Tart Cherry Prevents Bone Loss through Inhibition of RANKL in TNF-Overexpressing Mice

Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Diet
2.2. Inflammatory Cytokine Assay
2.3. Microcomputed Tomography (µCT)
2.4. Molecular Assessments
2.5. Mechanical Testing
2.6. Statistical Analyses
3. Results
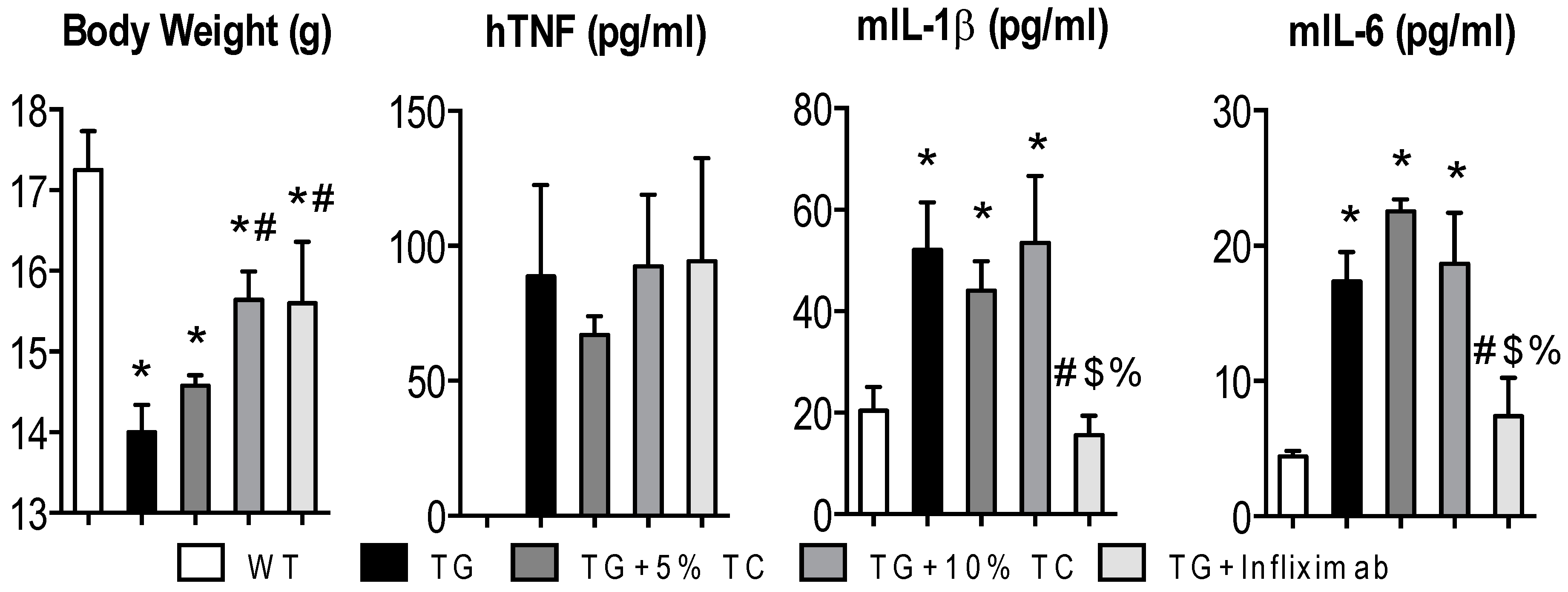
3.1. Although a High-Dose TC Diet Prevents TNF-Mediated Loss of Body Weight, It Does Not Suppress Elevated Proinflammatory Cytokines
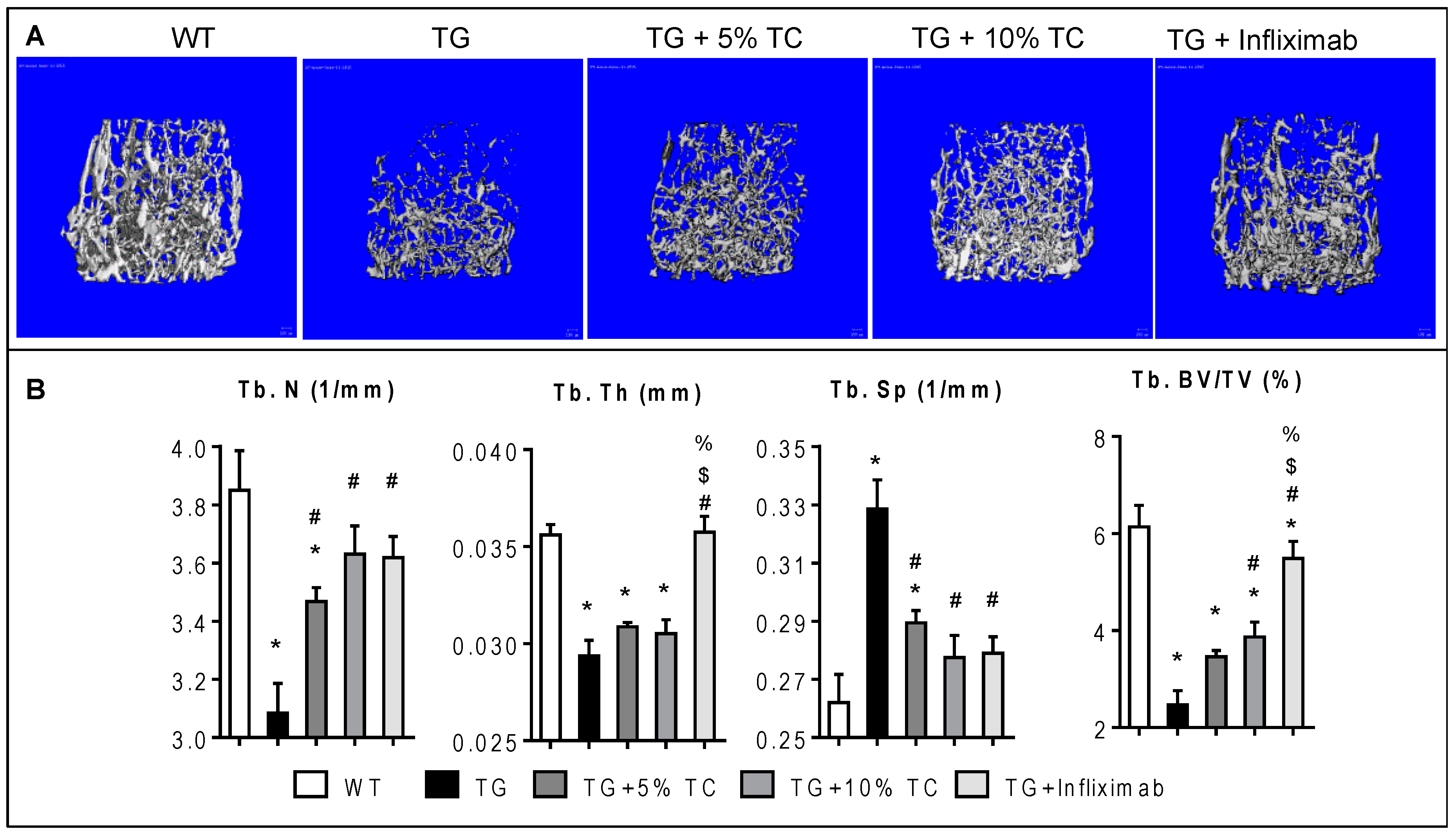
3.2. TC Diet Dose-Dependently Protects Trabecular Bone from Inflammation-Induced Bone Destruction
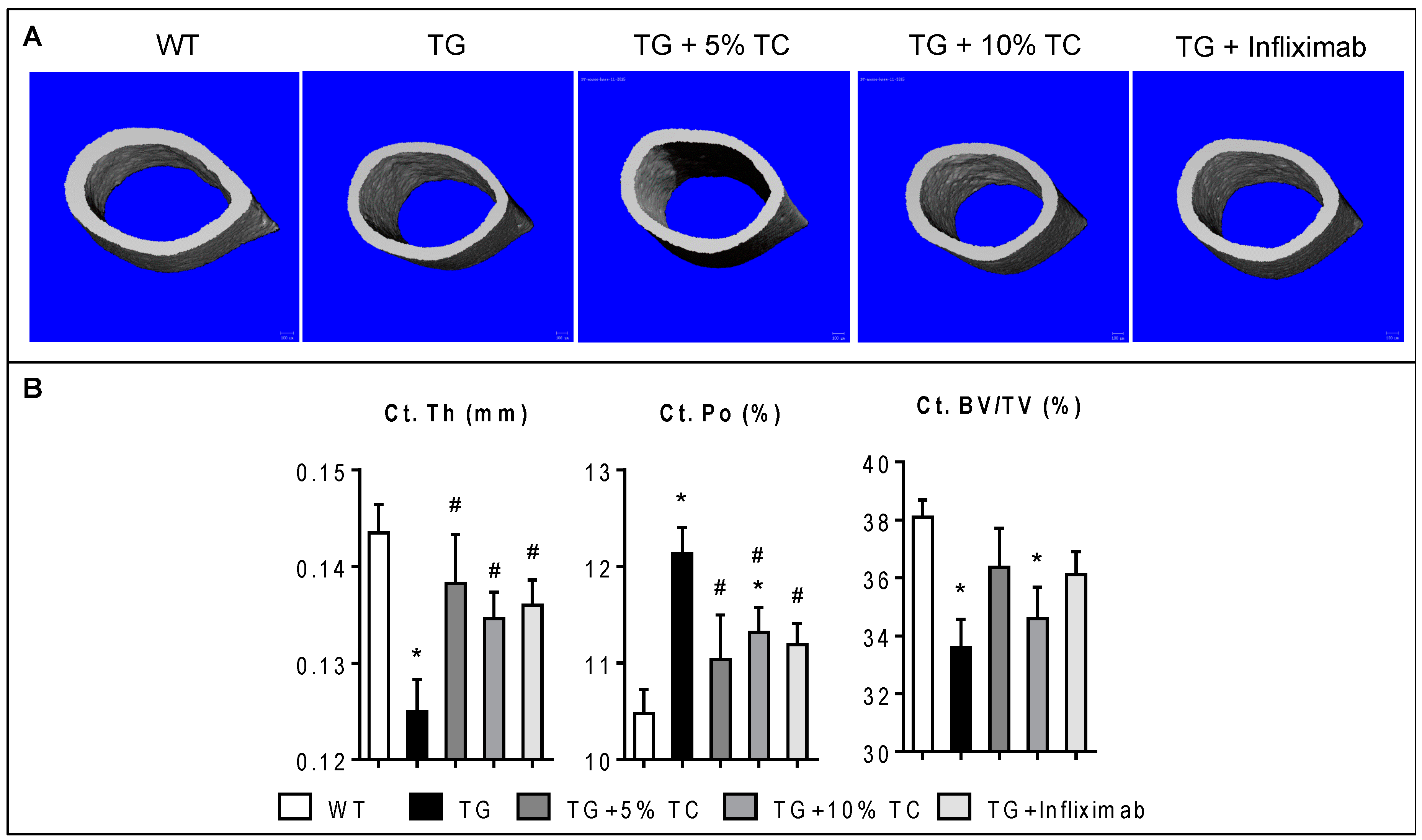
3.3. TC Diet Prevents a TNF-Mediated Reduction of Cortical Thickness and Increase of Cortical Porosity
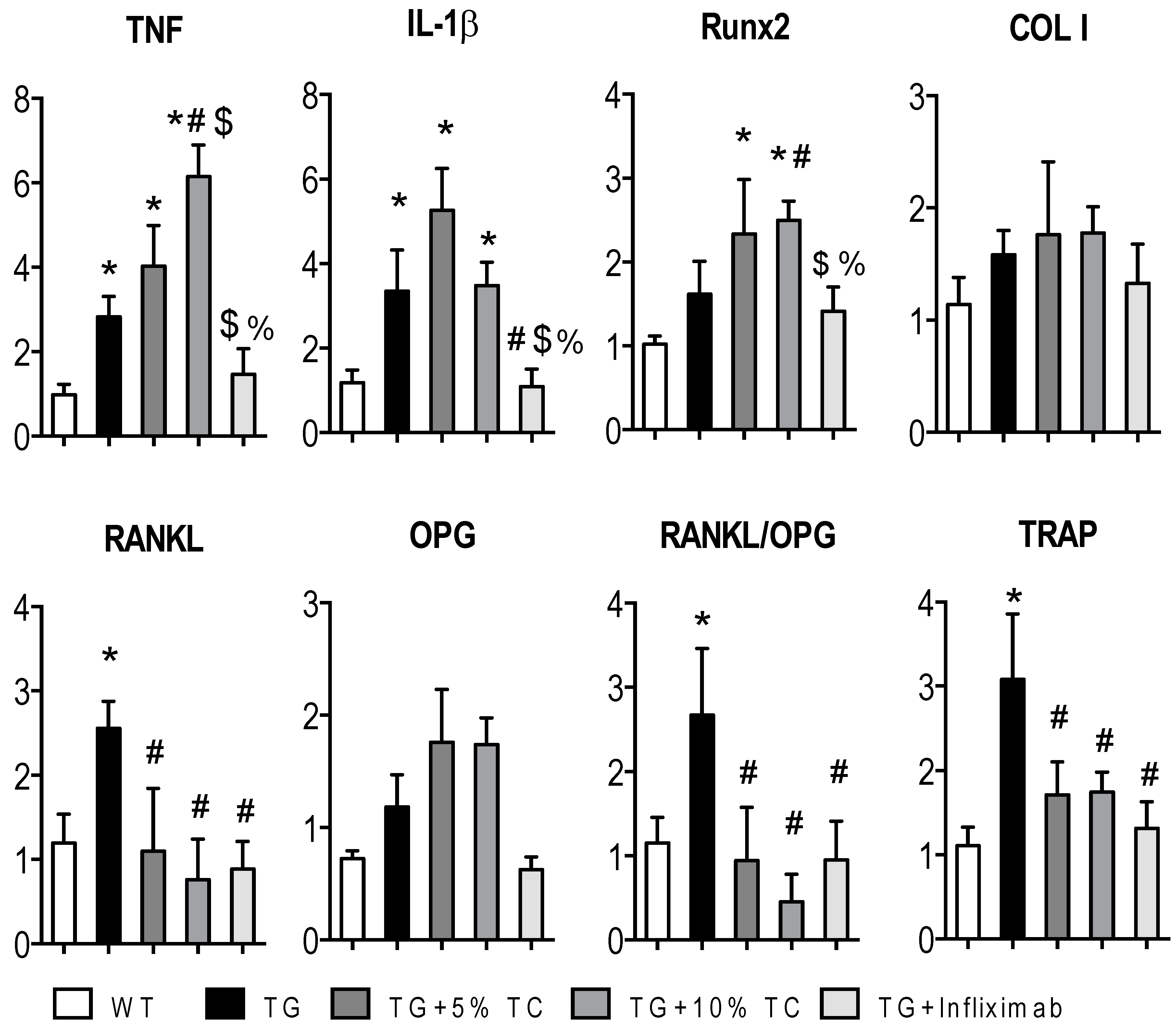
3.4. TC Modulates Gene Expressions Altered by TNF during Bone Destruction
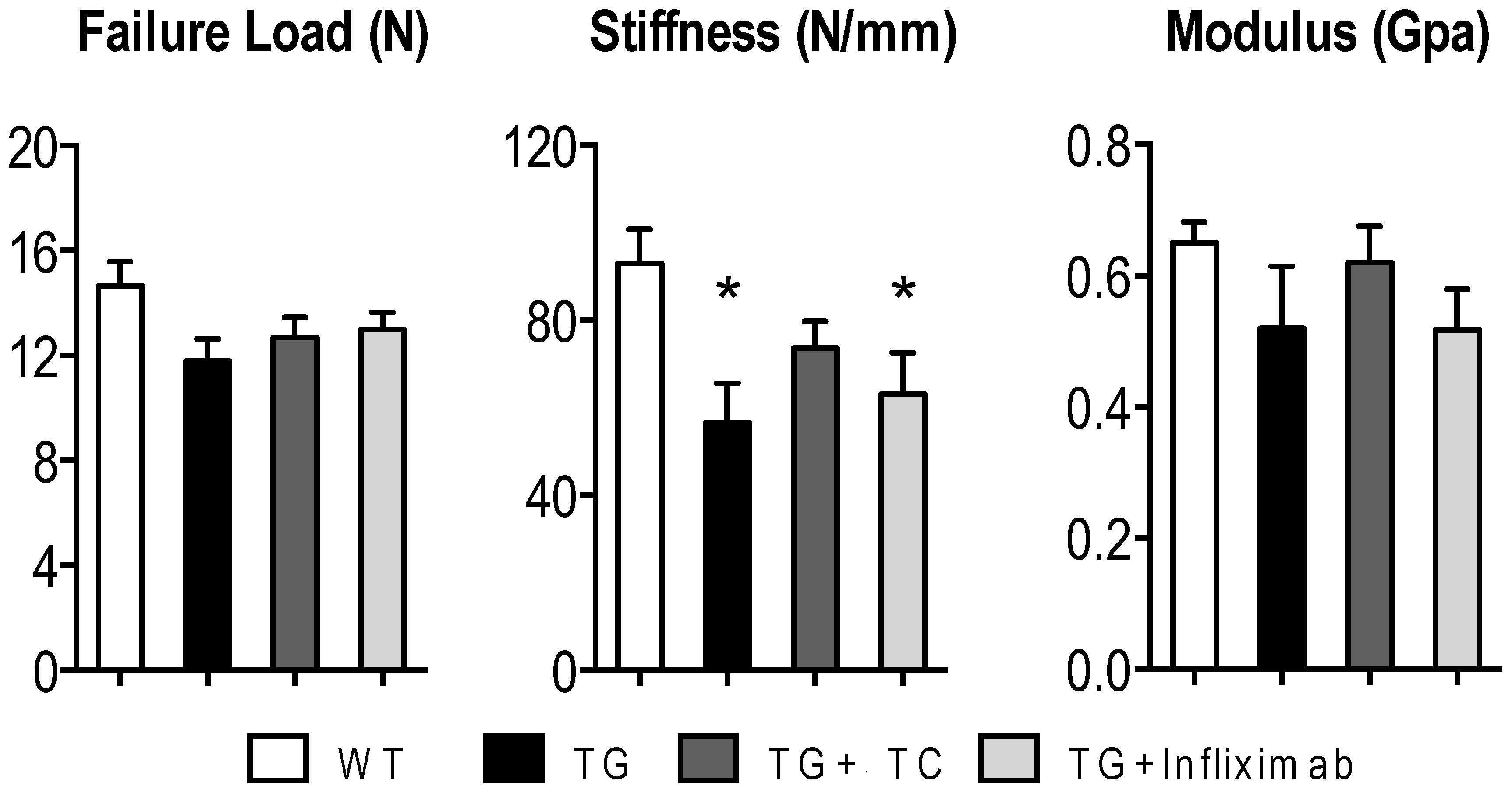
3.5. TC Diet Modestly Improves TNF-Mediated Reduction of Mechanical Strength
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Myasoedova, E.; Crowson, C.S.; Kremers, H.M.; Therneau, T.M.; Gabriel, S.E. Is the incidence of rheumatoid arthritis rising? Results from Olmsted County, Minnesota, 1955–2007. Arthritis Rheum. 2010, 62, 1576–1582. [Google Scholar] [CrossRef] [PubMed]
- Haugeberg, G.; Uhlig, T.; Falch, J.A.; Halse, J.I.; Kvien, T.K. Bone mineral density and frequency of osteoporosis in female patients with rheumatoid arthritis: Results from 394 patients in the Oslo County Rheumatoid Arthritis register. Arthritis Rheum. 2000, 43, 522–530. [Google Scholar] [CrossRef]
- Rossini, M.; Bagnato, G.; Frediani, B.; Iagnocco, A.; G, L.A.M.; Minisola, G.; Caminiti, M.; Varenna, M.; Adami, S. Relationship of focal erosions, bone mineral density, and parathyroid hormone in rheumatoid arthritis. J. Rheumatol. 2011, 38, 997–1002. [Google Scholar] [CrossRef] [PubMed]
- Forsblad-d’Elia, H.; Carlsten, H. Hormone replacement therapy in postmenopausal women with rheumatoid arthritis stabilises bone mineral density by digital X-ray radiogrammetry in a randomised controlled trial. Ann. Rheum. Dis. 2011, 70, 1167–1168. [Google Scholar] [CrossRef] [PubMed]
- Wright, N.C.; Lisse, J.R.; Walitt, B.T.; Eaton, C.B.; Chen, Z.; Women’s Health Initiative, I. Arthritis increases the risk for fractures—Results from the Women’s Health Initiative. J. Rheumatol. 2011, 38, 1680–1688. [Google Scholar] [CrossRef] [PubMed]
- van Staa, T.P.; Geusens, P.; Bijlsma, J.W.; Leufkens, H.G.; Cooper, C. Clinical assessment of the long-term risk of fracture in patients with rheumatoid arthritis. Arthritis Rheum. 2006, 54, 3104–3112. [Google Scholar] [CrossRef] [PubMed]
- Vis, M.; Haavardsholm, E.A.; Boyesen, P.; Haugeberg, G.; Uhlig, T.; Hoff, M.; Woolf, A.; Dijkmans, B.; Lems, W.; Kvien, T.K. High incidence of vertebral and non-vertebral fractures in the OSTRA cohort study: A 5-year follow-up study in postmenopausal women with rheumatoid arthritis. Osteoporos. Int. 2011, 22, 2413–2419. [Google Scholar] [CrossRef]
- Wijbrandts, C.A.; Klaasen, R.; Dijkgraaf, M.G.; Gerlag, D.M.; van Eck-Smit, B.L.; Tak, P.P. Bone mineral density in rheumatoid arthritis patients 1 year after adalimumab therapy: Arrest of bone loss. Ann. Rheum. Dis. 2009, 68, 373–376. [Google Scholar] [CrossRef] [PubMed]
- Marotte, H.; Pallot-Prades, B.; Grange, L.; Gaudin, P.; Alexandre, C.; Miossec, P. A 1-year case-control study in patients with rheumatoid arthritis indicates prevention of loss of bone mineral density in both responders and nonresponders to infliximab. Arthritis Res. 2007, 9, R61. [Google Scholar] [CrossRef]
- Hoff, M.; Kvien, T.K.; Kalvesten, J.; Elden, A.; Kavanaugh, A.; Haugeberg, G. Adalimumab reduces hand bone loss in rheumatoid arthritis independent of clinical response: Subanalysis of the PREMIER study. BMC Musculoskelet. Disord. 2011, 12, 54. [Google Scholar] [CrossRef]
- Krieckaert, C.L.; Nurmohamed, M.T.; Wolbink, G.; Lems, W.F. Changes in bone mineral density during long-term treatment with adalimumab in patients with rheumatoid arthritis: A cohort study. Rheumatology 2013, 52, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Saidenberg-Kermanach, N.; Corrado, A.; Lemeiter, D.; deVernejoul, M.C.; Boissier, M.C.; Cohen-Solal, M.E. TNF-alpha antibodies and osteoprotegerin decrease systemic bone loss associated with inflammation through distinct mechanisms in collagen-induced arthritis. Bone 2004, 35, 1200–1207. [Google Scholar] [CrossRef] [PubMed]
- Redlich, K.; Gortz, B.; Hayer, S.; Zwerina, J.; Doerr, N.; Kostenuik, P.; Bergmeister, H.; Kollias, G.; Steiner, G.; Smolen, J.S.; et al. Repair of local bone erosions and reversal of systemic bone loss upon therapy with anti-tumor necrosis factor in combination with osteoprotegerin or parathyroid hormone in tumor necrosis factor-mediated arthritis. Am. J. Pathol. 2004, 164, 543–555. [Google Scholar] [CrossRef]
- Mirza, F.; Lorenzo, J.; Drissi, H.; Lee, F.Y.; Soung, D.Y. Dried plum alleviates symptoms of inflammatory arthritis in TNF transgenic mice. J. Nutr. Biochem. 2018, 52, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Engvall, I.L.; Tengstrand, B.; Brismar, K.; Hafstrom, I. Infliximab therapy increases body fat mass in early rheumatoid arthritis independently of changes in disease activity and levels of leptin and adiponectin: A randomised study over 21 months. Arthritis Res. 2010, 12, R197. [Google Scholar] [CrossRef] [PubMed]
- Serelis, J.; Kontogianni, M.D.; Katsiougiannis, S.; Bletsa, M.; Tektonidou, M.G.; Skopouli, F.N. Effect of anti-TNF treatment on body composition and serum adiponectin levels of women with rheumatoid arthritis. Clin. Rheumatol. 2008, 27, 795–797. [Google Scholar] [CrossRef] [PubMed]
- Saag, K.G.; Teng, G.G.; Patkar, N.M.; Anuntiyo, J.; Finney, C.; Curtis, J.R.; Paulus, H.E.; Mudano, A.; Pisu, M.; Elkins-Melton, M.; et al. American College of Rheumatology 2008 recommendations for the use of nonbiologic and biologic disease-modifying antirheumatic drugs in rheumatoid arthritis. Arthritis Rheum. 2008, 59, 762–784. [Google Scholar] [CrossRef] [PubMed]
- Bonerz, D.; Wurth, K.; Dietrich, H.; Will, F. Analytical characterization and the impact of ageing on anthocyanin composition and degradation in juices from five sour cherry cultivars. Eur. Food Res. Technol. 2007, 224, 355–364. [Google Scholar] [CrossRef]
- Kirakosyan, A.; Seymour, E.M.; Llanes, D.E.U.; Kaufman, P.B.; Bolling, S.F. Chemical profile and antioxidant capacities of tart cherry products. Food Chem. 2009, 115, 20–25. [Google Scholar] [CrossRef]
- Bell, P.G.; Walshe, I.H.; Davison, G.W.; Stevenson, E.; Howatson, G. Montmorency cherries reduce the oxidative stress and inflammatory responses to repeated days high-intensity stochastic cycling. Nutrients 2014, 6, 829–843. [Google Scholar] [CrossRef] [PubMed]
- Ou, B.; Bosak, K.N.; Brickner, P.R.; Iezzoni, D.G.; Seymour, E.M. Processed tart cherry products--comparative phytochemical content, in vitro antioxidant capacity and in vitro anti-inflammatory activity. J. Food Sci. 2012, 77, H105–H112. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Nair, M.G.; Claycombe, K.J. Synergistic inhibition of interleukin-6 production in adipose stem cells by tart cherry anthocyanins and atorvastatin. Phytomedicine 2012, 19, 878–881. [Google Scholar] [CrossRef] [PubMed]
- Traustadottir, T.; Davies, S.S.; Stock, A.A.; Su, Y.; Heward, C.B.; Roberts, L.J., 2nd; Harman, S.M. Tart cherry juice decreases oxidative stress in healthy older men and women. J. Nutr. 2009, 139, 1896–1900. [Google Scholar] [CrossRef] [PubMed]
- Haidari, F., Jr.; Mohammad Shahi, M.; Keshavarz, S.A.; Rashidi, M.R. Inhibitory Effects of Tart Cherry (Prunus cerasus) Juice on Xanthine Oxidoreductase Activity and its Hypouricemic and Antioxidant Effects on Rats. Malays. J. Nutr. 2009, 15, 53–64. [Google Scholar] [PubMed]
- Tall, J.M.; Seeram, N.P.; Zhao, C.; Nair, M.G.; Meyer, R.A.; Raja, S.N. Tart cherry anthocyanins suppress inflammation-induced pain behavior in rat. Behav. Brain Res. 2004, 153, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Seeram, N.P.; Momin, R.A.; Nair, M.G.; Bourquin, L.D. Cyclooxygenase inhibitory and antioxidant cyanidin glycosides in cherries and berries. Phytomedicine 2001, 8, 362–369. [Google Scholar] [CrossRef]
- Wang, H.; Nair, M.G.; Strasburg, G.M.; Booren, A.M.; Gray, J.I. Antioxidant polyphenols from tart cherries (Prunus cerasus). J. Agric. Food Chem. 1999, 47, 840–844. [Google Scholar] [CrossRef]
- Wang, H.; Nair, M.G.; Strasburg, G.M.; Booren, A.M.; Gray, J.I. Novel antioxidant compounds from tart cherries (Prunus cerasus). J. Nat. Prod. 1999, 62, 86–88. [Google Scholar] [CrossRef]
- Schumacher, H.R.; Pullman-Mooar, S.; Gupta, S.R.; Dinnella, J.E.; Kim, R.; McHugh, M.P. Randomized double-blind crossover study of the efficacy of a tart cherry juice blend in treatment of osteoarthritis (OA) of the knee. Osteoarthr. Cartil. 2013, 21, 1035–1041. [Google Scholar] [CrossRef]
- Cho, M.L.; Heo, Y.J.; Park, M.K.; Oh, H.J.; Park, J.S.; Woo, Y.J.; Ju, J.H.; Park, S.H.; Kim, H.Y.; Min, J.K. Grape seed proanthocyanidin extract (GSPE) attenuates collagen-induced arthritis. Immunol. Lett. 2009, 124, 102–110. [Google Scholar] [CrossRef]
- He, Y.H.; Xiao, C.; Wang, Y.S.; Zhao, L.H.; Zhao, H.Y.; Tong, Y.; Zhou, J.; Jia, H.W.; Lu, C.; Li, X.M.; et al. Antioxidant and anti-inflammatory effects of cyanidin from cherries on rat adjuvant-induced arthritis. Zhongguo Zhong Yao Za Zhi 2005, 30, 1602–1605. [Google Scholar] [PubMed]
- Choi, E.J.; Bae, S.C.; Yu, R.; Youn, J.; Sung, M.K. Dietary vitamin E and quercetin modulate inflammatory responses of collagen-induced arthritis in mice. J. Med. Food 2009, 12, 770–775. [Google Scholar] [CrossRef] [PubMed]
- Keffer, J.; Probert, L.; Cazlaris, H.; Georgopoulos, S.; Kaslaris, E.; Kioussis, D.; Kollias, G. Transgenic mice expressing human tumour necrosis factor: A predictive genetic model of arthritis. EMBO J. 1991, 10, 4025–4031. [Google Scholar] [CrossRef]
- Schett, G.; Redlich, K.; Hayer, S.; Zwerina, J.; Bolon, B.; Dunstan, C.; Gortz, B.; Schulz, A.; Bergmeister, H.; Kollias, G.; et al. Osteoprotegerin protects against generalized bone loss in tumor necrosis factor-transgenic mice. Arthritis Rheum. 2003, 48, 2042–2051. [Google Scholar] [CrossRef] [PubMed]
- Bohm, C.; Derer, A.; Axmann, R.; Hillienhoff, U.; Zaiss, M.M.; Luther, J.; Zech, C.; Stock, M.; Scholtysek, C.; Engelke, K.; et al. RSK2 protects mice against TNF-induced bone loss. J. Cell. Sci. 2012, 125, 2160–2171. [Google Scholar] [CrossRef] [PubMed]
- Zwerina, J.; Redlich, K.; Polzer, K.; Joosten, L.; Kronke, G.; Distler, J.; Hess, A.; Pundt, N.; Pap, T.; Hoffmann, O.; et al. TNF-induced structural joint damage is mediated by IL-1. Proc. Natl. Acad. Sci. USA 2007, 104, 11742–11747. [Google Scholar] [CrossRef]
- Stamer, D.K.; Nizami, S.A.; Lee, F.Y.; Soung D, Y. Whole grape alleviates inflammatory arthritis through inhibition of tumor necrosis factor. J. Funct. Foods 2017, 35, 458–565. [Google Scholar] [CrossRef]
- Soung do, Y.; Kalinowski, J.; Baniwal, S.K.; Jacome-Galarza, C.E.; Frenkel, B.; Lorenzo, J.; Drissi, H. Runx1-mediated regulation of osteoclast differentiation and function. Mol. Endocrinol. 2014, 28, 546–553. [Google Scholar] [CrossRef]
- Soung do, Y.; Gentile, M.A.; Duong le, T.; Drissi, H. Effects of pharmacological inhibition of cathepsin K on fracture repair in mice. Bone 2013, 55, 248–255. [Google Scholar] [CrossRef]
- Tascau, L.; Gardner, T.; Anan, H.; Yongpravat, C.; Cardozo, C.P.; Bauman, W.A.; Lee, F.Y.; Oh, D.S.; Tawfeek, H.A. Activation of Protein Kinase A in Mature Osteoblasts Promotes a Major Bone Anabolic Response. Endocrinology 2016, 157, 112–126. [Google Scholar] [CrossRef]
- Nakamura, Y.; Suzuki, T.; Kato, H. Denosumab significantly improves bone mineral density with or without bisphosphonate pre-treatment in osteoporosis with rheumatoid arthritis: Denosumab improves bone mineral density in osteoporosis with rheumatoid arthritis. Arch. Osteoporos. 2017, 12, 80. [Google Scholar] [CrossRef] [PubMed]
- Mochizuki, T.; Yano, K.; Ikari, K.; Kawakami, K.; Hiroshima, R.; Koenuma, N.; Ishibashi, M.; Momohara, S. Effects of denosumab treatment on bone mineral density and joint destruction in patients with rheumatoid arthritis. J. Bone Miner. MeTable 2018, 36, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Ebina, K.; Hirao, M.; Hashimoto, J.; Hagihara, K.; Kashii, M.; Kitaguchi, K.; Matsuoka, H.; Iwahashi, T.; Chijimatsu, R.; Yoshikawa, H. Assessment of the effects of switching oral bisphosphonates to denosumab or daily teriparatide in patients with rheumatoid arthritis. J. Bone Miner. MeTable 2018, 36, 478–487. [Google Scholar] [CrossRef] [PubMed]
- Martin, K.R.; Burrell, L. 100% Tart cherry juice reduces pro-inflammatory biomarkers in overweight and obese subjects. FASEB J. 2010, 24, 15. [Google Scholar]
- Seymour, E.M.; Lewis, S.K.; Urcuyo-Llanes, D.E.; Tanone, II.; Kirakosyan, A.; Kaufman, P.B.; Bolling, S.F. Regular tart cherry intake alters abdominal adiposity, adipose gene transcription, and inflammation in obesity-prone rats fed a high fat diet. J. Med. Food 2009, 12, 935–942. [Google Scholar] [CrossRef] [PubMed]
- Pang, J.L.; Ricupero, D.A.; Huang, S.; Fatma, N.; Singh, D.P.; Romero, J.R.; Chattopadhyay, N. Differential activity of kaempferol and quercetin in attenuating tumor necrosis factor receptor family signaling in bone cells. Biochem. Pharmacol. 2006, 71, 818–826. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Park, M.K.; Oh, H.J.; Woo, Y.J.; Lim, M.A.; Lee, J.H.; Ju, J.H.; Jung, Y.O.; Lee, Z.H.; Park, S.H.; et al. Grape-seed proanthocyanidin extract as suppressors of bone destruction in inflammatory autoimmune arthritis. PLoS ONE 2012, 7, e51377. [Google Scholar] [CrossRef] [PubMed]
- Moriwaki, S.; Suzuki, K.; Muramatsu, M.; Nomura, A.; Inoue, F.; Into, T.; Yoshiko, Y.; Niida, S. Delphinidin, one of the major anthocyanidins, prevents bone loss through the inhibition of excessive osteoclastogenesis in osteoporosis model mice. PLoS ONE 2014, 9, e97177. [Google Scholar] [CrossRef] [PubMed]
- Atkins, G.J.; Welldon, K.J.; Halbout, P.; Findlay, D.M. Strontium ranelate treatment of human primary osteoblasts promotes an osteocyte-like phenotype while eliciting an osteoprotegerin response. Osteoporos. Int. 2009, 20, 653–664. [Google Scholar] [CrossRef] [PubMed]
- Brennan, T.C.; Rybchyn, M.S.; Green, W.; Atwa, S.; Conigrave, A.D.; Mason, R.S. Osteoblasts play key roles in the mechanisms of action of strontium ranelate. Br. J. Pharmacol. 2009, 157, 1291–1300. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Schneeweiss, S.; Liu, J.; Solomon, D.H. Effects of disease-modifying antirheumatic drugs on nonvertebral fracture risk in rheumatoid arthritis: A population-based cohort study. J. Bone Miner. Res. 2012, 27, 789–796. [Google Scholar] [CrossRef] [PubMed]
- Hsu, H.; Lacey, D.L.; Dunstan, C.R.; Solovyev, I.; Colombero, A.; Timms, E.; Tan, H.L.; Elliott, G.; Kelley, M.J.; Sarosi, I.; et al. Tumor necrosis factor receptor family member RANK mediates osteoclast differentiation and activation induced by osteoprotegerin ligand. Proc. Natl. Acad. Sci. USA 1999, 96, 3540–3545. [Google Scholar] [CrossRef] [PubMed]
- McClung, M.R.; Lewiecki, E.M.; Cohen, S.B.; Bolognese, M.A.; Woodson, G.C.; Moffett, A.H.; Peacock, M.; Miller, P.D.; Lederman, S.N.; Chesnut, C.H.; et al. Denosumab in postmenopausal women with low bone mineral density. N. Engl. J. Med. 2006, 354, 821–831. [Google Scholar] [CrossRef]
- Yamane, H.; Sakai, A.; Mori, T.; Tanaka, S.; Moridera, K.; Nakamura, T. The anabolic action of intermittent PTH in combination with cathepsin K inhibitor or alendronate differs depending on the remodeling status in bone in ovariectomized mice. Bone 2009, 44, 1055–1062. [Google Scholar] [CrossRef]
- Park-Wyllie, L.Y.; Mamdani, M.M.; Juurlink, D.N.; Hawker, G.A.; Gunraj, N.; Austin, P.C.; Whelan, D.B.; Weiler, P.J.; Laupacis, A. Bisphosphonate use and the risk of subtrochanteric or femoral shaft fractures in older women. JAMA 2011, 305, 783–789. [Google Scholar] [CrossRef] [PubMed]
- Odvina, C.V.; Zerwekh, J.E.; Rao, D.S.; Maalouf, N.; Gottschalk, F.A.; Pak, C.Y. Severely suppressed bone turnover: A potential complication of alendronate therapy. J. Clin. Endocrinol. MeTable 2005, 90, 1294–1301. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | 1 0% | 5% | 10% | |
|---|---|---|---|---|
| g/kg Diet | ||||
| TC Carbohydrates Proteins Fat Fiber 2 Mineral mix 3 Vitamin mix Choline bitartrate L-cystine 4 TBHQ, antioxidants | Cornstarch Maltodextrin Sucrose Casein Soybean oil Cellulose | 0 397.5 132 100.00 200 70.00 50.00 35 10 2.5 3 0.014 | 50 391.3 132 66.36 194.8 69.59 45.49 35 10 2.5 3 0.014 | 100 385.1 132 32.72 189.5 69.18 40.97 35 10 2.5 3 0.014 |
| Calculated nutrients | ||||
| Kcal/g | 4 | 4 | 4 | |
| Macronutrients, % by weight | ||||
| Protein Carbohydrate Fat Sugar (sucrose + TC) | 18 60 7 13 | 18 60 7 13 | 18 60 7 13 | |
| Target Name | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| TNF IL-1β Runx2 COL I RANKL OPG TRAP GAPDH | CCCCAAAGGGATGAGAAGTT ATGAAGGGCTGCTTCCAAA CCACCACTCACTACCACACG ACTGGTACATCAGCCCGAAC CAGAAGGAACTGCAACACATTG GTGTGGAATAGATGTCACCCT GATGACTTTGCCAGTCAGCA GGTCGGTGTGAACGGATTT | GGTCTGGGCCATAGAACTGA GGACAGCCCAGGTCAAAG CACTCTGGCTTTGGGAAGAG AATCCATCGGTCATGCTCTC CTCCTGAGAAGCGCTGTG AAGAAGGCCTCTTCACACAG AACTGCTTTTTGAGCCAGGA GACCAGGCGCCCAATAC |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moon, N.; Effiong, L.; Song, L.; Gardner, T.R.; Soung, D.Y. Tart Cherry Prevents Bone Loss through Inhibition of RANKL in TNF-Overexpressing Mice. Nutrients 2019, 11, 63. https://doi.org/10.3390/nu11010063
Moon N, Effiong L, Song L, Gardner TR, Soung DY. Tart Cherry Prevents Bone Loss through Inhibition of RANKL in TNF-Overexpressing Mice. Nutrients. 2019; 11(1):63. https://doi.org/10.3390/nu11010063
Chicago/Turabian StyleMoon, Nicholas, Linda Effiong, Lee Song, Thomas R. Gardner, and Do Y. Soung. 2019. "Tart Cherry Prevents Bone Loss through Inhibition of RANKL in TNF-Overexpressing Mice" Nutrients 11, no. 1: 63. https://doi.org/10.3390/nu11010063
APA StyleMoon, N., Effiong, L., Song, L., Gardner, T. R., & Soung, D. Y. (2019). Tart Cherry Prevents Bone Loss through Inhibition of RANKL in TNF-Overexpressing Mice. Nutrients, 11(1), 63. https://doi.org/10.3390/nu11010063