Post Meal Energy Boluses Do Not Increase the Duration of Muscle Protein Synthesis Stimulation in Two Anabolic Resistant Situations

 ,
,  , , and
, , and {kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Glucocorticoid Treatment Study
2.1.1. Animal Housing, Surgery and Ethics Statement
2.1.2. Tracer Infusion Procedures
2.1.3. Samples Analysis
2.1.4. Calculations
2.1.5. Statistics and Analysis of Results
2.2. Aging Study
2.2.1. Animals and Diets
2.2.2. Analytical Procedures
2.2.3. Statistics and Analysis of Results
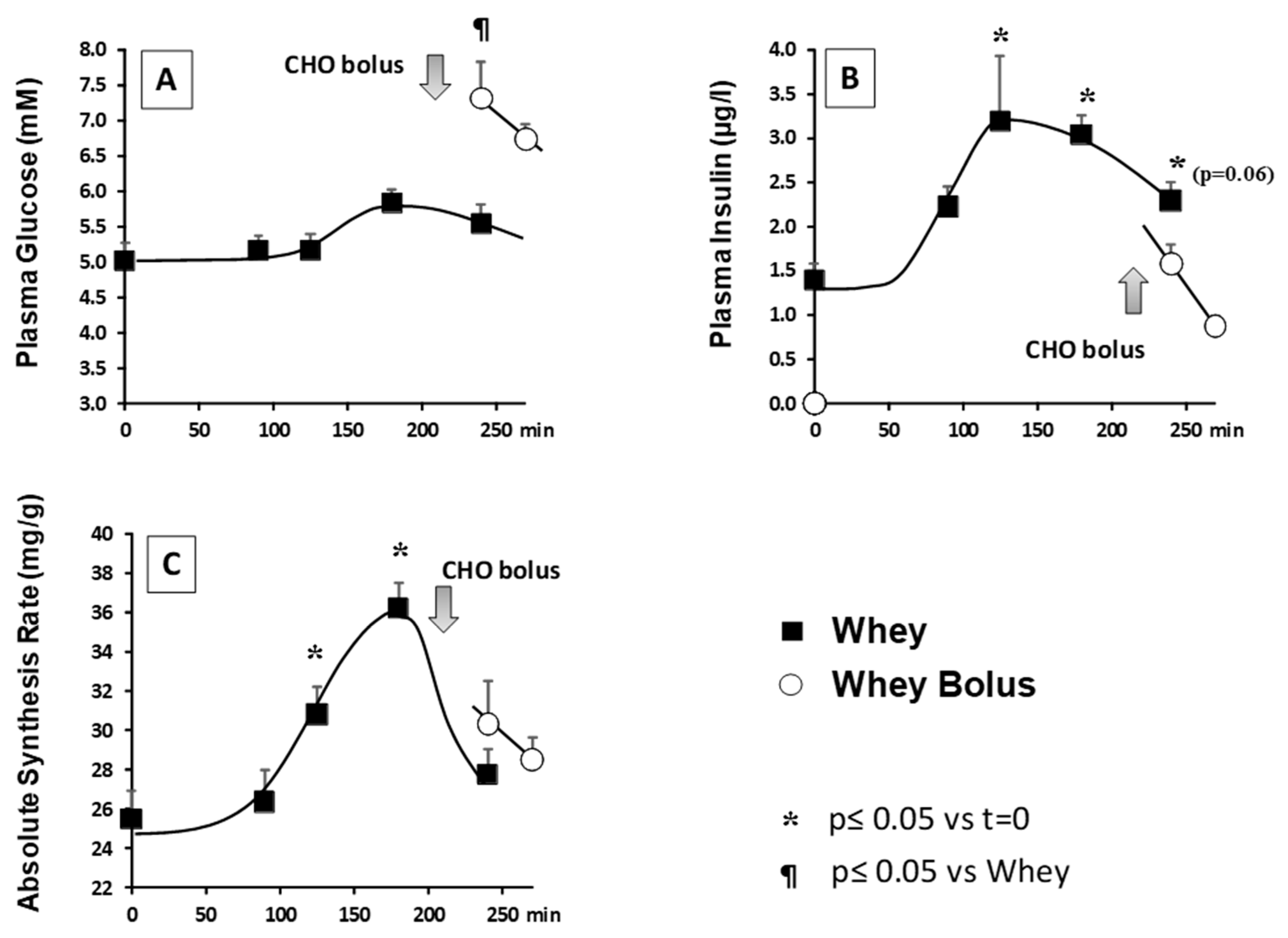
3. Results
3.1. Minipigs Treated with Glucocorticoids
3.1.1. Plasma Glucose, Insulin, and Arterial Amino Acid Concentrations
3.1.2. Muscle Protein Synthesis
3.2. Aging Study
Plasma Glucose and Insulin
3.3. Muscle Protein Synthesis
4. Discussion
Limiting Points of the Study
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dardevet, D.; Remond, D.; Peyron, M.A.; Papet, I.; Savary-Auzeloux, I.; Mosoni, L. Muscle wasting and resistance of muscle anabolism: The “anabolic threshold concept” for adapted nutritional strategies during sarcopenia. Sci. World J. 2012, 2012, 269531. [Google Scholar] [CrossRef]
- Atherton, P.J.; Wilkinson, D.J.; Smith, K. Chapter 9—Feeding Modulation of Amino Acid Utilization: Role of Insulin and Amino Acids in Skeletal Muscle. In The Molecular Nutrition of Amino Acids and Proteins; Dardevet, D., Ed.; Academic Press: Boston, MA, USA, 2016; pp. 109–124. [Google Scholar] [CrossRef]
- Prod’homme, M.; Rieu, I.; Balage, M.; Dardevet, D.; Grizard, J. Insulin and amino acids both strongly participate to the regulation of protein metabolism. Curr. Opin. Clin. Nutr. Metab. Care 2004, 7, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Mosoni, L.V.M.; Serrurier, B.; Prugnaud, J.; Obled, C.; Guezennec, C.Y.; Mirand, P.P. Altered response of protein synthesis to nutritional state and endurance training in old rats. Am. J. Physiol. 1995, 268, E328–E335. [Google Scholar] [CrossRef] [PubMed]
- Cuthbertson, D.; Smith, K.; Babraj, J.; Leese, G.; Waddell, T.; Atherton, P.; Wackerhage, H.; Taylor, P.M.; Rennie, M.J. Anabolic signaling deficits underlie amino acid resistance of wasting, aging muscle. FASEB J. 2004, 19, 422–424. [Google Scholar] [CrossRef]
- Horstman, A.; Olde Damink, S.; Schols, A.; van Loon, L. Is Cancer Cachexia Attributed to Impairments in Basal or Postprandial Muscle Protein Metabolism? Nutrients 2016, 8, 499. [Google Scholar] [CrossRef] [PubMed]
- Rudrappa, S.S.; Wilkinson, D.J.; Greenhaff, P.L.; Smith, K.; Idris, I.; Atherton, P.J. Human Skeletal Muscle Disuse Atrophy: Effects on Muscle Protein Synthesis, Breakdown, and Insulin Resistance—A Qualitative Review. Front. Physiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Magne, H.; Savary-Auzeloux, I.; Migné, C.; Peyron, M.A.; Combaret, L.; Rémond, D.; Dardevet, D. Contrarily to whey and high protein diets, dietary free leucine supplementation cannot reverse the lack of recovery of muscle mass after prolonged immobilization during ageing. J. Physiol. 2012, 590, 2035–2049. [Google Scholar] [CrossRef]
- Jarzaguet, M.; Polakof, S.; David, J.; Migné, C.; Joubrel, G.; Efstathiou, T.; Rémond, D.; Mosoni, L.; Dardevet, D. A meal with mixed soy/whey proteins is as efficient as a whey meal in counteracting the age-related muscle anabolic resistance only if the protein content and leucine levels are increased. Food Funct. 2018, 9, 6526–6534. [Google Scholar] [CrossRef]
- Revel, A.; Jarzaguet, M.; Peyron, M.A.; Papet, I.; Hafnaoui, N.; Migné, C.; Mosoni, L.; Polakof, S.; Savary-Auzeloux, I.; Rémond, D.; et al. At same leucine intake, a whey/plant protein blend is not as effective as whey to initiate a transient postprandial muscle anabolic response during a catabolic state in mini pigs. PLoS ONE 2017, 12, e0186204. [Google Scholar] [CrossRef]
- Rand, W.M.; Pellett, P.L.; Young, V.R. Meta-analysis of nitrogen balance studies for estimating protein requirements in healthy adults. Am. J. Clin. Nutr. 2003, 77, 109–127. [Google Scholar] [CrossRef] [PubMed]
- Dangin, M.; Guillet, C.; Garcia-Rodenas, C.; Gachon, P.; Bouteloup-Demange, C.; Reiffers-Magnani, K.; Fauquant, J.; Ballèvre, O.; Beaufrère, B. The Rate of Protein Digestion affects Protein Gain Differently during Aging in Humans. J. Physiol. 2003, 549, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Magne, H.; Savary-Auzeloux, I.; Rémond, D.; Dardevet, D. Nutritional strategies to counteract muscle atrophy caused by disuse and to improve recovery. Nutr. Res. Rev. 2013, 26, 149–165. [Google Scholar] [CrossRef]
- Ferrando, A.A.; Paddon-Jones, D.; Hays, N.P.; Kortebein, P.; Ronsen, O.; Williams, R.H.; McComb, A.; Symons, T.B.; Wolfe, R.R.; Evans, W. EAA supplementation to increase nitrogen intake improves muscle function during bed rest in the elderly. Clin. Nutr. 2010, 29, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Mosoni, L.; Gatineau, E.; Gatellier, P.; Migné, C.; Savary-Auzeloux, I.; Rémond, D.; Rocher, E.; Dardevet, D. High Whey Protein Intake Delayed the Loss of Lean Body Mass in Healthy Old Rats, whereas Protein Type and Polyphenol/Antioxidant Supplementation Had No Effects. PLoS ONE 2014, 9, e109098. [Google Scholar] [CrossRef]
- Verhoeven, S.; Vanschoonbeek, K.; Verdijk, L.B.; Koopman, R.; Wodzig, W.K.W.H.; Dendale, P.; van Loon, L.J.C. Long-term leucine supplementation does not increase muscle mass or strength in healthy elderly men. Am. J. Clin. Nutr. 2009, 89, 1468–1475. [Google Scholar] [CrossRef]
- Choudry, H.A.; Pan, M.; Karinch, A.M.; Souba, W.W. Branched-Chain Amino Acid-Enriched Nutritional Support in Surgical and Cancer Patients. J. Nutr. 2006, 136, S314–S318. [Google Scholar] [CrossRef]
- Nicastro, H.; Zanchi, N.E.; da Luz, C.R.; de Moraes, W.M.A.M.; Ramona, P.; de Siqueira Filho, M.A.; Chaves, D.F.S.; Medeiros, A.; Brum, P.C.; Dardevet, D.; et al. Effects of leucine supplementation and resistance exercise on dexamethasone-induced muscle atrophy and insulin resistance in rats. Nutrition 2012, 28, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Nicastro, H.; Artioli, G.G.; dos Santos Costa, A.; Solis, M.Y.; da Luz, C.R.; Blachier, F.; Lancha, A.H. An overview of the therapeutic effects of leucine supplementation on skeletal muscle under atrophic conditions. Amino Acids 2011, 40, 287–300. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, M.; Bos, C.; Léonil, J.; Airinei, G.; Luengo, C.; Daré, S.; Benamouzig, R.; Fouillet, H.; Fauquant, J.; Tomé, D.; et al. Compared with casein or total milk protein, digestion of milk soluble proteins is too rapid to sustain the anabolic postprandial amino acid requirement. Am. J. Clin. Nutr. 2006, 84, 1070–1079. [Google Scholar] [CrossRef]
- Atherton, P.J.; Etheridge, T.; Watt, P.W.; Wilkinson, D.; Selby, A.; Rankin, D. Muscle full effect after oral protein: Time-dependent concordance and discordance between human muscle protein synthesis and mTORC1 signaling. Am. J. Clin. Nutr. 2010, 92, 1080–1088. [Google Scholar] [CrossRef]
- Bohé, J.; Low, J.F.A.; Wolfe, R.R.; Rennie, M.J. Latency and duration of stimulation of human muscle protein synthesis during continuous infusion of amino acids. J. Physiol. 2001, 532, 575–579. [Google Scholar] [CrossRef]
- Norton, L.E.; Layman, D.K.; Bunpo, P.; Anthony, T.G.; Brana, D.V.; Garlick, P.J. The Leucine Content of a Complete Meal Directs Peak Activation but Not Duration of Skeletal Muscle Protein Synthesis and Mammalian Target of Rapamycin Signaling in Rats. J. Nutr. 2009, 139, 1103–1109. [Google Scholar] [CrossRef]
- Wilson, G.J.; Layman, D.K.; Moulton, C.J.; Norton, L.E.; Anthony, T.G.; Proud, C.G.; Rupassara, S.I.; Garlick, P.J. Leucine or carbohydrate supplementation reduces AMPK and eEF2 phosphorylation and extends postprandial muscle protein synthesis in rats. Am. J. Physiol. Endocrinol. Metab. 2011, 301, E1236–E1242. [Google Scholar] [CrossRef] [PubMed]
- Rémond, D.; Buffière, C.; Godin, J.P.; Mirand, P.P.; Obled, C.; Papet, I.; Dardevet, D.; Williamson, G.; Breuillé, D.; Faure, M. Intestinal Inflammation Increases Gastrointestinal Threonine Uptake and Mucin Synthesis in Enterally Fed Minipigs. J. Nutr. 2009, 139, 720–726. [Google Scholar] [CrossRef] [PubMed]
- Rieu, I.; Magne, H.; Savary-Auzeloux, I.; Averous, J.; Bos, C.; Peyron, M.A.; Combaret, L.; Dardevet, D. Reduction of low grade inflammation restores blunting of postprandial muscle anabolism and limits sarcopenia in old rats. J. Physiol. 2009, 587, 5483–5492. [Google Scholar] [CrossRef] [PubMed]
- Bruins, M.J.; Soeters, P.B.; Lamers, W.H.; Deutz, N.E. l-Arginine supplementation in pigs decreases liver protein turnover and increases hindquarter protein turnover both during and after endotoxemia. Am. J. Clin. Nutr. 2002, 75, 1031–1044. [Google Scholar] [CrossRef]
- Paddon-Jones, D.; Sheffield-Moore, M.; Zhang, X.J.; Volpi, E.; Wolf, S.E.; Aarsland, A.; Ferrando, A.A.; Wolfe, R.R. Amino acid ingestion improves muscle protein synthesis in the young and elderly. Am. J. Physiol. Endocrinol. Metab. 2004, 286, E321–E328. [Google Scholar] [CrossRef]
- Tourian, A.; Goddard, J.; Puck, T.T. Phenylalanine hydroxylase activity in mammalian cells. J. Cell. Physiol. 1969, 73, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Dardevet, D.; Sornet, C.; Balage, M.; Grizard, J. Stimulation of in vitro rat muscle protein synthesis by leucine decreases with age. J. Nutr. 2000, 130, 2630–2635. [Google Scholar] [CrossRef]
- Kimball, S.R.; Shantz, L.M.; Horetsky, R.L.; Jefferson, L.S. Leucine regulates translation of specific mRNAs in L6 myoblasts through mTOR-mediated changes in availability of eIF4E and phosphorylation of ribosomal protein S6. J. Biol. Chem. 1999, 274, 11647–11652. [Google Scholar] [CrossRef]
- Rieu, I.; Sornet, C.; Grizard, J.; Dardevet, D. Glucocorticoid excess induces a prolonged leucine resistance on muscle protein synthesis in old rats. Exp. Gerontol. 2004, 39, 1315–1321. [Google Scholar] [CrossRef]
- Paddon-Jones, D.; Sheffield-Moore, M.; Creson, D.L.; Sanford, A.P.; Wolf, S.E.; Wolfe, R.R.; Ferrando, A.A. Hypercortisolemia alters muscle protein anabolism following ingestion of essential amino acids. Am. J. Physiol. Endocrinol. Metab. 2003, 284, E946–E953. [Google Scholar] [CrossRef]
- Rieu, I.; Balage, M.; Sornet, C.; Giraudet, C.; Pujos, E.; Grizard, J.; Mosoni, L.; Dardevet, D. Leucine supplementation improves muscle protein synthesis in the elderly men independently of hyperaminoacidemia. J. Physiol. 2006, 575, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Woo, J. Nutritional interventions in sarcopenia: Where do we stand? Curr. Opin. Clin. Nutr. Metab. Care 2018, 21, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Trappe, S.; Creer, A.; Minchev, K.; Slivka, D.; Louis, E.; Luden, N.; Trappe, T. Human soleus single muscle fiber function with exercise or nutrition countermeasures during 60 days of bed rest. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 294, R939–R947. [Google Scholar] [CrossRef] [PubMed]
- Trappe, S.; Creer, A.; Slivka, D.; Minchev, K.; Trappe, T. Single muscle fiber function with concurrent exercise or nutrition countermeasures during 60 days of bed rest in women. J. Appl. Physiol. 2007, 103, 1242–1250. [Google Scholar] [CrossRef] [PubMed]
- Magne, H.; Savary-Auzeloux, I.; Peyron, M.A.; Migne, C.; Combaret, L.; Rémond, D.; Dardevet, D. Which Nutritional Protein Strategy to Recover Muscle Mass after Prolonged Immobilization during Aging? In Proceedings of the 34th Congress of the European Society of Clinical Nutrition and Metabolism (ESPEN), Barcelone, Spain, 8–11 September 2012; pp. 6–7. [Google Scholar]
- Verney, J.; Martin, V.; Ratel, S.; Chavanelle, V.; Bargetto, M.; Etienne, M.; Chaplais, E.; Le Ruyet, P.; Bonhomme, C.; Combaret, L.; et al. Soluble milk proteins improve muscle mass recovery after immobilization-induced muscle atrophy in old rats but do not improve muscle functional property restoration. J. Nutr. Health Aging 2016, 21, 1133–1141. [Google Scholar] [CrossRef] [PubMed]
- Phillips, B.E.; Hill, D.S.; Atherton, P.J. Regulation of muscle protein synthesis in humans. Curr. Opin. Clin. Nutr. Metab. Care 2012, 15, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Dideriksen, K.; Reitelseder, S.; Holm, L. Influence of Amino Acids, Dietary Protein, and Physical Activity on Muscle Mass Development in Humans. Nutrients 2013, 5, 852–876. [Google Scholar] [CrossRef]
- Wilson, G.; Moulton, C.; Garlick, P.; Anthony, T.; Layman, D. Post-Meal Responses of Elongation Factor 2 (eEF2) and Adenosine Monophosphate-Activated Protein Kinase (AMPK) to Leucine and Carbohydrate Supplements for Regulating Protein Synthesis Duration and Energy Homeostasis in Rat Skeletal Muscle. Nutrients 2012, 4, 1723–1739. [Google Scholar] [CrossRef]
- Chang, A.M.; Halter, J.B. Aging and insulin secretion. Am. J. Physiol. Endocrinol. Metab. 2003, 284, E7–E12. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Peng, Y.; Wang, X.; Fan, Y.; Qin, C.; Shi, L.; Tang, Y.; Cao, K.; Li, H.; Long, J.; et al. Mitochondrial Dysfunction Launches Dexamethasone-Induced Skeletal Muscle Atrophy via AMPK/FOXO3 Signaling. Mol. Pharm. 2016, 13, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Triolo, M.; Hood, D.A. Impact of Aging and Exercise on Mitochondrial Quality Control in Skeletal Muscle. Oxid. Med. Cell. Longev. 2017, 2017. [Google Scholar] [CrossRef] [PubMed]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mosoni, L.; Jarzaguet, M.; David, J.; Polakof, S.; Savary-Auzeloux, I.; Rémond, D.; Dardevet, D. Post Meal Energy Boluses Do Not Increase the Duration of Muscle Protein Synthesis Stimulation in Two Anabolic Resistant Situations. Nutrients 2019, 11, 727. https://doi.org/10.3390/nu11040727
Mosoni L, Jarzaguet M, David J, Polakof S, Savary-Auzeloux I, Rémond D, Dardevet D. Post Meal Energy Boluses Do Not Increase the Duration of Muscle Protein Synthesis Stimulation in Two Anabolic Resistant Situations. Nutrients. 2019; 11(4):727. https://doi.org/10.3390/nu11040727
Chicago/Turabian StyleMosoni, Laurent, Marianne Jarzaguet, Jérémie David, Sergio Polakof, Isabelle Savary-Auzeloux, Didier Rémond, and Dominique Dardevet. 2019. "Post Meal Energy Boluses Do Not Increase the Duration of Muscle Protein Synthesis Stimulation in Two Anabolic Resistant Situations" Nutrients 11, no. 4: 727. https://doi.org/10.3390/nu11040727
APA StyleMosoni, L., Jarzaguet, M., David, J., Polakof, S., Savary-Auzeloux, I., Rémond, D., & Dardevet, D. (2019). Post Meal Energy Boluses Do Not Increase the Duration of Muscle Protein Synthesis Stimulation in Two Anabolic Resistant Situations. Nutrients, 11(4), 727. https://doi.org/10.3390/nu11040727