Regulation of Skeletal Muscle Glucose Transport and Glucose Metabolism by Exercise Training

 ,
,
Abstract
1. Introduction
2. Models of Aerobic and Resistance Exercise Training
3. Aerobic and Resistance Exercise Training-Induced Adaptations
4. Regulation of Systemic Glucose Homeostasis
4.1. Regulation in Healthy Individuals
4.2. Dysregulation in Type 2 Diabetes
4.3. Effects of Aerobic Training in Type 2 Diabetes
4.4. Effects of Resistance Training in Type 2 Diabetes
5. Skeletal Muscle Glucose Transport
5.1. Regulation of Basal Glucose Transport
5.2. Regulation of Acute Insulin—And Exercise/Contraction-Stimulated Glucose Transport
5.3. Dysregulation of Insulin-Stimulated Glucose Transport in Type 2 Diabetes
5.4. Regulation of Aerobic Exercise Training-Induced Glucose Transport
5.5. Regulation of Resistance Exercise Training-Induced Glucose Transport
6. Skeletal Muscle Glucose Metabolism
6.1. Hexokinase
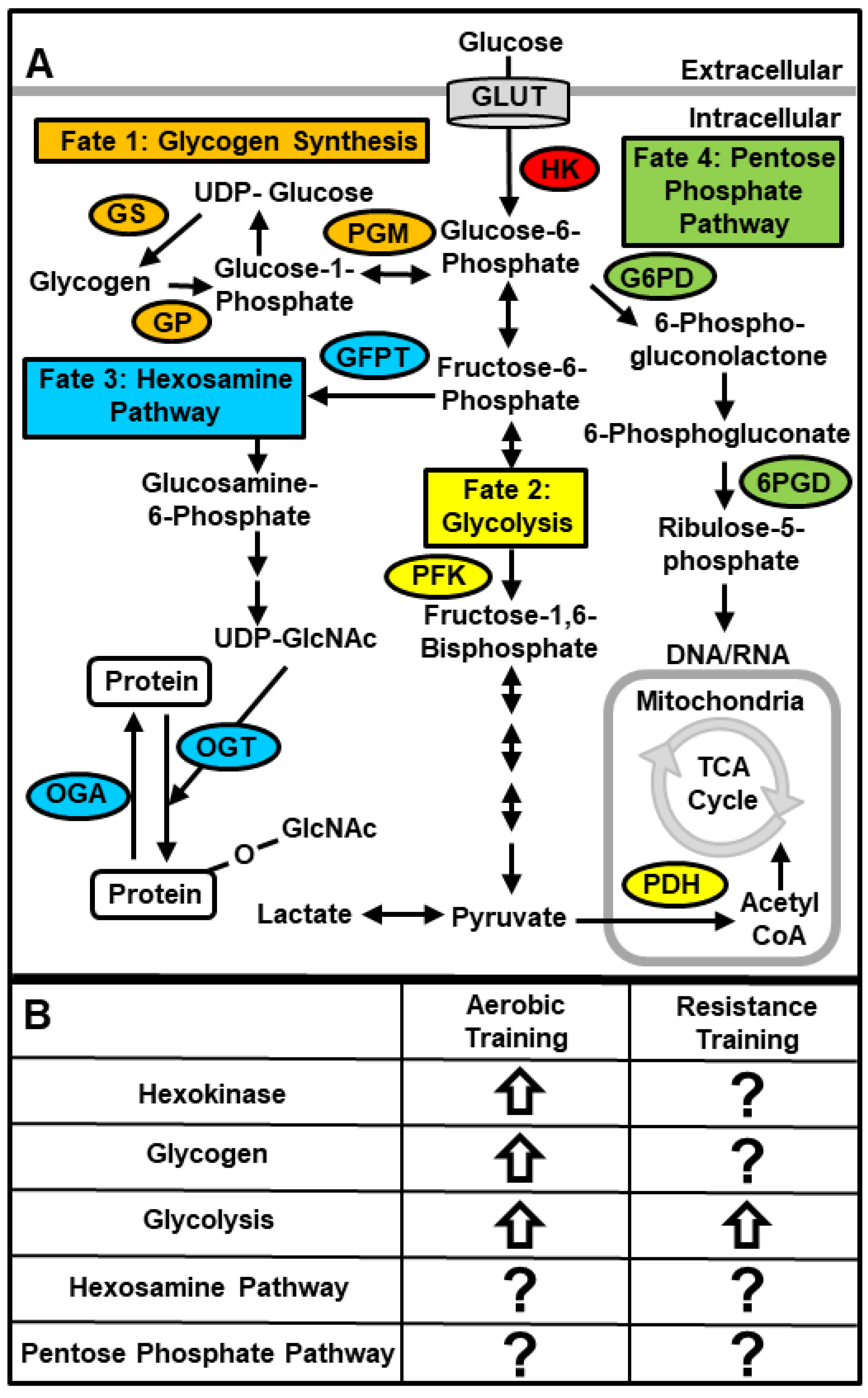
6.2. Cellular Fates of Glucose in Skeletal Muscle
6.2.1. Glycogen
6.2.2. Glycolytic Flux
6.2.3. Hexosamine Pathway
6.2.4. Pentose Phosphate Pathway
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Church, T.S.; Blair, S.N.; Cocreham, S.; Johannsen, N.; Johnson, W.; Kramer, K.; Mikus, C.R.; Myers, V.; Nauta, M.; Rodarte, R.Q.; et al. Effects of Aerobic and Resistance Training on Hemoglobin A1c Levels in Patients with Type 2 Diabetes: A Randomized Controlled Trial. JAMA 2010, 304, 2253–2262. [Google Scholar] [CrossRef] [PubMed]
- Jorge, M.L.M.P.; de Oliveira, V.N.; Resende, N.M.; Paraiso, L.F.; Calixto, A.; Diniz, A.L.D.; Resende, E.S.; Ropelle, E.R.; Carvalheira, J.B.; Espindola, F.S.; et al. The effects of aerobic, resistance, and combined exercise on metabolic control, inflammatory markers, adipocytokines, and muscle insulin signaling in patients with type 2 diabetes mellitus. Metabolism 2011, 60, 1244–1252. [Google Scholar] [CrossRef] [PubMed]
- Karstoft, K.; Winding, K.; Knudsen, S.H.; Nielsen, J.S.; Thomsen, C.; Pedersen, B.K.; Solomon, T.P.J. The Effects of Free-Living Interval-Walking Training on Glycemic Control, Body Composition, and Physical Fitness in Type 2 Diabetic Patients: A randomized, controlled trial. Diabetes Care 2013, 36, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Sigal, R.J.; Kenny, G.P.; Boulé, N.G.; Wells, G.A.; Prud’homme, D.; Fortier, M.; Reid, R.D.; Tulloch, H.; Coyle, D.; Phillips, P.; et al. Effects of Aerobic Training, Resistance Training, or Both on Glycemic Control in Type 2 Diabetes: A Randomized TrialEffects of Aerobic and Resistance Training on Glycemic Control in Type 2 Diabetes. Ann. Intern. Med. 2007, 147, 357–369. [Google Scholar] [CrossRef] [PubMed]
- Cortez, M.Y.; Torgan, C.E.; Brozinick, J.T.; Ivy, J.L. Insulin resistance of obese Zucker rats exercise trained at two different intensities. Am. J. Physiol. -Endocrinol. Metab. 1991, 261, E613–E619. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Khazaei, M.; Moien-Afshari, F.; Ang, L.S.; Granville, D.J.; Verchere, C.B.; Dunn, S.R.; McCue, P.; Mizisin, A.; Sharma, K.; et al. Moderate exercise attenuates caspase-3 activity, oxidative stress, and inhibits progression of diabetic renal disease in db/db mice. Am. Physiol. -Ren. Physiol. 2009, 296, F700–F708. [Google Scholar] [CrossRef]
- Ward, K.M.; Mahan, J.D.; Sherman, W.M. Aerobic training and diabetic nephropathy in the obese Zucker rat. Ann. Clin. Lab. Sci. 1994, 24, 266–277. [Google Scholar]
- Arora, E.; Shenoy, S.; Sandhu, J.S. Effects of resistance training on metabolic profile of adults with type 2 diabetes. Indian. J. Med. Res. 2009, 5, 515–519. [Google Scholar]
- Castaneda, C.; Layne, J.E.; Munoz-Orians, L.; Gordon, P.L.; Walsmith, J.; Foldvari, M.; Roubenoff, R.; Tucker, K.L.; Nelson, M.E. A Randomized Controlled Trial of Resistance Exercise Training to Improve Glycemic Control in Older Adults With Type 2 Diabetes. Diabetes Care 2002, 25, 2335–2341. [Google Scholar] [CrossRef]
- Dunstan, D.W.; Daly, R.M.; Owen, N.; Jolley, D.; de Courten, M.; Shaw, J.; Zimmet, P. High-Intensity Resistance Training Improves Glycemic Control in Older Patients With Type 2 Diabetes. Diabetes Care 2002, 25, 1729–1736. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Choi, M.J.; So, B.; Kim, H.; Seong, J.K.; Song, W. The Preventive Effects of 8 Weeks of Resistance Training on Glucose Tolerance and Muscle Fiber Type Composition in Zucker Rats. Diabetes Metab. J. 2015, 39, 424–433. [Google Scholar] [CrossRef] [PubMed]
- Prestes, J.; Leite, R.D.; Pereira, G.B.; Shiguemoto, G.E.; Bernardes, C.F.; Asano, R.Y.; Sales, M.M.; Bartholomeu Neto, J.; Perez, S.E.A. Resistance Training and Glycogen Content in Ovariectomized Rats. Int. J. Sports. Med. 2012, 33, 550–554. [Google Scholar] [CrossRef] [PubMed]
- Quines, C.B.; Jardim, N.S.; Araujo, P.C.O.; Cechella, J.L.; Prado, V.C.; Nogueira, C.W. Resistance training restores metabolic alterations induced by monosodium glutamate in a sex-dependent manner in male and female rats. J. Cell. Biochem. 2019, 120, 13426–13440. [Google Scholar] [CrossRef] [PubMed]
- Scheffer, D.L.; Silva, L.A.; Tromm, C.B.; da Rosa, G.L.; Silveira, P.C.L.; de Souza, C.T.; Latini, A.; Pinho, R.A. Impact of different resistance training protocols on muscular oxidative stress parameters. Appl. Physiol. Nutr. Metab. 2012, 37, 1239–1246. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, A. Work-induced growth of skeletal muscle in normal and hypophysectomized rats. Am. J. Physiol. -Leg. Content 1967, 213, 1193–1198. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, R.B.; Marum, P.; Tullson, P.; Saubert, C.W. Acute hypertrophic response of skeletal muscle to removal of synergists. J. Appl. Physiol. 1979, 46, 835–842. [Google Scholar] [CrossRef] [PubMed]
- Augert, G.; Van de Werve, G.; Le Marchand-Brustel, Y. Effect of work-induced hypertrophy on muscle glucose metabolism in lean and obese mice. Diabetologia 1985, 28, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, K.M.; Martinez, O.M.; Cheadle, W.G. Enzymatic changes in hypertrophied fast-twitch skeletal muscle. Pflugers Arch. 1976, 364, 229–234. [Google Scholar] [CrossRef]
- Bodine, S.C.; Baar, K. Analysis of Skeletal Muscle Hypertrophy in Models of Increased Loading. In Myogenesis: Methods and Protocols; DiMario, J.X., Ed.; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2012; pp. 213–229. ISBN 978-1-61779-343-1. [Google Scholar]
- Ferey, J.L.A.; Brault, J.J.; Smith, C.A.S.; Witczak, C.A. Constitutive activation of CaMKKα signaling is sufficient but not necessary for mTORC1 activation and growth in mouse skeletal muscle. Am. J. Physiol. -Endocrinol. Metab. 2014, 307, E686–E694. [Google Scholar] [CrossRef]
- Johnson, T.L.; Klueber, K.M. Skeletal muscle following tonic overload: Functional and structural analysis. Med. Sci. Sports Exerc. 1991, 23, 49. [Google Scholar] [CrossRef]
- Larkin, L.M.; Kuzon, W.M.; Halter, J.B. Synergist muscle ablation and recovery from nerve-repair grafting: Contractile and metabolic function. J. Appl. Physiol. 2000, 89, 1469–1476. [Google Scholar] [CrossRef] [PubMed]
- Sitnick, M.; Bodine, S.C.; Rutledge, J.C. Chronic high fat feeding attenuates load-induced hypertrophy in mice. J. Physiol. 2009, 587, 5753–5765. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, T.; Saotome, K.; Seino, S.; Shimojo, N.; Matsushita, A.; Iemitsu, M.; Ohshima, H.; Tanaka, K.; Mukai, C. Effects of a low-volume aerobic-type interval exercise on VO2max and cardiac mass. Med. Sci. Sports Exerc. 2014, 46, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Levy, W.C.; Cerqueira, M.D.; Harp, G.D.; Johannessen, K.-A.; Abrass, I.B.; Schwartz, R.S.; Stratton, J.R. Effect of endurance exercise training on heart rate variability at rest in healthy young and older men. Am. J. Cardiol. 1998, 82, 1236–1241. [Google Scholar] [CrossRef]
- Cornelissen, V.A.; Verheyden, B.; Aubert, A.E.; Fagard, R.H. Effects of aerobic training intensity on resting, exercise and post-exercise blood pressure, heart rate and heart-rate variability. J. Hum. Hypertens. 2010, 24, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Katch, V.L.; Sady, S.S.; Freedson, P. Biological variability in maximum aerobic power. Med. Sci. Sports Exerc. 1982, 14, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Solomon, T.P.J.; Malin, S.K.; Karstoft, K.; Knudsen, S.H.; Haus, J.M.; Laye, M.J.; Kirwan, J.P. Association Between Cardiorespiratory Fitness and the Determinants of Glycemic Control Across the Entire Glucose Tolerance Continuum. Diabetes Care 2015, 38, 921–929. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S.M. Short-Term Training: When Do Repeated Bouts of Resistance Exercise Become Training? Can. J. Appl. Physiol. 2000, 25, 185–193. [Google Scholar] [CrossRef]
- Toigo, M.; Boutellier, U. New fundamental resistance exercise determinants of molecular and cellular muscle adaptations. Eur. J. Appl. Physiol. 2006, 97, 643–663. [Google Scholar] [CrossRef]
- Schoenfeld, B.J. The Mechanisms of Muscle Hypertrophy and Their Application to Resistance Training. J. Strength Cond. Res. 2010, 24, 2857. [Google Scholar] [CrossRef]
- Treuth, M.S.; Ryan, A.S.; Pratley, R.E.; Rubin, M.A.; Miller, J.P.; Nicklas, B.J.; Sorkin, J.; Harman, S.M.; Goldberg, A.P.; Hurley, B.F. Effects of strength training on total and regional body composition in older men. J. Appl. Physiol. 1994, 77, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Duplanty, A.A.; Levitt, D.E.; Hill, D.W.; McFarlin, B.K.; DiMarco, N.M.; Vingren, J.L. Resistance Training Is Associated With Higher Bone Mineral Density Among Young Adult Male Distance Runners Independent of Physiological Factors. J. Strength Cond. Res. 2018, 32, 1594–1600. [Google Scholar] [CrossRef] [PubMed]
- English, K.L.; Loehr, J.A.; Lee, S.M.; Smith, S.M. Early-phase musculoskeletal adaptations to different levels of eccentric resistance after 8 weeks of lower body training. Eur. J. Appl. Physiol. 2014, 114, 2263–2280. [Google Scholar] [CrossRef] [PubMed]
- Braith, R.W.; Beck, D.T. Resistance exercise: Training adaptations and developing a safe exercise prescription. Heart Fail Rev. 2008, 13, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Association, A.D. 2. Classification and Diagnosis of Diabetes: Standards of Medical Care in Diabetes—2019. Diabetes Care 2019, 42, S13–S28. [Google Scholar] [CrossRef] [PubMed]
- Dimitriadis, G.; Mitrou, P.; Lambadiari, V.; Maratou, E.; Raptis, S.A. Insulin effects in muscle and adipose tissue. Diabetes Res. Clin. Pract. 2011, 93, S52–S59. [Google Scholar] [CrossRef]
- Han, H.-S.; Kang, G.; Kim, J.S.; Choi, B.H.; Koo, S.-H. Regulation of glucose metabolism from a liver-centric perspective. Exp. Mol. Med. 2016, 48, e218. [Google Scholar] [CrossRef] [PubMed]
- MacDonald Patrick E; Joseph Jamie W; Rorsman Patrik Glucose-sensing mechanisms in pancreatic β-cells. Philos. Trans. R. Soc. B: Biol. Sci. 2005, 360, 2211–2225. [CrossRef]
- DeFronzo, R.A.; Ferrannini, E.; Simonson, D.C. Fasting hyperglycemia in non-insulin-dependent diabetes mellitus: Contributions of excessive hepatic glucose production and impaired tissue glucose uptake. Metabolism 1989, 38, 387–395. [Google Scholar] [CrossRef]
- Garvey, W.T.; Maianu, L.; Huecksteadt, T.P.; Birnbaum, M.J.; Molina, J.M.; Ciaraldi, T.P. Pretranslational suppression of a glucose transporter protein causes insulin resistance in adipocytes from patients with non-insulin-dependent diabetes mellitus and obesity. J. Clin. Investig. 1991, 87, 1072–1081. [Google Scholar] [CrossRef]
- Kahn, B.B.; Charron, M.J.; Lodish, H.F.; Cushman, S.W.; Flier, J.S. Differential regulation of two glucose transporters in adipose cells from diabetic and insulin-treated diabetic rats. J. Clin. Investig. 1989, 84, 404–411. [Google Scholar] [CrossRef] [PubMed]
- Wallberg-Henriksson, H. Glucose transport into skeletal muscle. Influence of contractile activity, insulin, catecholamines and diabetes mellitus. Acta Physiol. Scand. Suppl. 1987, 564, 1–80. [Google Scholar] [PubMed]
- Zierath, J. In vitro studies of human skeletal muscle: Hormonal and metabolic regulation of glucose transport. Acta Physiol. Scand. Suppl. 1995, 626, 1–96. [Google Scholar] [PubMed]
- Stumvoll, M.; Goldstein, B.J.; van Haeften, T.W. Type 2 diabetes: Principles of pathogenesis and therapy. Lancet 2005, 365, 1333–1346. [Google Scholar] [CrossRef]
- Vijan, S. Type 2 Diabetes. Ann. Intern. Med. 2010, 152, ITC3-1. [Google Scholar] [CrossRef] [PubMed]
- Zaccardi, F.; Webb, D.R.; Yates, T.; Davies, M.J. Pathophysiology of type 1 and type 2 diabetes mellitus: A 90-year perspective. Postgrad. Med J. 2016, 92, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Bacchi, E.; Negri, C.; Zanolin, M.E.; Milanese, C.; Faccioli, N.; Trombetta, M.; Zoppini, G.; Cevese, A.; Bonadonna, R.C.; Schena, F.; et al. Metabolic Effects of Aerobic Training and Resistance Training in Type 2 Diabetic Subjects. Diabetes Care 2012, 35, 676. [Google Scholar] [CrossRef] [PubMed]
- Kadoglou, N.P.E.; Fotiadis, G.; Kapelouzou, A.; Kostakis, A.; Liapis, C.D.; Vrabas, I.S. The differential anti-inflammatory effects of exercise modalities and their association with early carotid atherosclerosis progression in patients with Type 2 diabetes. Diabetic Med. 2013, 30, e41–e50. [Google Scholar] [CrossRef] [PubMed]
- Yavari, A.; Najafipoor, F.; Aliasgharzadeh, A.; Niafar, M.; Mobasseri, M. Effect of aerobic exercise, resistance training or combined training on glycaemic control and cardiovascular risk factors in patients with type 2 diabetes. Biol. Sport 2012, 29, 135–143. [Google Scholar] [CrossRef]
- Najafipour, F.; Mobasseri, M.; Yavari, A.; Nadrian, H.; Aliasgarzadeh, A.; Abbasi, N.M.; Niafar, M.; Gharamaleki, J.H.; Sadra, V. Effect of regular exercise training on changes in HbA1c, BMI and VO2max among patients with type 2 diabetes mellitus: An 8-year trial. BMJ Open Diabetes Res. Care 2017, 5, e000414. [Google Scholar] [CrossRef]
- Sennott, J.; Morrissey, J.; Standley, P.R.; Broderick, T.L. Treadmill exercise training fails to reverse defects in glucose, insulin and muscle GLUT4 content in the db/db mouse model of diabetes. Pathophysiology 2008, 15, 173–179. [Google Scholar] [CrossRef] [PubMed]
- De Lemos, E.T.; Reis, F.; Baptista, S.; Pinto, R.; Sepodes, B.; Vala, H.; Rocha-Pereira, P.; Silva, A.S.; Teixeira, F. Exercise training is associated with improved levels of C-reactive protein and adiponectin in ZDF (type 2) diabetic rats. Med. Sci. Monit. 2007, 13, BR168–BR174. [Google Scholar] [PubMed]
- Teixeira de Lemos, E.; Pinto, R.; Oliveira, J.; Garrido, P.; Sereno, J.; Mascarenhas-Melo, F.; Páscoa-Pinheiro, J.; Teixeira, F.; Reis, F. Differential Effects of Acute (Extenuating) and Chronic (Training) Exercise on Inflammation and Oxidative Stress Status in an Animal Model of Type 2 Diabetes Mellitus. Available online: https://www.hindawi.com/journals/mi/2011/253061/abs/ (accessed on 5 September 2019).
- Király, M.A.; Bates, H.E.; Kaniuk, N.A.; Yue, J.T.Y.; Brumell, J.H.; Matthews, S.G.; Riddell, M.C.; Vranic, M. Swim training prevents hyperglycemia in ZDF rats: Mechanisms involved in the partial maintenance of β-cell function. Am. J. Physiol. -Endocrinol. Metab. 2008, 294, E271–E283. [Google Scholar] [CrossRef] [PubMed]
- Cauza, E.; Hanusch-Enserer, U.; Strasser, B.; Ludvik, B.; Metz-Schimmerl, S.; Pacini, G.; Wagner, O.; Georg, P.; Prager, R.; Kostner, K.; et al. The Relative Benefits of Endurance and Strength Training on the Metabolic Factors and Muscle Function of People With Type 2 Diabetes Mellitus. Arch. of Phys. Med. Rehabil. 2005, 86, 1527–1533. [Google Scholar] [CrossRef] [PubMed]
- Hangping, Z.; Xiaona, Q.; Qi, Z.; Qingchun, L.; Na, Y.; Lijin, J.; Siying, L.; Shuo, Z.; Xiaoming, Z.; Xiaoxia, L.; et al. The impact on glycemic control through progressive resistance training with bioDensityTM in Chinese elderly patients with type 2 diabetes: The PReTTy2 (Progressive Resistance Training in Type 2 Diabetes) Trial. Diabetes Res. Clin. Pract. 2019, 150, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Krüger, K.; Gessner, D.K.; Seimetz, M.; Banisch, J.; Ringseis, R.; Eder, K.; Weissmann, N.; Mooren, F.C. Functional and Muscular Adaptations in an Experimental Model for Isometric Strength Training in Mice. PLoS ONE 2013, 8, e79069. [Google Scholar] [CrossRef]
- DeFronzo, R.A.; Ferrannini, E.; Sato, Y.; Felig, P.; Wahren, J. Synergistic interaction between exercise and insulin on peripheral glucose uptake. J. Clin. Investig. 1981, 68, 1468–1474. [Google Scholar] [CrossRef]
- Baron, A.D.; Brechtel, G.; Wallace, P.; Edelman, S.V. Rates and tissue sites of non-insulin- and insulin-mediated glucose uptake in humans. Am. J. Physiol. 1988, 255, E769–E774. [Google Scholar] [CrossRef]
- Mueckler, M. Family of glucose-transporter genes. Implications for glucose homeostasis and diabetes. Diabetes 1990, 39, 6–11. [Google Scholar] [CrossRef]
- Mueckler, M.; Thorens, B. The SLC2 (GLUT) family of membrane transporters. Mol. Aspects Med. 2013, 34, 121–138. [Google Scholar] [CrossRef]
- Cura, A.J.; Carruthers, A. Role of monosaccharide transport proteins in carbohydrate assimilation, distribution, metabolism, and homeostasis. Compr. Physiol. 2012, 2, 863–914. [Google Scholar] [PubMed]
- Zhao, F.-Q.; Keating, A.F. Functional properties and genomics of glucose transporters. Curr. Genomics 2007, 8, 113–128. [Google Scholar] [CrossRef] [PubMed]
- Scheepers, A.; Joost, H.-G.; Schürmann, A. The glucose transporter families SGLT and GLUT: Molecular basis of normal and aberrant function. JPEN J Parenter Enter. Nutr. 2004, 28, 364–371. [Google Scholar] [CrossRef] [PubMed]
- Deng, D.; Yan, N. GLUT, SGLT, and SWEET: Structural and mechanistic investigations of the glucose transporters. Protein Sci. 2016, 25, 546–558. [Google Scholar] [CrossRef] [PubMed]
- Augustin, R. The protein family of glucose transport facilitators: It’s not only about glucose after all. IUBMB Life 2010, 62, 315–333. [Google Scholar] [CrossRef] [PubMed]
- Wright, E.M.; Loo, D.D.F.; Hirayama, B.A. Biology of human sodium glucose transporters. Physiol. Rev. 2011, 91, 733–794. [Google Scholar] [CrossRef] [PubMed]
- Miele, C.; Formisano, P.; Condorelli, G.; Caruso, M.; Oriente, F.; Andreozzi, F.; Tocchetti, C.G.; Riccardi, G.; Beguinot, F. Abnormal glucose transport and GLUT1 cell-surface content in fibroblasts and skeletal muscle from NIDDM and obese subjects. Diabetologia 1997, 40, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Al-Khalili, L.; Chibalin, A.V.; Kannisto, K.; Zhang, B.B.; Permert, J.; Holman, G.D.; Ehrenborg, E.; Ding, V.D.H.; Zierath, J.R.; Krook, A. Insulin action in cultured human skeletal muscle cells during differentiation: Assessment of cell surface GLUT4 and GLUT1 content. Cell. Mol. Life Sci. 2003, 60, 991–998. [Google Scholar] [CrossRef]
- Janot, M.; Audfray, A.; Loriol, C.; Germot, A.; Maftah, A.; Dupuy, F. Glycogenome expression dynamics during mouse C2C12 myoblast differentiation suggests a sequential reorganization of membrane glycoconjugates. BMC Genomics 2009, 10, 483. [Google Scholar] [CrossRef]
- Niu, W.; Bilan, P.J.; Ishikura, S.; Schertzer, J.D.; Contreras-Ferrat, A.; Fu, Z.; Liu, J.; Boguslavsky, S.; Foley, K.P.; Liu, Z.; et al. Contraction-related stimuli regulate GLUT4 traffic in C2C12-GLUT4myc skeletal muscle cells. Am. J. Physiol. Endocrinol. Metab. 2010, 298, E1058–E1071. [Google Scholar] [CrossRef]
- McMillin, S.L.; Schmidt, D.L.; Kahn, B.B.; Witczak, C.A. GLUT4 Is Not Necessary for Overload-Induced Glucose Uptake or Hypertrophic Growth in Mouse Skeletal Muscle. Diabetes 2017, 66, 1491–1500. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Hansen, P.A.; Marshall, B.A.; Holloszy, J.O.; Mueckler, M. Insulin unmasks a COOH-terminal Glut4 epitope and increases glucose transport across T-tubules in skeletal muscle. J. Cell Biol. 1996, 135, 415–430. [Google Scholar] [CrossRef] [PubMed]
- Andrisse, S.; Patel, G.D.; Chen, J.E.; Webber, A.M.; Spears, L.D.; Koehler, R.M.; Robinson-Hill, R.M.; Ching, J.K.; Jeong, I.; Fisher, J.S. ATM and GLUT1-S490 phosphorylation regulate GLUT1 mediated transport in skeletal muscle. PLoS ONE 2013, 8, e66027. [Google Scholar] [CrossRef] [PubMed]
- Marette, A.; Richardson, J.M.; Ramlal, T.; Balon, T.W.; Vranic, M.; Pessin, J.E.; Klip, A. Abundance, localization, and insulin-induced translocation of glucose transporters in red and white muscle. Am. J. Physiol. 1992, 263, C443–C452. [Google Scholar] [CrossRef] [PubMed]
- Handberg, A.; Kayser, L.; Høyer, P.E.; Vinten, J. A substantial part of GLUT-1 in crude membranes from muscle originates from perineurial sheaths. Am. J. Physiol. 1992, 262, E721–E727. [Google Scholar] [CrossRef]
- Yano, H.; Seino, Y.; Inagaki, N.; Hinokio, Y.; Yamamoto, T.; Yasuda, K.; Masuda, K.; Someya, Y.; Imura, H. Tissue distribution and species difference of the brain type glucose transporter (GLUT3). Biochem. Biophys. Res. Commun. 1991, 174, 470–477. [Google Scholar] [CrossRef]
- Stuart, C.A.; Wen, G.; Jiang, J. GLUT3 protein and mRNA in autopsy muscle specimens. Metab. Clin. Exp. 1999, 48, 876–880. [Google Scholar] [CrossRef]
- Haren, M.T.; Siddiqui, A.M.; Armbrecht, H.J.; Kevorkian, R.T.; Kim, M.J.; Haas, M.J.; Mazza, A.; Kumar, V.B.; Green, M.; Banks, W.A.; et al. Testosterone modulates gene expression pathways regulating nutrient accumulation, glucose metabolism and protein turnover in mouse skeletal muscle. Int. J. Androl. 2011, 34, 55–68. [Google Scholar] [CrossRef]
- Bilan, P.J.; Mitsumoto, Y.; Maher, F.; Simpson, I.A.; Klip, A. Detection of the GLUT3 facilitative glucose transporter in rat L6 muscle cells: Regulation by cellular differentiation, insulin and insulin-like growth factor-I. Biochem. Biophys. Res. Commun. 1992, 186, 1129–1137. [Google Scholar] [CrossRef]
- Teixeira, S.S.; Tamrakar, A.K.; Goulart-Silva, F.; Serrano-Nascimento, C.; Klip, A.; Nunes, M.T. Triiodothyronine acutely stimulates glucose transport into L6 muscle cells without increasing surface GLUT4, GLUT1, or GLUT3. Thyroid 2012, 22, 747–754. [Google Scholar] [CrossRef]
- Guillet-Deniau, I.; Leturque, A.; Girard, J. Expression and cellular localization of glucose transporters (GLUT1, GLUT3, GLUT4) during differentiation of myogenic cells isolated from rat foetuses. J. Cell. Sci. 1994, 107 Pt 3, 487–496. [Google Scholar]
- Shepherd, P.R.; Gould, G.W.; Colville, C.A.; McCoid, S.C.; Gibbs, E.M.; Kahn, B.B. Distribution of GLUT3 glucose transporter protein in human tissues. Biochem. Biophys. Res. Commun. 1992, 188, 149–154. [Google Scholar] [CrossRef]
- Gumà, A.; Zierath, J.R.; Wallberg-Henriksson, H.; Klip, A. Insulin induces translocation of GLUT-4 glucose transporters in human skeletal muscle. Am. J. Physiol. 1995, 268, E613–E622. [Google Scholar] [CrossRef] [PubMed]
- Kraegen, E.W.; Sowden, J.A.; Halstead, M.B.; Clark, P.W.; Rodnick, K.J.; Chisholm, D.J.; James, D.E. Glucose transporters and in vivo glucose uptake in skeletal and cardiac muscle: Fasting, insulin stimulation and immunoisolation studies of GLUT1 and GLUT4. Biochem. J. 1993, 295, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Zisman, A.; Peroni, O.D.; Abel, E.D.; Michael, M.D.; Mauvais-Jarvis, F.; Lowell, B.B.; Wojtaszewski, J.F.; Hirshman, M.F.; Virkamaki, A.; Goodyear, L.J.; et al. Targeted disruption of the glucose transporter 4 selectively in muscle causes insulin resistance and glucose intolerance. Nat. Med. 2000, 6, 924–928. [Google Scholar] [CrossRef] [PubMed]
- Brozinick, J.T.; Etgen, G.J.; Yaspelkis, B.B.; Kang, H.Y.; Ivy, J.L. Effects of exercise training on muscle GLUT-4 protein content and translocation in obese Zucker rats. Am. J. Physiol. 1993, 265, E419–E427. [Google Scholar] [CrossRef] [PubMed]
- Ploug, T.; van Deurs, B.; Ai, H.; Cushman, S.W.; Ralston, E. Analysis of GLUT4 distribution in whole skeletal muscle fibers: Identification of distinct storage compartments that are recruited by insulin and muscle contractions. J. Cell Biol. 1998, 142, 1429–1446. [Google Scholar] [CrossRef] [PubMed]
- Hundal, H.S.; Darakhshan, F.; Kristiansen, S.; Blakemore, S.J.; Richter, E.A. GLUT5 expression and fructose transport in human skeletal muscle. Adv. Exp. Med. Biol. 1998, 441, 35–45. [Google Scholar]
- Kristiansen, S.; Darakhshan, F.; Richter, E.A.; Hundal, H.S. Fructose transport and GLUT-5 protein in human sarcolemmal vesicles. Am. J. Physiol. 1997, 273, E543–E548. [Google Scholar] [CrossRef]
- Ryder, J.W.; Kawano, Y.; Chibalin, A.V.; Rincón, J.; Tsao, T.S.; Stenbit, A.E.; Combatsiaris, T.; Yang, J.; Holman, G.D.; Charron, M.J.; et al. In vitro analysis of the glucose-transport system in GLUT4-null skeletal muscle. Biochem. J. 1999, 342 Pt 2, 321–328. [Google Scholar] [CrossRef]
- Hajduch, E.; Litherland, G.J.; Turban, S.; Brot-Laroche, E.; Hundal, H.S. Insulin regulates the expression of the GLUT5 transporter in L6 skeletal muscle cells. FEBS Lett. 2003, 549, 77–82. [Google Scholar] [CrossRef]
- Darakhshan, F.; Hajduch, E.; Kristiansen, S.; Richter, E.A.; Hundal, H.S. Biochemical and functional characterization of the GLUT5 fructose transporter in rat skeletal muscle. Biochem. J. 1998, 336 Pt 2, 361–366. [Google Scholar] [CrossRef]
- Chaillou, T.; Lee, J.D.; England, J.H.; Esser, K.A.; McCarthy, J.J. Time course of gene expression during mouse skeletal muscle hypertrophy. J. Appl. Physiol. 2013, 115, 1065–1074. [Google Scholar] [CrossRef] [PubMed]
- Doege, H.; Schürmann, A.; Bahrenberg, G.; Brauers, A.; Joost, H.G. GLUT8, a novel member of the sugar transport facilitator family with glucose transport activity. J. Biol. Chem. 2000, 275, 16275–16280. [Google Scholar] [CrossRef] [PubMed]
- Dawson, P.A.; Mychaleckyj, J.C.; Fossey, S.C.; Mihic, S.J.; Craddock, A.L.; Bowden, D.W. Sequence and functional analysis of GLUT10: A glucose transporter in the Type 2 diabetes-linked region of chromosome 20q12-13.1. Mol. Genet. Metab. 2001, 74, 186–199. [Google Scholar] [CrossRef] [PubMed]
- Gaster, M.; Handberg, A.; Schürmann, A.; Joost, H.-G.; Beck-Nielsen, H.; Schrøder, H.D. GLUT11, but not GLUT8 or GLUT12, is expressed in human skeletal muscle in a fibre type-specific pattern. Pflugers Arch. 2004, 448, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Rogers, S.; Macheda, M.L.; Docherty, S.E.; Carty, M.D.; Henderson, M.A.; Soeller, W.C.; Gibbs, E.M.; James, D.E.; Best, J.D. Identification of a novel glucose transporter-like protein-GLUT-12. Am. J. Physiol. Endocrinol. Metab. 2002, 282, E733–E738. [Google Scholar] [CrossRef]
- Stuart, C.A.; Yin, D.; Howell, M.E.A.; Dykes, R.J.; Laffan, J.J.; Ferrando, A.A. Hexose transporter mRNAs for GLUT4, GLUT5, and GLUT12 predominate in human muscle. Am. J. Physiol. Endocrinol. Metab. 2006, 291, E1067–E1073. [Google Scholar] [CrossRef]
- Shaheen, F.; Grammatopoulos, D.K.; Müller, J.; Zammit, V.A.; Lehnert, H. Extra-nuclear telomerase reverse transcriptase (TERT) regulates glucose transport in skeletal muscle cells. Biochim. Biophys. Acta 2014, 1842, 1762–1769. [Google Scholar] [CrossRef]
- Purcell, S.H.; Aerni-Flessner, L.B.; Willcockson, A.R.; Diggs-Andrews, K.A.; Fisher, S.J.; Moley, K.H. Improved insulin sensitivity by GLUT12 overexpression in mice. Diabetes 2011, 60, 1478–1482. [Google Scholar] [CrossRef]
- Macheda, M.L.; Kelly, D.J.; Best, J.D.; Rogers, S. Expression during rat fetal development of GLUT12--a member of the class III hexose transporter family. Anat. Embryol. 2002, 205, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Williams, S.; Ho, S.; Loraine, H.; Hagan, D.; Whaley, J.M.; Feder, J.N. Quantitative PCR tissue expression profiling of the human SGLT2 gene and related family members. Diabetes Ther 2010, 1, 57–92. [Google Scholar] [CrossRef] [PubMed]
- Nagai, K.; Yoshida, S.; Konishi, H. Gender differences in the gene expression profiles of glucose transporter GLUT class I and SGLT in mouse tissues. Pharmazie 2014, 69, 856–859. [Google Scholar] [PubMed]
- Castaneda, F.; Layne, J.E.; Castaneda, C. Skeletal muscle sodium glucose co-transporters in older adults with type 2 diabetes undergoing resistance training. Int. J. Med. Sci. 2006, 3, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Díez-Sampedro, A.; Hirayama, B.A.; Osswald, C.; Gorboulev, V.; Baumgarten, K.; Volk, C.; Wright, E.M.; Koepsell, H. A glucose sensor hiding in a family of transporters. Proc. Natl. Acad. Sci. USA 2003, 100, 11753–11758. [Google Scholar] [CrossRef]
- Marshall, B.A.; Ren, J.M.; Johnson, D.W.; Gibbs, E.M.; Lillquist, J.S.; Soeller, W.C.; Holloszy, J.O.; Mueckler, M. Germline manipulation of glucose homeostasis via alteration of glucose transporter levels in skeletal muscle. J. Biol. Chem. 1993, 268, 18442–18445. [Google Scholar]
- Hansen, P.A.; Wang, W.; Marshall, B.A.; Holloszy, J.O.; Mueckler, M. Dissociation of GLUT4 translocation and insulin-stimulated glucose transport in transgenic mice overexpressing GLUT1 in skeletal muscle. J. Biol. Chem. 1998, 273, 18173–18179. [Google Scholar] [CrossRef]
- Deshmukh, A.S.; Murgia, M.; Nagaraj, N.; Treebak, J.T.; Cox, J.; Mann, M. Deep proteomics of mouse skeletal muscle enables quantitation of protein isoforms, metabolic pathways, and transcription factors. Mol. Cell Proteom. 2015, 14, 841–853. [Google Scholar] [CrossRef]
- Jaldin-Fincati, J.R.; Pavarotti, M.; Frendo-Cumbo, S.; Bilan, P.J.; Klip, A. Update on GLUT4 Vesicle Traffic: A Cornerstone of Insulin Action. Trends Endocrinol. Metab. 2017, 28, 597–611. [Google Scholar] [CrossRef]
- Foley, K.; Boguslavsky, S.; Klip, A. Endocytosis, recycling, and regulated exocytosis of glucose transporter 4. Biochemistry 2011, 50, 3048–3061. [Google Scholar] [CrossRef]
- Douen, A.G.; Ramlal, T.; Rastogi, S.; Bilan, P.J.; Cartee, G.D.; Vranic, M.; Holloszy, J.O.; Klip, A. Exercise induces recruitment of the “insulin-responsive glucose transporter”. Evidence for distinct intracellular insulin- and exercise-recruitable transporter pools in skeletal muscle. J. Biol. Chem. 1990, 265, 13427–13430. [Google Scholar] [PubMed]
- Whitfield, J.; Paglialunga, S.; Smith, B.K.; Miotto, P.M.; Simnett, G.; Robson, H.L.; Jain, S.S.; Herbst, E.A.F.; Desjardins, E.M.; Dyck, D.J.; et al. Ablating the protein TBC1D1 impairs contraction-induced sarcolemmal glucose transporter 4 redistribution but not insulin-mediated responses in rats. J. Biol. Chem. 2017, 292, 16653–16664. [Google Scholar] [CrossRef] [PubMed]
- Klip, A.; McGraw, T.E.; James, D.E. 30 sweet years of GLUT4. J. Biol. Chem. 2019, 30, 11369–11381. [Google Scholar] [CrossRef] [PubMed]
- Lauritzen, H.P.M.M. Insulin- and contraction-induced glucose transporter 4 traffic in muscle: Insights from a novel imaging approach. Exerc. Sport Sci. Rev. 2013, 41, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Czech, M.P. The GLUT4 glucose transporter. Cell Metab. 2007, 5, 237–252. [Google Scholar] [CrossRef] [PubMed]
- Bryant, N.J.; Govers, R.; James, D.E. Regulated transport of the glucose transporter GLUT4. Nat. Rev. Mol. Cell Biol. 2002, 3, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Lauritzen, H.P.M.M. In vivo imaging of GLUT4 translocation. Appl. Physiol. Nutr. Metab. 2009, 34, 420–423. [Google Scholar] [CrossRef] [PubMed]
- Ryder, J.W.; Yang, J.; Galuska, D.; Rincón, J.; Björnholm, M.; Krook, A.; Lund, S.; Pedersen, O.; Wallberg-Henriksson, H.; Zierath, J.R.; et al. Use of a novel impermeable biotinylated photolabeling reagent to assess insulin- and hypoxia-stimulated cell surface GLUT4 content in skeletal muscle from type 2 diabetic patients. Diabetes 2000, 49, 647–654. [Google Scholar] [CrossRef]
- Zierath, J.R.; He, L.; Gumà, A.; Odegoard Wahlström, E.; Klip, A.; Wallberg-Henriksson, H. Insulin action on glucose transport and plasma membrane GLUT4 content in skeletal muscle from patients with NIDDM. Diabetologia 1996, 39, 1180–1189. [Google Scholar] [CrossRef]
- Garvey, W.T.; Maianu, L.; Zhu, J.H.; Brechtel-Hook, G.; Wallace, P.; Baron, A.D. Evidence for defects in the trafficking and translocation of GLUT4 glucose transporters in skeletal muscle as a cause of human insulin resistance. J. Clin. Investig. 1998, 101, 2377–2386. [Google Scholar] [CrossRef]
- Shepherd, P.R.; Kahn, B.B. Glucose transporters and insulin action--implications for insulin resistance and diabetes mellitus. N. Engl. J. Med. 1999, 341, 248–257. [Google Scholar] [CrossRef] [PubMed]
- Jessen, N.; Goodyear, L.J. Contraction signaling to glucose transport in skeletal muscle. J. Appl. Physiol. 2005, 99, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, J.W.; Hirshman, M.F.; Gervino, E.V.; Ocel, J.V.; Forse, R.A.; Hoenig, S.J.; Aronson, D.; Goodyear, L.J.; Horton, E.S. Acute exercise induces GLUT4 translocation in skeletal muscle of normal human subjects and subjects with type 2 diabetes. Diabetes 1999, 48, 1192–1197. [Google Scholar] [CrossRef] [PubMed]
- Yeh, J.I.; Gulve, E.A.; Rameh, L.; Birnbaum, M.J. The effects of wortmannin on rat skeletal muscle. Dissociation of signaling pathways for insulin- and contraction-activated hexose transport. J. Biol. Chem. 1995, 270, 2107–2111. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.D.; Hansen, P.A.; Holloszy, J.O. Wortmannin inhibits insulin-stimulated but not contraction-stimulated glucose transport activity in skeletal muscle. FEBS Lett. 1995, 361, 51–54. [Google Scholar] [CrossRef]
- Richter, E.A.; Kristiansen, S.; Wojtaszewski, J.; Daugaard, J.R.; Asp, S.; Hespel, P.; Kiens, B. Training effects on muscle glucose transport during exercise. Adv. Exp. Med. Biol. 1998, 441, 107–116. [Google Scholar] [PubMed]
- Stuart, C.A.; Howell, M.E.A.; Baker, J.D.; Dykes, R.J.; Duffourc, M.M.; Ramsey, M.W.; Stone, M.H. Cycle training increased GLUT4 and activation of mammalian target of rapamycin in fast twitch muscle fibers. Med. Sci. Sports Exerc. 2010, 42, 96–106. [Google Scholar] [CrossRef]
- Houmard, J.A.; Shinebarger, M.H.; Dolan, P.L.; Leggett-Frazier, N.; Bruner, R.K.; McCammon, M.R.; Israel, R.G.; Dohm, G.L. Exercise training increases GLUT-4 protein concentration in previously sedentary middle-aged men. Am. J. Physiol. 1993, 264, E896–E901. [Google Scholar] [CrossRef]
- Dela, F.; Mikines, K.J.; von Linstow, M.; Secher, N.H.; Galbo, H. Effect of training on insulin-mediated glucose uptake in human muscle. Am. J. Physiol. 1992, 263, E1134–E1143. [Google Scholar] [CrossRef]
- Greiwe, J.S.; Hickner, R.C.; Hansen, P.A.; Racette, S.B.; Chen, M.M.; Holloszy, J.O. Effects of endurance exercise training on muscle glycogen accumulation in humans. J. Appl. Physiol. 1999, 87, 222–226. [Google Scholar] [CrossRef]
- Little, J.P.; Safdar, A.; Wilkin, G.P.; Tarnopolsky, M.A.; Gibala, M.J. A practical model of low-volume high-intensity interval training induces mitochondrial biogenesis in human skeletal muscle: Potential mechanisms. J. Physiol. (Lond.) 2010, 588, 1011–1022. [Google Scholar] [CrossRef] [PubMed]
- Short, K.R.; Vittone, J.L.; Bigelow, M.L.; Proctor, D.N.; Rizza, R.A.; Coenen-Schimke, J.M.; Nair, K.S. Impact of aerobic exercise training on age-related changes in insulin sensitivity and muscle oxidative capacity. Diabetes 2003, 52, 1888–1896. [Google Scholar] [CrossRef] [PubMed]
- Burgomaster, K.A.; Cermak, N.M.; Phillips, S.M.; Benton, C.R.; Bonen, A.; Gibala, M.J. Divergent response of metabolite transport proteins in human skeletal muscle after sprint interval training and detraining. Am. J. Physiol. -Regul., Integr. Comp. Physiol. 2007, 292, R1970–R1976. [Google Scholar] [CrossRef] [PubMed]
- Perry, C.G.R.; Heigenhauser, G.J.F.; Bonen, A.; Spriet, L.L. High-intensity aerobic interval training increases fat and carbohydrate metabolic capacities in human skeletal muscle. Appl. Physiol. Nutr. Metab. 2008, 33, 1112–1123. [Google Scholar] [CrossRef] [PubMed]
- Banks, E.A.; Brozinick, J.T.; Yaspelkis, B.B.; Kang, H.Y.; Ivy, J.L. Muscle glucose transport, GLUT-4 content, and degree of exercise training in obese Zucker rats. Am. J. Physiol. 1992, 263, E1010–E1015. [Google Scholar] [CrossRef] [PubMed]
- Neufer, P.D.; Shinebarger, M.H.; Dohm, G.L. Effect of training and detraining on skeletal muscle glucose transporter (GLUT4) content in rats. Can. J. Physiol. Pharm. 1992, 70, 1286–1290. [Google Scholar] [CrossRef] [PubMed]
- Goodyear, L.J.; Hirshman, M.F.; Valyou, P.M.; Horton, E.S. Glucose Transporter Number, Function, and Subcellular Distribution in Rat Skeletal Muscle After Exercise Training. Diabetes 1992, 41, 1091–1099. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, S.B.; Treebak, J.T.; Viollet, B.; Schjerling, P.; Vaulont, S.; Wojtaszewski, J.F.P.; Richter, E.A. Role of AMPKα2 in basal, training-, and AICAR-induced GLUT4, hexokinase II, and mitochondrial protein expression in mouse muscle. Am. J. Physiol. -Endocrinol. Metab. 2007, 292, E331–E339. [Google Scholar] [CrossRef]
- Rodnick, K.J.; Henriksen, E.J.; James, D.E.; Holloszy, J.O. Exercise training, glucose transporters, and glucose transport in rat skeletal muscles. Am. J. Physiol. 1992, 262, C9–C14. [Google Scholar] [CrossRef]
- Hansen, P.A.; Nolte, L.A.; Chen, M.M.; Holloszy, J.O. Increased GLUT-4 translocation mediates enhanced insulin sensitivity of muscle glucose transport after exercise. J. Appl. Physiol. 1998, 85, 1218–1222. [Google Scholar] [CrossRef]
- Frøsig, C.; Rose, A.J.; Treebak, J.T.; Kiens, B.; Richter, E.A.; Wojtaszewski, J.F.P. Effects of endurance exercise training on insulin signaling in human skeletal muscle: Interactions at the level of phosphatidylinositol 3-kinase, Akt, and AS160. Diabetes 2007, 56, 2093–2102. [Google Scholar] [CrossRef] [PubMed]
- Richter, E.A.; Garetto, L.P.; Goodman, M.N.; Ruderman, N.B. Muscle Glucose Metabolism following Exercise in the Rat: INCREASED SENSITIVITY TO INSULIN. J. Clin. Investig. 1982, 69, 785–793. [Google Scholar] [CrossRef] [PubMed]
- Iwabe, M.; Kawamoto, E.; Koshinaka, K.; Kawanaka, K. Increased postexercise insulin sensitivity is accompanied by increased AS160 phosphorylation in slow-twitch soleus muscle. Physiol. Rep. 2014, 2, e12162. [Google Scholar] [CrossRef] [PubMed]
- Cartee, G.D. Mechanisms for Greater Insulin-stimulated Glucose Uptake in Normal and Insulin Resistant Skeletal Muscle after Acute Exercise. Am. J. Physiol. Endocrinol. Metab. 2015, 309, E949–E959. [Google Scholar] [CrossRef] [PubMed]
- Seki, Y.; Berggren, J.R.; Houmard, J.A.; Charron, M.J. Glucose transporter expression in skeletal muscle of endurance-trained individuals. Med. Sci. Sports Exerc. 2006, 38, 1088–1092. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.E.; Ma, J.; Sacharidou, A.; Mi, W.; Salato, V.K.; Nguyen, N.; Jiang, Y.; Pascual, J.M.; North, P.E.; Shaul, P.W.; et al. A Protein Kinase C Phosphorylation Motif in GLUT1 Affects Glucose Transport and is Mutated in GLUT1 Deficiency Syndrome. Mol. Cell 2015, 58, 845–853. [Google Scholar] [CrossRef] [PubMed]
- Lisinski, I.; Schürmann, A.; Joost, H.G.; Cushman, S.W.; Al-Hasani, H. Targeting of GLUT6 (formerly GLUT9) and GLUT8 in rat adipose cells. Biochem. J. 2001, 358, 517–522. [Google Scholar] [CrossRef]
- Flessner, L.B.; Moley, K.H. Similar [DE]XXXL[LI] motifs differentially target GLUT8 and GLUT12 in Chinese hamster ovary cells. Traffic 2009, 10, 324–333. [Google Scholar] [CrossRef]
- Holten, M.K.; Zacho, M.; Gaster, M.; Juel, C.; Wojtaszewski, J.F.P.; Dela, F. Strength training increases insulin-mediated glucose uptake, GLUT4 content, and insulin signaling in skeletal muscle in patients with type 2 diabetes. Diabetes 2004, 53, 294–305. [Google Scholar] [CrossRef]
- Yaspelkis, B.B.; Singh, M.K.; Trevino, B.; Krisan, A.D.; Collins, D.E. Resistance training increases glucose uptake and transport in rat skeletal muscle. Acta Physiol. Scand. 2002, 175, 315–323. [Google Scholar] [CrossRef]
- Young, J.C.; Kandarian, S.C.; Kurowski, T.G. Skeletal muscle glucose uptake following overload-induced hypertrophy. Life Sci. 1992, 50, 1319–1325. [Google Scholar] [CrossRef]
- Pereira, R.O.; Wende, A.R.; Olsen, C.; Soto, J.; Rawlings, T.; Zhu, Y.; Riehle, C.; Abel, E.D. GLUT1 deficiency in cardiomyocytes does not accelerate the transition from compensated hypertrophy to heart failure. J. Mol. Cell. Cardiol. 2014, 72, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-C.; Huang, H.-Y.; Chang, C.-J.; Cheng, C.-H.; Chen, Y.-T. Mitochondrial GLUT10 facilitates dehydroascorbic acid import and protects cells against oxidative stress: Mechanistic insight into arterial tortuosity syndrome. Hum. Mol. Genet. 2010, 19, 3721–3733. [Google Scholar] [CrossRef] [PubMed]
- Syu, Y.-W.; Lai, H.-W.; Jiang, C.-L.; Tsai, H.-Y.; Lin, C.-C.; Lee, Y.-C. GLUT10 maintains the integrity of major arteries through regulation of redox homeostasis and mitochondrial function. Hum. Mol. Genet. 2018, 27, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Halseth, A.E.; Bracy, D.P.; Wasserman, D.H. Overexpression of hexokinase II increases insulinand exercise-stimulated muscle glucose uptake in vivo. Am. J. Physiol. -Endocrinol. Metab. 1999, 276, E70–E77. [Google Scholar] [CrossRef] [PubMed]
- O’Doherty, R.M.; Bracy, D.P.; Osawa, H.; Wasserman, D.H.; Granner, D.K. Rat skeletal muscle hexokinase II mRNA and activity are increased by a single bout of acute exercise. Am. J. Physiol. 1994, 266, E171–E178. [Google Scholar] [CrossRef]
- Koval, J.A.; DeFronzo, R.A.; O’Doherty, R.M.; Printz, R.; Ardehali, H.; Granner, D.K.; Mandarino, L.J. Regulation of hexokinase II activity and expression in human muscle by moderate exercise. Am. J. Physiol. -Endocrinol. Metab. 1998, 274, E304–E308. [Google Scholar] [CrossRef]
- Bonadonna, R.C.; Del Prato, S.; Bonora, E.; Saccomani, M.P.; Gulli, G.; Natali, A.; Frascerra, S.; Pecori, N.; Ferrannini, E.; Bier, D.; et al. Roles of glucose transport and glucose phosphorylation in muscle insulin resistance of NIDDM. Diabetes 1996, 45, 915–925. [Google Scholar] [CrossRef]
- Fueger, P.T.; Heikkinen, S.; Bracy, D.P.; Malabanan, C.M.; Pencek, R.R.; Laakso, M.; Wasserman, D.H. Hexokinase II partial knockout impairs exercise-stimulated glucose uptake in oxidative muscles of mice. Am. J. Physiol. -Endocrinol. Metab. 2003, 285, E958–E963. [Google Scholar] [CrossRef][Green Version]
- Fueger, P.T.; Lee-Young, R.S.; Shearer, J.; Bracy, D.P.; Heikkinen, S.; Laakso, M.; Rottman, J.N.; Wasserman, D.H. Phosphorylation Barriers to Skeletal and Cardiac Muscle Glucose Uptakes in High-Fat–Fed Mice. Diabetes 2007, 56, 2476–2484. [Google Scholar] [CrossRef]
- Barnard, R.J.; Peter, J.B. Effect of training and exhaustion on hexokinase activity of skeletal muscle. J. Appl. Physiol. 1969, 27, 691–695. [Google Scholar] [CrossRef] [PubMed]
- Lamb, D.; Peter, J.; Jeffress, R.; Wallace, H. Glycogen, hexokinase, and glycogen synthetase adaptations to exercise. Am. J. Physiol. -Leg. Content 1969, 217, 1628–1632. [Google Scholar] [CrossRef] [PubMed]
- Bylund, A.C.; Bjurö, T.; Cederblad, G.; Holm, J.; Lundholm, K.; Sjöström, M.; Ängquist, K.A.; Scherstén, T. Physical training in man—Keletal muscle metabolism in relation to muscle morphology and running ability. Eur. J. Appl. Physiol. Occup. Physiol. 1977, 36, 151–169. [Google Scholar] [CrossRef] [PubMed]
- Mandroukas, K.; Krotkiewski, M.; Hedberg, M.; Wroblewski, Z.; Björntorp, P.; Grimby, G. Physical training in obese women—Effects of muscle morphology, biochemistry and function. Eur. J. Appl. Physiol. Occup. Physiol. 1984, 52, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Röckl, K.S.C.; Hirshman, M.F.; Brandauer, J.; Fujii, N.; Witters, L.A.; Goodyear, L.J. Skeletal muscle adaptation to exercise training: AMP-activated protein kinase mediates muscle fiber type shift. Diabetes 2007, 56, 2062–2069. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, Y.; Gao, R.; Xing, Y.; Li, X.; Wang, Z. Serum metabolomic response to exercise training in spontaneously hypertensive rats. J. Am. Soc. Hypertens.: JASH 2017, 11, 428–436. [Google Scholar] [CrossRef]
- Real-Hohn, A.; Navegantes, C.; Ramos, K.; Ramos-Filho, D.; Cahuê, F.; Galina, A.; Salerno, V.P. The synergism of high-intensity intermittent exercise and every-other-day intermittent fasting regimen on energy metabolism adaptations includes hexokinase activity and mitochondrial efficiency. PLoS ONE 2018, 13, e0202784. [Google Scholar] [CrossRef]
- Brandt, N.; Dethlefsen, M.M.; Bangsbo, J.; Pilegaard, H. PGC-1α and exercise intensity dependent adaptations in mouse skeletal muscle. PLoS ONE 2017, 12, e0185993. [Google Scholar] [CrossRef]
- Steenberg, D.E.; Jørgensen, N.B.; Birk, J.B.; Sjøberg, K.A.; Kiens, B.; Richter, E.A.; Wojtaszewski, J.F.P. Exercise training reduces the insulin-sensitizing effect of a single bout of exercise in human skeletal muscle. J. Physiol. 2019, 597, 89–103. [Google Scholar] [CrossRef]
- Kleinert, M.; Parker, B.L.; Jensen, T.E.; Raun, S.H.; Pham, P.; Han, X.; James, D.E.; Richter, E.A.; Sylow, L. Quantitative proteomic characterization of cellular pathways associated with altered insulin sensitivity in skeletal muscle following high-fat diet feeding and exercise training. Sci. Rep. 2018, 8, 10723. [Google Scholar] [CrossRef]
- Thibault, M.C.; Simoneau, J.A.; Côté, C.; Boulay, M.R.; Lagassé, P.; Marcotte, M.; Bouchard, C. Inheritance of human muscle enzyme adaptation to isokinetic strength training. Hum. Hered. 1986, 36, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Green, H.J.; Dahly, A.; Shoemaker, K.; Goreham, C.; Bombardier, E.; Ball-Burnett, M. Serial effects of high-resistance and prolonged endurance training on Na+-K+ pump concentration and enzymatic activities in human vastus lateralis. Acta Physiol. Scand. 1999, 165, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Tesch, P.A.; Komi, P.V.; Häkkinen, K. Enzymatic adaptations consequent to long-term strength training. Int. J. Sports Med. 1987, 8 Suppl 1, 66–69. [Google Scholar] [CrossRef]
- Manchester, J.; Skurat, A.V.; Roach, P.; Hauschka, S.D.; Lawrence, J.C. Increased glycogen accumulation in transgenic mice over expressing glycogen synthase in skeletal muscle. Proc. Natl. Acad. Sci. USA 1996, 93, 10707–10711. [Google Scholar] [CrossRef] [PubMed]
- Xirouchaki, C.E.; Mangiafico, S.P.; Bate, K.; Ruan, Z.; Huang, A.M.; Tedjosiswoyo, B.W.; Lamont, B.; Pong, W.; Favaloro, J.; Blair, A.R.; et al. Impaired glucose metabolism and exercise capacity with muscle-specific glycogen synthase 1 (gys1) deletion in adult mice. Mol. Metab. 2016, 5, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Perseghin, G.; Price, T.; Petersen, K.; Roden, M.; Cline, G.; Gerow, K.; Rothman, D.; Shulman, G. Increased Glucose Transport—Phosphorylation and Muscle Glycogen Synthesis After Exercise Training in Insulin-Resistant. New Engl. J. Med. 1996, 335, 1357–1362. [Google Scholar] [CrossRef] [PubMed]
- Manabe, Y.; Gollisch, K.S.C.; Holton, L.; Kim, Y.-B.; Brandauer, J.; Fujii, N.L.; Hirshman, M.F.; Goodyear, L.J. Exercise training-induced adaptations associated with increases in skeletal muscle glycogen content. FEBS J. 2013, 280, 916–926. [Google Scholar] [CrossRef]
- Gollnick, P.D.; Armstrong, R.B.; Saltin, B.; Saubert, C.W.; Sembrowich, W.L.; Shepherd, R.E. Effect of training on enzyme activity and fiber composition of human skeletal muscle. J. Appl. Physiol. 1973, 34, 107–111. [Google Scholar] [CrossRef]
- Hickner, R.C.; Fisher, J.S.; Hansen, P.A.; Racette, S.B.; Mier, C.M.; Turner, M.J.; Holloszy, J.O. Muscle glycogen accumulation after endurance exercise in trained and untrained individuals. J. Appl. Physiol. 1997, 83, 897–903. [Google Scholar] [CrossRef]
- Bergström, J.; Hermansen, L.; Hultman, E.; Saltin, B. Diet, Muscle Glycogen and Physical Performance. Acta Physiol. Scand. 1967, 71, 140–150. [Google Scholar] [CrossRef]
- Murray, B.; Rosenbloom, C. Fundamentals of glycogen metabolism for coaches and athletes. Nutr. Rev. 2018, 76, 243–259. [Google Scholar] [CrossRef]
- Haun, C.T.; Vann, C.G.; Mobley, C.B.; Osburn, S.C.; Mumford, P.W.; Roberson, P.A.; Romero, M.A.; Fox, C.D.; Parry, H.A.; Kavazis, A.N.; et al. Pre-training Skeletal Muscle Fiber Size and Predominant Fiber Type Best Predict Hypertrophic Responses to 6 Weeks of Resistance Training in Previously Trained Young Men. Front. in Physiol. 2019, 10, 297. [Google Scholar] [CrossRef]
- Haun, C.T.; Vann, C.G.; Osburn, S.C.; Mumford, P.W.; Roberson, P.A.; Romero, M.A.; Fox, C.D.; Johnson, C.A.; Parry, H.A.; Kavazis, A.N.; et al. Muscle fiber hypertrophy in response to 6 weeks of high-volume resistance training in trained young men is largely attributed to sarcoplasmic hypertrophy. PLoS ONE 2019, 14, e0215267. [Google Scholar] [CrossRef]
- MacDougall, J.D.; Ward, G.R.; Sale, D.G.; Sutton, J.R. Biochemical adaptation of human skeletal muscle to heavy resistance training and immobilization. J. Appl. Physiol. 1977, 43, 700–703. [Google Scholar] [CrossRef]
- Tarui, S.; Giichi, O.; Ikura, Y.; Tanaka, T.; Suda, M.; Nishikawa, M. Phosphofructokinase deficiency in skeletal muscle. A new type of glycogenosis. Biochem. Biophys. Res. Commun. 1965, 19, 517–523. [Google Scholar] [CrossRef]
- García, M.; Pujol, A.; Ruzo, A.; Riu, E.; Ruberte, J.; Arbós, A.; Serafín, A.; Albella, B.; Felíu, J.E.; Bosch, F. Phosphofructo-1-kinase deficiency leads to a severe cardiac and hematological disorder in addition to skeletal muscle glycogenosis. PLoS Genet. 2009, 5, e1000615. [Google Scholar] [CrossRef] [PubMed]
- Svensson, K.; Dent, J.R.; Tahvilian, S.; Martins, V.F.; Sathe, A.; Ochala, J.; Patel, M.S.; Schenk, S. Defining the contribution of skeletal muscle pyruvate dehydrogenase α1 to exercise performance and insulin action. Am. J. Physiol. -Endocrinol. Metab. 2018, 315, E1034–E1045. [Google Scholar] [CrossRef] [PubMed]
- Metzger, J.M.; Fitts, R.H. Contractile and biochemical properties of diaphragm: Effects of exercise training and fatigue. J. Appl. Physiol. 1986, 60, 1752–1758. [Google Scholar] [CrossRef] [PubMed]
- Davidson, S.R.; Burnett, M.; Hoffman-Goetz, L. Training effects in mice after long-term voluntary exercise. Med. Sci. Sports Exerc. 2006, 38, 250–255. [Google Scholar] [CrossRef] [PubMed]
- LeBlanc, P.J.; Peters, S.J.; Tunstall, R.J.; Cameron-Smith, D.; Heigenhauser, G.J.F. Effects of aerobic training on pyruvate dehydrogenase and pyruvate dehydrogenase kinase in human skeletal muscle. J. Physiol. 2004, 557, 559–570. [Google Scholar] [CrossRef] [PubMed]
- Ngo, K.T.A.; Denis, C.; Saafi, M.A.; Feasson, L.; Verney, J. Endurance but not resistance training increases intra-myocellular lipid content and β-hydroxyacyl coenzyme A dehydroge in active elderly men. Acta Physiol 2012, 205, 133–144. [Google Scholar] [CrossRef]
- Love, D.C.; Hanover, J.A. The Hexosamine Signaling Pathway: Deciphering the “O-GlcNAc Code”. Sci. Signal. 2005, 2005, re13. [Google Scholar] [CrossRef]
- Shi, H.; Munk, A.; Nielsen, T.S.; Daughtry, M.R.; Larsson, L.; Li, S.; Høyer, K.F.; Geisler, H.W.; Sulek, K.; Kjøbsted, R.; et al. Skeletal muscle O-GlcNAc transferase is important for muscle energy homeostasis and whole-body insulin sensitivity. Mol. Metab. 2018, 11, 160–177. [Google Scholar] [CrossRef] [PubMed]
- Hebert, L.F.; Daniels, M.C.; Zhou, J.; Crook, E.D.; Turner, R.L.; Simmons, S.T.; Neidigh, J.L.; Zhu, J.S.; Baron, A.D.; McClain, D.A. Overexpression of glutamine:fructose-6-phosphate amidotransferase in transgenic mice leads to insulin resistance. J. Clin. Investig. 1996, 98, 930–936. [Google Scholar] [CrossRef] [PubMed]
- Toivonen, M.H.M.; Pöllänen, E.; Ahtiainen, M.; Suominen, H.; Taaffe, D.R.; Cheng, S.; Takala, T.; Kujala, U.M.; Tammi, M.I.; Sipilä, S.; et al. OGT and OGA expression in postmenopausal skeletal muscle associates with hormone replacement therapy and muscle cross-sectional area. Exp. Gerontol. 2013, 48, 1501–1504. [Google Scholar] [CrossRef] [PubMed]
- Hortemo, K.H.; Lunde, P.K.; Anonsen, J.H.; Kvaloy, H.; Munkvik, M.; Rehn, T.A.; Sjaastad, I.; Lunde, I.G.; Aronsen, J.M.; Sejersted, O.M. Exercise training increases protein O-GlcNAcylation in rat skeletal muscle. Physiol. Rep. 2016, 4, 1–20. [Google Scholar] [CrossRef]
- Meijer, A.E. The Pentose Phosphate Pathway in Skeletal Muscle Under Patho-Physiological Conditions. Prog. Histochem. Cytochem. 1991, 22, 1–118. [Google Scholar]
- Wagner, K.R.; Kauffman, F.C.; Max, S.R. The pentose phosphate pathway in regenerating skeletal muscle. Biochem. J. 1978, 170, 17–22. [Google Scholar] [CrossRef]
- Wu, C.-L.; Satomi, Y.; Walsh, K. RNA-seq and metabolomic analyses of Akt1-mediated muscle growth reveals regulation of regenerative pathways and changes in the muscle secretome. BMC Genomics 2017, 18, 181. [Google Scholar] [CrossRef]
- Battistuzzi, G.; D’Urso, M.; Toniolo, D.; Persico, G.M.; Luzzatto, L. Tissue-specific levels of human glucose-6-phosphate dehydrogenase correlate with methylation of specific sites at the 3′ end of the gene. Proc. Natl. Acad. Sci. USA 1985, 82, 1465–1469. [Google Scholar] [CrossRef]
- Glock, G.E.; Mclean, P. Levels of Enzymes of the Direct Oxidative Pathway of Carbohydrate Metabolism in Mammalian Tissues and Tumours. Biochem. J. 1954, 56, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Schwane, J.A.; Armstrong, R.B. Effect of training on skeletal muscle injury from downhill running in rats. J. Appl. Physiol. 1983, 55, 969–975. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, S.; Hayashi, T.; Egawa, T. The Effects of Caffeine on Metabolomic Responses to Muscle Contraction in Rat Skeletal Muscle. Nutrients 2019, 11, 1819. [Google Scholar] [CrossRef] [PubMed]
- Lira, V.A.; Benton, C.R.; Yan, Z.; Bonen, A. PGC-1α regulation by exercise training and its influences on muscle function and insulin sensitivity. Am. J. Physiol. -Endocrinol. Metab. 2010, 299, E145–E161. [Google Scholar] [CrossRef] [PubMed]
- Hatazawa, Y.; Senoo, N.; Tadaishi, M.; Ogawa, Y.; Ezaki, O.; Kamei, Y.; Miura, S. Metabolomic Analysis of the Skeletal Muscle of Mice Overexpressing PGC-1α. PLoS ONE 2015, 10, e0129084. [Google Scholar] [CrossRef]
- Schiaffino, S.; Mammucari, C. Regulation of skeletal muscle growth by the IGF1-Akt/PKB pathway: Insights from genetic models. Skelet. Muscle 2011, 1, 4. [Google Scholar] [CrossRef] [PubMed]
- Manning, B.D.; Toker, A. AKT/PKB Signaling: Navigating the Network. Cell 2017, 169, 381–405. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Transporter | Gene | Muscle Models | References |
|---|---|---|---|
| GLUT1 | SLC2A1 | Human muscle | [69,70] |
| C2C12 | [71,72] | ||
| Mouse muscle | [73,74] | ||
| L6 myotubes | [72,75] | ||
| Rat muscle | [76,77] | ||
| GLUT3 | SLC2A3 | Human muscle | [78,79] |
| C2C12 | [71] | ||
| Mouse muscle | [73,80] | ||
| L6 myotubes | [81,82] | ||
| Rat muscle | [83,84] | ||
| GLUT4 | SLC2A4 | Human muscle | [85,86] |
| C2C12 | [71,72] | ||
| Mouse muscle | [87,88] | ||
| L6 myotubes | [82] | ||
| Rat muscle | [76,89] | ||
| GLUT5 | SLC2A5 | Human muscle | [90,91] |
| C2C12 | [71] | ||
| Mouse muscle | [92] | ||
| L6 myotubes | [93] | ||
| Rat muscle | [94] | ||
| GLUT6 | SLC2A6 | C2C12 | [71] |
| Mouse muscle | [73,95] | ||
| GLUT8 | SLC2A8 | Human muscle | [96] |
| Mouse muscle | [95] | ||
| GLUT10 | SLC2A10 | Human muscle | [97] |
| Mouse muscle | [73,95] | ||
| GLUT11 | SLC2A11 | Human vastus lateralis | [98] |
| (slow-twitch fibers) | |||
| GLUT12 | SLC2A12 | Human muscle | [99,100] |
| C2C12 | [71,101] | ||
| Mouse muscle | [102] | ||
| Rat muscle | [103] | ||
| SGLT1 | SLC5A1 | Human muscle | [104] |
| Mouse muscle | [105] | ||
| SGLT2 | SLC5A2 | Mouse muscle | [105] |
| SGLT3 | SLC5A4 | Human muscle | [106] |
| Mouse muscle | [107] | ||
| SGLT4 | SLC5A9 | Human muscle | [104] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Evans, P.L.; McMillin, S.L.; Weyrauch, L.A.; Witczak, C.A. Regulation of Skeletal Muscle Glucose Transport and Glucose Metabolism by Exercise Training. Nutrients 2019, 11, 2432. https://doi.org/10.3390/nu11102432
Evans PL, McMillin SL, Weyrauch LA, Witczak CA. Regulation of Skeletal Muscle Glucose Transport and Glucose Metabolism by Exercise Training. Nutrients. 2019; 11(10):2432. https://doi.org/10.3390/nu11102432
Chicago/Turabian StyleEvans, Parker L., Shawna L. McMillin, Luke A. Weyrauch, and Carol A. Witczak. 2019. "Regulation of Skeletal Muscle Glucose Transport and Glucose Metabolism by Exercise Training" Nutrients 11, no. 10: 2432. https://doi.org/10.3390/nu11102432
APA StyleEvans, P. L., McMillin, S. L., Weyrauch, L. A., & Witczak, C. A. (2019). Regulation of Skeletal Muscle Glucose Transport and Glucose Metabolism by Exercise Training. Nutrients, 11(10), 2432. https://doi.org/10.3390/nu11102432