Dietary Components That May Influence the Disturbed Gut Microbiota in Chronic Kidney Disease


 , , and
, , and
Abstract

1. Introduction
2. Probiotics
3. Paraprobiotics and Postbiotics
4. Prebiotics
5. Synbiotics
6. Bioactive Compounds
7. Low Protein Diet
8. Dietary Patterns and Microbiota
9. Other Diet Components
10. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Petrova, P.; Petrov, K. Prebiotic-Probiotic Relationship: The Genetic Fundamentals of Polysaccharides Conversion by Bifidobacterium and Lactobacillus Genera. Food Bioconvers. 2017, 2, 237–278. [Google Scholar]
- Aagaard, K.; Ma, J.; Antony, K.M.; Ganu, R.; Petrosino, J.; Versalovic, J. The Placenta Harbors a Unique Microbiome Kjersti. Sci. Transl. Med. 2014, 6, 1–22. [Google Scholar]
- Tang, W.H.W.; Kitai, T.; Hazen, S.L.; Clinic, C. Gut Microbiota in Cardiovascular Health and Disease. Circ. Res. 2017, 120, 1183–1196. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, stability and resilience of the human gut microbiota. Nature 2012, 489, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Vaziri, N.D.; Zhao, Y.Y.; Pahl, M.V. Altered intestinal microbial flora and impaired epithelial barrier structure and function in CKD: The nature, mechanisms, consequences and potential treatment. Nephrol. Dial. Transplant. 2015, 31, 737–746. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.; Piceno, Y.M.; Desantis, T.Z.; Pahl, M.; Andersen, G.L.; Vaziri, N.D. Expansion of urease- and uricase-containing, indole- and p-cresol-forming and contraction of short chain fatty acid-producing intestinal microbiota in ESRD. Am. J. Nephrol. 2014, 39, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Koppe, L.; Mafra, D.; Fouque, D. Probiotics and chronic kidney disease. Kidney Int. 2015, 88, 958–966. [Google Scholar] [CrossRef] [PubMed]
- Ramezani, A.; Massy, Z.A.; Meijers, B.; Evenepoel, P.; Vanholder, R.; Raj, D.S. Role of the Gut Microbiome in Uremia: A Potential Therapeutic Target. Am. J. Kidney Dis. 2016, 67, 483–498. [Google Scholar] [CrossRef] [PubMed]
- Pisano, A.; D’Arrigo, G.; Coppolino, G.; Bolignano, D. Biotic Supplements for Renal Patients: A Systematic Review and Meta-Analysis. Nutrients 2018, 10, 1224. [Google Scholar] [CrossRef] [PubMed]
- Lobach, A.R.; Roberts, A.; Rowland, I.R. Assessing the in vivo data on low/no-calorie sweeteners and the gut microbiota. Food Chem. Toxicol. 2019, 124, 385–399. [Google Scholar] [CrossRef] [PubMed]
- Sonnenburg, J.L.; Bäckhed, F. Diet–microbiota interactions as moderators of human metabolism. Nature 2016, 535, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Ranganathan, N.; Ranganathan, P.; Friedman, E.A.; Joseph, A.; Delano, B.; Goldfarb, D.S.; Tam, P.; Rao, A.V.; Anteyi, E.; Musso, C.G. Pilot study of probiotic dietary supplementation for promoting healthy kidney function in patients with chronic kidney disease. Adv. Ther. 2010, 27, 634–647. [Google Scholar] [CrossRef] [PubMed]
- Borges, N.A.; Carmo, F.L.; Stockler-Pinto, M.B.; de Brito, J.S.; Dolenga, C.J.; Ferreira, D.C.; Nakao, L.S.; Rosado, A.; Fouque, D.; Mafra, D. Probiotic Supplementation in Chronic Kidney Disease: A Double-blind, Randomized, Placebo-controlled Trial. J. Ren. Nutr. 2018, 28, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Vaziri, N.D.; Liu, S.M.; Lau, W.L.; Khazaeli, M.; Nazertehrani, S.; Farzaneh, S.H.; Kieffer, D.A.; Adams, S.H.; Martin, R.J. High Amylose Resistant Starch Diet Ameliorates Oxidative Stress, Inflammation, and Progression of Chronic Kidney Disease. PLoS ONE 2014, 9, e114881. [Google Scholar] [CrossRef] [PubMed]
- Claro, L.M.; Moreno-Amaral, A.N.; Gadotti, A.C.; Dolenga, C.J.; Nakao, L.S.; Azevedo, M.L.V.; de Noronha, L.; Olandoski, M.; de Moraes, T.P.; Stinghen, A.E.M.; et al. The impact of uremic toxicity induced inflammatory response on the cardiovascular burden in chronic kidney disease. Toxins 2018, 10, 384. [Google Scholar] [CrossRef] [PubMed]
- Esgalhado, M.; Kemp, J.A.; Azevedo, R.; Paiva, B.R.; Stockler-Pinto, M.B.; Dolenga, C.J.; Borges, N.A.; Nakao, L.S.; Mafra, D. Could resistant starch supplementation improve inflammatory and oxidative stress biomarkers and uremic toxins levels in hemodialysis patients? A pilot randomized controlled trial. Food Funct. 2018, 9, 6508–6516. [Google Scholar] [CrossRef] [PubMed]
- Rossi, M.; Johnson, D.W.; Morrison, M.; Pascoe, E.M.; Coombes, J.S.; Forbes, J.M.; Szeto, C.-C.; McWhinney, B.C.; Ungerer, J.P.J.; Campbell, K.L. Synbiotics Easing Renal Failure by Improving Gut Microbiology (SYNERGY): A Randomized Trial. Clin. J. Am. Soc. Nephrol. 2016, 11, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Alvarenga, L.A.; De Oliveira Leal, V.; Borges, N.A.; Silva De Aguiar, A.; Faxén-Irving, G.; Stenvinkel, P.; Lindholm, B.; Denise, M. Curcumin—A promising nutritional strategy for chronic kidney disease patients. J. Funct. Foods 2018, 40, 715–721. [Google Scholar] [CrossRef]
- Martins, I.C.V.S.; Borges, N.A.; Stenvinkel, P.; Lindholm, B.; Rogez, H.; Pinheiro, M.C.N.; Nascimento, J.L.M.; Mafra, D. The value of the Brazilian açai fruit as a therapeutic nutritional strategy for chronic kidney disease patients. Int. Urol. Nephrol. 2018, 50, 2207–2220. [Google Scholar] [CrossRef] [PubMed]
- Black, A.P.; Anjos, J.S.; Cardozo, L.; Carmo, F.L.; Dolenga, C.J.; Nakao, L.S.; de Carvalho Ferreira, D.; Rosado, A.; Carraro Eduardo, J.C.; Mafra, D. Does Low-Protein Diet Influence the Uremic Toxin Serum Levels From the Gut Microbiota in Nondialysis Chronic Kidney Disease Patients? J. Ren. Nutr. 2018, 28, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S. Expert consensus document: The international scientific association for probiotics and prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Siciliano, R.A.; Mazzeo, M.F. Molecular mechanisms of probiotic action: A proteomic perspective. Curr. Opin. Microbiol. 2012, 15, 390–396. [Google Scholar] [CrossRef] [PubMed]
- Alatriste, P.V.M.; Arronte, R.U.; Espinosa, C.O.G.; Cuevas, M.E. Effect of probiotics on human blood urea levels in patients with chronic renal failure. Nutr. Hosp. 2014, 29, 582–590. [Google Scholar]
- Barros, A.F.; Borges, N.A.; Nakao, L.S.; Dolenga, C.J.; do Carmo, F.L.; de Carvalho Ferreira, D.; Stenvinkel, P.; Bergman, P.; Lindholm, B.; Mafra, D. Effects of probiotic supplementation on inflammatory biomarkers and uremic toxins in non-dialysis chronic kidney patients: A double-blind, randomized, placebo-controlled trial. J. Funct. Foods 2018, 46, 378–383. [Google Scholar]
- Taki, K.; Takayama, F.; Niwa, T. Beneficial effects of Bifidobacteria in a gastroresistant seamless capsule on hyperhomocysteinemia in hemodialysis patients. J. Ren. Nutr. 2005, 15, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Eidi, F.; Poor-reza Gholi, F.; Ostadrahimi, A.; Dalili, N.; Samadian, F.; Barzegari, A. Effect of Lactobacillus Rhamnosus on serum uremic toxins (phenol and P-Cresol) in hemodialysis patients: A double blind randomized clinical trial. Clin. Nutr. ESPEN 2018, 28, 158–164. [Google Scholar] [CrossRef] [PubMed]
- McFarlane, C.; Ramos, C.I.; Johnson, D.W.; Campbell, K.L. Prebiotic, Probiotic, and Synbiotic Supplementation in Chronic Kidney Disease: A Systematic Review and Meta-analysis. J. Ren. Nutr. 2018, 18, S1051–S2276. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, R.; Pechenyak, B.; Vyas, U.; Ranganathan, P.; Weinberg, A.; Liang, P.; Mallappallil, M.C.; Norin, A.J.; Friedman, E.A.; Saggi, S.J. Randomized Controlled Trial of Strain-Specific Probiotic Formulation (Renadyl) in Dialysis Patients. Biomed Res. Int. 2014, 2014, 568571. [Google Scholar] [CrossRef] [PubMed]
- Wang, I.K.; Wu, Y.Y.; Yang, Y.F.; Ting, I.W.; Lin, C.C.; Yen, T.H.; Chen, J.H.; Wang, C.H.; Huang, C.C.; Lin, H.C. The effect of probiotics on serum levels of cytokine and endotoxin in peritoneal dialysis patients: A randomised, double-blind, placebo-controlled trial. Benef. Microbes 2015, 6, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Soleimani, A.; Mojarrad, M.Z.; Bahmani, F.; Taghizadeh, M.; Ramezani, M.; Tajabadi-Ebrahimi, M.; Jafari, P.; Esmaillzadeh, A.; Asemi, Z. Probiotic supplementation in diabetic hemodialysis patients has beneficial metabolic effects. Kidney Int. 2016, 91, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Shariaty, Z.; Shan, G.R.M.; Farajollahi, M.; Amerian, M.; Pour, N.B. The effects of probiotic supplement on hemoglobin in chronic renal failure patients under hemodialysis: A randomized clinical trial. J. Res. Med. Sci. 2017, 22, 1–5. [Google Scholar]
- Sánchez, B.; Delgado, S.; Blanco-Míguez, A.; Lourenço, A.; Gueimonde, M.; Margolles, A. Probiotics, gut microbiota, and their influence on host health and disease. Mol. Nutr. Food Res. 2017, 61, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Polk, D.B. Probiotics and immune health. Curr. Opin. Gastroenterol. 2011, 27, 496–501. [Google Scholar] [CrossRef] [PubMed]
- Lau, W.L.; Kalantar-zadeh, K.; Vaziri, N.D. The gut as a source of inflammation in chronic kidney disease. Nephron 2015, 130, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Grimoud, J.; Durand, H.; Souza, S.; Monsan, P.; Ouarné, F.; Theodorou, V.; Roques, C. In vitro screening of probiotics and synbiotics according to anti-inflammatory and anti-proliferative effects. Int. J. Food Microbiol. 2010, 144, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Jensen, H.; Drømtorp, S.M.; Axelsson, L.; Grimmer, S. Immunomodulation of Monocytes by Probiotic and Selected Lactic Acid Bacteria. Probiotics Antimicrob. Proteins 2015, 7, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Rocha-Ramírez, L.M.; Pérez-Solano, R.A.; Castañón-Alonso, S.L.; Moreno Guerrero, S.S.; Ramírez Pacheco, A.; García Garibay, M.; Eslava, C. Probiotic Lactobacillus Strains Stimulate the Inflammatory Response and Activate Human Macrophages. J. Immunol. Res. 2017, 2017, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Gareau, M.G.; Sherman, P.M.; Walker, W.A. Study on the method for evaluating ecological health of the main river network in the Yellow River’s estuary area. Nat. Rev. Gastroenterol. Hepatol. 2010, 7, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Toalá, J.E.; Garcia-Varela, R.; Garcia, H.S.; Mata-Haro, V.; González-Córdova, A.F.; Vallejo-Cordoba, B.; Hernández-Mendoza, A. Postbiotics: An evolving term within the functional foods field. Trends Food Sci. Technol. 2018, 75, 105–114. [Google Scholar] [CrossRef]
- Fisberg, R.M.; Marchioni, D.M.L.; Colucci, A.C.A. Avaliação do consumo alimentar e da ingestão de nutrientes na prática clínica. Arq Bras Endocrinol. Metabol. 2009, 53, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Cosola, C.; Rocchetti, M.T.; Cupisti, A.; Gesualdo, L. Microbiota metabolites: Pivotal players of cardiovascular damage in chronic kidney disease. Pharmacol. Res. 2018, 130, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. The International Scientific Association for Probiotics and Prebiotics (ISAPP) Consensus on the Definition and Scope of Prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14. [Google Scholar] [CrossRef]
- Slavin, J. Fiber and prebiotics: Mechanisms and health benefits. Nutrients 2013, 5, 1417–1435. [Google Scholar] [CrossRef] [PubMed]
- Distrutti, E.; Monaldi, L.; Ricci, P.; Fiorucci, S. Gut microbiota role in irritable bowel syndrome: New therapeutic strategies. World J. Gastroenterol. 2016, 22, 2219–2241. [Google Scholar] [CrossRef] [PubMed]
- Ramezani, A.; Raj, D.S. The Gut Microbiome, Kidney Disease, and Targeted Interventions. J. Am. Soc. Nephrol. 2013, 25, 657–670. [Google Scholar] [CrossRef] [PubMed]
- Barczynska, R.; Bandurska, K.; Slizewska, K.; Litwin, M.; Szalecki, M.; Libudzisz, Z.; Szalecki, M.; Libudzisz, Z.; Kapusniak, J. Intestinal microbiota, obesity and prebiotics. Pol. J. Microbiol. 2015, 64, 93–100. [Google Scholar] [PubMed]
- Valcheva, R.; Dieleman, L.A. Prebiotics: Definition and protective mechanisms. Best Pract. Res. Clin. Gastroenterol. 2016, 30, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Kieffer, D.A.; Piccolo, B.D.; Vaziri, N.D.; Liu, S.; Lau, W.L.; Khazaeli, M.; Nazertehrani, S.; Moore, M.E.; Marco, M.L.; et al. Resistant starch alters gut microbiome and metabolomic profiles concurrent with amelioration of chronic kidney disease in rats. Am. J. Physiol. Physiol. 2016, 310, F857–F871. [Google Scholar] [CrossRef] [PubMed]
- Hung, T.V.; Suzuki, T. Dietary fermentable fibers attenuate chronic kidney disease in mice by protecting the intestinal barrier. J. Nutr. 2018, 148, 552–561. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Li, Q.; Henning, S.M.; Zhong, J.; Hsu, M.; Lee, R.; Long, J.; Chan, B.; Nagami, G.T.; Heber, D.; Li, Z. Effects of Prebiotic Fiber Xylooligosaccharide in Adenine-Induced Nephropathy in Mice. Mol. Nutr. Food Res. 2018, 62, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Tayebi-Khosroshahi, H.; Habibzadeh, A.; Niknafs, B.; Ghotaslou, R.; Yeganeh Sefidan, F.; Ghojazadeh, M.; Moghaddaszadeh, M.; Parkhide, S. The effect of lactulose supplementation on fecal microflora of patients with chronic kidney disease; a randomized clinical trial. J. Ren. Inj. Prev. 2016, 5, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Sirich, T.L.; Plummer, N.S.; Gardner, C.D.; Hostetter, T.H.; Meyer, T.W. Effect of Increasing Dietary Fiber on Plasma Levels of Colon-Derived Solutes in Hemodialysis Patients. Clin. J. Am. Soc. Nephrol. 2014, 9, 1603–1610. [Google Scholar] [CrossRef] [PubMed]
- Salmean, Y.A.; Segal, M.S.; Palii, S.P.; Dahl, W.J. Fiber supplementation lowers plasma p-cresol in chronic kidney disease patients. J. Ren. Nutr. 2015, 25, 316–320. [Google Scholar] [CrossRef] [PubMed]
- Koppe, L.; Pillon, N.J.; Vella, R.E.; Croze, M.L.; Pelletier, C.C.; Chambert, S.; Massy, Z.; Glorieux, G.; Vanholder, R.; Dugenet, Y.; et al. p-Cresyl Sulfate Promotes Insulin Resistance Associated with CKD. J. Am. Soc. Nephrol. 2012, 24, 88–99. [Google Scholar] [CrossRef] [PubMed]
- Al Za’Abi, M.; Al Salam, S.; Al Suleimani, Y.; Manoj, P.; Nemmar, A.; Ali, B.H. Gum Acacia Improves Renal Function and Ameliorates Systemic Inflammation, Oxidative and Nitrosative Stress in Streptozotocin-Induced Diabetes in Rats with Adenine-Induced Chronic Kidney Disease. Cell. Physiol. Biochem. 2018, 45, 2293–2304. [Google Scholar] [CrossRef] [PubMed]
- Meijers, B.K.I.; De Preter, V.; Verbeke, K.; Vanrenterghem, Y.; Evenepoel, P. P-Cresyl sulfate serum concentrations in haemodialysis patients are reduced by the prebiotic oligofructose-enriched inulin. Nephrol. Dial. Transplant. 2010, 25, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Poesen, R.; Evenepoel, P.; Loor, H.; Delcour, J.A.; Courtin, C.M.; Kuypers, D.; Augustijns, P.; Verbeke, K.; Meijers, B. The Influence of Prebiotic Arabinoxylan Oligosaccharides on Microbiota Derived Uremic Retention Solutes in Patients with Chronic Kidney Disease: A Randomized Controlled Trial. PLoS ONE 2016, 11, e0153893. [Google Scholar] [CrossRef] [PubMed]
- Younes, H.; Egret, N.; Hadj-Abdelkader, M.; Rémésy, C.; Demigné, C.; Gueret, C.; Deteix, P.; Alphonse, J.C. Fermentable carbohydrate supplementation alters nitrogen excretion in chronic renal failure. J. Ren. Nutr. 2006, 16, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Tayebi Khosroshahi, H.; Vaziri, N.D.; Abedi, B.; Asl, B.H.; Ghojazadeh, M.; Jing, W.; Vatankhah, A.M. Effect of high amylose resistant starch (HAM-RS2) supplementation on biomarkers of inflammation and oxidative stress in hemodialysis patients: A randomized clinical trial. Hemodial. Int. 2018, 22, 492–500. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Roberfroid, M.B. Dietary Modulation of the Human Colonic Microbiota: Introducing the Concept of Prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [CrossRef] [PubMed]
- Markowiak, P.; Ślizewska, K. Effects of probiotics, prebiotics, and synbiotics on human health. Nutrients 2017, 9, 1–30. [Google Scholar]
- Oliveira, G.; González-Molero, I. Actualización de probióticos, prebióticos y simbióticos en nutrición clínica. Endocrinol. Nutr. 2016, 63, 482–494. [Google Scholar] [CrossRef] [PubMed]
- Schrezenmeir, J.; Vrese, M. Probiotics, prebiotics, and synbiotics—Approaching a definition. Am. J. Clin. Nutr. 2018, 73, 361S–364S. [Google Scholar] [CrossRef] [PubMed]
- Vaziri, N.D. Effect of Synbiotic Therapy on Gut–Derived Uremic Toxins and the Intestinal Microbiome in Patients with CKD. Clin. J. Am. Soc. Nephrol. 2016, 11, 199–201. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Mora, J.; Martínez-Hernández, N.E.; Castro-Alarcón, N.; Parra-Rojas, I.; Viramontes-Hörner, D.; García-García, G.; Parra-Rojas, I.; Castro-Alarcón, N. Effects of a Symbiotic on Gut Microbiota in Mexican Patients With End-Stage Renal Disease. J. Ren. Nutr. 2014, 24, 330–335. [Google Scholar] [CrossRef] [PubMed]
- Viramontes-Hörner, D.; Márquez-Sandoval, F.; Martín-del-Campo, F.; Vizmanos-Lamotte, B.; Sandoval-Rodríguez, A.; Armendáriz-Borunda, J.; García-Bejarano, H.; Renoirte-López, K.; García-García, G. Effect of a symbiotic gel (Lactobacillus acidophilus + Bifidobacterium lactis + Inulin) on presence and severity of gastrointestinal symptoms in hemodialysis patients. J. Ren. Nutr. 2015, 25, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Dehghani, H.; Heidari, F.; Mozaffari-Khosravi, H.; Nouri-Majelan, N.; Dehghani, A. Synbiotic Supplementations for Azotemia in Patients with Chronic Kidney Disease: A Randomized Controlled Trial. Iran. J. Kidney Dis. 2016, 10, 351–357. [Google Scholar] [PubMed]
- Pavan, M. Influence of prebiotic and probiotic supplementation on the progression of chronic kidney disease Malleshappa. Minerva Urol. Nefrol. 2016, 68, 222–226. [Google Scholar] [PubMed]
- Guida, B.; Germanò, R.; Trio, R.; Russo, D.; Memoli, B.; Grumetto, L.; Barbato, F.; Cataldi, M. Effect of short-term synbiotic treatment on plasma p-cresol levels in patients with chronic renal failure: A randomized clinical trial. Nutr. Metab. Cardiovasc. Dis. 2014, 24, 1043–1049. [Google Scholar] [CrossRef] [PubMed]
- Nakabayashi, I.; Nakamura, M.; Kawakami, K.; Ohta, T.; Kato, I.; Uchida, K.; Yoshida, M. Effects of synbiotic treatment on serum level of p-cresol in haemodialysis patients: A preliminary study. Nephrol. Dial. Transplant. 2011, 26, 1094–1098. [Google Scholar] [CrossRef] [PubMed]
- Cicero, A.F.G.; Fogacci, F.; Colletti, A. Food and plant bioactives for reducing cardiometabolic disease risk: An evidence based approach. Food Funct. 2017, 8, 2076–2088. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Chandrashekharappa, S.; Bodduluri, S.R.; Baby, B.V.; Hegde, B.; Kotla, N.G.; Hiwale, A.A.; Saiyed, T.; Patel, P.; Vijay-Kumar, M.; et al. Enhancement of the gut barrier integrity by a microbial metabolite through the Nrf2 pathway. Nat. Commun. 2019, 10, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Anhê, F.F.; Varin, T.V.; Barz, M.L.; Desjardins, Y.; Levy, E.; Roy, D.; Marette, A. Gut Microbiota Dysbiosis in Obesity-Linked Metabolic Diseases and Prebiotic Potential of Polyphenol-Rich Extracts. Curr. Obes. Rep. 2015, 4, 389–400. [Google Scholar] [CrossRef] [PubMed]
- Anhê, F.F.; Pilon, G.; Roy, D.; Desjardins, Y.; Levy, E.; Marette, A. Triggering Akkermansia with dietary polyphenols: A new weapon to combat the metabolic syndrome? Gut Microbes 2016, 7, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Henning, S.M.; Lee, R.P.; Lu, Q.Y.; Summanen, P.H.; Thames, G.; Corbett, K.; Downes, J.; Tseng, C.H.; Finegold, S.M. Pomegranate extract induces ellagitannin metabolite formation and changes stool microbiota in healthy volunteers. Food Funct. 2015, 6, 2487–2495. [Google Scholar] [CrossRef] [PubMed]
- Choya, Y.Y.; Quifer-Rada, P.; Holstege, D.M.; Frese, S.A.; Calvert, C.C.; Mills, D.A.; Lamuela-Raventos, R.M.; Waterhouse, A.L. Phenolic metabolites and substantial microbiome changes in pig feces by ingesting grape seed proanthocyanidins. Food Funct. 2014, 5, 2298–2308. [Google Scholar] [CrossRef] [PubMed]
- Roopchand, D.E.; Carmody, R.N.; Kuhn, P.; Moskal, K.; Rojas-Silva, P.; Turnbaugh, P.J.; Raskin, I. Dietary polyphenols promote growth of the gut bacterium akkermansia muciniphila and attenuate high-fat diet-induced metabolic syndrome. Diabetes 2015, 64, 2847–2858. [Google Scholar] [CrossRef] [PubMed]
- Etxeberria, U.; Fernández-Quintela, A.; Milagro, F.I.; Aguirre, L.; Martínez, J.A.; Portillo, M.P. Impact of polyphenols and polyphenol-rich dietary sources on gut microbiota composition. J. Agric. Food Chem. 2013, 61, 9517–9533. [Google Scholar] [CrossRef] [PubMed]
- Fang, J. Bioavailability of anthocyanins. Drug. Metab. Rev. 2013, 2532, 2465–2487. [Google Scholar] [CrossRef] [PubMed]
- Esposito, D.; Damsud, T.; Wilson, M.; Grace, M.H.; Strauch, R.; Li, X.; Lila, M.A.; Komarnytsky, S. Black Currant Anthocyanins Attenuate Weight Gain and Improve Glucose Metabolism in Diet-Induced Obese Mice with Intact, but Not Disrupted, Gut Microbiome. J. Agric. Food Chem. 2015, 63, 6172–6180. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.S.; Gehr, T.W.B.; Ghosh, S. Curcumin and chronic kidney disease (CKD): Major mode of action through stimulating endogenous intestinal alkaline phosphatase. Molecules 2014, 19, 20139–20156. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Liu, L.; Ji, H.F. Regulative effects of curcumin spice administration on gut microbiota and its pharmacological implications. Food Nutr. Res. 2017, 61, 1361780. [Google Scholar] [CrossRef] [PubMed]
- Ohno, M.; Nishida, A.; Sugitani, Y.; Nishino, K.; Inatomi, O.; Sugimoto, M.; Kawahara, M.; Andoh, A. Nanoparticle curcumin ameliorates experimental colitis via modulation of gut microbiota and induction of regulatory T cells. PLoS ONE 2017, 12, e0185999. [Google Scholar] [CrossRef] [PubMed]
- Clavel, T.; Fallani, M.; Lepage, P.; Levenez, F.; Mathey, J.; Rochet, V.; Sérézat, M.; Sutren, M.; Henderson, G.; Bennetau-Pelissero, C.; et al. Isoflavones and Functional Foods Alter the Dominant Intestinal Microbiota in Postmenopausal Women. J. Nutr. 2005, 135, 2786–2792. [Google Scholar] [CrossRef] [PubMed]
- Queipo-Ortuño, M.I.; Boto-Ordonez, M.; Murri, M.; Gomez-Zumaquero, J.M.; Clemente-Postigo, M.; Estruch, R.; Diaz, F.C.; Andres-Lacueva, C. Influence of red wine polyphenols on the gut microbiota ecology. Am. J. Clin. Nutr. 2012, 95, 1323–1334. [Google Scholar] [CrossRef] [PubMed]
- Song, M.Y.; Wang, J.; Eom, T.; Kim, H. Schisandra chinensis fruit modulates the gut microbiota composition in association with metabolic markers in obese women: A randomized, double-blind placebo-controlled study. Nutr. Res. 2015, 35, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Eid, N.; Osmanova, H.; Natchez, C.; Walton, G.; Costabile, A.; Gibson, G.; Rowland, I.; Spencer, J.P. Impact of palm date consumption on microbiota growth and large intestinal health: A randomised, controlled, cross-over, human intervention study. Br. J. Nutr. 2015, 114, 1226–1236. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Indias, I.; Sánchez-Alcoholado, L.; Pérez-Martínez, P.; Andrés-Lacueva, C.; Cardona, F.; Tinahones, F.; Queipo-Ortuño, M.I. Red wine polyphenols modulate fecal microbiota and reduce markers of the metabolic syndrome in obese patients. Food Funct. 2015, 7, 1775–1787. [Google Scholar] [CrossRef] [PubMed]
- Janssens, P.L.H.R.; Penders, J.; Hursel, R.; Budding, A.E.; Savelkoul, P.H.M.; Westerterp-Plantenga, M.S. Long-Term Green Tea Supplementation Does Not Change the Human Gut Microbiota. PLoS ONE 2016, 11, e0153134. [Google Scholar] [CrossRef] [PubMed]
- Barroso, E.; Muñoz-González, I.; Jiménez, E.; Bartolomé, B.; Moreno-Arribas, V.; Peláez, C.; Del Carmen Martínez-Cuesta, M.; Requena, T. Phylogenetic profile of gut microbiota in healthy adults after moderate intake of red wine Elvira. Mol. Nutr. Food Res. 2017, 61, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Most, J.; Penders, J.; Lucchesi, M.; Goossens, G.H.; Blaak, E.E. Gut microbiota composition in relation to the metabolic response to 12-week combined polyphenol supplementation in overweight men and women. Eur. J. Clin. Nutr. 2017, 71, 1040–1045. [Google Scholar] [CrossRef] [PubMed]
- Peterson, C.T.; Vaughn, A.R.; Sharma, V.; Chopra, D.; Mills, P.J.; Peterson, S.N.; Sivamani, R.K. Effects of Turmeric and Curcumin Dietary Supplementation on Human Gut Microbiota: A Double-Blind, Randomized, Placebo-Controlled Pilot Study. J. Evid.-Based Integr. Med. 2018, 23, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Mafra, D.; Barros, A.F.; Fouque, D. Dietary protein metabolism by gut microbiota and its consequences for chronic kidney disease patients. Future Microbiol. 2013, 8, 1317–1323. [Google Scholar] [CrossRef] [PubMed]
- Kopple, J.D. National kidney foundation K/DOQI clinical practice guidelines for nutrition in chronic renal failure. Am. J. Kidney Dis. 2001, 37, S66–S70. [Google Scholar] [CrossRef] [PubMed]
- Marzocco, S.; Dal Piaz, F.; Di Micco, L.; Torraca, S.; Sirico, M.L.; Tartaglia, D.; Autore, G.; Di Iorio, B. Very Low Protein Diet Reduces Indoxyl Sulfate Levels in Chronic Kidney Disease. Blood Purif. 2013, 35, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Mantrana, I.; Selma-Royo, M.; Alcantara, C.; Collado, M.C. Shifts on gut microbiota associated to mediterranean diet adherence and specific dietary intakes on general adult population. Front. Microbiol. 2018, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Klinder, A.; Shen, Q.; Heppel, S.; Lovegrove, J.A.; Rowland, I.; Tuohy, K.M. Impact of increasing fruit and vegetables and flavonoid intake on the human gut microbiota. Food Funct. 2016, 7, 1788–1796. [Google Scholar] [CrossRef] [PubMed]
- Berryman, C.E.; Agarwal, S.; Lieberman, H.R.; Fulgoni, V.L.; Pasiakos, S.M. Diets higher in animal and plant protein are associated with lower adiposity and do not impair kidney function in US adults. Am. J. Clin. Nutr. 2016, 104, 743–749. [Google Scholar] [CrossRef] [PubMed]
- Kandouz, S.; Mohamed, A.S.; Zheng, Y.; Sandeman, S.; Davenport, A. Reduced protein bound uraemic toxins in vegetarian kidney failure patients treated by haemodiafiltration. Hemodial. Int. 2016, 20, 610–617. [Google Scholar] [CrossRef] [PubMed]
- Patel, K.P.; Luo, F.J.G.; Plummer, N.S.; Hostetter, T.H.; Meyer, T.W. The production of p-Cresol sulfate and indoxyl sulfate in vegetarians versus omnivores. Clin. J. Am. Soc. Nephrol. 2012, 7, 982–988. [Google Scholar] [CrossRef] [PubMed]
- Mafra, D.; Borges, N.A.; Cardozo, L.F.M.; Anjos, J.S.; Black, A.P.; Moraes, C.; Bergman, P.; Lindholm, B.; Stenvinkel, P. Red meat intake in chronic kidney disease patients: Two sides of the coin. Nutrition 2018, 46, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Koppe, L.; Fouque, D.; Soulage, C.O. The role of gut microbiota and diet on uremic retention solutes production in the context of chronic kidney disease. Toxins 2018, 10, 155. [Google Scholar] [CrossRef] [PubMed]
- Koppe, L.; Fouque, D.; Soulage, C.O. Metabolic Abnormalities in Diabetes and Kidney Disease: Role of Uremic Toxins. Curr. Diabetes Rep. 2018, 18, 97. [Google Scholar] [CrossRef] [PubMed]
- Del Chierico, F.; Vernocchi, P.; Dallapiccola, B.; Putignani, L. Mediterranean diet and health: Food effects on gut microbiota and disease control. Int. J. Mol. Sci. 2014, 15, 11678–11699. [Google Scholar] [CrossRef] [PubMed]
- Montemurno, E.; Dalfino, C.C.G.; Daidone, G.; Gesualdo, M.D. What Would You Like to Eat, Mr CKD Microbiota? A Mediterranean Diet, please! Kidney Blood Press. Res. 2014, 39, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Lamuel-Raventos, R.M.; Onge, M.P.S. Prebiotic nut compounds and human microbiota. Crit. Rev. Food Sci. Nutr. 2017, 57, 3154–3163. [Google Scholar] [CrossRef] [PubMed]
- Martín-Peláez, S.; Covas, M.I.; Fitó, M.; Kušar, A.; Pravst, I. Health effects of olive oil polyphenols: Recent advances and possibilities for the use of health claims. Mol. Nutr. Food Res. 2013, 57, 760–771. [Google Scholar] [CrossRef] [PubMed]
- Filippis, F.; Pellegrini, N.; Vannini, L.; Jeffery, I.B.; Storia, A.L.; Laghi, L.; Serrazanetti, D.I.; Di Cagno, R.; Ferrocino, I.; Lazzi, C.; et al. High-level adherence to a Mediterranean diet beneficially impacts the gut microbiota and associated metabolome. Gut Microbiota 2016, 65, 1812–1821. [Google Scholar] [CrossRef] [PubMed]
- Mitsou, E.K.; Kakali, A.; Mountzouris, S.A.K.C.; Panagiotakos, M.Y.D.B.; Kyriacou, A. Adherence to the Mediterranean diet is associated with the gut microbiota pattern and gastrointestinal characteristics in an adult population. Br. J. Nutr. 2017, 117, 1645–1655. [Google Scholar] [CrossRef] [PubMed]
- Bell, V.; Ferrão, J.; Pimentel, L.; Pintado, M.; Fernandes, T. One Health, Fermented Foods, and Gut Microbiota. Foods 2018, 7, 195. [Google Scholar] [CrossRef] [PubMed]
- Aslam, H.; Green, J.; Jacka, F.N.; Collier, F.; Berk, M.; Pasco, J.; Dawson, S.L. Fermented foods, the gut and mental health: A mechanistic overview with implications for depression and anxiety. Nutr. Neurosci. 2018, 0, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Senger, D.R.; Li, D.; Jaminet, S.C.; Cao, S. Activation of the Nrf2 cell defense pathway by ancient foods: Disease prevention by important molecules and microbes lost from the modern western diet. PLoS ONE 2016, 11, e0148042. [Google Scholar] [CrossRef] [PubMed]
- Zinöcker, M.K.; Lindseth, I.A. The western diet–microbiome-host interaction and its role in metabolic disease. Nutrients 2018, 10, 365. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Bäckhed, F.; Fulton, L.; Gordon, J.I. Diet-Induced Obesity Is Linked to Marked but Reversible Alterations in the Mouse Distal Gut Microbiome. Cell Host Microbe 2008, 3, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Spencer, M.; Gupta, A.; Dam, L.; Shannon, C.; Menees, S.; Chey, W.D. Artificial Sweeteners: A Systematic Review and Primer for Gastroenterologists. J. Neurogastroenterol. Motil. 2016, 22, 2093–2879. [Google Scholar] [CrossRef] [PubMed]
- Dudek-Wicher, R.K.; Junka, A.; Bartoszewicz, M. The influence of antibiotics and dietary components on gut microbiota. Prz Gastroenterol. 2018, 13, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.P.; Browman, D.; Herzog, H.; Gregory Neely, G. Non-nutritive sweeteners possess a bacteriostatic effect and alter gut microbiota in mice. PLoS ONE 2018, 13, e0199080. [Google Scholar] [CrossRef] [PubMed]
- Zopf, Y.; Reljic, D.; Dieterich, W. Dietary Effects on Microbiota—New Trends with Gluten-Free or Paleo Diet. Med. Sci. 2018, 6, 92. [Google Scholar] [CrossRef] [PubMed]
- Palma, G.; Nadal, I.; Collado, Y.S. Effects of a gluten-free diet on gut microbiota and immune function in healthy adult humans. Gut Microbes 2010, 1, 135–137. [Google Scholar] [CrossRef] [PubMed]
- Laparra, J.M.; Sanz, Y. Interactions of gut microbiota with functional food components and nutraceuticals. Pharmacol. Res. 2010, 61, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Mahalhal, A.; Williams, J.M.; Johnson, S.; Ellaby, N.; Duckworth, C.A.; Burkitt, M.D.; Liu, X.; Hold, G.L.; Campbell, B.J.; Pritchard, D.M.; et al. Oral iron exacerbates colitis and influences the intestinal microbiome. PLoS ONE 2018, 13, e0202460. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, B.; Li, H. Gut Microbiota and Iron: The Crucial Actors in Health and Disease. Pharmaceuticals 2018, 11, 98. [Google Scholar] [CrossRef] [PubMed]
- Kortman, G.A.M.; Reijnders, D.; Swinkels, D.W. Oral iron supplementation: Potential implications for the gut microbiome and metabolome in patients with CKD. Hemodial. Int. 2017, 21, S28–S36. [Google Scholar] [CrossRef] [PubMed]
- Crespo-Salgado, J.; Vehaskari, V.M.; Stewart, T.; Ferris, M.; Zhang, Q.; Wang, G.; Blanchard, E.E.; Taylor, C.M.; Kallash, M.; Greenbaum, L.A.; et al. Intestinal microbiota in pediatric patients with end stage renal disease: A Midwest Pediatric Nephrology Consortium study. Microbiome 2016. [Google Scholar] [CrossRef] [PubMed]
- Crowley, E.K.; Long-Smith, C.M.; Murphy, A.; Patterson, E.; Murphy, K.; O’Gorman, D.M.; Stanton, C.; Nolan, Y.M. Dietary Supplementation with a Magnesium-Rich Marine Mineral Blend Enhances the Diversity of Gastrointestinal Microbiota. Mar. Drugs 2018, 16, 216. [Google Scholar] [CrossRef] [PubMed]
- Chaplin, A.; Parra, P.; Laraichi, S.; Serra, F.; Palou, A. Calcium supplementation modulates gut microbiota in a prebiotic manner in dietary obese mice. Mol. Nutr. Food Res. 2016, 60, 468–480. [Google Scholar] [CrossRef] [PubMed]
- Miao, Y.Y.; Xu, C.M.; Xia, M.; Zhu, H.Q.; Chen, Y.Q. Relationship between Gut Microbiota and Phosphorus Metabolism in Hemodialysis Patients: A Preliminary Exploration. Chin. Med. J. 2018, 131, 2792–2799. [Google Scholar] [PubMed]
- Lau, W.L.; Savoj, J.; Nakata, M.B.; Vaziri, N.D. Altered microbiome in chronic kidney disease: Systemic effects of gut-derived uremic toxins. Clin. Sci. 2018, 132, 509–522. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Xiang, S.; Ye, K.; Zheng, Y.; Feng, X.; Zhu, X.; Chen, J.; Chen, Y. Cobalamin (Vitamin B12) induced a shift in microbial composition and metabolic activity in an in vitrocolon simulation. Front. Microbiol. 2018, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Liu, X.; Xiong, X.Q.; Yang, T.; Cui, T.; Hou, N.L.; Lai, X.; Liu, S.; Guo, M.; Liang, X.H.; et al. Effect of Vitamin A supplementation on gut microbiota in children with autism spectrum disorders—A pilot study. BMC Microbiol. 2017, 17, 204. [Google Scholar] [CrossRef] [PubMed]
- Sassi, F.; Tamone, C.; D’Amelio, P. Vitamin D: Nutrient, Hormone, and Immunomodulator. Nutrients 2018, 10, 1656. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.S.; Ko, I.K.; Yoo, J.J. Bioactive Compounds for the Treatment of Renal Disease. Yonsei Med. J. 2018, 59, 1015. [Google Scholar] [CrossRef] [PubMed]


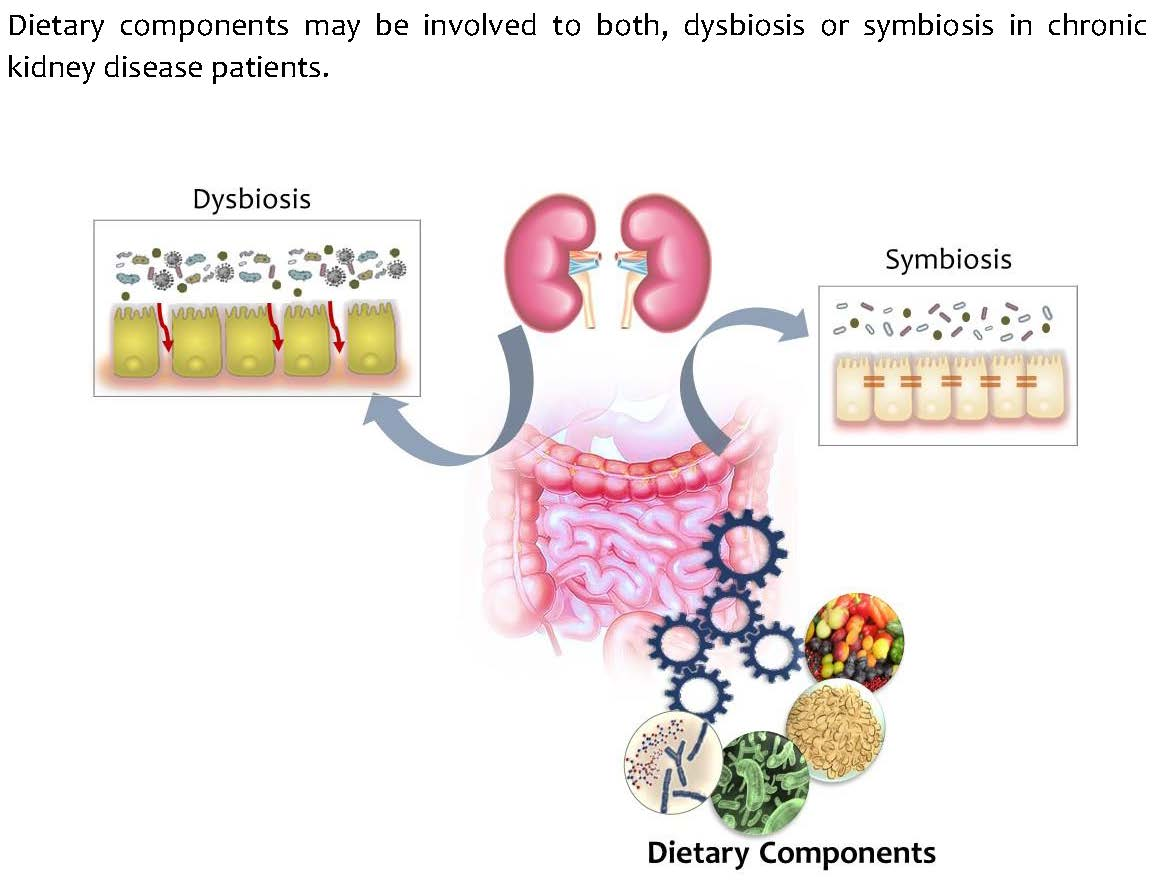{kind=link}
{kind=link}
| References | Study Design, Sample, Follow-up | Intervention | Results |
|---|---|---|---|
| Taki et al., 2005 [25] | Non-controlled trial; 27 HD patients; 3 months | 3 × 109 CFU/day of B. longum for 4 weeks, 6.0 × 109 CFU/day from 4th to 8th week, 12.0 × 109 CFU/day from 8th to 12th week. | ↓ IS, homocystein and triglyceride serum levels ↑ folate serum levels |
| Ranganathan et al., 2010 [12] | RCT with crossover, multicenter; 46 non-dialysis CKD patients; 6 months | 9 × 1010 CFU/day of a probiotic mix: S. thermophilus, L. acidophilus, and B. longum | ↓ urea serum levels Overall improvement in quality of life ↔ Cr and or uric acid |
| Alatriste et al., 2014 [23] | RCT; 30 non-dialysis CKD patients; 2 months | 8 × 109 CFU/day vs. 16 × 109 CFU/day of L. casei Shirota | ↓ serum urea – in dose of 16 × 109 CFU |
| Natarajan et al., 2014 [28] | RCT with crossover; 22 HD patients; 6 months | 1.8 × 1011 CFU/day of a probiotic mix: S. thermophilus, L. acidophilus, and B. longum | ↔ Uremic toxins or inflammatory markers. |
| Wang et al., 2015 [29] | Randomized, double-blind, placebo-controlled clinical trial; 39 peritoneal dialysis patients;6 months | 109 CFU/day of B. bifidum, 109 CFU/day of B. catenulatum, 109 CFU/day of B. longum, and 109 CFU/day of L. plantarum | ↓ serum TNF-α, IL-5, IL-6, and endotoxin ↑ serum IL-10 |
| Soleimani et al., 2016 [30] | RCT; 60 diabetic patients on HD; 3 months | 2 × 109 CFU. L. acidophilus, L. casei and B. bifidum | ↓ plasma glucose, serum insulin, HOMA-IR, HOMA-B, HbA1c, hs-CRP, MDA, SGA scores, TIBC ↑ plasma total antioxidant capacity ↔ lipid profiles, biomarkers of inflammation and oxidative stress |
| Shariaty et al., 2017 [31] | RCT; 34 HD patients; 3 months | 3 × 1010 CFU of L. acidophilus, 3 × 109 CFU of L. casei, 7 × 109 CFU of L. rhamnosus, 5 × 108 CFU of L. bulgaricus, 2 × 1010 CFU of B. breve, 1 × 109 CFU of B. longum, 3 × 108 CFU of S. thermophilus | ↔ Hb or CRP levels |
| Borges et al., 2017 [13] | RCT; 33 HD patients; 3 months | 9 × 1010 CFU/day of a probiotic mix: S. thermophilus, L. acidophilus, and B. longum | ↑ plasma IS, K, urea; ↓ fecal pH ↔ p-CS, IAA, inflammatory markers (CRP and IL-6) or gut microbiota profile |
| Barros et al., 2018 [24] | RCT; 22 non-dialysis CKD patients; 3 months | 9 × 1010 CFU/day of a probiotic mix: S. thermophilus, L. acidophilus, and B. longum | ↑ IL-6 plasma levels ↔ Uremic toxins (TMAO, IS, p-CS and IAA), CRP, LPS or calprotectin. |
| Eidi et al., 2018 [26] | RCT; 42 HD patients; 1 month | 1.6 × 107 CFU/day of L. Rhamnosus | ↓ phenol and p-cresol serum levels |
| References | Study Design, Sample, Follow-up | Prebiotics | Results |
|---|---|---|---|
| Younes et al, 2006 [58] | RCT with crossover; 9 non-dialysis CKD patients; 5 weeks | 40 g/day fermentable carbohydrate (25 g whole-meal bread + 4.5 g inulin + 10.5 g crude potato starch) | ↑ stool weight, fecal and urinary urea, fiber intake No change in eGFR |
| Meijers et al, 2010 [56] | Non-randomized, single-center, open-label phase; 22 HD patients; 4 weeks | 10–20 g/day of oligofructose-enriched inulin | ↓ 20% serum p-CS, generation rate and BUN ↔ IS serum or generation rates |
| Sirich et al, 2014 [52] | RCR; 56 HD patients; 6 weeks | 15 g/day of high-amylose corn starch | ↓ IS and a trend to p-CS free plasma ↔ body weight, CRP |
| Salmean et al, 2015 [53] | Single-blind, placebo controlled; 13 non-dialysis CKD patients; 6 weeks | 10 g/day of pea hull fiber (Best Pea Fiber; Best Cooking Pulses, Portage la Prairie, Manitoba, Canada) + 15 g/day inulin | ↓ 20% total plasma p-CS, ↑ stool frequency, fiber intake ↔ CRP, cystatin C, BUN, ammonia, eGFR |
| Poesen et al, 2016 [57] | RCT with crossover; 40 non-dialysis CKD patients; 4 weeks | 10 g twice/day of arabinoxylan oligosaccharide | ↓serum TMAO ↔ IS, p-Cs, p-Cglucuronide and phenylacetylglutamine, HOMA-IR |
| Tayebi-Khosroshahi et al, 2016 [51] | RCT; 32 non-dialysis CKD patients; 8 weeks | 30 mm thrice/day of lactulose syrup | ↑ fecal bifidobacteria and lactobacillus counts ↓ Cr |
| Tayebi Khosroshahi et al, 2018 [59] | RCT; 46 HD patients; 4 weeks | 20–25 g/day of high amylose maize resistant starch | ↓ TNF-α, IL-6, MDA, severity constipation, serum urea and creatinine ↔ IL-1β, hs-CRP, total antioxidant activity |
| Esgalhado et al, 2018 [16] | RCT; 31 HD patients; 4 weeks | 16 g/day resistant starch (Hi-Maize® 260) | ↓ IL-6, TBARS, IS and a trend to protein carbonylation; ↑ fiber intake ↔ CRP, p-CS |
| References | Study Design, Sample, Follow-up | Intervention | Results |
|---|---|---|---|
| Nakabayashi et al., 2011 [70] | Clinical trial; 9 HD patients; 2 weeks | 1 × 108 Lactobacillus casei strain Shirota and Bifidobacterium breve strain Yakult + 4 g of prebiotic: 1.67 g or more galacto-oligosaccharides and <1.36 g of lactose and monosaccharide | ↓ serum p-CS levels ↔ phenol or IS |
| Cruz-Mora et al., 2014 [65] | RCT; 18 HD patients; 2 months | (Lactobacillus acidophilus and Bifidobacterium bifidum), for total as probiotic of 2.0 × 1012 CFU 2.31 g of a prebiotic fiber (inulin); 1.5 g of omega-3 fatty acids (eicosapentaenoic and docosahexaenoic acid) and vitamins (complex B, folic acid, ascorbic acid, and vitamin E) | ↑ Bifidobacterium counts ↓Gastrointestinal symptoms scores |
| Guida et al, 2014 [69] | RCT; 30 non-dialyzed CKD patients; 1 month | 5 × 109 Lactobacillus plantarum, 2 × 109 Lactobacillus casei subsp. rhamnosus and 2 × 109 Lactobacillus gasseri, 1 × 109 Bifidobacterium infantis and 1 × 109 Bifidobacterium longum, 1 × 109 Lactobacillus acidophilus, 1 × 109 Lactobacillus salivarius and 1 × 109 Lactobacillus sporogenes and 5 × 109 Streptococcus thermophilus) + prebiotic: 2.2 g inulin and 1.3 g of tapioca-resistant starch | ↓ plasma p-CS |
| Viramontes-Horner et al, 2015 [66] | RCT; 42 HD patients; 2 months | Symbiotic gel (containing Lactobacillus acidophilus NCFM and Bifidobacterium lactis Bi-07 for a total of 11 × 106 CFU + 2.31 g of a prebiotic fiber inulin + 1.5 g of omega-3 fatty acids and vitamins of complex B, folic acid, ascorbic acid, and vitamin E) | ↓ Episodes of vomit, heart- burn, and stomachache, gastrointestinal symptons. No change in the prevalence of malnutrition, CRP and TNFα levels |
| Dehghani et al., 2016 [67] | RCT; 66 non-dialysis CKD patients; 6 weeks | 2 × 500 mg containing 7 strains of probiotics: Lactobacilus casei, Lactobacilus acidophilus, Lactobacilus bulgarigus, Lactobacilus rhamnosus, Bifidobacterium breve, Bifidobacterium longum, Sterptococus thermophilus + FOS | ↓ blood urea nitrogen ↔ Cr, uric acid |
| Pavan, 2016 [68] | Prospective observational study with randomized control, open-label design; 24 non-dialysis CKD patients; 6 months | 15 billion cells/cfU of each one: Streptococcus thermophiles, Lactobacilllus acidophilus, Bifidobacterium longum + 100 mg Fructooligosaccharides | ↓ GFR |
| Rossi et al, 2016 [17] | RCT with crossover; 31 non-dialyzed CKD patients; 6 weeks | High–molecular weight inulin, FOS and GOS and the probiotic component including 9 different strains across the Lactobacillus, Bifidobacteria, and Streptococcus genera | ↑Bifidobacterium and ↓ Ruminococcaceae ↓ p-CS ↑albuminuria by 38 mg/24 h ↔ IS, eGFR, IL-1b, IL-6, IL-10, TNF-a, serum oxidative stress biomarkers (F2- isoprostanes and glutathione peroxidase), and LPS |
| References | Study Design, Sample, Follow-up | Intervention | Results |
|---|---|---|---|
| Clavel et al. (2005) [84] | RCT; 39 postmenopausal women; 1 month | 100 mg/day of isoflavones supplemented in cereal bars and gelified milk | ↑ Lactobacillus-Enterococcus ↑ Faecalibacterium prausnitzii ↑ Bifidobacterium ↔ Atopobium ↔ Bacteroides |
| Queipo-Ortuno et al. (2012) [85] | RCT;10 healthy male volunteers; 20 days | Group 1: de-alcoholized red wine (272 mL/day) Group 2: red wine (272 mL/day) Group 3: gin (100 mL/day) | Group 1: ↑ Fusobacteria ↓ Bacteroidetes and Firmicutes Group 2: ↑ Proteobacteria, Fusobacteria, Firmicutes, Bacteroidetes, Bacteroides, Prevotella and the B. uniformis. Group 3: ↑ Clostridium and the Clostridium histolyticum; ↓ Prevotella; ↓ CRP; total cholesterol |
| Song et al. (2015) [86] | RCT; 28 obese women; 3 months | 2 pouches in a day, equivalent of 6.7 g dried Schisandra chinensis fruit | ↑ Akkermansia, Roseburia, Bacteroides, Prevotella, and Bifidobacterium ↓ Ruminococcus ↓ blood glucose, triglycerides |
| Eid et al. (2015) [87] | RCT; 21 healthy volunteers; 21 days | 50 g of palm date | ↔ growth of the faecal microbiota |
| Moreno-Indias et al. (2015) [88] | RCT; 10 metabolic syndrome in obese patients; 1 month | red wine (272 mL per day) or de-alcoholized red wine (272 mL per day) | Red wine and de-alcoholized red wine: ↓ Clostridium and the Clostridium histolyticum ↑ Blautia coccoides–Eubacterium rectale, Faecalibacterium prausnitzii, Roseburia and Lactobacillus ↓ LPS |
| Janssens et al. (2016) [89] | RCT; 58 Caucasian men and women; 3 months | green tea (>0.56 g/d epigallocatechin-gallate + 0.28*0.45 g/d caffeine) capsules | ↔ growth of the faecal microbiota |
| Li et al. (2015) [75] | RCT; 20 healthy participants; 1 month | 1g of pomegranate extract daily | ↑ Actinobacteria, Butyrivibrio, Enterobacter, Escherichia, Lactobacillus, Prevotella, Serratia and Veillonella. ↓ Firmicutes and Collinsell |
| Barroso et al. (2017) [90] | RCT; 15 healthy volunteers; 28 days | 250 mL of red wine per day | ↑ Slackia, Gordonibacter, Oscillatoria, Veillonella and Oenococcus |
| Most et al. (2017) [91] | RCT; 37 overweight and obese men and women; 3 months | epigallocatechin-3-gallate (282 mg/day) and resveratrol (80 mg/day) | ↓ Bacteroidetes and Faecalibacterium prausnitzii in men ↔ Bacteroidetes and Faecalibacterium prausnitzii in women |
| Peterson et al. (2018) [92] | RCT; 14 healthy volunteers; 2 months | Group 1: turmeric tablets contained 1000 mg turmeric root (Curcuma longa) plus 1.25 mg black pepper–derived extract of piperine alkaloid. Group 2: curcumin tablets contained 1000 mg of curcumin (Curcumin C3 Complex) plus 1.25 mg black pepper. | Group 1: ↑ in observed species 7% (156 vs. 167) ↓ Eisenbergiella tayi; ↑ Alistipes putredinis Group 2: ↑ 69% (127 vs. 215) in detected species ↓ Coprococcus catus; ↑ Raoultella electrica; (Clostridium) xylanolyticum; Collinsella aerofaciens; Kluyvera intermedia |
| References | Study Design, Sample, Follow-up | Intervention | Results |
|---|---|---|---|
| CKD patients | |||
| Marzocco et al., 2013 [95] | RCT with crossover; 32 non-dialysis CKD patients; 1 week | VLPD (0.3 g/kg bw/day) + ketoanalogues LPD (0.6 g/kg bw/day) | VLPD changed the IS level with a reduction of 37% when compared to LPD |
| Kandouz et al., 2016 [99] | Cross-sectional; 138 HD patients from a cohort were analyzed and 16 patients were strict vegetarians | Vegetarian diet | ↓ IS and p-CS levels; ↓ serum urea, and phosphate and estimated urea nitrogen intake before HD |
| Black et al., 2018 [20] | Longitudinal; 30 non-dialysis CKD patients; 6 months | LPD (0.6 g/kg/day) | ↓ p-CS plasma levels Change in the intestinal microbiota profile ↔ IS, IAA |
| Mafra et al, 2018 [101] | Prospective pilot study; 9 non-dialysis CKD patients; 6 months | LPD (0.6 g protein/kg day) | ↓ TMAO plasma levels |
| Patel et al., 2012 [100] | 15 healthy vegetarian individuals 11 health individuals – normal diet | Vegetarian diet | ↓ p-CS and IS production rates |
| Non CKD patients | |||
| De Filippis et al, 2016 [108] | Cross-sectional survey; 51 vegetarians, 51 vegans, 51 omnivores | Adherence to the Mediterranean diet | Associations between consumption of vegetable-based diets and higher levels of short-chain fecal fatty acids, Prevotella and fiber-degrading Firmicutes; ↓ urinary TMAO levels in vegetarian and vegan diet |
| Mitsou et al, 2017 [109] | Cross-sectional study, 120 healthy participants | Adherence to the Mediterranean diet | ↓ Escherichia Coli counts, ↑ Bifidobacteria |
| Garcia-Mantrana et al, 2018 [96] | Cross-sectional study; 27 healthy volunteers | Adherence to the Mediterranean diet | ↑ Bifidobacterial counts, ↑ concentration of acetate, propionate, and butyrate in fecal samples, ↑ Bacteroidetes and a lower Firmicutes–Bacteroidetes ratio |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mafra, D.; Borges, N.; Alvarenga, L.; Esgalhado, M.; Cardozo, L.; Lindholm, B.; Stenvinkel, P. Dietary Components That May Influence the Disturbed Gut Microbiota in Chronic Kidney Disease. Nutrients 2019, 11, 496. https://doi.org/10.3390/nu11030496
Mafra D, Borges N, Alvarenga L, Esgalhado M, Cardozo L, Lindholm B, Stenvinkel P. Dietary Components That May Influence the Disturbed Gut Microbiota in Chronic Kidney Disease. Nutrients. 2019; 11(3):496. https://doi.org/10.3390/nu11030496
Chicago/Turabian StyleMafra, Denise, Natália Borges, Livia Alvarenga, Marta Esgalhado, Ludmila Cardozo, Bengt Lindholm, and Peter Stenvinkel. 2019. "Dietary Components That May Influence the Disturbed Gut Microbiota in Chronic Kidney Disease" Nutrients 11, no. 3: 496. https://doi.org/10.3390/nu11030496
APA StyleMafra, D., Borges, N., Alvarenga, L., Esgalhado, M., Cardozo, L., Lindholm, B., & Stenvinkel, P. (2019). Dietary Components That May Influence the Disturbed Gut Microbiota in Chronic Kidney Disease. Nutrients, 11(3), 496. https://doi.org/10.3390/nu11030496