Metabolic Trajectories Following Contrasting Prudent and Western Diets from Food Provisions: Identifying Robust Biomarkers of Short-Term Changes in Habitual Diet

 , , ,
, , , 
Abstract

1. Introduction
2. Experimental
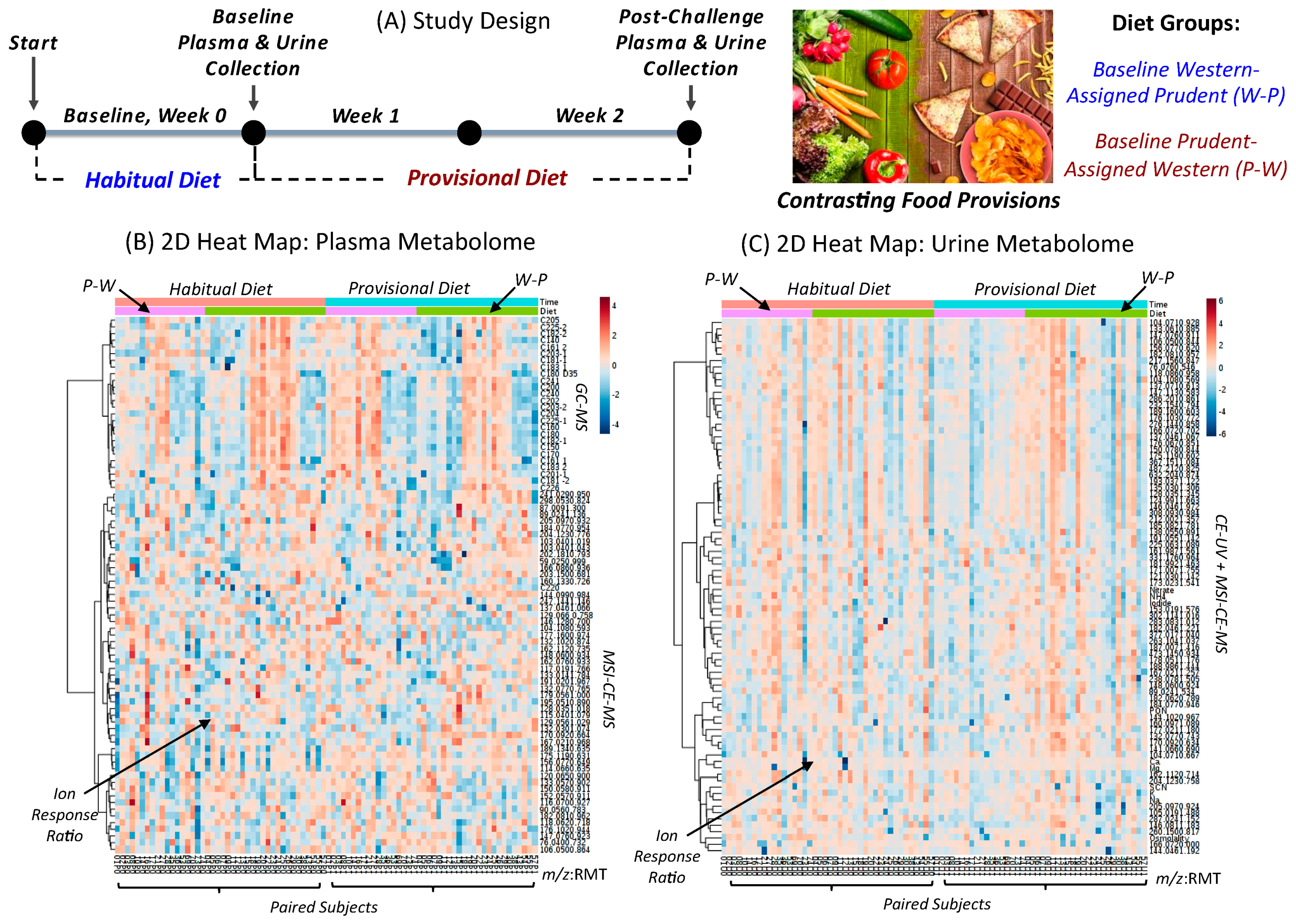
2.1. Study Design, Participant Eligibility and Dietary Self-reporting
2.2. Chemicals and Reagents
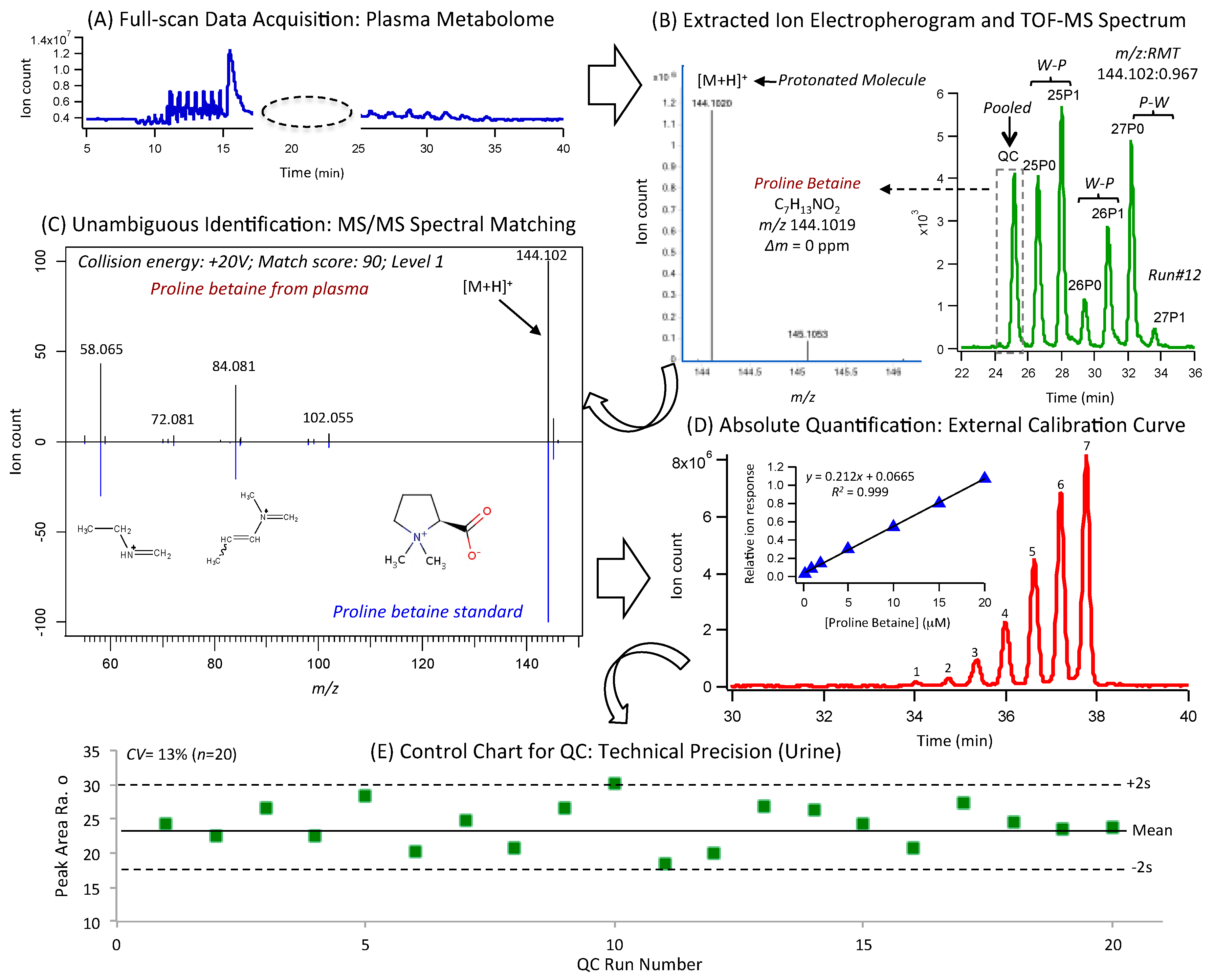
2.3. Nontargeted Metabolite Profiling of Plasma and Urine by MSI-CE-MS
2.4. Unknown Metabolite Identification by MS/MS
2.5. Total Plasma Fatty Acid Determination by GC-MS
2.6. Targeted Urinary Electrolyte Analysis
2.7. Data Preprocessing and Statistical Analysis
3. Results
3.1. Study Design, Baseline Habitual Diet and Metabolomics Workflow
3.2. Changes in Dietary Intake and Biomarker Classification
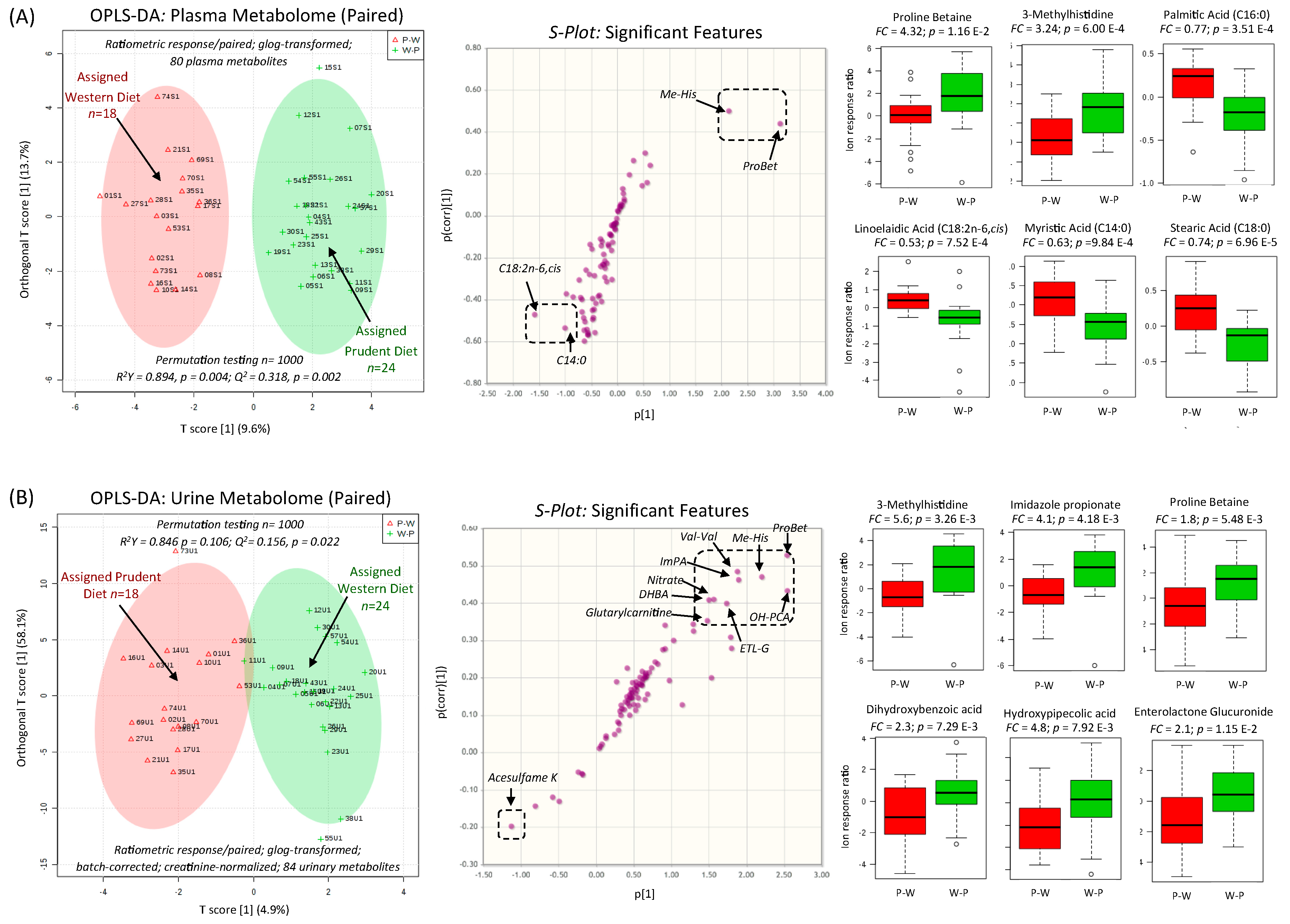
3.3. Biomarkers of Contrasting Diets and Correlation with Diet Records
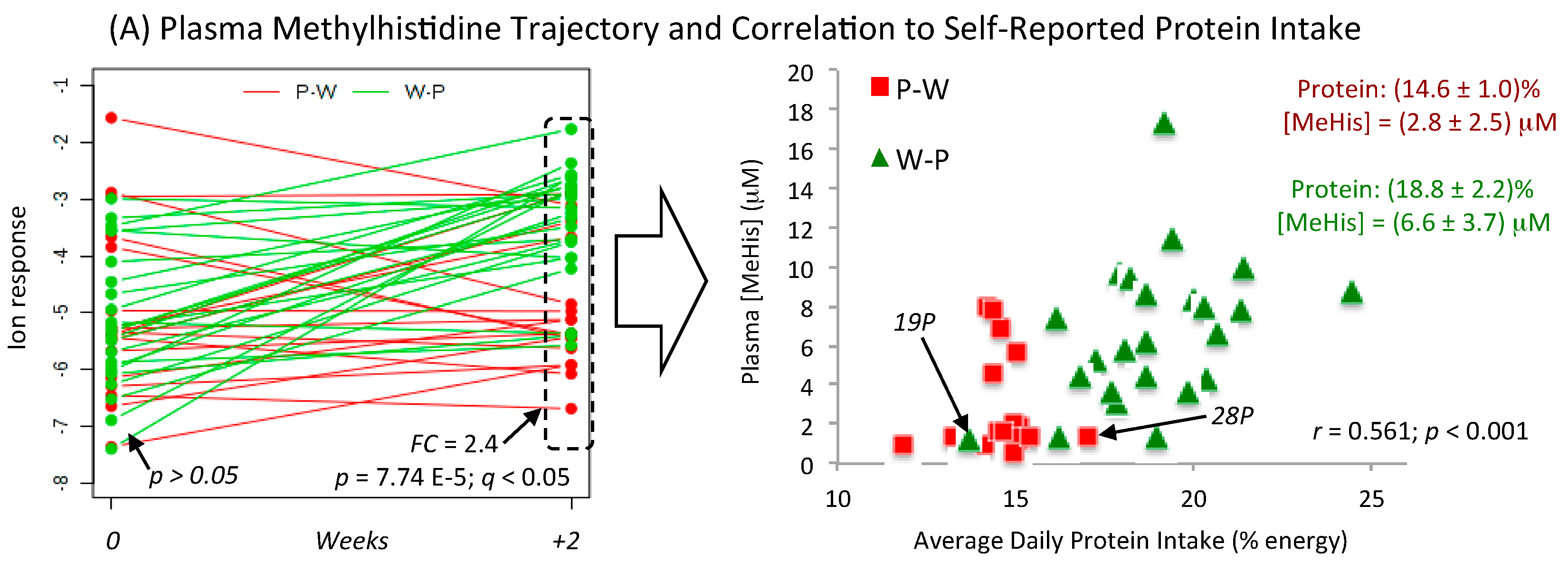
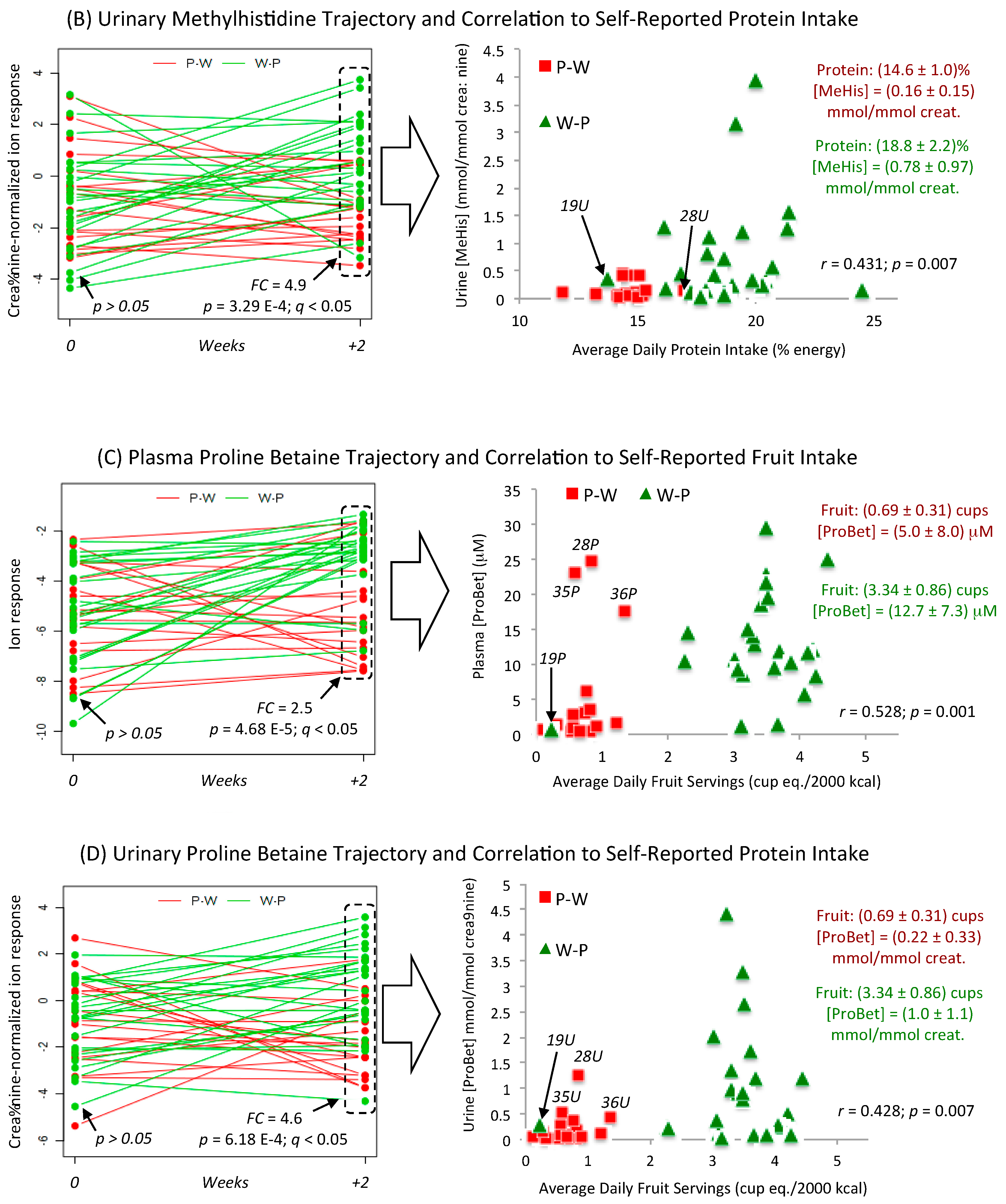
3.4. Metabolic Trajectories and Metabolite Correlation Analysis
4. Discussion
4.1. Contrasting Diets from Food Provisions
4.2. Robust Biomarkers of a Prudent Diet Measured in Both Plasma and Urine
4.3. Novel Biomarkers Identified Following a Prudent Diet
4.4. Novel Biomarkers Identified Following a Western Diet
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Ethics Approval
Acknowledgments
Conflicts of Interest
References
- Hossain, P.; Kawar, B.; El Nahas, P. Obesity and diabetes in the developing world—A growing challenge. N. Engl. J. Med. 2007, 356, 213–215. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Healthy Diet. In Fact Sheet No. 394; World Health Organization: Geneva, Switzerland, 2017; Available online: https://www.who.int/en/news-room/fact-sheets/detail/healthy-diet (accessed on 23 October 2018).
- Mente, A.; Yusuf, S. Evolving evidence about diet and health. Lancet Public Health 2018, 17, e408–e409. [Google Scholar] [CrossRef]
- World Health Organization. Cardiovascular diseases (CVDs). In Fact Sheets; World Health Organization: Geneva, Switzerland, 2017; Available online: http://www.who.int/mediacentre/factsheets/fs317/en/ (accessed on 17 May 2017).
- Cordain, L.; Eaton, S.B.; Sebastian, A.; Mann, N.; Lindeberg, S.; Watkins, B.A.; O’Keefe, J.H.; Brand-Miller, J. Origins and evolution of the Western diet: Health Implications for the 21st century. Am. J. Clin. Nutr. 2005, 81, 341–354. [Google Scholar] [CrossRef] [PubMed]
- Al-Hamad, D.; Raman, D. Metabolic syndrome in children and adolescents. Transl. Pediatr. 2017, 6, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Wood, P.D.; Stefanick, M.L.; Williams, P.T.; Haskell, W.L. The effects on plasma lipoproteins of a Prudent weight-reducing diet, with or without exercise, in overweight men and women. N. Engl. J. Med. 1991, 325, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Dehghan, M.; Mente, A.; Zhang, X.; Swaminathan, S.; Li, W.; Mohan, V.; Iqbal, R.; Kumar, R.; Wentzel-Viljoen, E.; Rosengren, A.; et al. Associations of fats and carbohydrate intake with cardiovascular disease and mortality in 18 countries from five continents (PURE): A prospective cohort study. Lancet 2017, 390, 2050–2062. [Google Scholar] [CrossRef]
- Ramsden, C.E.; Zamora, D.; Majchrzak-Hong, S.; Faurot, K.R.; Broste, S.K.; Frantz, R.P.; Davis, J.M.; Ringel, A.; Suchindran, C.M.; Hibbeln, J.R. Re-evaluation of the traditional diet-heart hypothesis: Analysis of recovered data from Minnesota Coronary Experiment (1968–1973). BMJ 2016, 353, i1246. [Google Scholar] [CrossRef] [PubMed]
- Ioannidis, J.P.A. The challenge of reforming nutritional epidemiological research. JAMA 2018, 320, 969–970. [Google Scholar] [CrossRef] [PubMed]
- Naska, A.; Lagiou, A.; Lagiou, P. Dietary assessment methods in epidemiological research: Current state of the art and future prospects [version 1; referees: 3 approved]. F1000Research 2017, 6, 926. [Google Scholar] [CrossRef]
- Scalbert, A.; Brennan, L.; Manach, C.; Andres-Lacueva, C.; Dragsted, L.O.; Draper, J.; Rappaport, S.M.; van der Hooft, J.J.; Wishart, D.S. The food metabolome: A window over dietary exposure. Am. J. Clin. Nutr. 2014, 99, 1286–1308. [Google Scholar] [CrossRef]
- Rattray, N.J.W.; Deziel, N.C.; Wallach, J.D.; Khan, S.A.; Vasiliou, V.; Ioannidis, J.P.A.; Johnson, C.H. Beyond genomics: Understanding exposotypes through metabolomics. Hum. Genom. 2018, 12, 4. [Google Scholar] [CrossRef] [PubMed]
- Guasch-Ferre, M.; Bhupathiraiu, S.N.; Hu, F.B. Use of metabolomics in improving assessment of dietary intake. Clin. Chem. 2018, 64, 82–98. [Google Scholar] [CrossRef] [PubMed]
- Brennan, L.; Hu, F.B. Metabolomics-based dietary biomarkers in nutritional epidemiology-Current status and future opportunities. Mol. Nutr. Food Res. 2018, 170, e1701064. [Google Scholar] [CrossRef] [PubMed]
- Brennan, L. Moving toward objective biomarkers of dietary intake. J. Nutr. 2018, 148, 821–822. [Google Scholar] [CrossRef] [PubMed]
- Andersen, M.B.; Rinnan, Å.; Manach, C.; Poulsen, S.K.; Pujos-Guillot, E.; Larsen, T.M.; Astrup, A.; Dragsted, L.O. Untargeted metabolomics as a screening tool for estimating compliance to a dietary pattern. J. Proteome Res. 2014, 13, 1405–1418. [Google Scholar] [CrossRef] [PubMed]
- Playdon, M.C.; Sampson, J.N.; Cross, A.J.; Sinha, R.; Guertin, K.A.; Moy, K.A.; Rothman, N.; Irwin, M.L.; Mayne, S.T.; Stolzenberg-Solomon, R.; et al. Comparing metabolite profiles of habitual diet in serum and urine. Am. J. Clin Nutr. 2016, 104, 776–789. [Google Scholar] [CrossRef] [PubMed]
- Esko, T.; Hirschhorn, J.N.; Feldman, H.A.; Hsu, Y.H.; Deik, A.A.; Clish, C.B.; Ebbeling, C.B.; Ludwig, D.S. Metabolomic profiles as reliable biomarkers of dietary composition. Am. J. Clin. Nutr. 2017, 105, 547–554. [Google Scholar] [CrossRef]
- Gibbons, H.; Carr, E.; McNulty, B.A.; Nugent, A.P.; Walton, J.; Flynn, A.; Gibney, M.J.; Brennan, L. Metabolomic-based identification of clusters that reflect dietary patterns. Mol. Nutr. Food Res. 2017, 61, 1601050. [Google Scholar] [CrossRef]
- Guertin, K.A.; Moore, S.C.; Sampson, J.N.; Huang, W.Y.; Xiao, Q.; Stolzenberg-Solomon, R.Z.; Sinha, R.; Cross, A.J. Metabolomics in nutritional epidemiology: Identifying metabolites associated with diet and quantifying their potential to uncover diet-disease relations in populations. Am. J. Clin. Nutr. 2014, 100, 208–217. [Google Scholar] [CrossRef]
- Hanhineva, K.; Lankinen, M.A.; Pedret, A.; Schwab, U.; Kolehmainen, M.; Paananen, J.; de Mello, V.; Sola, R.; Lehtonen, M.; Poutanen, K.; et al. Nontargeted metabolite profiling discriminates diet-specific biomarkers for consumption of whole grains, fatty fish, and bilberries in a randomized controlled trial. J. Nutr. 2015, 145, 7–17. [Google Scholar] [CrossRef]
- Khakimov, B.; Poulsen, S.K.; Savorani, F.; Acar, E.; Gürdeniz, G.; Larsen, T.M.; Astrup, A.; Dragsted, L.O.; Engelsen, S.B. New Nordic diet versus average Danish diet: A randomized controlled trial revealed healthy long-term effects of the new Nordic diet by GC-MS blood plasma metabolomics. J. Proteome Res. 2016, 15, 1939–1954. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Perez, I.; Posma, J.M.; Gibson, R.; Chambers, E.S.; Hansen, T.H.; Vestergaard, H.; Hansen, T.; Beckmann, M.; Pedersen, O.; Elliott, P.; et al. Objective assessment of dietary patterns by use of metabolic phenotyping: A randomised, controlled, crossover trial. Lancet Diabetes Endocrinol. 2017, 5, 184–195. [Google Scholar] [CrossRef]
- Zulyniak, M.A.; de Souza, R.J.; Mente, A.; Kandasamy, S.; Nundy, M.; Desai, D.; Raman, K.; Hasso, R.; Paré, G.; Beyene, J.; et al. A randomized controlled trial of the effects of a Prudent diet on cardiovascular risk factors, gene expression, and DNA methylation—The Diet and Genetic Intervention (DIGEST) pilot study. BMC Nutr. 2016, 2, 34. [Google Scholar] [CrossRef]
- Hu, F.B.; Rimm, E.B.; Stampfer, M.J.; Ascherio, A.; Spiegelman, D.; Willett, W.C. Prospective study of major dietary patterns and risk of coronary heart disease in men. Am. J. Clin. Nutr. 2000, 72, 912–921. [Google Scholar] [CrossRef] [PubMed]
- Kerver, J.M.; Yang, E.J.; Bianchi, L.; Song, W.O. Dietary patterns associated with risk factors for cardiovascular disease in healthy US adults. Am. J. Clin. Nutr. 2003, 78, 1103–1110. [Google Scholar] [CrossRef] [PubMed]
- Kuehnbaum, N.L.; Kormendi, A.; Britz-McKibbin, P. Multisegment injection-capillary electrophoresis-mass spectrometry: A high-throughput platform for metabolomics with high data fidelity. Anal. Chem. 2013, 85, 10664–10669. [Google Scholar] [CrossRef]
- Yamamoto, M.; Ly, R.; Gill, B.; Zhu, Y.; Moran-Mirabal, J.; Britz-McKibbin, P. Robust and high-throughput method for anionic metabolite profiling: Preventing polyimide aminolysis and capillary breakages under alkaline conditions in capillary electrophoresis-mass spectrometry. Anal. Chem. 2016, 88, 10710–10719. [Google Scholar] [CrossRef] [PubMed]
- Nori de Macedo, A.; Mathiaparanam, S.; Brick, L.; Keenan, K.; Gonska, T.; Pedder, L.; Hill, S.; Britz-McKibbin, P. The sweat metabolome of screen-positive cystic fibrosis infants: Revealing mechanisms beyond impaired chloride transport. ACS Cent. Sci. 2017, 3, 904–913. [Google Scholar] [CrossRef]
- DiBattista, A.; McIntosh, N.; Lamoureux, M.; Al-Dirbashi, O.Y.; Chakraborty, P.; Britz-McKibbin, P. Metabolic signatures of cystic fibrosis identified in dried blood spots for newborn screening without carrier identification. J. Proteome Res. 2019, 18, 841–854. [Google Scholar] [CrossRef]
- Saoi, M.; Percival, M.; Nemr, C.; Li, A.; Gibala, M.J.; Britz-McKibbin, P. Characterization of the human skeletal muscle metabolome for elucidating the mechanisms of bicarbonate ingestion on strenuous interval exercise. Anal. Chem. 2019, 91, 4709–4718. [Google Scholar] [CrossRef]
- Mahieu, N.G.; Patti, G.J. Systems-level annotation of a metabolomics data set reduces 25,000 features to fewer than 1000 unique metabolites. Anal. Chem. 2017, 89, 10397–10406. [Google Scholar] [CrossRef] [PubMed]
- Broadhurst, D.I.; Kell, D.B. Statistical strategies for avoiding false discoveries in metabolomics and related experiments. Metabolomics 2006, 2, 171–196. [Google Scholar] [CrossRef]
- Dunn, W.B.; Erban, A.; Weber, R.J.M.; Creek, D.J.; Brown, M.; Breitling, R.; Hankemeier, T.; Goodacre, R.; Neumann, S.; Kopka, J.; et al. Mass appeal: Metabolite identification in mass spectrometry-focused untargeted metabolomics. Metabolomics 2013, 9, 44–66. [Google Scholar] [CrossRef]
- Allen, F.; Pon, A.; Wilson, M.; Greiner, R.; Wishart, W. CFM-ID: A web server for annotation, spectrum prediction and metabolite identification from tandem mass spectra. Nucleic Acids Res. 2014, 42, W94–W99. [Google Scholar] [CrossRef] [PubMed]
- Jafari, N.; Ahmed, R.; Gloyd, M.; Bloomfield, J.; Britz-McKibbin, P.; Melacini, G. Allosteric sensing of fatty acid binding by NMR: Application to human serum albumin. J. Med. Chem. 2016, 59, 7457–7465. [Google Scholar] [CrossRef]
- Nori de Macedo, A.N.; Teo, K.; Mente, A.; McQueen, M.J.; Zeidler, J.; Poirier, P.; Lear, S.A.; Wielgosz, A.; Britz-McKibbin, P. A robust method for iodine status determination in epidemiological studies by capillary electrophoresis. Anal. Chem. 2014, 86, 10010–10015. [Google Scholar] [CrossRef] [PubMed]
- Wehrens, R.; Hageman, J.A.; van Eeuwijk, F.; Kooke, R.; Flood, P.J.; Wijnker, E.; Keurentjes, J.J.B.; Lommen, A.; van Eekelen, H.D.L.M.; Hall, R.D.; et al. Improved batch correction in untargeted MS-based metabolomics. Metabolomics 2016, 12, 88. [Google Scholar] [CrossRef]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef]
- Tai, B.Y.; Speed, T.P. A multivariate empirical Bayes statistic for replicated microarray time course data. Ann. Stat. 2006, 34, 2387–2412. [Google Scholar] [CrossRef]
- Salek, R.M.; Steinbeck, C.; Viant, M.R.; Goodacre, R.; Dunn, W.B. The role of reporting standards for metabolite annotation and identification in metabolomic studies. Gigascience 2013, 2, 13. [Google Scholar] [CrossRef]
- GBD 2017 Diet Collaborators. Health effects of dietary risks in 195 countries, 1990–2017: A systematic analysis for the global burden of disease study 2017. Lancet 2019, 393, 1958–1972. [Google Scholar] [CrossRef]
- Moore, L.B.; Liu, S.V.; Halliday, T.M.; Neilson, A.P.; Hedrick, V.E.; Davy, B.M. Urinary excretion of sodium, nitrogen, and sugar amounts are valid biomarkers of dietary sodium, protein, and high sugar intake in nonobese adolescents. J. Nutr. 2017, 147, 2364–2373. [Google Scholar] [CrossRef] [PubMed]
- Stark, K.D.; Van Elswyk, M.E.; Higgins, M.R.; Weatherford, C.A.; Salem, N., Jr. Global survey of the omega-3 fatty acids, docosahexaenoic acid and eicosapentaenoic acid in the blood stream of healthy adults. Prog. Lipid Res. 2016, 63, 132–152. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Huybrechts, I.; Vereecken, C.; Mouratidou, T.; Valtuena, J.; Kersting, M.; Gonzalez-Gross, M.; Bolca, S.; Warmberg, J.; Cuenca-Garcia, M.; et al. Dietary fiber intake and its association with indicators of adiposity and serum biomarkers in European adolescents: The HELENA study. J. Nutr. 2015, 54, 771–782. [Google Scholar] [CrossRef] [PubMed]
- Marks, L. Policies for a Prudent diet. Food Policy 1985, 10, 166–174. [Google Scholar] [CrossRef]
- Hu, F.B. Association between dietary patterns and plasma biomarkers of obesity and cardiovascular disease risk. J. Am. Diet. Assoc. 2003, 73, 61–67. [Google Scholar]
- Heinzmann, S.S.; Brown, I.J.; Chan, Q.; Bictash, M.; Dumas, M.E.; Kochhar, S.; Stamler, J.; Holmes, E.; Elliott, P.; Nicholson, J.K. Metabolic profiling strategy for discovery of nutritional biomarkers: Proline betaine as a marker of citrus consumption. Am. J. Clin. Nutr. 2010, 92, 436–443. [Google Scholar] [CrossRef]
- Fung, T.T.; Rimm, E.B.; Spiegelman, D.; Rifai, N.; Tofler, G.H.; Willett Lang, R.; Lang, T.; Bader, M.; Beusch, A.; Schlagbauer, V.; et al. High-throughput quantitation of proline betaine in foods and suitability as a valid biomarker for citrus consumption. J. Agric. Food Chem. 2017, 65, 1613–1619. [Google Scholar]
- Long, C.L.; Dillard, D.R.; Bodzin, J.H.; Geiger, J.W.; Blakemore, W.S. Validity of 3-methylhistidine excretion as an indicator of skeletal muscle protein breakdown in humans. Metabolism 1988, 37, 844–849. [Google Scholar] [CrossRef]
- Kochlik, B.; Gerbracht, C.; Grune, T.; Weber, D. The influence of dietary habits and meat consumption on plasma 3-methylhistidine: A potential marker for muscle protein turnover. Mol. Nutr. Food Res. 2018, 62, e1701062. [Google Scholar] [CrossRef]
- Atkinson, W.; Downer, P.; Lever, M.; Chambers, S.T.; George, P.M. Effects of orange juice and proline betaine on glycine betaine and homocysteine in healthy male subjects. Eur. J. Nutr. 2007, 46, 446–452. [Google Scholar] [CrossRef] [PubMed]
- Sen, N.P.; McGeer, P.L.; Paul, R.M. Imidazolepropionic acid as a urinary metabolite of L-histidine. Biochem. Biophys. Res. Commun. 1962, 9, 257–261. [Google Scholar] [CrossRef]
- Koh, A.; Molinaro, A.; Ståhlman, M.; Khan, M.T.; Schmidt, C.; Mannerås-Holm, L.; Wu, H.; Carreras, A.; Jeong, H.; Olofsson, L.E.; et al. Microbially produced imidazole propionate impairs insulin signaling through mTORC1. Cell 2018, 175, 947–961. [Google Scholar] [CrossRef] [PubMed]
- Kunii, Y.; Otsuka, M.; Kashino, S.; Takeuchi, H.; Ohmori, S. 4-hydroxypipecolic acid and pipecolic acid in acacia species: Their determination by high-performance liquid chromatography, its application to leguminous plants, and configuration of 4-hydroxypipecolic acid. J. Agric. Food Chem. 1996, 44, 483–487. [Google Scholar] [CrossRef]
- Servillo, L.; Giovane, A.; Balestrieri, M.L.; Ferrari, G.; Cautela, D.; Castaldo, D. Occurrence of pipecolic acid and pipecolic acid betaine (homostachydrine) in citrus genus plants. J. Agric. Food Chem. 2012, 60, 315–321. [Google Scholar] [CrossRef]
- Dancis, J.; Hutzler, J. The significance of hyperpipecolatemia in Zellweger syndrome. Am. J. Hum. Genet. 1986, 38, 707–711. [Google Scholar] [PubMed]
- Fujita, T.; Hada, T.; Higashino, K. Origin of D- and L-pipecolic acid in human physiological fluids: A study of the catabolic mechanism to pipecolic acid using the lysine loading test. Clin. Chim. Acta 1999, 287, 145–156. [Google Scholar] [CrossRef]
- Johnson, C.H.; Manna, S.K.; Krausz, K.W.; Bonzo, J.A.; Divelbiss, R.D.; Hollingshead, M.G.; Gonzalez, F.J. Global metabolomics reveals urinary biomarkers of breast cancer in a MCF-7 xenograft mouse model. Metabolites 2013, 3, 658–672. [Google Scholar] [CrossRef]
- Knust, U.; Hull, W.E.; Spiegelhalder, B.; Bartsch, H.; Strowitzki, T.; Owen, R.W. Analysis of enterolignan glucuronides in serum and urine by HPLC-ESI-MS. Food Chem. Toxicol. 2006, 44, 1038–1049. [Google Scholar] [CrossRef]
- Lampe, J.W.; Atkinson, C.; Hullar, M.A. Assessing exposure to lignans and their metabolites in humans. J. AOAC Int. 2006, 89, 1174–1181. [Google Scholar]
- Rodríguez-García, C.; Sánchez-Quesada, C.; Toledo, E.; Delgado-Rodríguez, M.; Gaforio, J.J. Naturally lignan-rich foods: A dietary tool for health promotion? Molecules 2019, 24, 917. [Google Scholar] [CrossRef] [PubMed]
- Khakimov, B.; Jespersen, B.M.; Engelsen, S.B. Comprehensive and comparative metabolomic profiling of wheat, barley, oat and rye using gas chromatography-mass spectrometry and advanced chemometrics. Foods 2014, 3, 569–585. [Google Scholar] [CrossRef] [PubMed]
- Johansson-Persson, A.; Barri, T.; Ulmius, M.; Onning, G.; Dragsted, L.O. LC-QTOF/MS metabolomic profiles in human plasma after a 5-week high dietary fiber intake. Anal. Bioanal. Chem. 2013, 405, 4799–4809. [Google Scholar] [CrossRef] [PubMed]
- Logue, C.; Dowey, L.R.C.; Strain, J.J.; Verhagen, H.; McClean, S.; Gallagher, A.M. Application of liquid chromatography-tandem mass spectrometry to determine urinary concentrations of five commonly used low-calorie sweeteners: A novel biomarker approach for assessing recent intakes? J. Agric. Food Chem. 2017, 65, 4516–4525. [Google Scholar] [CrossRef]
- Hord, N.G.; Tang, Y.; Bryan, N.S. Food sources of nitrates and nitrites: The physiologic context for potential health benefits. Am. J. Clin. Nutr. 2009, 90, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Spritzler, F.A. Low-carbohydrate, whole-foods approach to managing diabetes and prediabetes. Diabetes Spectr. 2012, 25, 238–243. [Google Scholar] [CrossRef][Green Version]
- Nie, C.; He, T.; Zhang, W.; Zhang, G.; Ma, X. Branched-chain amino acids: Beyond nutrition metabolism. Int. J. Mol. Sci. 2018, 19, 954. [Google Scholar] [CrossRef] [PubMed]
- Holeček, M. Branched-chain amino acids in health and disease: Metabolism, alterations in blood plasma, and as supplements. Nutr. Metab. 2018, 15, 33. [Google Scholar] [CrossRef]
- Flores-Guerrero, J.L.; Osté, M.C.J.; Kieneker, L.M.; Gruppen, E.G.; Wolak-Dinsmore, J.; Otvos, J.D.; Connelly, M.A.; Bakker, S.J.L.; Dullaart, R.P.F. Plasma branched-chain amino acids and risk of incident type 2 diabetes: Results from the PREVEND prospective cohort study. J. Clin. Med. 2018, 7, 513. [Google Scholar] [CrossRef]
- Merz, B.; Frommherz, L.; Rist, M.J.; Kulling, S.E.; Bub, A.; Watzl, B. Dietary pattern and plasma BCAA-variations in healthy men and women-Results from the KarMeN study. Nutrients 2018, 10, 623. [Google Scholar] [CrossRef]
- Bouchard-Mercier, A.; Rudkowska, I.; Lemieux, S.; Couture, P.; Vohl, M.-C. The metabolic signature associated with the Western dietary pattern: A cross-sectional study. Nutr. J. 2013, 12, 158. [Google Scholar] [CrossRef] [PubMed]
- Koeth, R.A.; Wang, Z.; Levison, B.S.; Buffa, J.A.; Org, E.; Sheehy, B.T.; Britt, E.B.; Fu, X.; Wu, Y.; Li, L.; et al. Intestinal microbiota metabolism of L-carnitine, A nutrient in red meat, promotes atherosclerosis. Nat. Med. 2013, 19, 576–585. [Google Scholar] [CrossRef] [PubMed]
- Koeth, R.A.; Lam-Galvez, B.R.; Kirsop, J.; Wang, Z.; Levison, B.S.; Gu, X.; Copeland, M.F.; Bartlett, D.; Cody, D.B.; Dai, H.J.; et al. L-carnitine in omnivorous diets induces an atherogenic gut microbial pathway in humans. J. Clin. Investig. 2019, 129, 373–387. [Google Scholar] [CrossRef] [PubMed]
- Fielding, R.; Riede, L.; Lugo, J.P.; Bellamine, A. L-carnitine supplementation in recovery after exercise. Nutrients 2018, 10, 349. [Google Scholar] [CrossRef] [PubMed]
- Patterson, E.; Wall, R.; Fitzgerald, G.F.; Ross, R.P.; Stanton, C. Health implications of high dietary omega-6 polyunsaturated fatty acids. J. Nutr. Metab. 2012, 2012, 539426. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.S.; Sharp, S.J.; Imamura, F.; Koulman, A.; Schulze, M.B.; Ye, Z.; Griffin, J.; Guevara, M.; Huerta, J.M.; Kröger, J.; et al. Association between plasma phospholipid saturated fatty acids and metabolic markers of lipid, hepatic, inflammation and glycaemic pathways in eight european countries: A cross-sectional analysis in the EPIC-InterAct study. BMC Med. 2017, 15, 203. [Google Scholar] [CrossRef] [PubMed]
- Mente, A.; Dehghan, M.; Rangarajan, S.; McQueen, M.; Dagenais, G.; Wielgosz, A.; Lear, S.; Li, W.; Chen, H.; Yi, S.; et al. Association of dietary nutrients with blood lipids and blood pressure in 18 countries: A cross-sectional analysis from the PURE study. Lancet Diabetes Endocrinol. 2017, 5, 774–787. [Google Scholar] [CrossRef]
- Risérus, U.; Marklund, M. Milk fat biomarkers and cardiometabolic disease. Curr. Opin. Lipidol. 2017, 28, 46–51. [Google Scholar] [CrossRef] [PubMed]
- DiNicolantonio, J.J.; O’Keefe, J.H. Omega-6 vegetable oils as a driver of coronary heart disease: The oxidized linoleic acid hypothesis. Open Heart J. 2018, 5, e000898. [Google Scholar] [CrossRef] [PubMed]
- Wanders, A.J.; Zock, P.L.; Brouwer, I.A. Trans fat intake and its dietary sources in general populations worldwide: A systematic review. Nutrients 2017, 9, 840. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diet Category a | W-P, n = 24 | P-W, n = 18 | p for Comparison/Outcome b |
|---|---|---|---|
| Δ Insoluble fiber intake (g/2000 kcal) | (14.0 ± 5.3) | (−5.0 ± 3.5) | p = 1.4 × 10−15; Greater insol. fiber intake in Prudent arm |
| Δ Mg intake (mg/2000 kcal) | (189 ± 89) | (−134 ± 70) | p = 3.5 × 10−15; Greater Mg intake in Prudent arm |
| Δ Fruits + vegetables intake (servings/2000 kcal/day) | (3.6 ± 1.4) | (−1.8 ± 1.3) | p = 7.3 × 10−15; Greater fruits + vegetables intake in Prudent arm |
| Δ Total fiber intake (g/2000 kcal) | (16.6 ± 8.4) | (−13.4 ± 8.1) | p = 5.2 × 10−14; Greater total fiber intake in Prudent arm |
| Δ Energy from sat. fat (%) | (−5.4 ± 3.2) | (4.6 ± 2.4) | p = 1.8 × 10−13; Greater intake of sat. fat in Western arm |
| Δ K intake (mg/2000 kcal) | (1338 ± 617) | (−854 ± 667) | p = 2.5 × 10−13; Greater K intake in Prudent arm |
| Δ Vegetable intake (cup eq./2000 kcal) | (1.8 ± 0.80) | (−0.91 ± 0.92) | p = 2.4 × 10−12; Greater K intake in Prudent arm |
| Δ Vitamin E (mg/2000 kcal) | (7.7 ± 5.3) | (−7.0 ± 4.0) | p = 5.1 × 10−12; Higher intake of vitamin E in Prudent arm |
| Δ Poly:sat (ratio) | (0.47 ± 0.21) | (−0.14 ± 0.18) | p = 8.2 × 10−12; Greater intake of Poly:sat in Prudent arm |
| Δ Vitamin C (mg/2000 kcal) | (149 ± 69) | (−40 ± 54) | p = 1.2 × 10−11; Higher intake of vitamin C in Prudent arm |
| Δ Soluble fiber intake (g/2000 kcal) | (3.9 ± 2.1) | (−1.5 ± 1.5) | p = 2.3 × 10−11; Greater total fiber intake in Prudent arm |
| Δ Fruit intake (cup eq./2000 kcal) | (1.79 ± 0.93) | (−0.92 ± 0.99) | p = 5.9 × 10−11; Greater fruits intake in Prudent arm |
| Δ Energy from fat (%) | (−7.5 ± 5.6) | (5.6 ± 5.6) | p = 9.0 × 10−10; Greater intake of total fat in Western arm |
| Δ Na intake (mg/2000 kcal) | (−694 ± 590) | (754 ± 658) | p = 6.4 × 10−9; Greater Na intake in Western arm |
| Δ Vitamin A (μg/2000 kcal) | (12,973 ± 56,344) | (−7,847 ± 14,060) | p = 1.4 × 10−7; Higher intake of vitamin A in Prudent arm |
| Δ Energy from sugar (%) | (8.9 ± 5.4) | (−1.5 ± 5.8) | p = 7.3 × 10−7; Higher sugar intake in Prudent arm |
| Δ Energy from protein (%) | (1.9 ± 3.6) | (−3.2 ± 2.7) | p = 1.5 × 10−5; Greater intake of protein in Prudent arm |
| Δ Energy from carbohydrates (%) | (8.5 ± 7.8) | (−0.35 ± 5.7) | p = 2.9 × 10−4; Greater intake of total carbs in Prudent arm |
| Δ Cholesterol b (mg/2000 kcal) | (−101 ± 140) | (54 ± 110) | p = 4.8 × 10−4; Greater intake of cholesterol in Western arm |
| Δ Energy from trans fat (%) | (−0.26 ± 0.55) | (0.27 ± 0.23) | p = 6.4 × 10−4; Greater intake of trans fats in Western arm |
| Metabolite/ID | Identifier | T2a | F-value b | p-value b | Rc | p-value c | Food Record d |
|---|---|---|---|---|---|---|---|
| Proline betaine (ProBet) HMDB04827 | 144.102:0.984 (+) MSI-CE-MS C7H13NO2 Level 1 | 24.6 | 8.7 | 0.007 | −0.601 −0.544 −0.528 0.528 0.518 | <0.001 <0.001 0.001 0.001 0.001 | Change %fat trans fat %energy Sat. fat %energy Fruits; Vitamin C Fruits + Vegetables |
| 3-Methylhistidine (Me-His) HMDB00479 | 170.092:0.664 (+) MSI-CE-MS C7H11N2O3 Level 1 | 24.9 | 14.0 | 0.001 | 0.573 0.561 0.553 0.546 0.534 | <0.001 <0.001 <0.001 <0.001 0.001 | Magnesium Protein %energy Insoluble Fiber Potassium Fiber; Fruits + Vegetables |
| Proline (Pro) HMDB00162 | 116.070:0.927 (+) MSI-CE-MS C5H9NO2 Level 1 | 14.6 | 5.9 | 0.020 | 0.495 −0.412 −0.378 −0.362 −0.359 | 0.002 0.010 0.019 0.026 0.027 | trans fat %energy Fruits + Vegetables Vegetables Fruits Protein %energy |
| Carnitine (C0) HMDB00062 | 162.112:0.735 (+) MSI-CE-MS C7H15NO3 Level 1 | 12.2 | 8.9 | 0.005 | −0.464 0.426 −0.404 −0.386 −0.368 | 0.003 0.008 0.012 0.017 0.023 | Poly:sat trans fat %energy Fruits + Vegetables Vitamin E Vitamin C |
| Deoxycarnitine or γ-Butyrobetaine (dC0) HMDB01161 | 146.128:0.700 (+) MSI-CE-MS C7H16NO2 Level 1 | 11.9 | 7.9 | 0.008 | 0.367 0.366 −0.352 0.340 −0.336 | 0.024 0.024 0.030 0.037 0.039 | Change %fat Cholesterol Magnesium Sodium Poly:sat |
| Linoelaidic acid (C18:2n-6trans) HMDB06270 | 294/67.1:15.289 GC-MS C18H32O2 Level 1 | 10.3 | 21.5 | <0.001 | −0.579 −0.555 −0.486 0.485 0.464 | <0.001 <0.001 0.002 0.002 0.003 | Poly:sat Fruits + Vegetables Vitamin C Sat. fat %energy trans fat %energy |
| Pentadecanoic acid (C15:0) HMDB000673 | 294/67.1:14.171 GC-MS C18H32O2 Level 1 | 9.9 | 16.8 | <0.001 | −0.471 0.408 −0.403 −0.379 0.379 | 0.003 0.011 0.012 0.019 0.019 | Poly:sat Change %fat Fruits + Vegetables Vitamin A Change %sat. fat |
| Alanine (Ala) HMDB00161 | 90.056:0.783 (+) MSI-CE-MS C3H7NO2 Level 1 | 9.6 | 6.2 | 0.018 | 0.452 0.439 0.428 −0.395 0.386 | 0.004 0.006 0.007 0.014 0.017 | Change %sat. fat Change %fat trans fat %energy Protein %energy Sat. fat %energy |
| Ketoleucine or 4-Methyl-2-oxopentanoic acid (kLeu) HMDB00695 | 129.056:1.209 (−) MSI-CE-MS C6H10O3 Level 1 | 7.7 | 4.4 | 0.043 | 0.493 −0.459 0.456 0.453 0.452 | 0.002 0.004 0.004 0.004 0.004 | Fruits + Vegetables Sat. fat %energy Fruits Poly:sat Protein %energy |
| 3-Hydroxybutyric acid (OH-BA) HMDB00357 | 103.040:1.043 (−) MSI-CE-MS C4H8O3 Level 1 | 7.6 | 2.9 | 0.097 | 0.437 −0.429 0.425 0.419 0.415 | 0.006 0.007 0.008 0.009 0.01 | Fruits trans fat %energy Poly:sat Vitamin A Fruits + Vegetables |
| α-Linoleic acid (C18:3n-6cis) HMDB001388 | 292/79.1:15.096 GC-MS C18H30O2 Level 1 | 7.0 | 11.6 | 0.002 | −0.441 −0.397 −0.391 0.391 −0.387 | 0.006 0.013 0.015 0.015 0.016 | Poly:sat Vitamin A Fruits + Vegetables trans fat %energy Vitamin E |
| Ketovaline or α-Isovaleric acid (kVal) HMDB00019 | 115.040:1.079 (−) MSI-CE-MS C5H8O3 Level 1 | 6.3 | 2.4 | 0.125 | 0.489 0.472 0.466 0.458 0.451 | 0.002 0.003 0.003 0.004 0.004 | Protein %energy Fiber (total) Fruits + Vegetables Vitamin E Poly:sat |
| Myristic acid (14:0) HMDB00826 | 242/74.1:10.336 GC-MS C15H30O2 Level 1 | 5.0 | 15.2 | <0.001 | −0.535 −0.512 0.503 0.465 −0.463 | 0.001 0.001 0.001 0.003 0.009 | Poly:sat Fruits + Vegetables Change %fat Change %sat. fat Vitamin A |
| Linoleic acid (C18:2n-6cis) HMDB000673 | 294/67.1:14.171 GC-MSC18H32O2 Level 1 | 2.6 | 16.4 | <0.001 | −0.438 0.420 0.412 −0.382 −0.370 | 0.006 0.009 0.005 0.018 0.022 | Poly:sat Change %fat Change %sat. fat Fruits + Vegetables Vitamin A |
| Metabolite/ID | Identifier | T2a | F-test b | p-value b | rc | p-value c | Food Record d |
|---|---|---|---|---|---|---|---|
| 3-Methylhistidine (Me-His) HMDB00479 | 170.092:0.664 (+) MSI-CE-MS C7H11N2O3 Level 1 | 17.9 | 7.8 | 0.008 | 0.524 0.517 0.457 −0.432 0.431 | 0.001 0.001 0.004 0.007 0.007 | Fiber (total) Fruits + Vegetables Vitamin E trans fat %energy Protein %energy |
| 5-Hydroxypipecolic acid (OH-PCA) * HMDB0029246 | 146.081:1.180 (+) MSI-CE-MS C6H11NO3 Level 2 | 16.3 | 1.1 | 0.293 | −0.468 0.397 0.390 0.381 0.374 | 0.003 0.013 0.016 0.018 0.021 | Change fat Fiber (total) Fruits + Vegetables Vitamin E Poly:sat |
| Imidazole propionic acid (ImPA) HMDB02271 | 141.066:0.690 (+) MSI-CE-MS C6H8N2O2 Level 2 | 16.1 | 10.8 | 0.002 | 0.515 0.511 0.471 0.463 0.444 | 0.001 0.001 0.003 0.003 0.005 | Fiber (total) Fruits + Vegetables Protein %energy Vitamin E Poly:sat |
| Proline betaine (ProBet) HMDB04827 | 144.099:0.984 (+) MSI-CE-MS C7H13NO2 Level 1 | 15.5 | 10.8 | 0.002 | 0.487 −0.487 0.482 0.480 0.469 | 0.002 0.002 0.002 0.002 0.003 | Poly:sat trans fat %energy Fiber Fruits + Vegetables Fiber (insoluble) |
| Valinyl-valine (Val-Val) HMDB0029140 | 217.156:0.847 (+) MSI-CE-MS C10H20N2O3 Level 3 | 10.9 | 3.8 | 0.060 | 0.320 0.320 | 0.050 0.050 | Poly:sat Vitamin E |
| Enterolactone glucuronide (ETL-G) HMDB0240377 | 473.145:0.934 (−) MSI-CE-MS C24H25O10 Level 2 | 8.0 | 7.3 | 0.010 | −0.434 0.387 0.340 0.332 0.316 | 0.006 0.016 0.037 0.042 0.054 | Fat Vitamin C Fruits Fruits + Vegetables Vegetables |
| Dihydroxybenzoic acid (DHBA) * HMDB0001856 | 153.019:1.576 (−) MSI-CE-MS C7H6O4 Level 2 | 8.0 | 7.3 | 0.010 | −0.403 0.383 0.355 0.324 0.310 | 0.012 0.018 0.029 0.047 0.058 | Fat Sugar % energy Vitamin C Vegetables Fruits + Vegetables |
| Dimethylglycine (DMG) HMDB0000092 | 104.108:0.569 (+) MSI-CE-MS C4H9NO2 Level 1 | 2.9 | 3.6 | 0.065 | 0.356 0.322 | 0.028 0.049 | Fruits + Vegetables Fiber (total) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wellington, N.; Shanmuganathan, M.; de Souza, R.J.; Zulyniak, M.A.; Azab, S.; Bloomfield, J.; Mell, A.; Ly, R.; Desai, D.; Anand, S.S.; et al. Metabolic Trajectories Following Contrasting Prudent and Western Diets from Food Provisions: Identifying Robust Biomarkers of Short-Term Changes in Habitual Diet. Nutrients 2019, 11, 2407. https://doi.org/10.3390/nu11102407
Wellington N, Shanmuganathan M, de Souza RJ, Zulyniak MA, Azab S, Bloomfield J, Mell A, Ly R, Desai D, Anand SS, et al. Metabolic Trajectories Following Contrasting Prudent and Western Diets from Food Provisions: Identifying Robust Biomarkers of Short-Term Changes in Habitual Diet. Nutrients. 2019; 11(10):2407. https://doi.org/10.3390/nu11102407
Chicago/Turabian StyleWellington, Nadine, Meera Shanmuganathan, Russell J. de Souza, Michael A. Zulyniak, Sandi Azab, Jonathon Bloomfield, Alicia Mell, Ritchie Ly, Dipika Desai, Sonia S. Anand, and et al. 2019. "Metabolic Trajectories Following Contrasting Prudent and Western Diets from Food Provisions: Identifying Robust Biomarkers of Short-Term Changes in Habitual Diet" Nutrients 11, no. 10: 2407. https://doi.org/10.3390/nu11102407
APA StyleWellington, N., Shanmuganathan, M., de Souza, R. J., Zulyniak, M. A., Azab, S., Bloomfield, J., Mell, A., Ly, R., Desai, D., Anand, S. S., & Britz-McKibbin, P. (2019). Metabolic Trajectories Following Contrasting Prudent and Western Diets from Food Provisions: Identifying Robust Biomarkers of Short-Term Changes in Habitual Diet. Nutrients, 11(10), 2407. https://doi.org/10.3390/nu11102407