Metagenomic Insights into the Effects of Fructooligosaccharides (FOS) on the Composition of Luminal and Mucosal Microbiota in C57BL/6J Mice, Especially the Bifidobacterium Composition

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Animals and Sample Collection
2.3. Determination of the Weight of Cecum
2.4. Determination of the pH and SCFAs Level of the Intestinal Content
2.5. DNA Extraction and PCR Amplification of the V3–V4 Region and the groEL Gene
2.6. Quantification, Sequencing, and Bioinformatic Analysis
2.7. Data Statistics and Analysis
3. Results
3.1. Cecal Weights in Different Groups
3.2. pH and SCFAs Levels of the Cecal and Colonic Contents
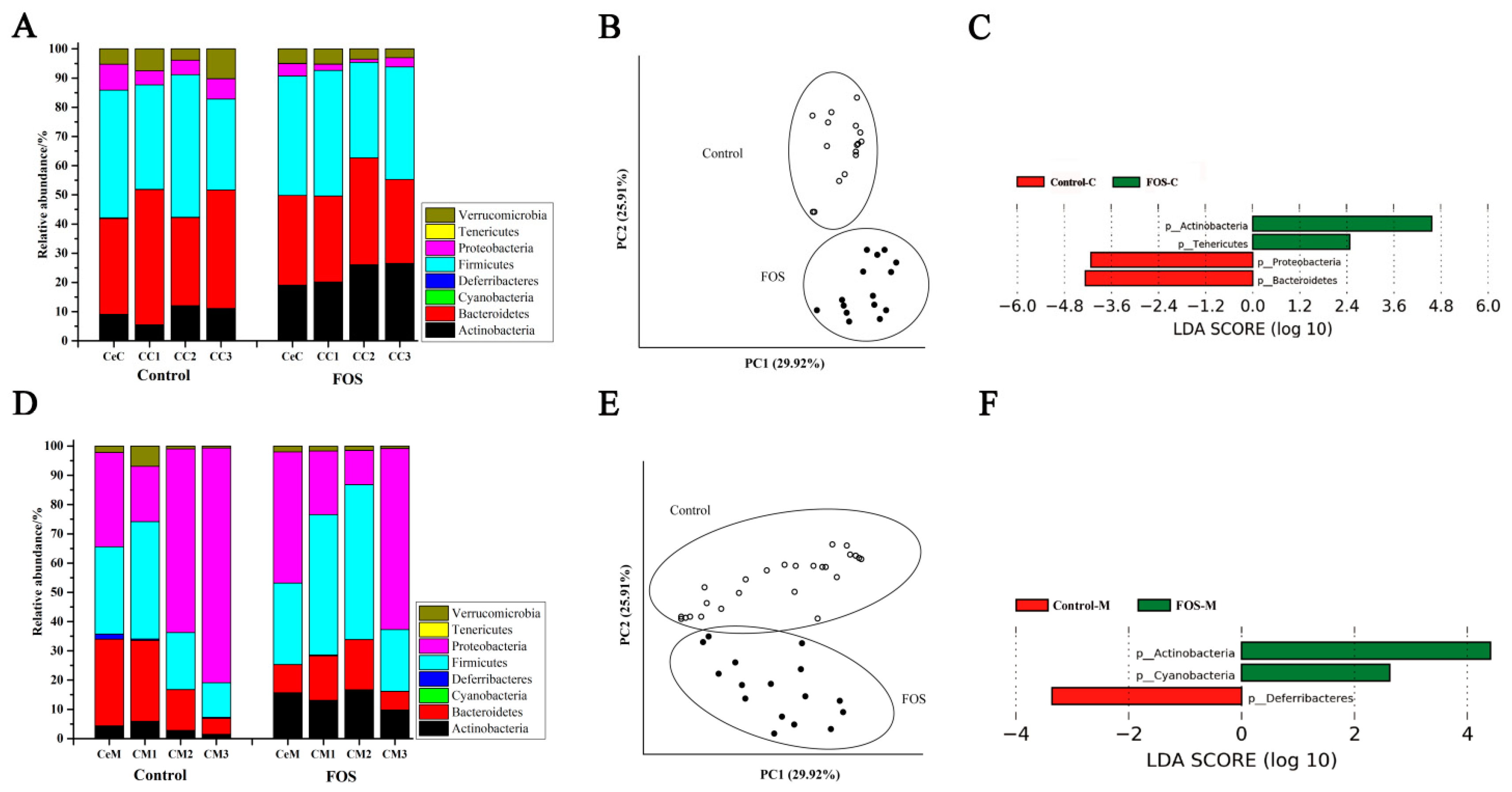
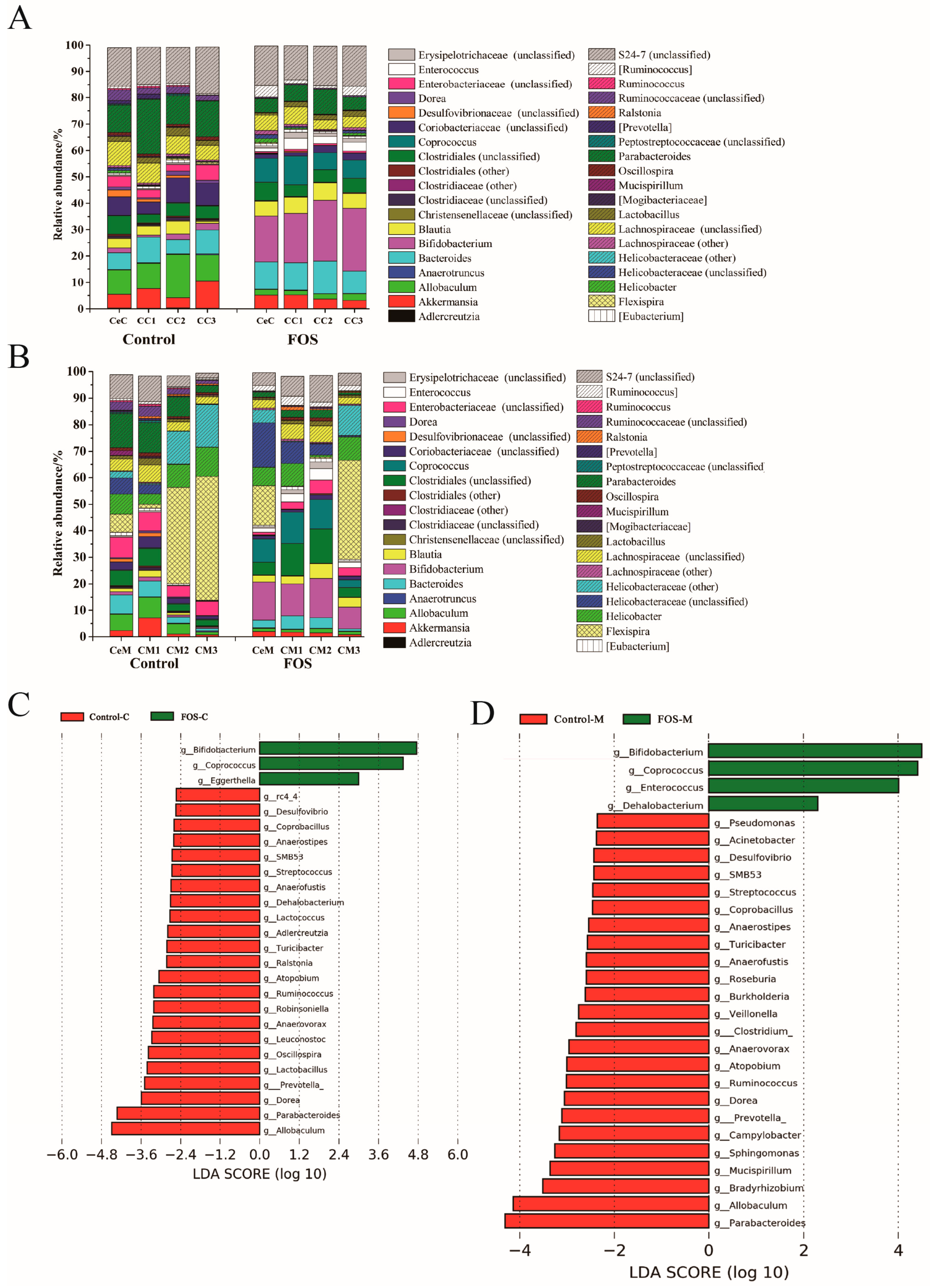
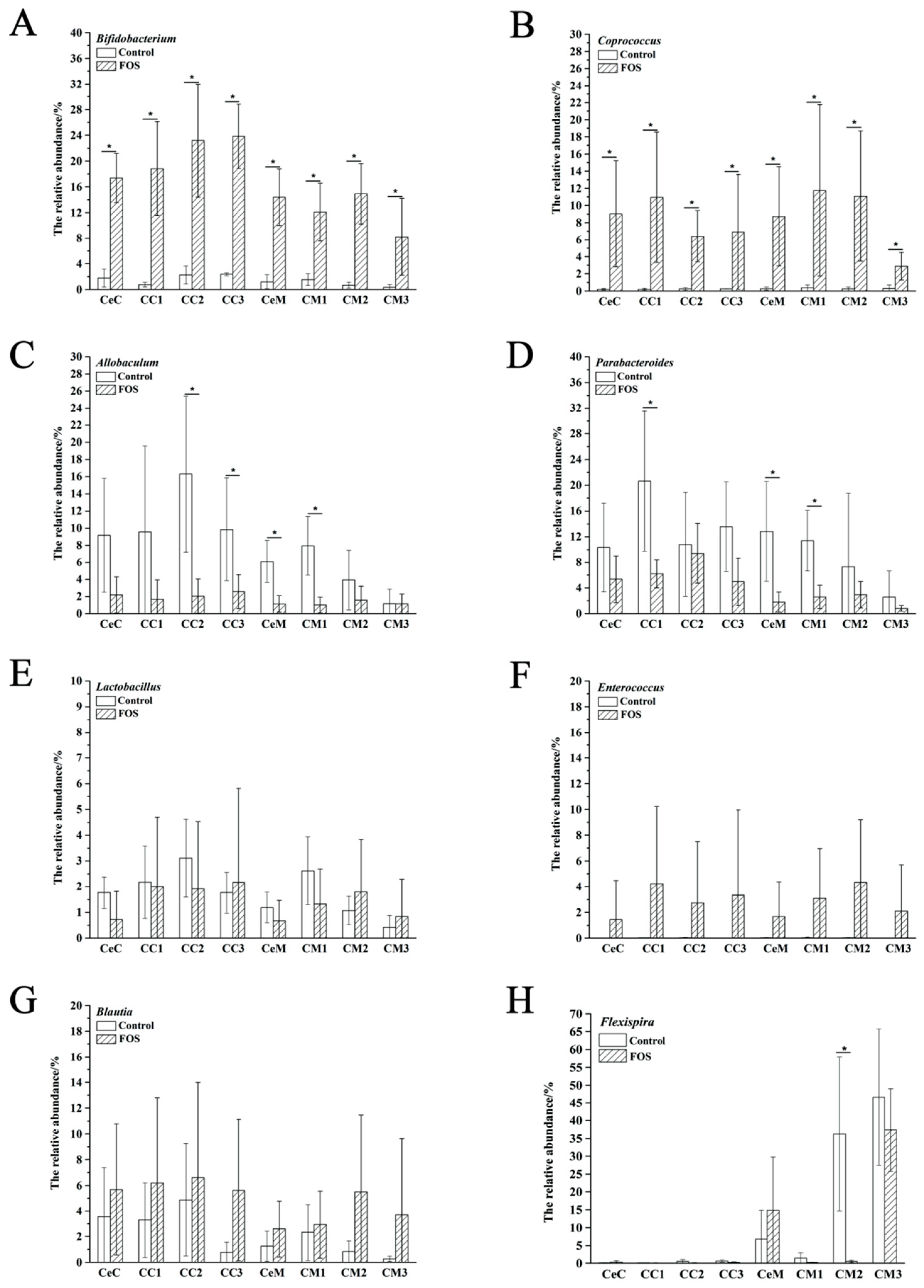
3.3. Effects of FOS on the Composition of Luminal and Mucosal Microbiota in Cecum and Colon
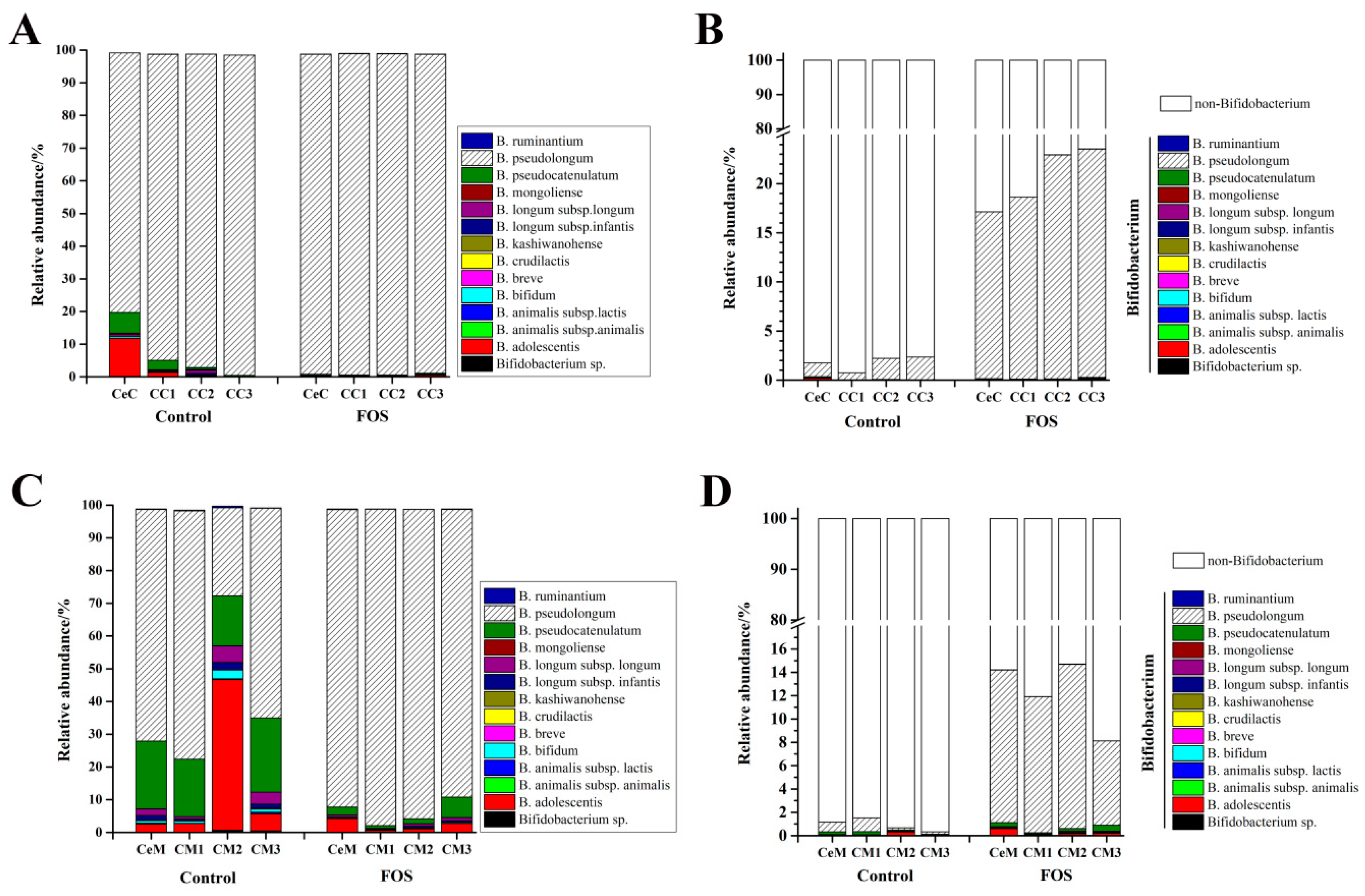
3.4. Effects of FOS on the Composition of Bifidobacterium in the Luminal Contents and Mucosa
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Derrien, M.; Vlieg, V.H.; Johan, E.T. Fate, activity, and impact of ingested bacteria within the human gut microbiota. Trends Microbiol. 2015, 23, 354–366. [Google Scholar] [CrossRef] [PubMed]
- Rakoff-Nahoum, S.; Coyne, M.J.; Comstock, L.E. An ecological network of polysaccharide utilization among human intestinal symbionts. Curr. Biol. 2014, 24, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Faith, J.J.; Guruge, J.L.; Charbonneau, M.; Subramanian, S.; Seedorf, H.; Goodman, A.L.; Clemente, J.C.; Knight, R.; Heath, A.C.; Leibel, R.L. The Long-Term Stability of the Human Gut Microbiota. Science 2013, 341, 1237439. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, G.P.; Lee, S.M.; Mazmanian, S.K. Gut biogeography of the bacterial microbiota. Nat. Rev. Microbiol. 2015, 14, 20–32. [Google Scholar] [CrossRef]
- Dubos, R.; Schaedler, R.W.; Costello, R.; Hoet, P. Indigenous, Indigenous, normal, and autochthonous flora of the gastrointestinal tract. J. Exp. Med. 1965, 122, 67–76. [Google Scholar] [CrossRef]
- Albenberg, L.; Esipova, T.V.; Judge, C.P.; Bittinger, K.; Chen, J.; Laughlin, A.; Grunberg, S.; Baldassano, R.N.; Lewis, J.D.; Li, H. Correlation between intraluminal oxygen gradient and radial partitioning of intestinal microbiota. Gastroenterology 2014, 147, 1055–1063. [Google Scholar] [CrossRef]
- Mao, B.Y.; Li, D.Y.; Zhao, J.X.; Liu, X.M.; Gu, Z.N.; Chen, Y.Q.; Zhang, H.; Chen, W. In vitro fermentation of fructooligosaccharides with human gut bacteria. Food Funct. 2015, 6, 947–954. [Google Scholar] [CrossRef]
- Eckburg, P.B.; Bik, E.M.; Bernstein, C.N.; Purdom, E.; Dethlefsen, L.; Sargent, M.; Gill, S.R.; Nelson, K.E.; Relman, D.A. Diversity of the Human Intestinal Microbial Flora. Science 2005, 308, 1635–1638. [Google Scholar] [CrossRef]
- Mao, B.Y.; Li, D.Y.; Zhao, J.X.; Ai, C.Q.; Zhang, H.; Chen, W. Lactulose differently modulates the composition of luminal and mucosal microbiota in C57BL/6J mice. J. Agric. Food Chem. 2016, 64, 6240–6247. [Google Scholar] [CrossRef]
- Besten, G.D.; Eunen, V.K.; Groen, A.K.; Venema, K.; Reijingoud, D.J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef]
- Giese, E.C.; Hirosi, T.; Silva, R.D.; Aneli, D.M.B. Produção, propriedades e aplicações de oligossacarídeos. Semin. Cienc. Agrar. 2011, 32, 683–700. [Google Scholar] [CrossRef][Green Version]
- Gibson, G.R.; Beatty, E.R.; Wang, X.; Cummings, J.H. Selective stimulation of bifidobacteria in the human colon by oligofructose and inulin. Gastroenterology 1995, 108, 975–982. [Google Scholar] [CrossRef]
- Wang, X.; Gibson, G.R. Effects of the in vitro fermentation of oligofructose and inulin by bacteria growing in the human large intestine. J. Appl. Bacteriol. 1993, 75, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Endo, H.; Tamura, K.; Fukasawa, T.; Kanegae, M.; Koga, J. Comparison of fructooligosaccharide utilization by Lactobacillus and Bacteroides species. Biosci. Biotech. Bioch. 2012, 76, 176–179. [Google Scholar] [CrossRef]
- Mao, B.Y.; Gu, J.Y.; Li, D.Y.; Cui, S.M.; Zhao, J.X.; Zhang, H.; Chen, W. Effects of Different Doses of Fructooligosaccharides (FOS) on the Composition of Mice Fecal Microbiota, Especially the Bifidobacterium Composition. Nutrients 2018, 10, 1105. [Google Scholar] [CrossRef]
- Hidaka, T.; Eida, T.; Takizawa, T.; Tokunaga, T.; Tashiro, Y. Effects of fructooligosaccharides on intestinal flora and human health. Biosci. Microflora 1986, 5, 37–50. [Google Scholar] [CrossRef]
- Pokusaeva, K.; Fitzgerald, G.F.; Sinderen, D.V. Carbohydrate metabolism in Bifidobacteria. Genes Nutri. 2011, 6, 285–306. [Google Scholar] [CrossRef]
- Mao, B.Y.; Li, D.Y.; Zhao, J.X.; Liu, X.M.; Gu, Z.N.; Chen, Y.Q.; Zhang, H.; Chen, W. Metagenomic insights into the effects of fructo-oligosaccharides (FOS) on the composition of fecal microbiota in mice. J. Agric. Food Chem. 2015, 63, 856–863. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Wang, L.; Hu, L.; Yan, S.; Jiang, T.; Fang, S.; Wang, G.; Zhao, J.; Zhang, H.; Chen, W. Effects of different oligosaccharides at various dosages on the composition of gut microbiota and short-chain fatty acids in mice with constipation. Food Funct. 2017, 8, 1966–1978. [Google Scholar] [CrossRef]
- Pan, X.D.; Chen, F.Q.; Tian-Xing, W.U.; Tang, H.G.; Zhao, Z.Y. Prebiotic oligosaccharides change the concentrations of short-chain fatty acids and the microbial population of mouse bowel. J. Zhejiang Univ. Sci. B 2009, 10, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Collares, E.F.; Rossi, M.A.; Macedo, A.S. Experimental dilatation of the cecum and colon in rats. II. Reversion after induction by the continuous administration of lactose. Arq. Gastroenterol. 1985, 22, 192–195. [Google Scholar] [CrossRef] [PubMed]
- Genda, T.; Kondo, T.; Hino, S.; Sugiura, S.; Nishimura, N.; Morita, T. The Impact of Fructo-Oligosaccharides on Gut Permeability and Inflammatory Responses in the Cecal Mucosa Quite Differs between Rats Fed Semi-Purified and Non-Purified Diets. J. Nutr. Sci. Vitaminol. 2018, 64, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Tandon, D.; Haque, M.M.; Gote, M.; Jain, M.; Bhaduri, A.; Dubey, A.K.; Mande, S.S. A prospective randomized, double-blind, placebo-controlled, dose-response relationship study to investigate efficacy of fructo-oligosaccharides (FOS) on human gut microflora. Sci. Rep. 2019, 9, 5473. [Google Scholar] [CrossRef]
- Scott, K.P.; Martin, J.C.; Duncan, S.H.; Flint, H.J. Prebiotic stimulation of human colonic butyrate-producing bacteria and bifidobacteria, in vitro. FEMS Microbiol. Ecol. 2014, 87, 30–40. [Google Scholar] [CrossRef]
- Rivière, A.; Selak, M.; Lantin, D.; Leroy, F.; Vuyst, L.D. Bifidobacteria and Butyrate-Producing Colon Bacteria: Importance and Strategies for Their Stimulation in the Human Gut. Front. Microbiol. 2016, 7, 979. [Google Scholar] [CrossRef]
- Derrien, M.; Passel, M.W.V.; Bovenkamp, J.H.V.D.; Schipper, R.G.; Vos, W.M.D.; Dekker, J. Mucin-bacterial interactions in the human oral cavity and digestive tract. Gut Microbes 2010, 1, 254–268. [Google Scholar] [CrossRef]
- Linden, S.K.; Sutton, P.; Karlsson, N.G.; Korolik, V.; Mcguckin, M.A. Mucins in the mucosal barrier to infection. Mucosal Immunol. 2008, 1, 183–197. [Google Scholar] [CrossRef]
- Schauer, D.B.; Ghori, N.; Falkow, S. Isolation and characterization of “Flexispira rappini” from laboratory mice. J. Clin. Microbiol. 1993, 31, 2709–2714. [Google Scholar] [CrossRef]
- Romero, S.; Archer, J.R.; Hamacher, M.E.; Bologna, S.M.; Schell, R.F. Case report of an unclassified microaerophilic bacterium associated with gastroenteritis. J. Clin. Microbiol. 1988, 26, 142–143. [Google Scholar]
- Yoshioka, H.; Fujita, K.; Sakata, H.; Murono, K.; Iseki, K.I. Development of the Normal Intestinal Flora and Clinical Significance in Infants and Children. Bifidobact. Microflora 1991, 10, 11–17. [Google Scholar] [CrossRef][Green Version]
- Welch, J.L.M.; Hasegawa, Y.; Mcnulty, N.P.; Gordon, J.I.; Borisy, G.G. Spatial organization of a model 15-member human gut microbiota established in gnotobiotic mice. Proc. Natl. Acad. Sci. USA 2017, 114, E9105. [Google Scholar] [CrossRef] [PubMed]
- Marx, S.P.; Winkler, S.; Hartmeier, W. Metabolization of β-(2,6)-linked fructose-oligosaccharides by different bifidobacteria. FEMS Microbiol. Lett. 2000, 182, 163–169. [Google Scholar] [CrossRef]
- Blay, L.; Michel, C.G.; Hervé, M.B.; Cherbut, C. Prolonged Intake of Fructo-Oligosaccharides Induces a Short-Term Elevation of Lactic Acid-Producing Bacteria and a Persistent Increase in Cecal Butyrate in Rats. J. Nutr. 1999, 129, 2231–2235. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Control Group | FOS Group | p1 |
|---|---|---|---|
| Total weight | 0.28 ± 0.04 b | 0.87 ± 0.16 a | <0.01 |
| Wall weight | 0.10 ± 0.03 b | 0.26 ± 0.05 a | <0.05 |
| Content weight | 0.18 ± 0.03 b | 0.61 ± 0.07 a | <0.01 |
| Item | pH | p1 | ||
|---|---|---|---|---|
| Control Group | FOS Group | |||
| Cecum | 8.03 ± 0.12 a | 6.13 ± 0.06 b | <0.001 | |
| Colon | Proximal | 7.46 ± 0.15 a | 5.93 ± 0.66 b | <0.05 |
| Middle | 7.62 ± 0.26 a | 6.12 ± 0.29 b | <0.001 | |
| Distal | 7.11 ± 0.42 a | 6.43 ± 0.26 b | <0.05 | |
| SCFAs | Control Group | FOS Group | p2 |
|---|---|---|---|
| Acetic acid | 7.41 ± 2.60 b | 31.17 ± 11.08 a | <0.001 |
| Propionic acid | 3.23 ± 0.81 b | 10.17 ± 4.71 a | <0.01 |
| Butyric acid | 1.55 ± 0.36 b | 3.58 ± 0.88 a | <0.01 |
| Isobutyric acid | 3.04 ± 0.73 b | 14.60 ± 7.15 a | <0.01 |
| Valeric acid | 0.77 ± 0.23 b | 1.82 ± 0.44 a | <0.01 |
| Isovaleric acid | 1.12 ± 0.28 b | 2.59 ± 0.64 a | <0.01 |
| Total | 17.12 ± 4.10 b | 63.93 ± 15.74 a | <0.01 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gu, J.; Mao, B.; Cui, S.; Liu, X.; Zhang, H.; Zhao, J.; Chen, W. Metagenomic Insights into the Effects of Fructooligosaccharides (FOS) on the Composition of Luminal and Mucosal Microbiota in C57BL/6J Mice, Especially the Bifidobacterium Composition. Nutrients 2019, 11, 2431. https://doi.org/10.3390/nu11102431
Gu J, Mao B, Cui S, Liu X, Zhang H, Zhao J, Chen W. Metagenomic Insights into the Effects of Fructooligosaccharides (FOS) on the Composition of Luminal and Mucosal Microbiota in C57BL/6J Mice, Especially the Bifidobacterium Composition. Nutrients. 2019; 11(10):2431. https://doi.org/10.3390/nu11102431
Chicago/Turabian StyleGu, Jiayu, Bingyong Mao, Shumao Cui, Xuemei Liu, Hao Zhang, Jianxin Zhao, and Wei Chen. 2019. "Metagenomic Insights into the Effects of Fructooligosaccharides (FOS) on the Composition of Luminal and Mucosal Microbiota in C57BL/6J Mice, Especially the Bifidobacterium Composition" Nutrients 11, no. 10: 2431. https://doi.org/10.3390/nu11102431
APA StyleGu, J., Mao, B., Cui, S., Liu, X., Zhang, H., Zhao, J., & Chen, W. (2019). Metagenomic Insights into the Effects of Fructooligosaccharides (FOS) on the Composition of Luminal and Mucosal Microbiota in C57BL/6J Mice, Especially the Bifidobacterium Composition. Nutrients, 11(10), 2431. https://doi.org/10.3390/nu11102431