Glycomacropeptide Bioactivity and Health: A Review Highlighting Action Mechanisms and Signaling Pathways



Abstract
1. Introduction
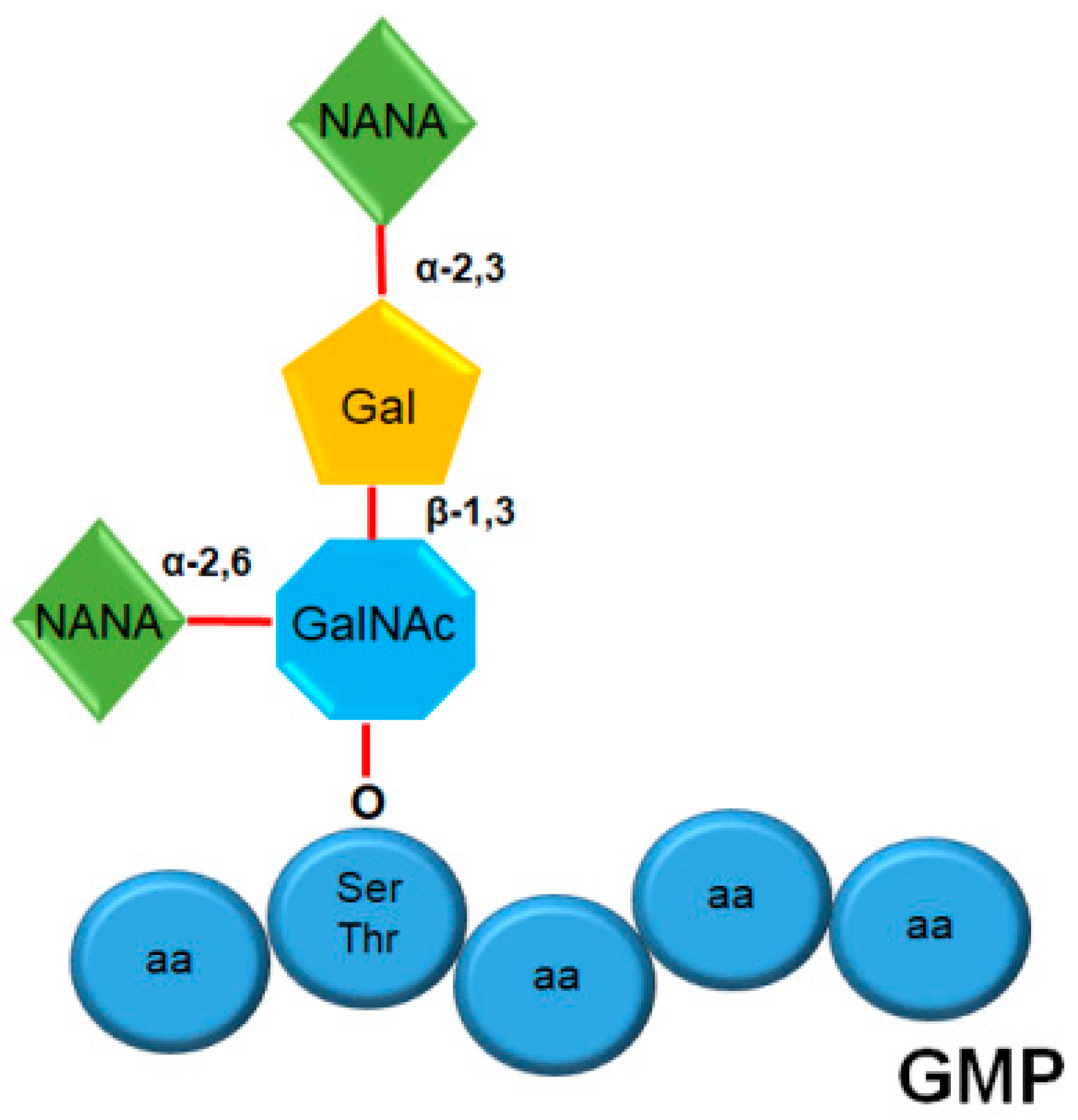
2. Structure and Composition of GMP
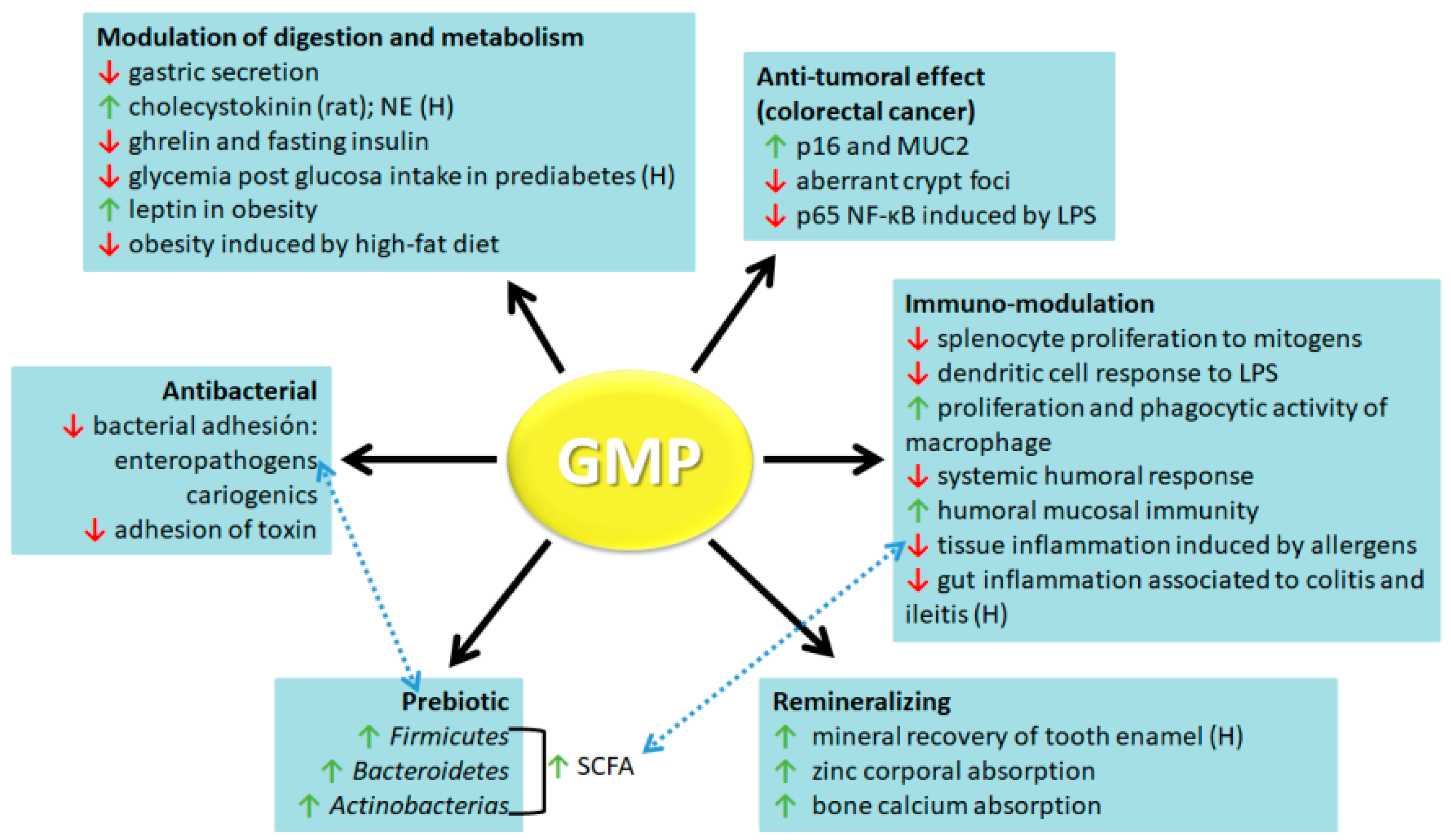
3. GMP Biological Activities
3.1. Antibacterial
3.2. Prebiotic
3.3. Remineralizing
3.4. Modulator of Digestion and Metabolism
3.5. Anti-Tumoral
3.6. Immuno-Modulation
4. Signaling Pathways Regulated or Activated by GMP or GMP-Hydrolyzed
4.1. Signaling Related with Immuno-Modulatory Activity
4.2. Signaling Associated with Metabolism Modulating Activity
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Moughan, P.J.; Rutherfurd, S.M.; Montoya, C.A.; Dave, L.A. Food-derived bioactive peptides—A new paradigm. Nutr. Res. Rev. 2014, 27, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, D.P.; Mohapatra, S.; Misra, S.; Sahu, P.S. Milk derived bioactive peptides and their impact on human health—A review. Saudi J. Biol. Sci. 2016, 23, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Thomä-Worringer, C.; Sørensen, J.; López-Fandiño, R. Health effects and technological features of caseinomacropeptide. Int. Dairy J. 2006, 16, 1324–1333. [Google Scholar] [CrossRef]
- Eigel, W.N.; Butler, J.E.; Emstrom, C.A.; Farrell, H.M., Jr.; Harwalkar, V.R.; Jenness, R.; Whitney, R.M. Nomenclature of proteins of cow’s milk fifth revision. J. Dairy Sci. 1984, 67, 1599–1631. [Google Scholar] [CrossRef]
- Brody, E.P. Biological activities of bovine glycomacropeptide. Br. J. Nutr. 2000, 84, 39–46. [Google Scholar] [CrossRef]
- Farrell, H.M., Jr.; Jimenez-Flores, R.; Bleck, G.T.; Brown, E.M.; Butler, J.E.; Creamer, L.K.; Hicks, C.L.; Hollar, C.M.; Ng-Kwai-Hang, K.F.; Swaisgood, H.E. Nomenclature of the proteins of cows’ milk—Sixth revision. J. Dairy Sci. 2004, 87, 1641–1674. [Google Scholar] [CrossRef]
- Yvon, M.; Beucher, S.; Guilloteau, P.; Le Huerou-Luron, I.; Corring, T. Effects of caseinomacropeptide (CMP) on digestion regulation. Reprod. Nutr. Dev. 1994, 34, 527–537. [Google Scholar] [CrossRef]
- Mercier, J.C.; Brignon, G.; Ribadeau-Dumas, B. Structure primaire de la caseine κ-bovine B Sequence complete. Eur. J. Biochem. 1973, 35, 222–235. [Google Scholar] [CrossRef]
- Ney, D.M.; Stroup, B.M.; Clayton, M.K.; Murali, S.G.; Rice, G.M.; Rohr, F.; Levy, H.L. Glycomacropeptide for nutritional management of phenylketonuria: A randomized, controlled, crossover trial. Am. J. Clin. Nutr. 2016, 104, 334–345. [Google Scholar] [CrossRef]
- Van Spronsen, F.J.; van Wegberg, A.M.; Ahring, K.; Bélanger-Quintana, A.; Blau, N.; Bosch, A.M.; Burlina, A.; Campistol, J.; Feillet, F.; Giżewska, M.; et al. Key European guidelines for the diagnosis and management of patients with phenylketonuria. Lancet Diabetes Endocrinol. 2017, 5, 743–756. [Google Scholar] [CrossRef]
- Abd El-Salama, M.H.; El-Shibinya, S.; Buchheimb, W. Characteristics and potential uses of the casein macropeptide. J. Dairy Sci. 1996, 6, 327–341. [Google Scholar] [CrossRef]
- Tomczak, L.; Lazzari, E.; Bibleski, I. Isolation and purification of milk whey glycomacropeptide. Boletim CEPPA 2007, 25, 121–132. [Google Scholar] [CrossRef]
- Neelima; Sharma, R.; Rajput, Y.S.; Mann, B. Chemical and functional properties of glycomacropeptide (GMP) and its role in the detection of cheese whey adulteration in milk: A review. Dairy Sci. Technol. 2013, 93, 21–43. [Google Scholar] [CrossRef] [PubMed]
- Kreuß, M.; Strixner, T.; Kulozik, U. The effect of glycosylation on the interfacial properties of bovine caseinomacropeptide. Food Hydrocoll. 2009, 23, 1818–1826. [Google Scholar] [CrossRef]
- Minikiewicz, P.; Slangen, C.J.; Lagerwerf, F.M.; Haverkamp, J.; Rollema, H.S.; Visser, S. Reversed-phase high-performance liquid chromatographic separation of bovine kappa-casein macropeptide and characterization of isolated fractions. J. Chromatogr. A 1996, 743, 123–135. [Google Scholar] [CrossRef]
- Huang, L.J.; Lin, J.H.; Tsai, J.H.; Chu, Y.Y.; Chen, Y.W.; Chen, S.L.; Chen, S.H. Identification of protein O-glycosylation site and corresponding glycans using liquid chromatography-tandem mass spectrometry via mapping accurate mass and retention time shift. J. Chromatogr. A 2014, 1371, 113–145. [Google Scholar] [CrossRef]
- Pisano, A.; Packe, N.H.; Redmond, J.W.; Williams, K.L.; Gooley, A.A. Characterization of O-linked glycosylation motifs in the glycopeptide domain of bovine kappa-casein. Glycobiology 1994, 4, 837–844. [Google Scholar] [CrossRef]
- Saito, T.; Itoh, T. Variations and distributions of O-glycosidically linked sugar chains in bovine kappa-casein. J. Dairy Sci. 1992, 75, 1768–1774. [Google Scholar] [CrossRef]
- Isoda, H.; Kawasaki, Y.; Tanimoto, M.; Dosako, S.; Idota, T. Use of Compounds Containing or Binding Sialic Acid to Neutralize Bacterial Toxins. European Patent EP0385118A2, 5 September 1990. [Google Scholar]
- Kawasaki, Y.; Isoda, H.; Tanioto, M.; Dosako, S.; Idota, T.; Ahiko, K. Inhibition by lactoferrin and κ-casein glycomacropetide of binding of cholera toxin to its receptor. Biosci. Biotechnol. Biochem. 1992, 56, 195–198. [Google Scholar] [CrossRef]
- Nakajima, K.; Tamura, N.; Kobayashi-Hattori, K.; Yoshida, T.; Hara-Kudo, Y.; Ikedo, M.; Sugita-Konishi, Y.; Hattori, M. Prevention of intestinal infection by glycomacropeptide. Biosci. Biotechnol. Biochem. 2005, 69, 2294–2301. [Google Scholar] [CrossRef]
- Rhoades, A.R.; Gibson, G.R.; Formentín, K.; Beer, M.; Greenberg, N.; Rastall, R.A. Caseinoglycomacropeptide Inhibits Adhesion of Pathogenic Escherichia coli Strains to Human Cells in Culture. J. Dairy Sci. 2005, 88, 3455–3459. [Google Scholar] [CrossRef]
- Brück, W.M.; Kelleher, S.L.; Gibson, G.R.; Graverholt, G.; Lönnerdal, B.L. The effects of a-lactalbumin and glycomacropeptide on the association of CaCo-2 cells by enteropathogenic Escherichia coli, Salmonella typhimurium and Shigella flexneri. FEMS Microbiol. Lett. 2006, 259, 158–162. [Google Scholar] [CrossRef]
- Hermes, R.G.; Molist, F.; Pérez, J.F.; Gómez de Segura, A.; Ywasaki, M.; Davin, R.; Nofrarías, M.; Korhonen, T.K.; Virkola, R.; Martín-Orúe, S.M. Casein glycomacropeptide in the diet may reduce Escherichia coli attachment to the intestinal mucosa and increase the intestinal lactobacilli of early weaned piglets after an enterotoxigenic E. coli K88 challenge. Br. J. Nutr. 2013, 109, 1001–1012. [Google Scholar] [CrossRef]
- Feeney, S.; Tyan, J.T.; Kilcoyne, M.; Joshi, L.; Hickey, R. Glycomacropeptide reduces intestinal epithelial cell barrier dysfunction and adhesion of entero-hemorrhagic and entero-pathogenic Escherichia coli in vitro. Food 2017, 6, 93. [Google Scholar] [CrossRef]
- Rong, Y.; Lu, Z.; Zhang, H.; Zhan, L.; Song, D.; Wang, Y. Effects of casein glycomacropeptide supplementation on growth performance, intestinal morphology, intestinal barrier permeability and inflammatory responses in Escherichia coli K88 challenged piglets. Anim. Nutr. 2015, 1, 54–59. [Google Scholar] [CrossRef]
- Neeser, J.R. Dental Anti-Plaque and Anti-Caries Agent. U.S. Patent US4992420A, 12 February 1991. [Google Scholar]
- Schüpbach, P.; Neeser, J.R.; Golliard, M.; Rouvet, M.; Guggenheim, B. Incorporation of caseinoglycomacropeptide and caseinophosphopeptide into the salivary pellicle inhibits adherence of mutans streptococci. J. Dent. Res. 1996, 75, 1779–1788. [Google Scholar] [CrossRef]
- Azuma, N.; Yamauchi, K.; Mitsuoka, T. Bifidus growth-promoting activity of a glycomacropeptide derived from human K-casein. Agric. Biol. Chem. 1984, 48, 2159–2162. [Google Scholar] [CrossRef]
- Idota, T.; Kawakami, H.; Nakajima, I. Growth-promoting effects of N-actylneuraminic acid-containing substances on bifidobacteria. Biosci. Biotechnol. Biochem. 1994, 58, 1720–1722. [Google Scholar] [CrossRef]
- Janer, C.; Peláez, C.; Requena, T. Caseinomacropeptide and whey protein concentrate enhance Bifidobacterium lactis growth in milk. Food Chem. 2004, 86, 263–267. [Google Scholar] [CrossRef]
- Robitaille, G. Growth-promoting effects of caseinomacropeptide from cow and goat milk on probiotics. J. Dairy Res. 2013, 80, 58–63. [Google Scholar] [CrossRef]
- Ntemeri, A.; Chonchúir, F.N.; O’Callagan, T.F.; Stanton, C.; Ross, R.P.; O’Toole, P.W. Glycomacropeptide sustains microbiota diversity and promotes specific taxa in an artificial colon model of elderly gut microbiota. J. Agric. Food Chem. 2017, 65, 1836–1846. [Google Scholar] [CrossRef]
- O’Riordan, N.; O’Callagan, J.; Buttò, L.F.; Kilcoyne, M.; Joshi, L.; Hickey, R.M. Bovine glycomacropeptide promotes the growth of Bifidobacterium longum ssp. infntis anda modulates its gene expression. J. Dairy Sci. 2018, 101, 6730–6741. [Google Scholar] [CrossRef]
- Chen, Q.; Cao, J.; Jia, Y.; Liu, X.; Yan, Y.; Pang, G. Modulation of mice fecal microbiota by administration of casein glycomacropeptide. Microbiol. Res. 2012, 3, e3. [Google Scholar] [CrossRef]
- Sawin, E.A.; De Wolfe, T.J.; Aktas, B.; Stroup, B.M.; Murali, S.G.; Steele, J.L.; Ney, D.M. Glycomacropeptide is a prebiotic that reduces Desulfovibrio bacteria, increases cecal short-chain fatty acids, and is anti-inflammatory in mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 309, G590–G601. [Google Scholar] [CrossRef]
- Zhang, Y.P.; Gaffar, A. Fluoride Free Dental Remineralization. U.S. Patent US6207138B1, 27 March 2001. [Google Scholar] [CrossRef]
- Kelleher, S.L.; Chatterton, D.; Nielsen, K.; Lönnerdal, B. Glycomacropeptide and α-lactalbumin supplementation of infant formula affects growth and nutritional status in infant rhesus monkeys. Am. J. Clin. Nutr. 2003, 7, 1261–1268. [Google Scholar] [CrossRef]
- Solverson, P.; Murali, S.G.; Litscher, S.J.; Blank, R.D.; Ney, D.M. Low bone strength is a manifestation of phenylketonuria in mice and is attenuated by a glycomacropeptide diet. PLoS ONE 2012, 7, e45165. [Google Scholar] [CrossRef]
- Burns, P.; Binetti, A.; Torti, P.; Kulozik, U.; Forzani, L.; Renzulli, E.; Vinderola, G.; Reinheimer, J. Administration of caseinomacropeptide-enriched extract to mice enhances the calcium content of femur in a low-calcium diet. Int. Dairy J. 2015, 44, 15–20. [Google Scholar] [CrossRef]
- Sawin, E.A.; Stroup, B.M.; Murali, S.G.; O’Neill, L.M.; Ntambi, J.M.; Ney, D.N. Differential effects of dietary fat content and protein source on bone phenotype and fatty acid oxidation in female C57Bl/6 mice. PLoS ONE 2016, 11, e0163234. [Google Scholar] [CrossRef]
- Shlygin, G.K.; Vasilevskaya, L.S.; Chernikov, M.P.; Nikol’skaya, G.V. Antigastrin action of glycomaropeptide. Byulleten Eksperimental’noi Biologii i. Meditsiny 1971, 72, 9–13. [Google Scholar]
- Vasilevskaia, L.S.; Stan, E.I.; Chernikov, M.P.; Shlygin, G.K. Inhibiting action of glycomacropeptide on stomach secretion induced by various humoral stimulants. Vopr. Pitan. 1977, 4, 21–24. [Google Scholar]
- Stan, E.I.; Groĭsman, S.D.; Krasil’shchikov, K.B.; Chernikov, M.P. Effect of kappa-casein glycomacropeptide on gastrointestinal motility in dogs. Biulleten’eksperimental’noi Biologii i Meditsiny 1983, 96, 10–12. [Google Scholar]
- Stan, E.; Chernikov, M.P. Physiological activity of kappa-casein glycomacropeptide. Vopr. Med. Khim. 1979, 25, 348–352. [Google Scholar]
- Chernikov, M.P.; Stan, E.I.; Vasilevskaia, L.S.; Shlygin, G.K. Heterogeneity of the kappa-casein glycomacropeptide and physiological activity of its fractions. Vopr. Pitan. 1979, 5, 22–26. [Google Scholar]
- Beucher, S.; Levenez, F.; Yvon, M.; Corring, T. Effects of gastric digestive products from casein on CCK release by intestinal cells in rat. J. Nutr. Biochem. 1994, 5, 578–584. [Google Scholar] [CrossRef]
- Pedersen, N.L.; Nagain-Domaine, C.; Mahé, S.; Chariot, J.; Rozé, C.; Tomé, D. Caseinomacropeptide specifically stimulates exocrine pancreatic secretion in the anesthetized rat. Peptides 2000, 21, 1527–1535. [Google Scholar] [CrossRef]
- Gustafson, D.R.; McMahon, D.J.; Morey, J.; Nan, R. Appetite is not influenced by a unique milk peptide: Caseinomacropeptide (CMP). Appetite 2001, 36, 157–163. [Google Scholar] [CrossRef]
- Burton-Freeman, B.M. Glycomacropeptide (GMP) is not critical to whey-induced satiety, but may have a unique role in energy intake regulation through cholecystokinin (CCK). Physiol. Behav. 2008, 93, 379–387. [Google Scholar] [CrossRef]
- Keogh, J.B.; Clifton, P. The effect of meal replacements high in glycomacropeptide on weight loss and markers of cardiovascular disease risk. Am. J. Clin. Nutr. 2008, 87, 1602–1605. [Google Scholar] [CrossRef]
- Keogh, J.B.; Wooton, B.W.; Taylor, C.M.; Janakievski, F.; Desilva, K.; Clifton, P.M. Effect of glycomacropeptide fractions on cholecystokinin and food intake. Br. J. Nutr. 2010, 104, 286–290. [Google Scholar] [CrossRef]
- MacLeod, E.L.; Clayton, M.K.; van Clacar, S.C.; Ney, D.M. Breakfast with glycomacropeptide compared with amino acids suppresses plasma ghrelin levels in individuals with phenylketonuria. Mol. Genet. Metab. 2010, 100, 303–308. [Google Scholar] [CrossRef]
- Royle, P.J.; McIntosh, G.H.; Clifton, P.M. Whey protein isolate and glycomacropeptide decrease weight gain and alter body composition in male Wistar rats. Br. J. Nutr. 2008, 100, 88–93. [Google Scholar] [CrossRef]
- Hoefle, A.S.; Bangert, A.M.; Rist, M.J.; Gedrich, K.; Lee, Y.M.; Skurk, T.; Danier, J.; Schwarzenbolz, U.; Daniel, H. Postprandial metabolic responses to ingestion of bovine glycomacropeptide compared to a whey protein isolate in prediabetic volunteers. Eur. J. Nutr. 2018. [Google Scholar] [CrossRef]
- Xu, S.P.; Mao, X.Y.; Ren, F.Z.; Che, H.L. Attenuating effect of casein glycomacropeptide on proliferation, differentiation, and lipid accumulation of in vitro Sprague-Dawley rat preadipocytes. J. Dairy Sci. 2011, 94, 676–683. [Google Scholar] [CrossRef]
- Xu, S.P.; Mao, X.Y.; Cheng, X.; Chen, B. Ameliorating effects of casein glycomacropeptide on obesity induced by high-fat diet in male Sprague-Dawley rats. Food Chem. Toxicol. 2013, 56, 1–7. [Google Scholar] [CrossRef]
- Chen, Q.; Liang, Y.; Zhu, C.; Yan, Y.; Pang, G. Effects of casein glycomacropeptide on the early development of primary colorectal cancer in rats. Food Sci. Hum. Wellness 2013, 2, 113–118. [Google Scholar] [CrossRef]
- Gong, J.; Chen, Q.; Yan, Y.; Pang, G. Effect of casein glycomacropeptide on subunit p65 of nuclear transcription factor-B in lipopolysaccharide-stimulated human colorectal tumor HT-29 cells. Food Sci. Hum. Wellness 2014, 3, 51–55. [Google Scholar] [CrossRef]
- Otani, H.; Monnai, M.; Hosono, A. Bovine k-casein as inhibitor of the proliferation of mouse splenocytes induced by lipopolysaccharide stimulation. Milchwissenschaft 1992, 47, 512–515. [Google Scholar]
- Otani, H.; Hata, I. Inhibition of proliferative responses of mouse spleen lymphocytes and rabbit Peyer’s patch cells by bovine milk caseins and their digests. J. Dairy Res. 1995, 62, 339–348. [Google Scholar] [CrossRef]
- Otani, H.; Monnai, M. Inhibition of proliferative responses of mouse spleen lymphocytes by bovine milk k-casein digests. Food Agric. Immunol. 1993, 5, 219–229. [Google Scholar] [CrossRef]
- Otani, H.; Monnai, M.; Kawasaki, Y.; Kawakami, H.; Tanimoto, M. Inhibition of mitogen-induced proliferative responses of lymphocytes by bovine k-caseinoglycopeptides having different carbohydrate chains. J. Dairy Res. 1995, 62, 349–357. [Google Scholar] [CrossRef]
- Mikkelsen, T.L.; Bakman, S.; Sørensen, E.S.; Barkholt, V.; Frøkiaer, H. Sialic acid-containing milk proteins show differential immunomodulatory activities independent of sialic acid. J. Agric. Food Chem. 2005, 53, 7673–7680. [Google Scholar] [CrossRef]
- Otani, H.; Monnai, M. Induction of an interleukin-1 receptor antagonist-like component produced from mouse spleen cells by bovine kappa-caseinoglycopeptide. Biosci. Biotechnol. Biochem. 1995, 59, 1166–1168. [Google Scholar] [CrossRef]
- Monnai, M.; Otani, H. Effect of bovine k-caseinoglycopeptide on secretion of interleukin-1 family cytokines by P388D1 cells, a line derived from mouse monocyte/macrophage. Milchwissenschaft 1997, 52, 192–196. [Google Scholar] [CrossRef]
- Otani, H.; Horimoto, Y.; Monnai, M. Suppression of interleukin-2 receptor expression on mouse CD4+T Cells by bovine k-caseinoglycopeptide. Biosci. Biotechnol. Biochem. 1996, 60, 1017–1019. [Google Scholar] [CrossRef]
- Requena, P.; González, R.; López-Posadas, R.; Abadía-Molina, A.; Suárez, M.D.; Zarzuelo A de Medina, F.S.; Martínez-Augustin, O. The intestinal antiinflammatory agent glycomacropeptide has immunomodulatory actions on rat splenocytes. Biochem. Pharmacol. 2010, 79, 1797–1804. [Google Scholar] [CrossRef]
- Monnai, M.; Horimoto, Y.; Otani, H. Immunomodificatory effect of dietary bovine k-caseinoglycopeptide on serum antibody levels and proliferative responses of lymphocytes in mice. Milchwissenschaft 1998, 53, 129–132. [Google Scholar]
- Jiménez, M.; Chávez, N.A.; Salinas, E. Pretreatment with glycomacropeptide reduces allergen sensitization, alleviates immediate cutaneous hypersensitivity and protects from anaphylaxis. Clin. Exp. Immunol. 2012, 170, 18–27. [Google Scholar] [CrossRef]
- Li, E.W.; Mine, Y. Immunoenhancing effects of bovine glycomacropeptide and its derivatives on the proliferative response and phagocytic activities of human macrophagelike cells, U937. J. Agric. Food Chem. 2004, 52, 2704–2708. [Google Scholar] [CrossRef]
- Rusu, D.; Drouin, R.; Pouliot, Y.; Gauthier, S.; Poubelle, P.E. A bovine whey protein extract can enhance innate immunity by priming normal human blood neutrophils. J. Nutr. 2009, 139, 386–393. [Google Scholar] [CrossRef]
- Yun, S.S.; Sugita-Konishi, Y.; Kumagai, S.; Yamauchi, K. Glycomacropeptide from cheese whey protein concentrate enhances IgA production by lipopolysaccharide-stimulated spleen cells. Anim. Sci. Technol (Jpn.) 1996, 67, 458–462. [Google Scholar] [CrossRef]
- Ye, L.; Chen, Q.; Li, W.; Yan, Y.; Zhao, P.; Pang, G.; Hu, Z. Effect of Casein Glycomacropeptide on Phagocytic Cells and Intestinal Mucosa Immune Cells in Mice. Food Sci. 2014, 35, 234–240. [Google Scholar]
- Cui, Y.; Zhu, C.; Ming, Z.; Cao, J.; Yan, Y.; Zhao, P.; Pang, G.; Deng, Z.; Yao, Y.; Chen, Q. Molecular mechanisms by which casein glycomacropeptide maintains internal homeostasis in mice with experimental ulcerative colitis. PLoS ONE 2017, 12, e0181075. [Google Scholar] [CrossRef]
- Daddaoua, A.; Puerta, V.; Zarzuelo, A.; Suárez, M.D.; Sánchez de Medina, F.; Martínez-Augustin, O. Bovine glycomacropeptide is anti-inflammatory in rats with hapten-induced colitis. J. Nutr. 2005, 135, 1164–1170. [Google Scholar] [CrossRef]
- Requena, P.; Daddaoua, A.; Martínez-Plata, E.; González, M.; Zarzuelo, A.; Suárez, M.D.; Sánchez de Medina, F.; Martínez-Augustin, O. Bovine glycomacropeptide ameliorates experimental rat ileitis by mechanisms involving downregulation of interleukin 17. Br. J. Pharmacol. 2008, 154, 825–832. [Google Scholar] [CrossRef]
- López-Posadas, R.; Requena, P.; González, R.; Suárez, M.D.; Zarzuelo, A.; Sánchez de Medina, F.; Martínez-Augustin, O. Bovine glycomacropeptide has intestinal antiinflammatory effects in rats with dextran sulfate-induced colitis. J. Nutr. 2010, 140, 2014–2019. [Google Scholar] [CrossRef]
- Requena, P.; Daddaoua, A.; Guadix, E.; Zarzuelo, A.; Suárez, M.D.; Sánchez de Medina, F.; Martínez-Augustin, O. Bovine glycomacropeptide induces cytokine production in human monocytes through the stimulation of the MAPK and the NF-kappaB signal transduction pathways. Br. J. Pharmacol. 2009, 157, 1232–1240. [Google Scholar] [CrossRef]
- Ortega-González, M.; Capitán-Cañadas, F.; Requena, P.; Ocón, B.; Romero-Calvo, I.; Aranda, C.; Suárez, M.D.; Zarzuelo, A.; Sánchez de Medina, F.; Martínez-Augustin, O. Validation of bovine glycomacropeptide as an intestinal anti-inflammatory nutraceutical in the lymphocyte-transfer model of colitis. Br. J. Nutr. 2014, 111, 1202–1212. [Google Scholar] [CrossRef]
- Hvas, C.L.; Dige, A.; Bendix, M.; Wernlund, P.G.; Christensen, L.A.; Dahlerup, J.F.; Agnholt, J. Casein glycomacropeptide for active distal ulcerative colitis: A randomized pilot study. Eur. J. Clin. Investig. 2016, 46, 555–563. [Google Scholar] [CrossRef]
- Jiménez, M.; Cervantes-García, D.; Muñoz, Y.H.; García, A.; Haro, L.M., Jr.; Salinas, E. Novel Mechanisms Underlying the Therapeutic Effect of Glycomacropeptide on Allergy: Change in Gut Microbiota, Upregulation of TGF-β, and Inhibition of Mast Cells. Int. Arch. Allergy Immunol. 2016, 17, 217–226. [Google Scholar] [CrossRef]
- Roldán, N.R.; Jiménez, M.; Cervantes-García, D.; Marín, E.; Salinas, E. Glycomacropeptide administration attenuates airway inflammation and remodeling associated to allergic asthma in rat. Inflamm. Res. 2016, 65, 273–283. [Google Scholar] [CrossRef]
- Muñoz, F.C.; Cervantes, M.M.; Cervantes-García, D.; Jiménez, M.; Ventura-Juárez, J.; Salinas, E. Glycomacropeptide Attenuates Inflammation, Pruritus, and Th2 Response Associated with Atopic Dermatitis Induced by 2,4-Dinitrochlorobenzene in Rat. J. Immunol. Res. 2017, 2017, 6935402. [Google Scholar] [CrossRef]
- Galli, S.J.; Tsai, M.; Piliponsky, A.M. The development of allergic inflammation. Nature 2008, 454, 445–454. [Google Scholar] [CrossRef]
- Cheng, X.; Gao, D.; Chen, B.; Mao, X. Endotoxin-Binding Peptides Derived from Casein Glycomacropeptide Inhibit Lipopolysaccharide-Stimulated Inflammatory Responses via Blockade of NF-κB activation in macrophages. Nutrients 2015, 7, 3119–3137. [Google Scholar] [CrossRef]
- Li, T.; Gao, D.; Du, M.; Cheng, X.; Mao, X. Casein glycomacropeptide hydrolysates inhibit PGE2 production and COX2 expression in LPS-stimulated RAW 264.7 macrophage cells via Akt mediated NF-κB and MAPK pathways. Food Funct. 2018, 9, 2524–2532. [Google Scholar] [CrossRef]
- Cheng, X.; Gao, D.X.; Song, J.J.; Rena, F.Z.; Mao, X.Y. Casein glycomacropeptide hydrolysate exerts cytoprotection against H2O2-induced oxidative stress in RAW 264.7 macrophages via ROS dependent heme oxygenase-1 expression. RSC Adv. 2015, 5, 4511–4523. [Google Scholar] [CrossRef]
- Li, T.; Chen, B.; Du, M.; Song, J.; Cheng, X.; Wang, X.; Mao, X. Casein Glycomacropeptide Hydrolysates Exert Cytoprotective Effect against Cellular Oxidative Stress by Up-Regulating HO-1 Expression in HepG2 Cells. Nutrients. 2017, 9, 31. [Google Scholar] [CrossRef]
- Song, J.J.; Wang, Q.; Du, M.; Li, T.G.; Chen, B.; Mao, X.Y. Casein glycomacropeptide-derived peptide IPPKKNQDKTE ameliorates high glucose-induced insulin resistance in HepG2 cells via activation of AMPK signaling. Mol. Nutr. Food Res. 2017, 61, 1–12. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Specie and Strain | GMP Doses and Treatment | Effects and No Effects, Relative to Comparison Groups | References | |
|---|---|---|---|---|---|
| Colitis | TNBS- induced | Female Wistar rats | 500 mg/kg/day; Pre- and post-treatment | Effects: ↓ Colonic weight:length ratio ↓ Necrosis extension ↓ Damage score ↑ Food intake ↓ Weight loss ↓ Colonic AP activity ↓ iNOS expression in colon ↓ mRNA levels of IL-1β, IL-1ra, MUC4 and TFF3 in colon Intensity of effects: pre-treatment > post-treatment. No effects on: - Glutathione reduced levels in colon - Colonic mRNA expression of TGF-β | [76] |
| DSS-induced | Female Wistar rats | 500 mg/kg/day; Pre-treatment | Effects: ↓ Damage score ↑ Food intake ↓ ConA-stimulated IFN-γ secretion on mesenteric node cells ↓ mRNA levels of IL-1β, IL-6, IL-23, IL-17, IL-10, TGF- β, and Foxp3 in colon No effects on: - Lost of body weight - Increased AP and MPO colonic activity - Augmented IL-2 secretion stimulated by ConA on mesenteric node cells | [78] | |
| Lymphocyte transfer model | Female C57BL/6 mice | 15 mg/day; Post-treatment | No effects on: - Lost of body weight - Damage score - Increased colonic weight:length ratio - Augmented AP and MPO colonic activity - Increased mRNA levels of REG3γ, S100A8, CXCL1, and IL-1β in colon - Increased TNF-α, IL-17, IL-10, and IL-6 secretion stimulated by ConA on mesenteric node cells | [80] | |
| DSS-induced | Female C57BL/6 mice | 15 mg/day; Pre-treatment | Effects: ↑ Up-regulated IL-10 secretion by ConA-stimulated mesenteric node cells No effects on: - Lost of body weight - Damage score - Increased colonic weight–length ratio and colon length - Augmented AP and MPO colonic activity - Increased IL-17 secretion stimulated by ConA on mesenteric node cells | [80] | |
| OXZ-induced | Male BALB/c mice | 50 mg/kg/day; Treatment | Effects: ↓ Weight loss ↓ Intestinal morphological injury score ↓ Increased expression of CD4, CD8, and MAdCAM-1 in lamina propria, and MEKK1 and Smad7 in intestinal tissue ↑ Decreased levels of IgAs in lamina propria No effects on: - Smad3 levels in intestinal tissue | [75] | |
| Ileitis | TNBS-induced | Female Wistar rats | 500 mg/kg/day; Pre-treatment | Effects: ↓ Intestinal necrosis extension ↓ Damage score ↓ AP and MPO intestinal activity ↓ iNOS expression in ileum ↓ IL-1β, TNF-α, IL-1ra, IL-17, and TFF3 mRNA levels in ileal tissue No effects on: - Decreased weight loss or increased food intake - Increased COX-2 levels - Increased secretion of TNF-α, IFN-β, and IL-2 by in vitro stimulated mesenteric lymph node cells - Increased levels of RNA to Foxp3 in ileum | [77] |
| Hives and anafilaxis | OVA-induced | Male Wistar rats | 500 mg/kg/day; Pre-treatment | Effects: ↓ Allergen-specific IgE ↓ Splenocyte proliferation induced by allergen and ConA ↓ IL-13 and ↑ TGF-β splenocyte secretion stimulated by allergen ↑ Intestinal Lactobacillus, Bifidobacterium, and Bacteroides ↓ Intensity of inflammatory reaction and cutaneous mast-cell activation induced by i.d. allergen injection ↓ Score of anaphylactic shock and ↑ rectal temperature and rate of survival, induced by i.v. allergen injection No effects on: - Increased secretion of IL-10 by splenocytes in response to allergen - Number of cutaneous mast cells | [70,82] |
| Asthma | OVA-induced | Male Wistar rats | 500 mg/kg/day; Pre-treatment | Effects: ↓ Allergen-specific IgE ↓ Eosinophils and ↑ lymphocytes in blood ↓ Total cells in BALF, particularly basophils, neutrophils and lymphocytes ↓ Lung eosinophil infiltration, goblet cell hyperplasia and collagen deposition ↓ Lung TGF-β expression ↓ IL-5 and IL-13, but ↑ IL-10 mRNA levels in lung No effects on: - Increased eosinophils number in BALF | [83] |
| Atopic dermatitis | DNCB-induced | Male Wistar rats | 500 mg/kg/day; Pre- and post-treatment | Effects: ↓ Total IgE ↓ Cutaneous inflammatory process induced by topical application of allergen ↓ Pruritus ↓ Skin eosinophil infiltration and mast-cell hyperplasia ↓ IL-4, IL-5, and IL-13, but ↑ IL-10 mRNA levels in skin. Intensity of effects: pre-treatment > post-treatment. No effects on: - Pruritus when GMP was administered as post-treatment | [84] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Córdova-Dávalos, L.E.; Jiménez, M.; Salinas, E. Glycomacropeptide Bioactivity and Health: A Review Highlighting Action Mechanisms and Signaling Pathways. Nutrients 2019, 11, 598. https://doi.org/10.3390/nu11030598
Córdova-Dávalos LE, Jiménez M, Salinas E. Glycomacropeptide Bioactivity and Health: A Review Highlighting Action Mechanisms and Signaling Pathways. Nutrients. 2019; 11(3):598. https://doi.org/10.3390/nu11030598
Chicago/Turabian StyleCórdova-Dávalos, Laura Elena, Mariela Jiménez, and Eva Salinas. 2019. "Glycomacropeptide Bioactivity and Health: A Review Highlighting Action Mechanisms and Signaling Pathways" Nutrients 11, no. 3: 598. https://doi.org/10.3390/nu11030598
APA StyleCórdova-Dávalos, L. E., Jiménez, M., & Salinas, E. (2019). Glycomacropeptide Bioactivity and Health: A Review Highlighting Action Mechanisms and Signaling Pathways. Nutrients, 11(3), 598. https://doi.org/10.3390/nu11030598