The Effect of Macronutrients on Reproductive Hormones in Overweight and Obese Men: A Pilot Study


Abstract
1. Introduction
2. Materials and Methods
2.1. Cohort
2.2. Anthropometric Measures
2.3. Habitual Behaviour Patterns
2.3.1. Dietary Intake Data
2.3.2. Habitual Sleep and Exercise Patterns
2.4. Intervention Protocol
2.4.1. Baseline (Fasting) Protocol
2.4.2. Feeding Protocol
2.5. Statistical Analysis
3. Results
3.1. Subjects
3.2. Hormone Analysis
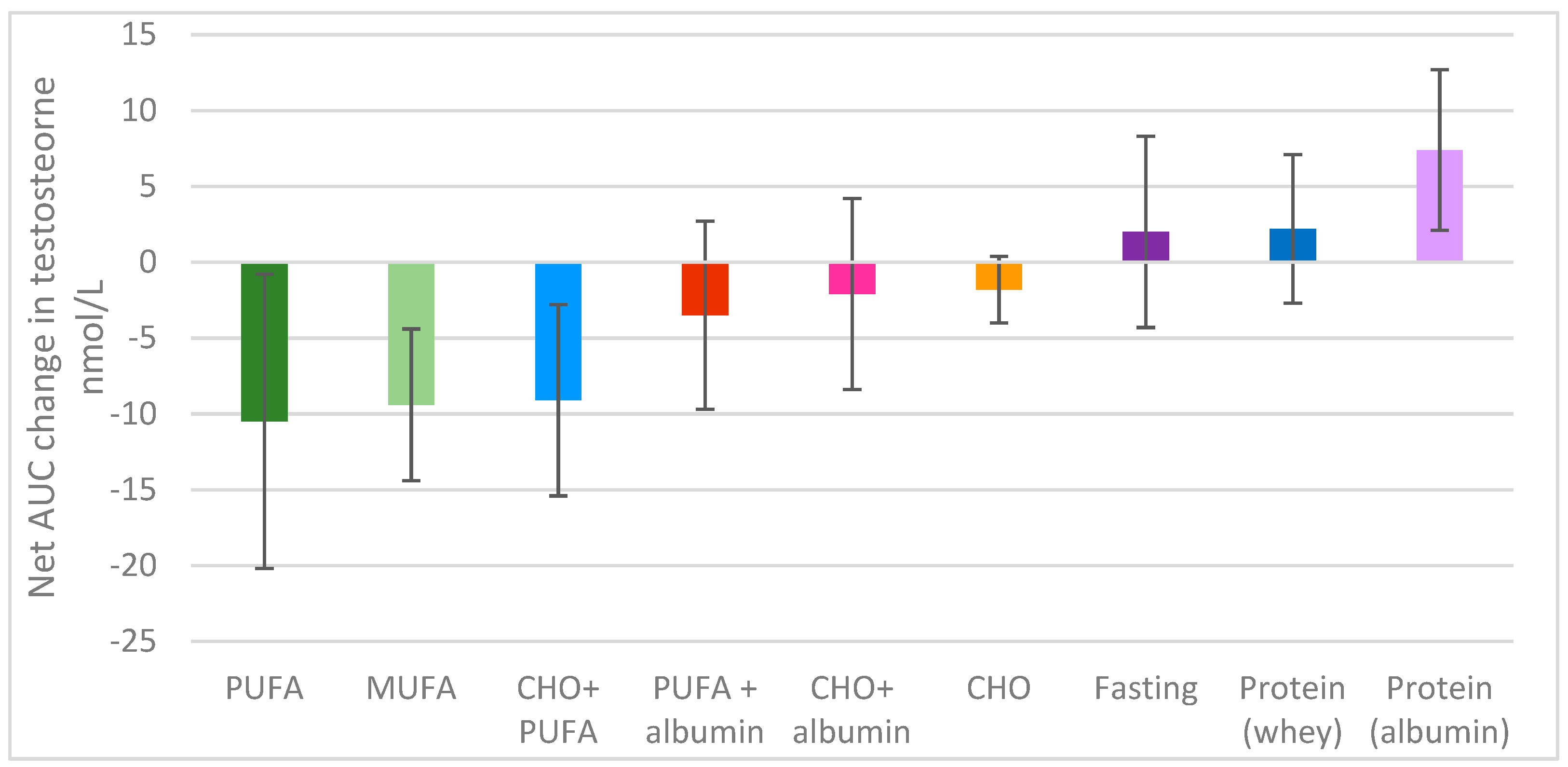
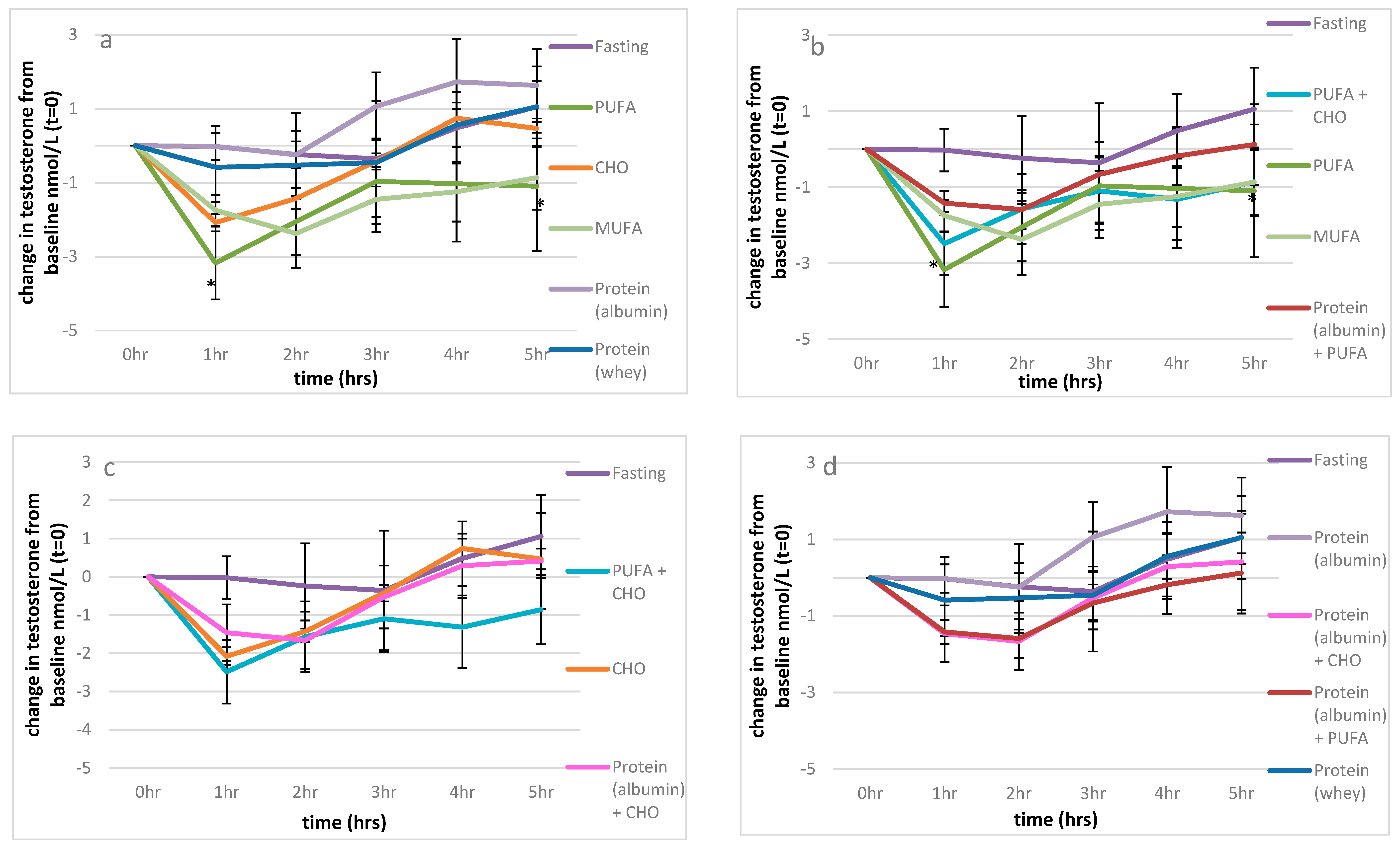
3.2.1. Testosterone
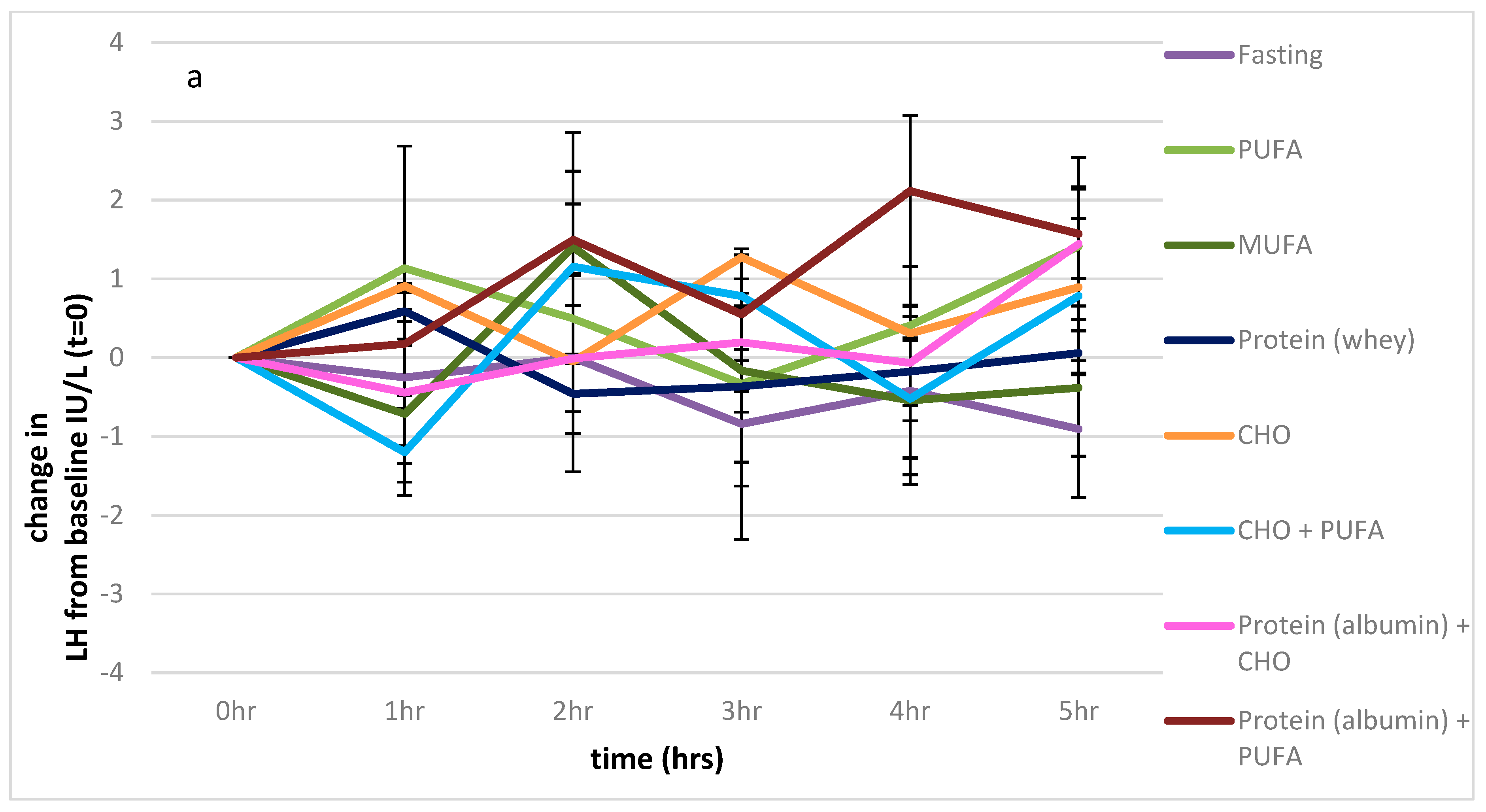
3.2.2. LH
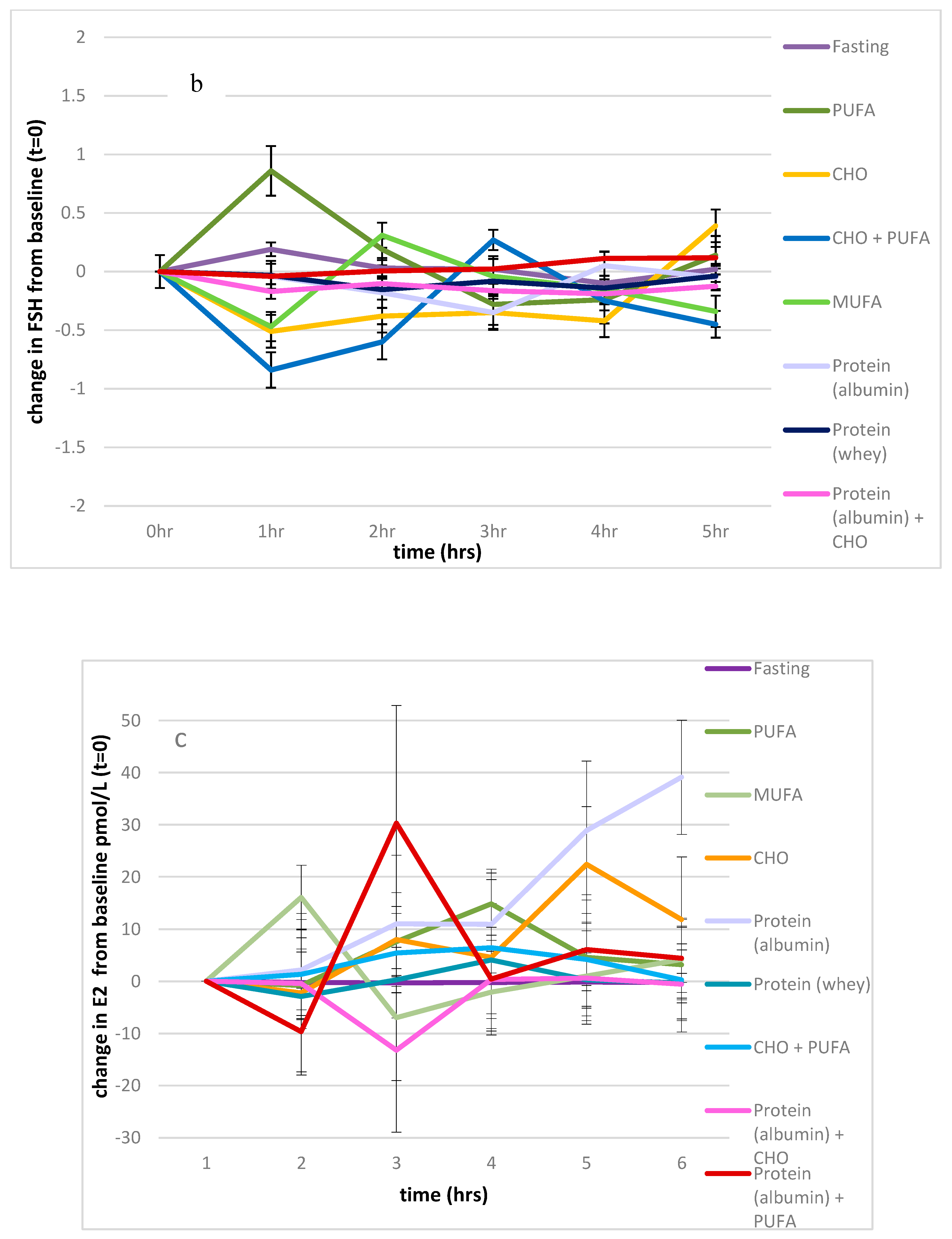
3.2.3. FSH
3.2.4. Oestradiol
3.3. Habitual Dietary Intake Data
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organisation: Obesity and Overweight. 2018. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 12 April 2019).
- Palmer, N.O.; Bakos, H.W.; Fullston, T.; Lane, M. Impact of obesity on male fertility, sperm function and molecular composition. Spermatogenesis 2012, 2, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Australian Institute of Health and Welfare: A Picture of Overweight and Obesity in Australia 2019. Available online: https://www.aihw.gov.au/getmedia/172fba28-785e-4a08-ab37-2da3bbae40b8/aihw-phe-216.pdf.aspx?inline=true (accessed on 20 August 2019).
- De Lorenzo, A.; Noce, A.; Moriconi, E.; Rampello, T.; Marrone, G.; Di Daniele, N.; Rovella, V. MOSH Syndrome (Male Obesity Secondary Hypogonadism): Clinical Assessment and Possible Therapeutic Approaches. Nutrients 2018, 10, 474. [Google Scholar] [CrossRef] [PubMed]
- Allen, N.E.; Key, J.K. The effects of diet on circulating sex hormone levels in men. Nutr. Res. Rev. 2007, 13, 159–184. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.-Y.; Chen, Y.C.; Lin, P.; Shih, C.-K.; Bai, C.-H.; Yuan, K.-C.; Lee, S.-Y.; Chang, J.-S. Testosterone-Associated Dietary Pattern Predicts Low Testosterone Levels and Hypogonadism. Nutrients 2018, 10, 1786. [Google Scholar] [CrossRef] [PubMed]
- Ghanim, H.; Abuaysheh, S.; Sia, C.L.; Korzeniewski, K.; Chaudhuri, A.; Fernandez-Real, J.M.; Dandona, P. Diminished androgen and estrogen receptors and aromatase levels in hypogonadal diabetic men: Reversal with testosterone. Diabetes Care 2018, 178, 277. [Google Scholar] [CrossRef] [PubMed]
- Grossmann, M. Hypogonadism and male obesity: Focus on unresolved questions. Clin. Endocrinol. 2018, 89, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Hales, D. Testicular macrophage modulation of Leydig cell steroidogenesis. J. Reprod. Immunol. 2002, 57, 3–18. [Google Scholar] [CrossRef]
- Tremellen, K.; Mcphee, N.; Pearce, K. Metabolic endotoxaemia related inflammation is associated with hypogonadism in overweight men. Basic Clin. Androl. 2017, 27, 5. [Google Scholar] [CrossRef]
- Tremellen, K.; McPhee, N.; Pearce, K.; Benson, S.; Schedlowski, M.; Engler, H. Endotoxin-initiated inflammation reduces testosterone production in men of reproductive age. Am. J. Physiol.-Endoc. Metab. 2018, 314, E206–E213. [Google Scholar] [CrossRef]
- Gregersen, S.; Samocha-Bonet, D.; Heilbronn, L.K.; Campbell, L.V. Inflammatory and Oxidative Stress Responses to High-Carbohydrate and High-Fat Meals in Healthy Humans. J. Nutr. Metab. 2012, 2012, 238056. [Google Scholar] [CrossRef]
- Erridge, C.; Attina, T.; Spickett, C.M.; Webb, D.J. A high-fat meal induces low-grade endotoxemia: Evidence of a novel mechanism of postprandial inflammation. Am. J. Clin. Nutr. 2007, 86, 1286–1292. [Google Scholar] [CrossRef] [PubMed]
- Eslamian, G.; Amirjannati, N.; Rashidkhani, B.; Sadeghi, M.-R.; Hekmatdoost, A. Intake of food groups and idiopathic asthenozoospermia: A case–control study. Hum. Reprod. 2012, 27, 3328–3336. [Google Scholar] [CrossRef] [PubMed]
- Mendiola, J.; Torres-Cantero, A.M.; Grau, J.; Ten, J.; Bernabeu, R. Food intake and its relationship with semen quality: A case-control study. Fertil. Steril. 2009, 91, 812–818. [Google Scholar] [CrossRef] [PubMed]
- Vujkovic, M.; de Vries, J.H.; Dohle, G.R.; Bonsel, G.J.; Lindemans, J.; Macklon, N.S.; van der Spek, P.J.; Steegers, E.A.P.; Steegers-Theunissen, R.P.M. Associations between dietary patterns and semen quality in men undergoing IVF/ICSI treatment. Hum. Reprod. 2009, 24, 1304–1312. [Google Scholar] [CrossRef]
- Gaskins, A.J.; Afeiche, M.C.; Hauser, R.; Williams, P.L.; Gillman, M.W.; Tanrikut, C.; Petrozza, J.C.; Chavarro, J.E. Paternal physical and sedentary activities in relation to semen quality and reproductive outcomes among couples from a fertility center. Hum. Reprod. 2014, 29, 2575–2582. [Google Scholar] [CrossRef]
- Karayiannis, D.; Kontogianni, M.D.; Mendorou, C.; Douka, L.; Mastrominas, M.; Yiannakouris, N. Association between adherence to the Mediterranean diet and semen quality parameters in male partners of couples attempting fertility. Hum. Reprod. 2016, 32, 215–222. [Google Scholar] [CrossRef]
- Jurewicz, J.; Radwan, M.; Sobala, W.; Radwan, P.; Bochenek, M.; Hanke, W. Dietary Patterns and Their Relationship with Semen Quality. Am. J. Men’s Health 2018, 12, 575–583. [Google Scholar] [CrossRef]
- Attaman, J.A.; Toth Thomas, L.; Furtado, J.; Campos, H.; Hauser, R.; Chavarro, J.E. Dietary fat and semen quality among men attending a fertility clinic. Hum. Reprod. 2012, 27, 1466–1474. [Google Scholar] [CrossRef]
- Jensen, T.K.; Heitmann, B.L.; Jensen, M.B.; Halldorsson, T.I.; Andersson, A.-M.; Skakkebæk, N.E.; Joensen, U.N.; Lauritsen, M.P.; Christiansen, P.; Dalgård, C.; et al. High dietary intake of saturated fat is associated with reduced semen quality among 701 young Danish men from the general population. Am. J. Clin. Nutr. 2012, 97, 411–418. [Google Scholar] [CrossRef]
- Chiu, Y.H.; Afeiche, M.C.; Gaskins, A.J.; Williams, P.L.; Mendiola, J.; Jørgensen, N.; Swan, S.H.; Chavarro, J.E. Sugar-sweetened beverage intake in relation to semen quality and reproductive hormone levels in young men. Hum. Reprod. 2014, 29, 1575–1584. [Google Scholar] [CrossRef]
- Volek, J.S.; Gmez, A.L.; Love, D.M.; Avery, N.G.; Sharman, M.J.; Kraemer, W.J. Effects of a high-fat diet on postabsorptive and postprandial testosterone responses to a fat-rich meal. Metabolism 2001, 50, 1351–1355. [Google Scholar] [CrossRef] [PubMed]
- Habito, R.C.; Montalto, J.; Leslie, E.; Ball, M.J. Effects of replacing meat with soyabean in the diet on sex hormone concentrations in healthy adult males. Br. J. Nutr. 2000, 84, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Meikle, A.W.; Benson, S.J.; Liu, X.H.; Boam, W.D.; Stringham, J.D. Nonesterified fatty acids modulate steroidogenesis in mouse Leydig cells. Am. J. Physiol. -Endocrinol. Metab. 1989, 257, E937–E942. [Google Scholar] [CrossRef] [PubMed]
- World Health Organisation. BMI Classifications. 2017. Available online: http://apps.who.int/bmi/index.jsp?introPage=intro_3.html (accessed on 15 June 2019).
- Black, A.E. Critical evaluation of energy intake using the Goldberg cut-off for energy intake:basal metabolic rate. A practical guide to its calculation, use and limitations. Int. J. Obes. 2000, 24, 1119. [Google Scholar] [CrossRef]
- Buysse, D.J.; Reynolds, C.F., III; Monk, T.H.; Berman, S.R.; Kupfer, D.J. The Pittsburgh sleep quality index: A new instrument for psychiatric practice and research. Psychiatry Res. 1989, 28, 193–213. [Google Scholar] [CrossRef]
- Baecke, J.A.; Burema, J.; Frijters, J.E. A short questionnaire for the measurement of habitual physical activity in epidemiological studies. Am. J. Clin. Nutr. 1982, 36, 936–942. [Google Scholar] [CrossRef]
- Tremellen, K.; Hill, A.; Pearce, K. Mechanistic insights into the aetiology of post-prandial decline in testosterone in reproductive-aged men. Andrologia 2019, e13418. [Google Scholar] [CrossRef]
- Martínez-González Miguel, A.; Gea, A.; Ruiz-Canela, M. The Mediterranean Diet and Cardiovascular Health. Circ. Res. 2019, 124, 779–798. [Google Scholar] [CrossRef]
- Iranmanesh, A.; Lawson, D.; Veldhuis, J.D. Glucose ingestion acutely lowers pulsatile LH and basal testosterone secretion in men. Am. J. Physiol.-Endoc. Metab. 2012, 302, E724–E730. [Google Scholar] [CrossRef]
- Caronia, L.M.; Dwyer, A.A.; Hayden, D.; Amati, F.; Pitteloud, N.; Hayes, F.J. Abrupt decrease in serum testosterone levels after an oral glucose load in men: Implications for screening for hypogonadism. Clin. Endocrinol. 2013, 78, 291–296. [Google Scholar] [CrossRef]
- Guo, Y.; Wang, A.; Liu, X.; Li, E. Effects of resveratrol on reducing spermatogenic dysfunction caused by high-intensity exercise. Reprod. Biol. Endocrinol. 2019, 17, 42. [Google Scholar] [CrossRef] [PubMed]
- Cutillas-Tolín, A.; Mínguez-Alarcón, L.; Mendiola, J.; López-Espín, J.J.; Jørgensen, N.; Navarrete-Muñoz, E.M.; Torres-Cantero, A.M.; Chavarro, J.E. Mediterranean and western dietary patterns are related to markers of testicular function among healthy men. Hum. Reprod. 2015, 30, 2945–2955. [Google Scholar] [CrossRef] [PubMed]
- Fantus, R.J.; Halpern, J.A.; Chang, C.; Keeter, M.K.; Bennett, N.E.; Helfand, B.; Brannigan, R.E. The association between popular diets and serum testosterone among men in the United States. J. Urol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Rajpathak, S.N.; Gunter, M.J.; Wylie-Rosett, J.; Ho, G.Y.F.; Kaplan, R.C.; Muzumdar, R.; Rohan, T.E.; Strickler, H.D. The role of insulin-like growth factor-I and its binding proteins in glucose homeostasis and type 2 diabetes. Diabetes-Metab. Res. Rev. 2009, 25, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Yoshizawa, A.; Clemmons, D.R. Testosterone and Insulin-like Growth Factor (IGF) I Interact in Controlling IGF-Binding Protein Production in Androgen-Responsive Foreskin Fibroblasts1. J. Clin. Endocrinol. Metab. 2000, 85, 1627–1633. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Corona, G.; Rastrelli, G.; Monami, M.; Melani, C.; Balzi, D.; Sforza, A.; Forti, G.; Mannucci, E.; Maggi, M. Body Mass Index Regulates Hypogonadism-Associated CV Risk: Results from a Cohort of Subjects with Erectile Dysfunction. J. Sex. Med. 2011, 8, 2098–2105. [Google Scholar] [CrossRef] [PubMed]
- Habito, R.C.; Ball, M.J. Postprandial changes in sex hormones after meals of different composition. Metabolism 2001, 50, 505–511. [Google Scholar] [CrossRef]
- Wood, R.I.; Stanton, S.J. Testosterone and sport: Current perspectives. Horm. Behav. 2012, 61, 147–155. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CHO | Protein (Albumin) | Protein (Whey) | PUFA | MUFA | CHO + Albumin | CHO + PUFA | PUFA + Albumin | |
|---|---|---|---|---|---|---|---|---|
| Total energy (Kj) | 1800 | 847 | 739 | 1930 | 1872 | 2647 | 3730 | 847 |
| Total fat (g) | <1 | <1 | <1 | 52 | 51 | <1 | 52 | <1 |
| Saturated fat (g) | <1 | <1 | <1 | 8 | 8 | <1 | 8 | <1 |
| Polyunsaturated fat (g) | <1 | <1 | <1 | 31 | 4 | <1 | 31 | <1 |
| Monounsaturated fat (g) | <1 | <1 | <1 | 10 | 36 | <1 | 10 | <1 |
| trans Fatty acids (g) | <1 | <1 | <1 | 1 | 0 | <1 | 1 | <1 |
| Omega 3 fatty acid (g) | <1 | <1 | <1 | 4 | 0 | <1 | 4 | <1 |
| Omega 6 fatty acid (g) | <1 | <1 | <1 | 28 | 4 | <1 | 28 | <1 |
| Protein (g) | <1 | 41 | 41 | 1 | 0 | 41 | 1 | 41 |
| Total carbohydrate (g) | 100 | <1 | <1 | 6 | 0 | 100 | 100 | <1 |
| Sugar (g) | 76 | <1 | <1 | 0 | 0 | 76 | 76 | <1 |
| Fibre (g) | <1 | <1 | <1 | 0 | 0 | <1 | <1 | <1 |
| Sodium (mg) | <5 | 3 | 102 | 5 | 0 | 3 | 5 | 3 |
| Fasting | CHO | Protein (Albumin) | Protein (Whey) | PUFA | MUFA | CHO + Albumin | CHO + PUFA | PUFA + Albumin | p Value | |
|---|---|---|---|---|---|---|---|---|---|---|
| Age (yrs) | 37 ± 12.5 | 33.0 ± 11.0 | 33.6 ± 11.1 | 37.0 + 12 | 32.6 ± 11.1 | 33.0 ± 10.6 | 37.0 + 12 | 37.0 + 12 | 37.0 + 12 | 0.913 |
| BMI kg/m2 | 32.3 ± 8.2 | 31.3 ± 5.2 | 32.3 ± 4.6 | 30.7 + 5.1 | 30.3 ± 5.8 | 31.3 ± 5.2 | 30.7 + 5.1 | 30.7 + 5.1 | 30.7 + 5.1 | 0.939 |
| Sleep | 6.4 ± 2.3 | 5.8 ± 3.1 | 5.7 ± 3.4 | 6.6 ± 2.4 | 5.4 ± 3.2 | 5.4 ± 3.2 | 6.6 ± 2.4 | 6.6 ± 2.4 | 6.6 ± 2.4 | 0.998 |
| Physical activity | 5.5 + 2.1 | 5.2 ± 6.1 | 8.0 ± 6.1 | 5.8 ± 2.1 | 5.2 ± 1.9 | 5.2 ± 6.2 | 5.8 ± 2.1 | 5.8 ± 2.1 | 5.8 ± 2.1 | 0.782 |
| Dietary intake | ||||||||||
| Energy (MJ/d) | 10.2 ± 3.2 | 9.7 ± 3.2 | 9.7 ± 3.2 | 11.3 + 3.2 | 9.7 ± 3.2 | 9.9 ± 3.2 | 11.3 + 3.2 | 11.3 + 3.2 | 11.3 + 3.2 | 0.921 |
| Total fat (g) | 85 ± 15 | 79 ± 24 | 84 ± 23 + 24 | 84 + 13 | 95 ± 33 | 79 ± 24 | 84 + 13 | 84 + 13 | 84 + 13 | 0.134 |
| Saturated fat (g) | 29.2 ± 8.0 | 26.6 ± 11.4 | 26.6 + 11 | 29 ± 7 | 27 ± 11 | 28 ± 11 | 29 ± 7 | 29 ± 7 | 29 ± 7 | 0.203 |
| Total protein (g) | 92.3 ± 20.2 | 103 ± 39 | 103 + 39 | 93 ± 18 | 115 + 36 | 100 + 37 | 93 ± 18 | 93 ± 18 | 93 ± 18 | 0.222 |
| Total carbohydrates (g) | 329 + 108 | 291 ± 148 | 290 ± 147 | 317 ± 123 | 313 ± 134 | 294 ± 149 | 317 ± 123 | 317 ± 123 | 317 ± 123 | 0.965 |
| Sugar (g) | 143 ± 101 | 143 ± 157 | 141 ± 158 | 142 ± 87 | 143 ± 128 | 140 + 145 | 142 ± 87 | 142 ± 87 | 142 ± 87 | 0.864 |
| Fibre (g) | 23.6 ± 8.1 | 19 ± 9 | 19 ± 9 | 23.4 ± 7.3 | 26 ± 15 | 19 ± 10 | 23.4 ± 7.3 | 23.4 ± 7.3 | 23.4 ± 7.3 | 0.614 |
| Fasting | CHO | Protein (Albumin) | Protein (Whey) | PUFA | MUFA | CHO+ Albumin | CHO + PUFA | PUFA + Albumin | p Value | |
|---|---|---|---|---|---|---|---|---|---|---|
| Testosterone (nmol/L) | 11.2 ± 3.6 | 13.3 ± 3.1 | 12.8 ± 2.7 | 10.8 ± 3.3 | 15.5 ± 5.4 | 12.3 ± 2.5 | 11.0 ± 4.0 | 10.7 ± 2.5 | 10.7 ± 2.5 | 0.018 |
| LH (IU/L) | 5.5 ± 1.5 | 4.7 ± 1.7 | 5.4 ± 1.9 | 5.8 ± 2.6 | 5.0 ± 1.8 | 5.5 ± 2.4 | 5.0 ± 2.2 | 5.2 ± 2.1 | 4.7 ± 1.3 | 0.617 |
| Testosterone/LH | 2.2 ± 0.8 | 3.4 ± 2.1 | 2.6 ± 1.0 | 2.1 ± 0.8 | 3.3 ± 1.2 | 2.5 ± 0.9 | 2.5 ± 0.8 | 3.1 ± 1.7 | 2.2 ± 0.3 | 0.361 |
| Oestradiol (pmol/L) | 123 ± 17 | 125 ± 48 | 94 ± 28 | 48 ± 41 | 110 ± 30 | 104 ± 29 | 117 ± 36 | 123 ± 30 | 114 ± 43 | 0.007 |
| FSH (IU/L) | 3.2 ± 1.5 | 4.2 ± 1.7 | 3.9 ± 1.6 | 3.5 ± 1.4 | 4.5 ± 2.1 | 3.8 ± 2.0 | 2.7 ± 0.6 | 4.4 ± 1.9 | 3.3+1.5 | 0.001 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pearce, K.L.; Tremellen, K. The Effect of Macronutrients on Reproductive Hormones in Overweight and Obese Men: A Pilot Study. Nutrients 2019, 11, 3059. https://doi.org/10.3390/nu11123059
Pearce KL, Tremellen K. The Effect of Macronutrients on Reproductive Hormones in Overweight and Obese Men: A Pilot Study. Nutrients. 2019; 11(12):3059. https://doi.org/10.3390/nu11123059
Chicago/Turabian StylePearce, Karma L., and Kelton Tremellen. 2019. "The Effect of Macronutrients on Reproductive Hormones in Overweight and Obese Men: A Pilot Study" Nutrients 11, no. 12: 3059. https://doi.org/10.3390/nu11123059
APA StylePearce, K. L., & Tremellen, K. (2019). The Effect of Macronutrients on Reproductive Hormones in Overweight and Obese Men: A Pilot Study. Nutrients, 11(12), 3059. https://doi.org/10.3390/nu11123059