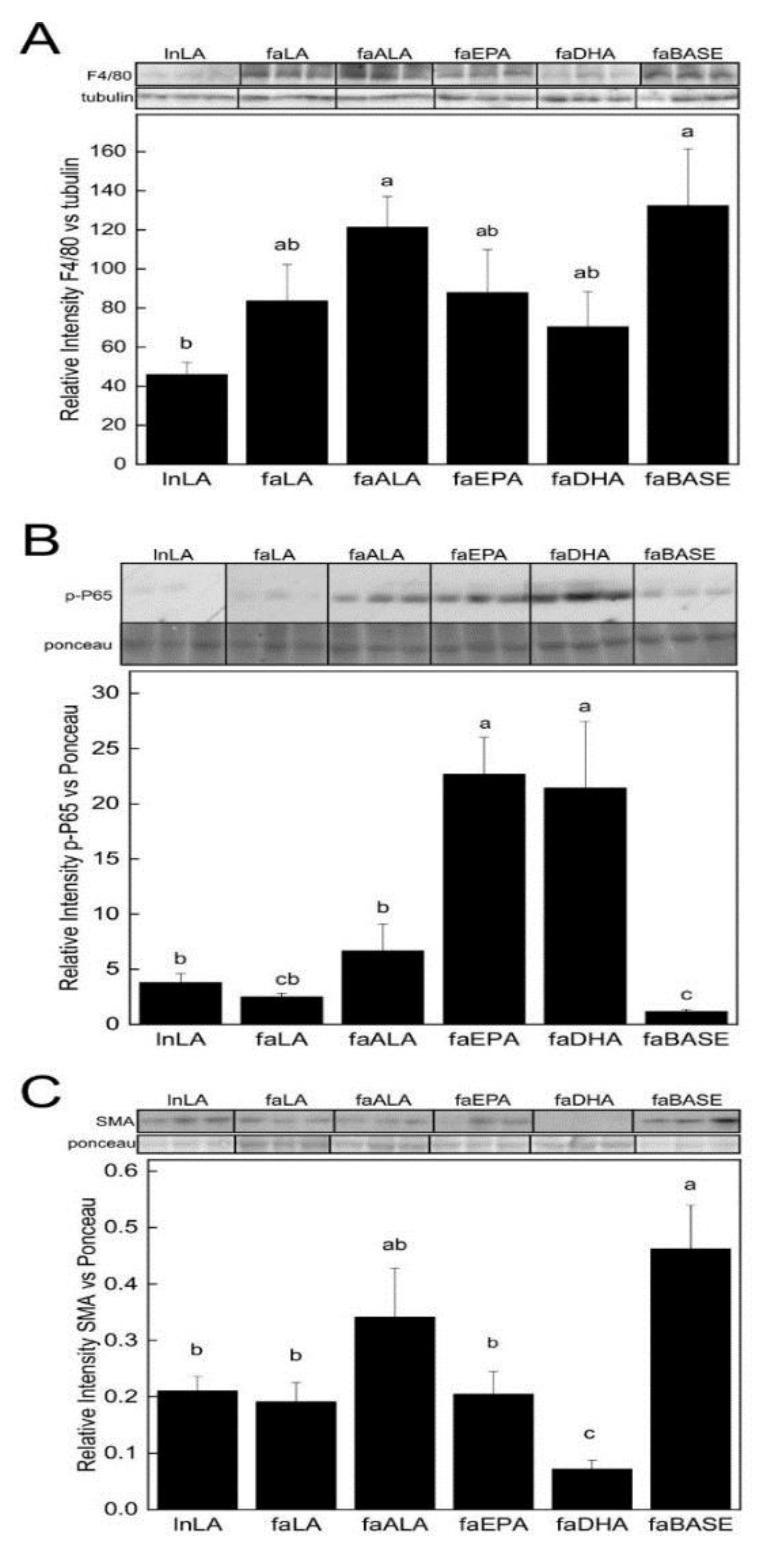
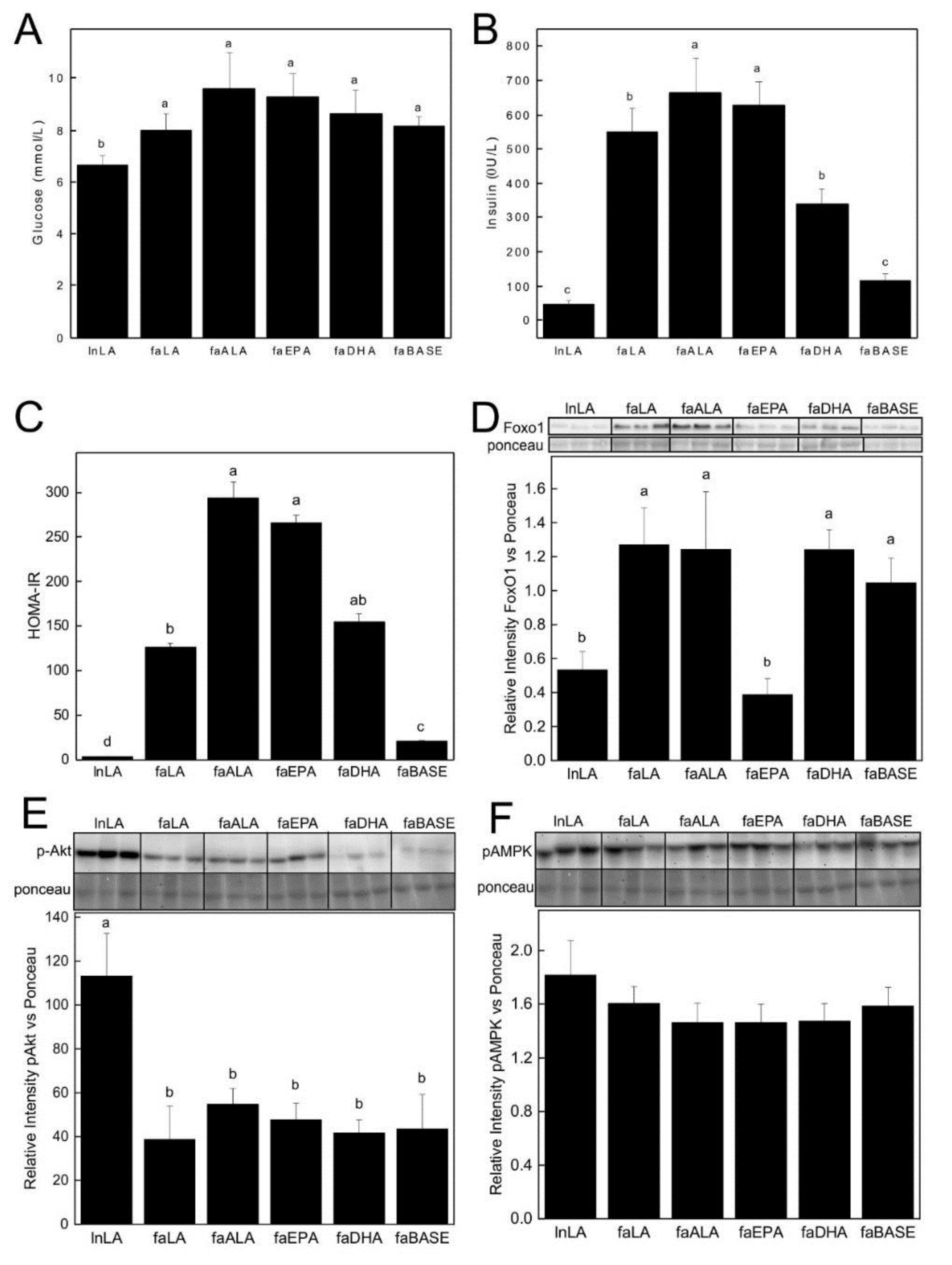
3.3. Biomarkers Related to Hepatic Function
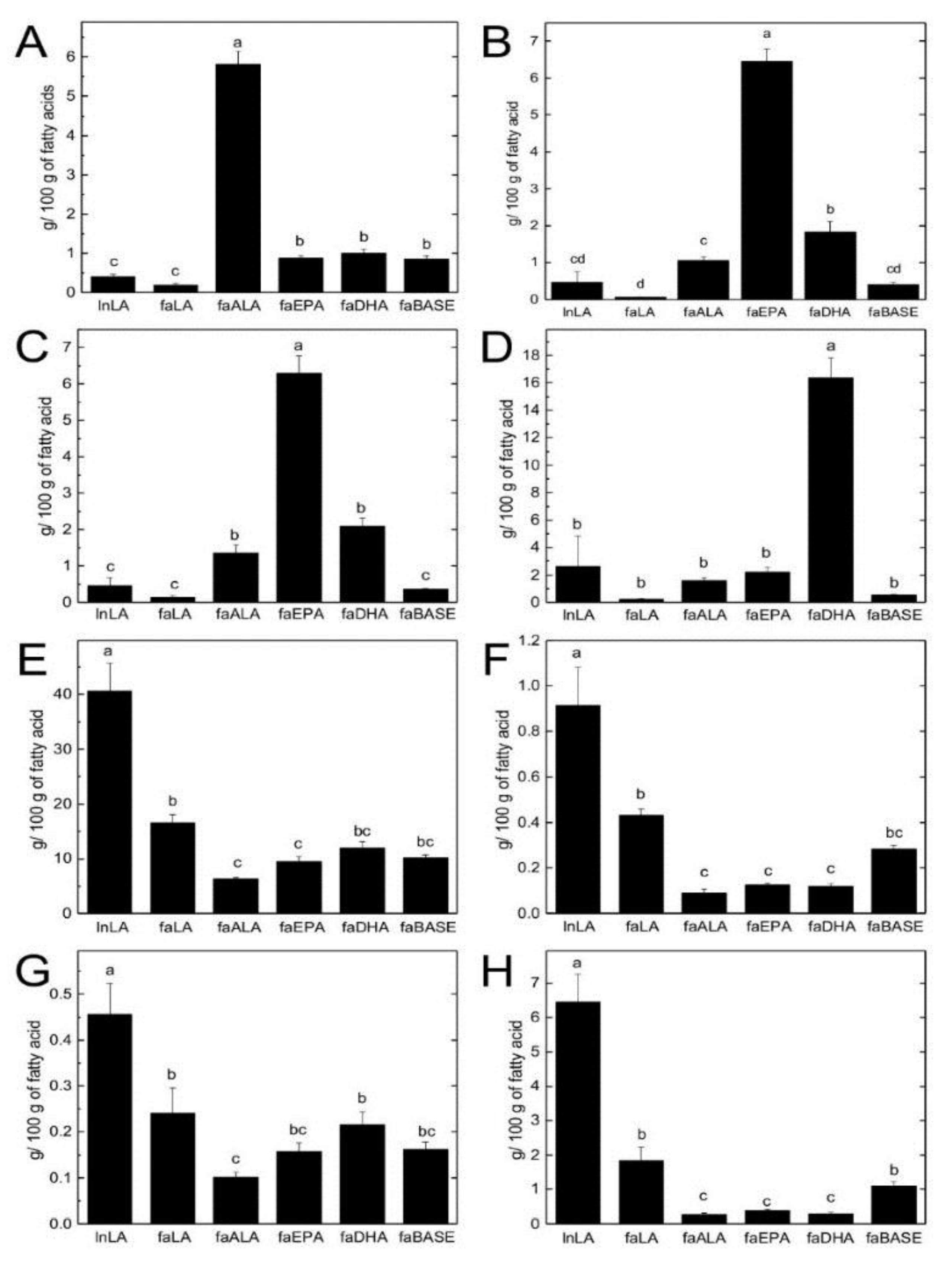
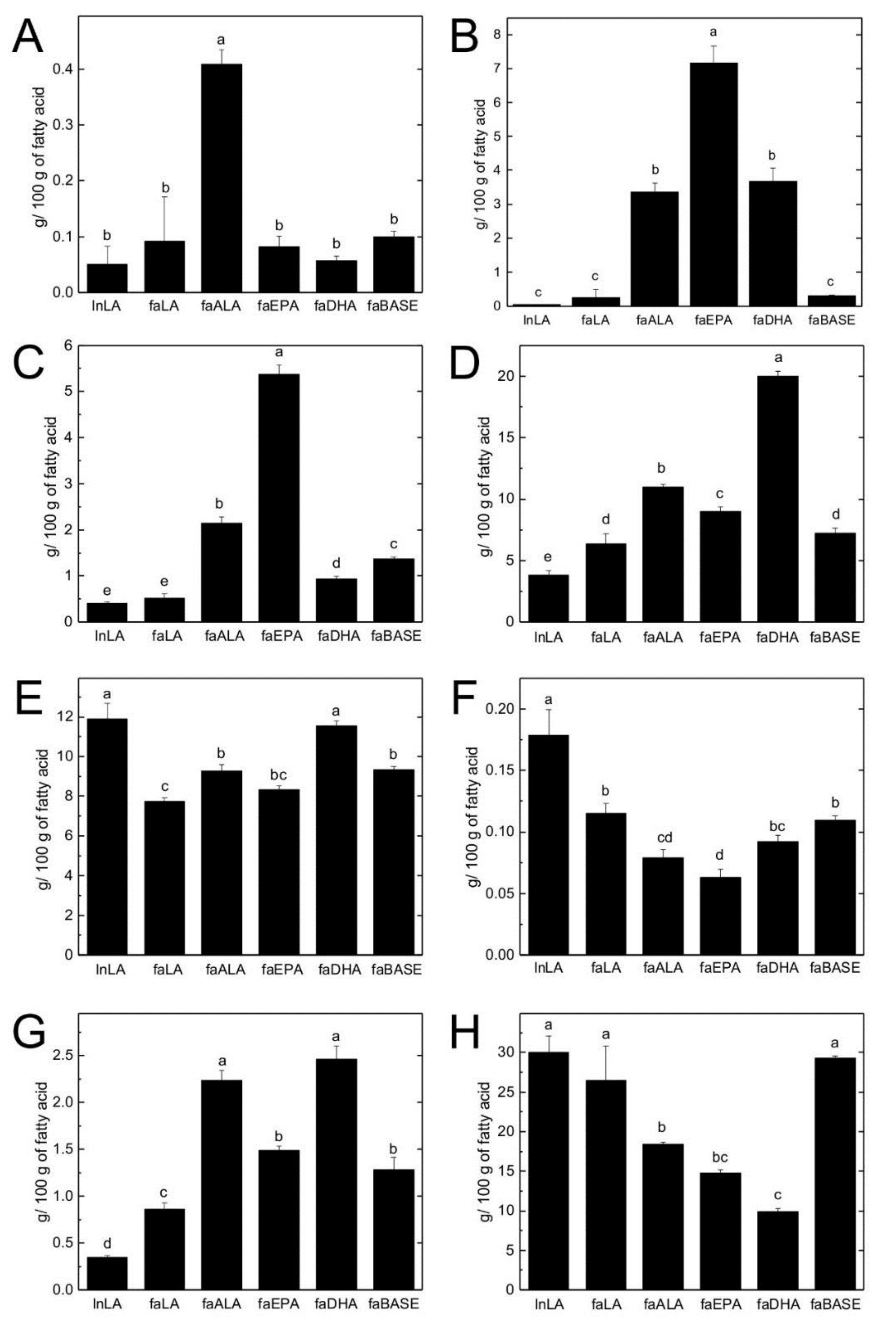
3.4. Hepatic TG and PL Fatty Acid Composition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Than, N.N.; Newsome, P.N. A concise review of non-alcoholic fatty liver disease. Atherosclerosis 2015, 239, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.M.; Koenig, A.B.; Abdelatif, D.; Fazel, Y.; Henry, L.; Wymer, M. Global epidemiology of nonalcoholic fatty liver disease-Meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology (Baltim. Md.) 2016, 64, 73–84. [Google Scholar] [CrossRef]
- Bril, F.; Cusi, K. Management of Nonalcoholic Fatty Liver Disease in Patients With Type 2 Diabetes: A Call to Action. Diabetes care 2017, 40, 419–430. [Google Scholar] [CrossRef]
- Shapiro, H.; Tehilla, M.; Attal-Singer, J.; Bruck, R.; Luzzatti, R.; Singer, P. The therapeutic potential of long-chain omega-3 fatty acids in nonalcoholic fatty liver disease. Clin. Nutr. (Edinb. Scotl.) 2011, 30, 6–19. [Google Scholar] [CrossRef]
- Di Minno, M.N.; Russolillo, A.; Lupoli, R.; Ambrosino, P.; Di Minno, A.; Tarantino, G. Omega-3 fatty acids for the treatment of non-alcoholic fatty liver disease. World J. Gastroenterol. 2012, 18, 5839–5847. [Google Scholar] [CrossRef] [PubMed]
- Scorletti, E.; Byrne, C.D. Omega-3 fatty acids and non-alcoholic fatty liver disease: Evidence of efficacy and mechanism of action. Mol. Aspects Med. 2018, 64, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Deckelbaum, R.J.; Torrejon, C. The omega-3 fatty acid nutritional landscape: Health benefits and sources. J. Nutr. 2012, 142, 587s–591s. [Google Scholar] [CrossRef] [PubMed]
- Depner, C.M.; Philbrick, K.A.; Jump, D.B. Docosahexaenoic acid attenuates hepatic inflammation, oxidative stress, and fibrosis without decreasing hepatosteatosis in a Ldlr(-/-) mouse model of western diet-induced nonalcoholic steatohepatitis. J. Nutr. 2013, 143, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Botelho, P.B.; Mariano Kda, R.; Rogero, M.M.; de Castro, I.A. Effect of Echium oil compared with marine oils on lipid profile and inhibition of hepatic steatosis in LDLr knockout mice. Lipids Health Dis. 2013, 12, 38. [Google Scholar] [CrossRef] [PubMed]
- Suzuki-Kemuriyama, N.; Matsuzaka, T.; Kuba, M.; Ohno, H.; Han, S.I.; Takeuchi, Y.; Isaka, M.; Kobayashi, K.; Iwasaki, H.; Yatoh, S.; et al. Different Effects of Eicosapentaenoic and Docosahexaenoic Acids on Atherogenic High-Fat Diet-Induced Non-Alcoholic Fatty Liver Disease in Mice. PLoS ONE 2016, 11, e0157580. [Google Scholar] [CrossRef]
- Hanke, D.; Zahradka, P.; Mohankumar, S.K.; Clark, J.L.; Taylor, C.G.; Zahradka, P.; Wright, B.; Weighell, W.; Blewett, H.; Baldwin, A.; et al. A diet high in alpha-linolenic acid and monounsaturated fatty acids attenuates hepatic steatosis and alters hepatic phospholipid fatty acid profile in diet-induced obese rats. Prostaglandins Leukot Essent Fat. Acids 2013, 89, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Yuan, F.; Wang, H.; Tian, Y.; He, L.; Shao, Y.; Li, N.; Liu, Z. Perilla Oil Supplementation Ameliorates High-Fat/High-Cholesterol Diet Induced Nonalcoholic Fatty Liver Disease in Rats via Enhanced Fecal Cholesterol and Bile Acid Excretion. BioMed. Res. Int. 2016, 2016, 2384561. [Google Scholar] [CrossRef]
- Han, H.; Qiu, F.; Zhao, H.; Tang, H.; Li, X.; Shi, D. Dietary Flaxseed Oil Prevents Western-Type Diet-Induced Nonalcoholic Fatty Liver Disease in Apolipoprotein-E Knockout Mice. Oxid. Med. Cellular Longev. 2017, 2017, 3256241. [Google Scholar] [CrossRef]
- Poudyal, H.; Panchal, S.K.; Ward, L.C.; Brown, L. Effects of ALA, EPA and DHA in high-carbohydrate, high-fat diet-induced metabolic syndrome in rats. J. Nutr. Biochem. 2013, 24, 1041–1052. [Google Scholar] [CrossRef] [PubMed]
- Stringer, D.M.; Zahradka, P.; Declercq, V.C.; Ryz, N.R.; Diakiw, R.; Burr, L.L.; Xie, X.; Taylor, C.G. Modulation of lipid droplet size and lipid droplet proteins by trans-10,cis-12 conjugated linoleic acid parallels improvements in hepatic steatosis in obese, insulin-resistant rats. Biochim. Et biophysi. Acta 2010, 1801, 1375–1385. [Google Scholar] [CrossRef] [PubMed]
- Wojcik, J.L.; Devassy, J.G.; Wu, Y.; Zahradka, P.; Taylor, C.G.; Aukema, H.M. Protein source in a high-protein diet modulates reductions in insulin resistance and hepatic steatosis in fa/fa Zucker rats. Obesity (Silver Spring Md.) 2016, 24, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C., Jr. AIN-93 purified diets for laboratory rodents: Final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef]
- Sankaran, D.; Lu, J.; Bankovic-Calic, N.; Ogborn, M.R.; Aukema, H.M. Modulation of renal injury in pcy mice by dietary fat containing n-3 fatty acids depends on the level and type of fat. Lipids 2004, 39, 207–214. [Google Scholar] [CrossRef]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef]
- Lepage, L.M.; Giesbrecht, J.A.; Taylor, C.G. Expression of T lymphocyte p56(lck), a zinc-finger signal transduction protein, is elevated by dietary zinc deficiency and diet restriction in mice. J. Nutr. 1999, 129, 620–627. [Google Scholar] [CrossRef]
- Noto, A.; Zahradka, P.; Yurkova, N.; Xie, X.; Nitschmann, E.; Ogborn, M.; Taylor, C.G. Conjugated linoleic acid reduces hepatic steatosis, improves liver function, and favorably modifies lipid metabolism in obese insulin-resistant rats. Lipids 2006, 41, 179–188. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Yeganeh, A.; Zahradka, P.; Taylor, C.G. Trans-10,cis-12 conjugated linoleic acid (t10-c12 CLA) treatment and caloric restriction differentially affect adipocyte cell turnover in obese and lean mice. J. Nutr. Biochem. 2017, 49, 123–132. [Google Scholar] [CrossRef]
- Ishihara, K.; Miyazaki, A.; Nabe, T.; Fushimi, H.; Iriyama, N.; Kanai, S.; Sato, T.; Uozumi, N.; Shimizu, T.; Akiba, S. Group IVA phospholipase A2 participates in the progression of hepatic fibrosis. FASEB J 2012, 26, 4111–4121. [Google Scholar] [CrossRef]
- Singh, S.B.; Kang, L.; Nawrocki, A.R.; Zhou, D.; Wu, M.; Previs, S.; Miller, C.; Liu, H.; Hines, C.D.; Madeira, M.; et al. The Fatty Acid Synthase Inhibitor Platensimycin Improves Insulin Resistance without Inducing Liver Steatosis in Mice and Monkeys. PLoS ONE 2016, 11, e0164133. [Google Scholar] [CrossRef]
- Musa-Veloso, K.; Venditti, C.; Lee, H.Y.; Darch, M.; Floyd, S.; West, S.; Simon, R. Systematic review and meta-analysis of controlled intervention studies on the effectiveness of long-chain omega-3 fatty acids in patients with nonalcoholic fatty liver disease. Nutr. Rev. 2018, 76, 581–602. [Google Scholar] [CrossRef]
- Ikejima, K.; Takei, Y.; Honda, H.; Hirose, M.; Yoshikawa, M.; Zhang, Y.J.; Lang, T.; Fukuda, T.; Yamashina, S.; Kitamura, T.; et al. Leptin receptor-mediated signaling regulates hepatic fibrogenesis and remodeling of extracellular matrix in the rat. Gastroenterology 2002, 122, 1399–1410. [Google Scholar] [CrossRef]
- Saito, T.; Toriniwa, Y.; Ishii, Y.; Uemura, A.; Miyajima, K.; Uno, K.; Shirai, Y.; Nakae, D.; Ohta, T. Hepatic lesions induced by feeding Western diets to Zucker fatty rats, an insulin-resistant model. J. Toxicol. Pathol. 2018, 31, 283–291. [Google Scholar] [CrossRef]
- Giffen, P.S.; Turton, J.; Andrews, C.M.; Barrett, P.; Clarke, C.J.; Fung, K.W.; Munday, M.R.; Roman, I.F.; Smyth, R.; Walshe, K.; et al. Markers of experimental acute inflammation in the Wistar Han rat with particular reference to haptoglobin and C-reactive protein. Archives Toxicol. 2003, 77, 392–402. [Google Scholar] [CrossRef]
- Barrios, V.; Almendro-Delia, M.; Facila, L.; Garcia-Moll, X.; Mazon, P.; Camafort, M.; Cepeda, J.M.; Mediavilla Garcia, J.D.; Pose Reino, A.; Suarez Fernandez, C. Rivaroxaban: Searching the integral vascular protection. Exp. Rev. Clin. Pharmacol. 2018, 11, 719–728. [Google Scholar] [CrossRef]
- Van der Heide, L.P.; Smidt, M.P. Regulation of FoxO activity by CBP/p300-mediated acetylation. Trends Biochem. Sci. 2005, 30, 81–86. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LA Diet | ALA Diet | EPA Diet | DHA Diet | |
|---|---|---|---|---|
| Diet Ingredients (g/kg) 1 | ||||
| Cornstarch | 348 | 348 | 348 | 348 |
| Maltodextrin | 132 | 132 | 132 | 132 |
| Sucrose | 100 | 100 | 100 | 100 |
| Egg white | 213 | 213 | 213 | 213 |
| Cellulose | 50 | 50 | 50 | 50 |
| AIN-93G-MX mineral mix | 35 | 35 | 35 | 35 |
| AIN-93-VX vitamin mix | 10 | 10 | 10 | 10 |
| Choline | 3 | 3 | 3 | 3 |
| Biotin mix 2 | 10 | 10 | 10 | 10 |
| Soybean oil | 0 | 0 | 67 | 67 |
| High linoleic safflower oil 3 | 100 | 0 | 0 | 0 |
| Flaxseed oil 4 | 0 | 87 | 0 | 0 |
| Canola oil 5 | 0 | 10 | 0 | 0 |
| Coconut oil 6 | 0 | 3 | 0 | 0 |
| EPA oil 7 | 0 | 0 | 33 | 0 |
| DHA oil 7 | 0 | 0 | 0 | 33 |
| Fatty Acid Composition (g/100 g lipid) 8 | ||||
| SFA | 10 | 11 | 10 | 10 |
| MUFA | 17 | 19 | 15 | 15 |
| PUFA | 72.3 | 70 | 75 | 75 |
| LA (C18:2n6) | 72 | 18 | 36 | 36 |
| ALA (C18:3n3) | 0.3 | 52 | 6 | 6 |
| EPA (C22:5n3) | 0 | 0 | 32 | 0 |
| DHA (C22:6n3) | 0 | 0 | 0 | 33 |
| Other PUFAS | 0 | 0 | 1 | 0 |
| n6-PUFA:n3-PUFA | 240:1 | 1:3 | 1:1.1 | 1:1.1 |
| lnLA | faLA | faALA | faEPA | faDHA | faBASE | |
|---|---|---|---|---|---|---|
| Body Weight and Feed Intake | ||||||
| Initial body weight (g) | 218 ± 6 c | 323 ± 15 a | 316 ± 11 a | 323 ± 12 a | 297 ± 13 a | 247 ± 4 b |
| Final body weight (g) | 423 ± 7 c | 663 ± 14 a | 635 ± 13 a | 593 ± 15 b | 568 ± 17 b | --- |
| Total weight gain (g) | 211 ± 7 d | 357 ± 19 a | 336 ± 13 ab | 293 ± 14 bc | 282 ± 13 c | ---- |
| Total feed intake (g) | 1219 ± 48 c | 1620 ± 38 a | 1551 ± 27 a | 1392 ± 31 b | 1370 ± 76 b | ---- |
| Feed efficiency ratio (g total feed intake/g weight gain) | 5.80 ± 0.25 a | 4.59 ± 0.16 b | 4.65 ± 0.13 b | 4.80 ± 0.14 b | 4.92 ± 0.2 b | ---- |
| Liver Weight | ||||||
| Liver weight (g) | 13.3 ± 0.7 c | 29.0 ± 1.4 b | 32.7 ± 1.2 a | 28.5 ± 1.2 b | 26.0 ± 0.8 b | 11.0 ± 0.2 d |
| Liver weight (g/100 g bwt) | 2.9 ± 0.1 b | 4.7 ± 0.4 a | 5.2 ± 0.2 a | 4.8 ± 0.2 a | 4.6 ± 0.2 a | 4.5 ± 0.1 a |
| Serum Biochemistry | ||||||
| ALT (U/L) | 47 ± 6 c | 123 ± 11 b | 241 ± 47 a | 99 ± 8 bc | 94 ± 11 bc | 77 ± 3 bc |
| AST (U/L) | 218 ± 17 b | 289 ± 15 b | 430 ± 58 a | 262 ± 20 b | 259 ± 32 b | 207 ± 18 b |
| Haptoglobin (mg/mL) | 1.51 ± 0.2 bc | 2.13 ± 0.2 b | 2.12 ± 0.3 b | 3.08 ± 0.3 a | 3.01 ± 0.2 a | 1.12 ± 0.1 c |
| Total cholesterol (mmol/L) | 2.59 ± 0.10 c | 7.02 ± 0.66 a | 7.29 ± 1.28 a | 3.45 ± 0.23 bc | 2.79 ± 0.28 bc | 4.33 ± 0.23 bc |
| LDL-C (mmol/L) | 0.38 ± 0.03 c | 1.56 ± 0.31 a | 0.81 ± 0.28 b | 0.25 ± 0.04 c | 0.17 ± 0.06 c | 0.22 ± 0.05 c |
| HDL-C (mmol/L) | 2.39 ± 0.06 d | 5.05 ± 0.45 a | 3.91 ± 0.48 b | 2.36 ± 0.13 cd | 1.72 ± 0.23 e | 2.94 ± 0.22 c |
| TG (mmol/L) | 0.87 ± 0.09 d | 4.66 ± 0.51 abc | 5.66 ± 0.53 a | 4.02 ± 0.51 c | 4.76 ± 0.80 bc | 4.81 ± 0.40 ab |
| lnLA | faLA | faALA | faEPA | faDHA | faBASE | |
|---|---|---|---|---|---|---|
| TG Fraction | ||||||
| Total SFA | 27.3 ± 2.5 d | 40.7 ± 1.4 ab | 42.1 ± 0.9 ab | 39.1 ± 0.9 bc | 31.9 ± 5.2 cd | 46.4 ± 0.7 a |
| Total MUFA | 18.4 ± 1.0 d | 37.3 ± 0.8 bc | 41.2 ± 0.6 a | 34.5 ± 1.1 c | 33.0 ± 2.4 c | 39.4 ± 1.1 ab |
| Total PUFA | 54.3 ± 3.3 a | 20.6 ± 2.2 d | 16.8 ± 0.9 de | 26.4 ± 2.1 c | 34.1 ± 3.1 b | 14.1 ± 0.9 e |
| Total n3-PUFA | 3.64 ± 2.5 c | 0.37 ± 0.1 c | 8.77 ± 0.6 b | 8.65 ± 0.8 b | 19.5 ± 1.8 a | 1.75 ± 0.2 c |
| Total n6-PUFA | 50.6 ± 5.7 a | 20.3 ± 2.2 b | 7.99 ± 0.3 d | 17.7 ± 1.3 bc | 14.6 ± 1.4 bcd | 12.4 ± 0.8 cd |
| n6/n3 Ratio | 45.2 ± 11.4 b | 64.2 ± 7.4 a | 0.92 ± 0.03 c | 2.09 ± 0.1 c | 0.75 ± 0.05 c | 7.20 ± 0.3 c |
| Phospholipid Fraction | ||||||
| Total SFA | 43.9 ± 0.1 | 46.0 ± 0.5 | 45.3 ± 0.7 | 45.5 ± 0.6 | 44.4 ± 0.4 | 43.5 ± 0.1 |
| Total MUFA | 5.89 ± 0.1 | 9.05 ± 2.5 | 6.62 ± 0.2 | 7.25 ± 0.2 | 6.07 ± 0.2 | 7.22 ± 1.9 |
| Total PUFA | 50.1 ± 0.1 a | 45.0 ± 2.6 b | 48.0 ± 0.6 ab | 47.2 ± 0.5 ab | 50.0 ± 0.3 a | 49.3 ± 1.6 a |
| Total n3-PUFA | 8.7 ± 0.4 d | 12.8 ± 5 bc | 17.4 ± 3.6 ab | 15.2 ± 0.3 abc | 21.5 ± 0.4 a | 4.85 ± 0.4 cd |
| Total n6-PUFA | 41.4 ± 0.4 a | 32.2 ± 5.6 b | 30.6 ± 3.5 b | 32.0 ± 0.2 b | 28.0 ± 0.4 b | 44.5 ± 1.4 a |
| n6/n3 Ratio | 4.81 ± 0.2 b | 4.36 ± 1.1 b | 2.11 ± 0.4 c | 2.10 ± 0.03 c | 1.30 ± 0.04 c | 9.39 ± 0.6 a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, L.; Zahradka, P.; Cordero-Monroy, L.; Wright, B.; Taylor, C.G. Dietary Docosahexaenoic Acid (DHA) and Eicosapentaenoic Acid (EPA) Operate by Different Mechanisms to Modulate Hepatic Steatosis and Hyperinsulemia in fa/fa Zucker Rats. Nutrients 2019, 11, 917. https://doi.org/10.3390/nu11040917
Hong L, Zahradka P, Cordero-Monroy L, Wright B, Taylor CG. Dietary Docosahexaenoic Acid (DHA) and Eicosapentaenoic Acid (EPA) Operate by Different Mechanisms to Modulate Hepatic Steatosis and Hyperinsulemia in fa/fa Zucker Rats. Nutrients. 2019; 11(4):917. https://doi.org/10.3390/nu11040917
Chicago/Turabian StyleHong, Lena, Peter Zahradka, Luis Cordero-Monroy, Brenda Wright, and Carla G. Taylor. 2019. "Dietary Docosahexaenoic Acid (DHA) and Eicosapentaenoic Acid (EPA) Operate by Different Mechanisms to Modulate Hepatic Steatosis and Hyperinsulemia in fa/fa Zucker Rats" Nutrients 11, no. 4: 917. https://doi.org/10.3390/nu11040917
APA StyleHong, L., Zahradka, P., Cordero-Monroy, L., Wright, B., & Taylor, C. G. (2019). Dietary Docosahexaenoic Acid (DHA) and Eicosapentaenoic Acid (EPA) Operate by Different Mechanisms to Modulate Hepatic Steatosis and Hyperinsulemia in fa/fa Zucker Rats. Nutrients, 11(4), 917. https://doi.org/10.3390/nu11040917