Improved Preventive Effects of Combined Bioactive Compounds Present in Different Blueberry Varieties as Compared to Single Phytochemicals



Abstract
1. Introduction
2. Material and Methods
2.1. Cell Culture
2.2. Preparation of Blueberry Extracts and Selection of Single Compounds
2.3. Treatment and Isolation of Caco-2 Cells
2.4. Cytotoxicity Assay
2.5. Alkaline Single-Cell Gel Electrophoresis (Comet Assay)
2.6. Electron Spin Resonance Spectroscopy Measurements
2.7. Real-Time qPCR
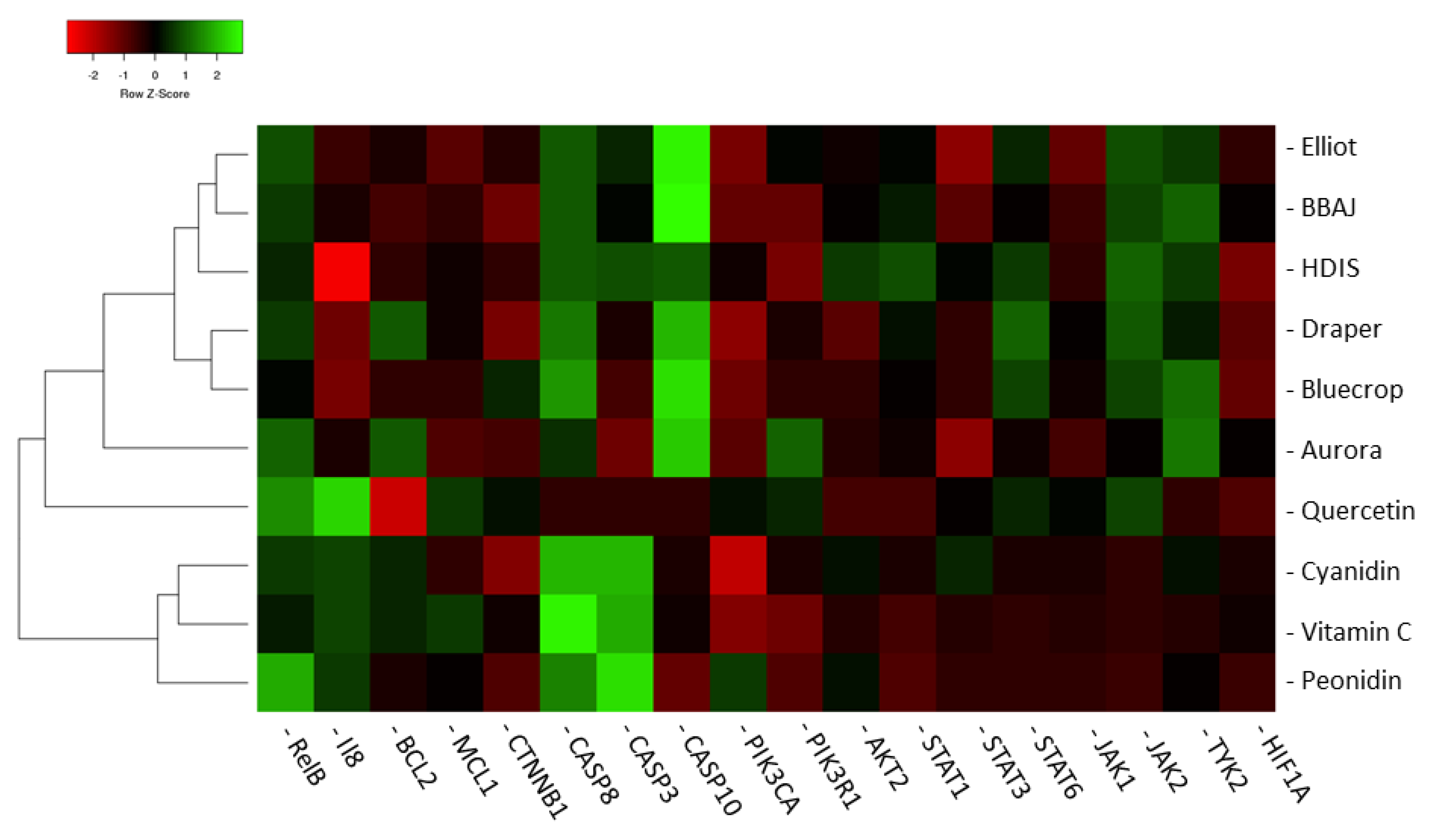
2.8. Hierarchical Clustering Analyses
2.9. Statistical Analyses
3. Results
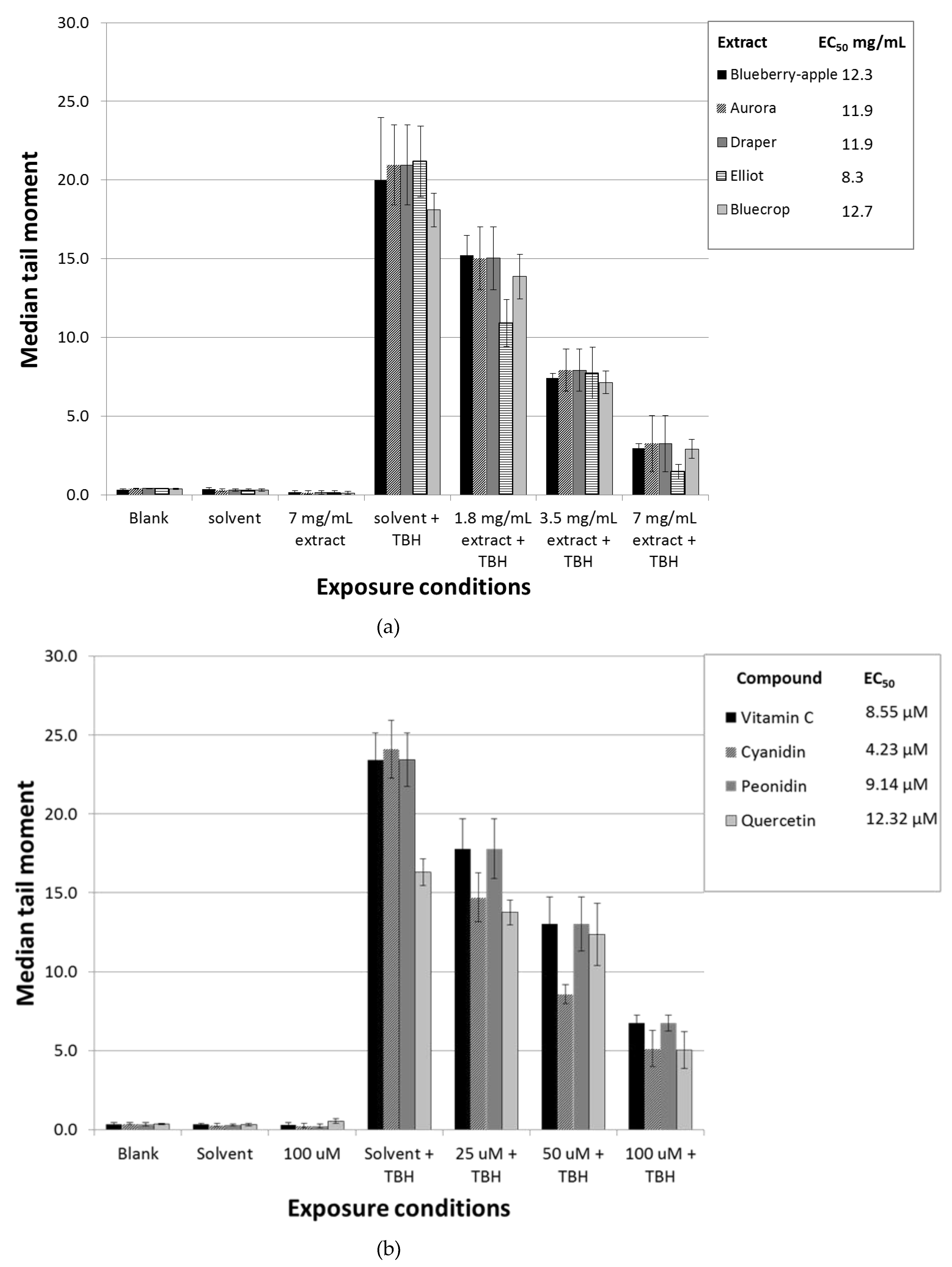
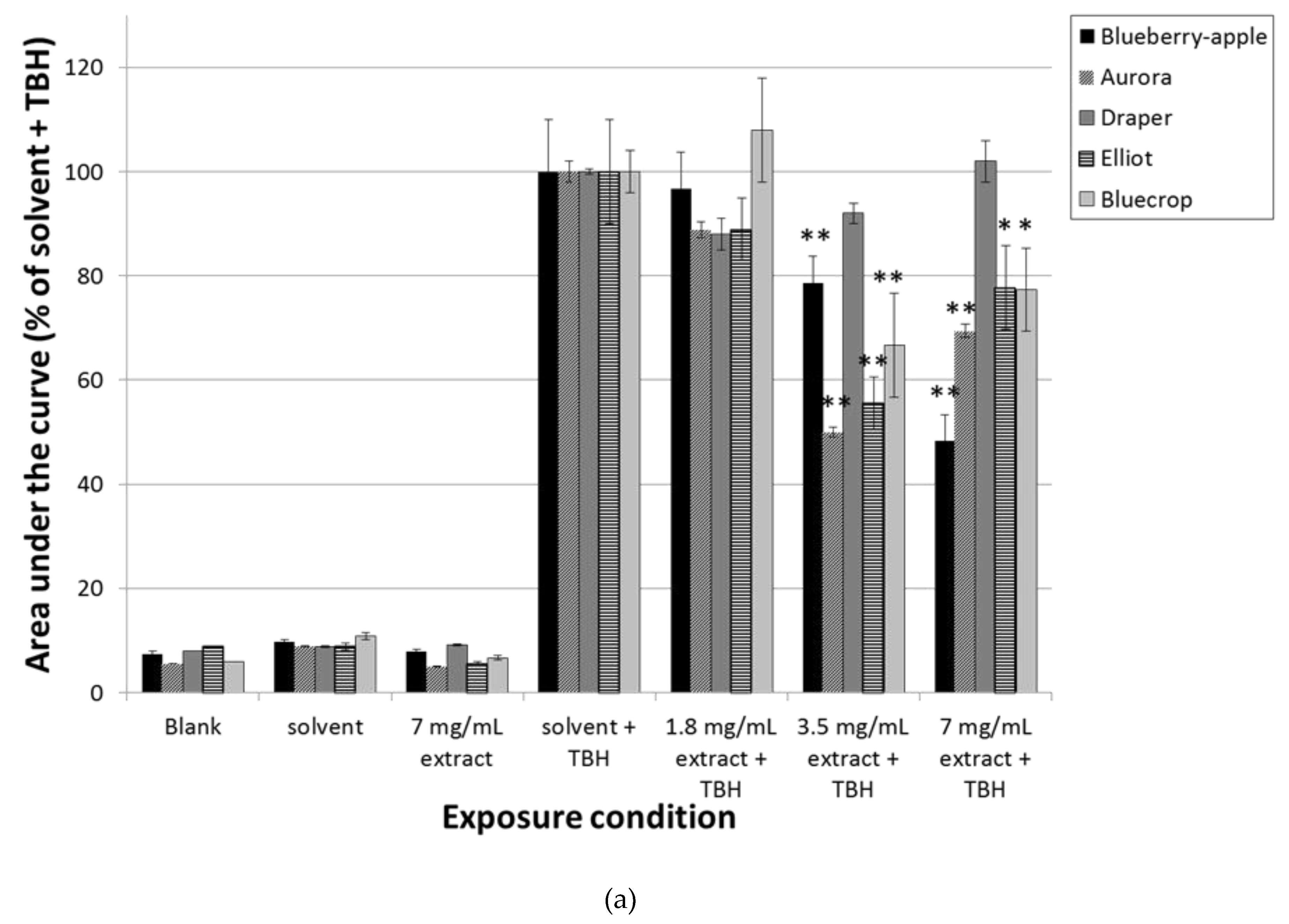
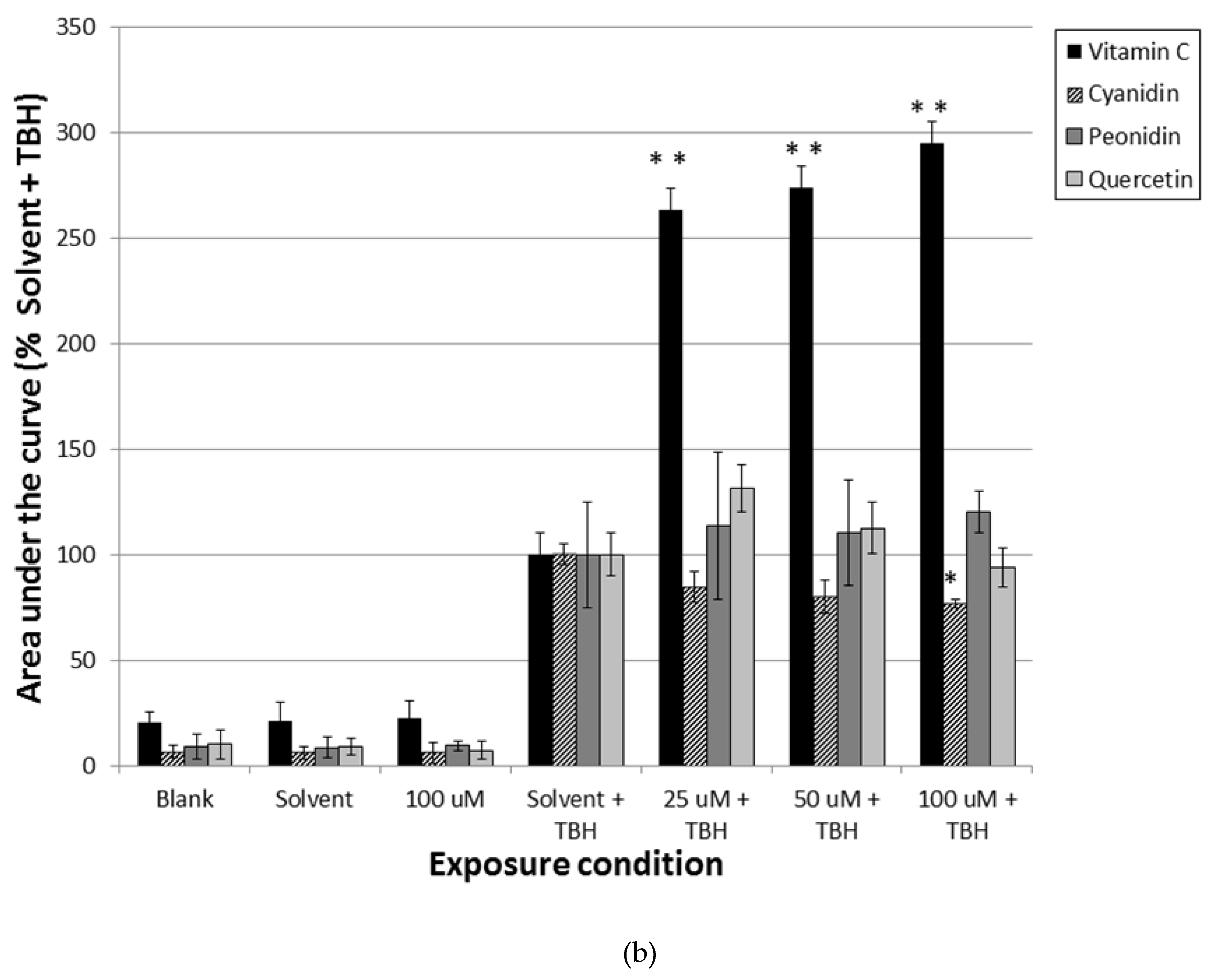
3.1. Pre-Incubation with Blueberry Extracts and Single Compounds Protect Caco-2 Cells against Oxidative DNA Damage
3.2. Pre-Incubation with Blueberry Extracts and Individual Phytochemicals Reduces Free Radical Generation in Caco-2 Cells
3.3. Modulation of Gene Expression by the Blueberry Extracts and Individual Phytochemicals
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Cancer Research Fund; American Institute for Cancer Research. Nutrition, Physical Activity, and the Prevention of Cancer: A Global Perspective; American Institute for Cancer Research: Washington, DC, USA, 2007; p. 517. [Google Scholar]
- De Kok, T.M.; van Breda, S.G.; Manson, M.M. Mechanisms of combined action of different chemopreventive dietary compounds: A review. Eur. J. Nutr. 2008, 47 (Suppl. 2), 51–59. [Google Scholar] [CrossRef] [PubMed]
- Kok, T.M.; Breda, S.G.; Briede, J.J. Genomics-based identification of molecular mechanisms behind the cancer preventive action of phytochemicals: Potential and challenges. Curr. Pharm. Biotechnol. 2012, 13, 255–264. [Google Scholar] [PubMed]
- Van Breda, S.G.J.; de Kok, T. Smart Combinations of Bioactive Compounds in Fruits and Vegetables May Guide New Strategies for Personalized Prevention of Chronic Diseases. Mol. Nutr. Food Res. 2017. [Google Scholar] [CrossRef] [PubMed]
- Van Breda, S.G.; van Agen, E.; Engels, L.G.; Moonen, E.J.; Kleinjans, J.C.; van Delft, J.H. Altered vegetable intake affects pivotal carcinogenesis pathways in colon mucosa from adenoma patients and controls. Carcinogenesis 2004, 25, 2207–2216. [Google Scholar] [CrossRef] [PubMed]
- Grainger, E.M.; Schwartz, S.J.; Wang, S.; Unlu, N.Z.; Boileau, T.W.; Ferketich, A.K.; Monk, J.P.; Gong, M.C.; Bahnson, R.R.; DeGroff, V.L.; et al. A combination of tomato and soy products for men with recurring prostate cancer and rising prostate specific antigen. Nutr. Cancer 2008, 60, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Thompson, H.J.; Heimendinger, J.; Diker, A.; O’Neill, C.; Haegele, A.; Meinecke, B.; Wolfe, P.; Sedlacek, S.; Zhu, Z.; Jiang, W. Dietary botanical diversity affects the reduction of oxidative biomarkers in women due to high vegetable and fruit intake. J. Nutr. 2006, 136, 2207–2212. [Google Scholar] [CrossRef] [PubMed]
- George, T.W.; Paterson, E.; Waroonphan, S.; Gordon, M.H.; Lovegrove, J.A. Effects of chronic consumption of fruit and vegetable puree-based drinks on vasodilation, plasma oxidative stability and antioxidant status. J. Hum. Nutr. Diet. 2012, 25, 477–487. [Google Scholar] [CrossRef]
- van Breda, S.G.; Wilms, L.C.; Gaj, S.; Jennen, D.G.; Briede, J.J.; Helsper, J.P.; Kleinjans, J.C.; de Kok, T.M. Can transcriptomics provide insight into the chemopreventive mechanisms of complex mixtures of phytochemicals in humans? Antioxid. Redox Signal. 2014, 20, 2107–2113. [Google Scholar] [CrossRef]
- Van Breda, S.G.; Wilms, L.C.; Gaj, S.; Jennen, D.G.; Briede, J.J.; Kleinjans, J.C.; de Kok, T.M. The exposome concept in a human nutrigenomics study: Evaluating the impact of exposure to a complex mixture of phytochemicals using transcriptomics signatures. Mutagenesis 2015, 30, 723–731. [Google Scholar] [CrossRef]
- Wilms, L.C.; Boots, A.W.; de Boer, V.C.; Maas, L.M.; Pachen, D.M.; Gottschalk, R.W.; Ketelslegers, H.B.; Godschalk, R.W.; Haenen, G.R.; van Schooten, F.J.; et al. Impact of multiple genetic polymorphisms on effects of a 4-week blueberry juice intervention on ex vivo induced lymphocytic DNA damage in human volunteers. Carcinogenesis 2007, 28, 1800–1806. [Google Scholar] [CrossRef]
- Hebels, D.G.; Jennen, D.G.; Kleinjans, J.C.; de Kok, T.M. Molecular signatures of N-nitroso compounds in Caco-2 cells: Implications for colon carcinogenesis. Toxicol. Sci. 2009, 108, 290–300. [Google Scholar] [CrossRef] [PubMed]
- Helsper, J.P. Antioxidantcapaciteit en Gehalten aan Anthocyanen en Fenolische Verbindingen in in Nederland Geteelde Rassen van Blauwe bes (Vaccinium corymbosum L.); Plant Research International: Wageningen, The Netherlands, 2008. [Google Scholar]
- Linskens, H.-F.; Jackson, J.F. (Eds.) High Performance Liquid Chromatography in Plant Sciences, 1st ed.; Springer: Berlin/Heidelberg, Germany, 1987; Volume 3. [Google Scholar]
- Singh, N.P.; McCoy, M.T.; Tice, R.R.; Schneider, E.L. A simple technique for quantitation of low levels of DNA damage in individual cells. Exp. Cell Res. 1988, 175, 184–191. [Google Scholar] [CrossRef]
- Tice, R.R.; Agurell, E.; Anderson, D.; Burlinson, B.; Hartmann, A.; Kobayashi, H.; Miyamae, Y.; Rojas, E.; Ryu, J.C.; Sasaki, Y.F. Single cell gel/comet assay: Guidelines for in vitro and in vivo genetic toxicology testing. Environ. Mol. Mutagen. 2000, 35, 206–221. [Google Scholar] [CrossRef]
- Tiku, M.L.; Yan, Y.P.; Chen, K.Y. Hydroxyl radical formation in chondrocytes and cartilage as detected by electron paramagnetic resonance spectroscopy using spin trapping reagents. Free Radic. Res. 1998, 29, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Hebels, D.G.; Briede, J.J.; Khampang, R.; Kleinjans, J.C.; de Kok, T.M. Radical mechanisms in nitrosamine- and nitrosamide-induced whole-genome gene expression modulations in Caco-2 cells. Toxicol. Sci. 2010, 116, 194–205. [Google Scholar] [CrossRef]
- Kimes, P.K.; Liu, Y.; Neil Hayes, D.; Marron, J.S. Statistical significance for hierarchical clustering. Biometrics 2017, 73, 811–821. [Google Scholar] [CrossRef] [PubMed]
- Noroozi, M.; Angerson, W.J.; Lean, M.E. Effects of flavonoids and vitamin C on oxidative DNA damage to human lymphocytes. Am. J. Clin. Nutr. 1998, 67, 1210–1218. [Google Scholar] [CrossRef]
- Amorini, A.M.; Fazzina, G.; Lazzarino, G.; Tavazzi, B.; Di Pierro, D.; Santucci, R.; Sinibaldi, F.; Galvano, F.; Galvano, G. Activity and mechanism of the antioxidant properties of cyanidin-3-O-β-glucopyranoside. Free Radic. Res. 2001, 35, 953–966. [Google Scholar] [CrossRef]
- Petruk, G.; Illiano, A.; Del Giudice, R.; Raiola, A.; Amoresano, A.; Rigano, M.M.; Piccoli, R.; Monti, D.M. Malvidin and cyanidin derivatives from acai fruit (Euterpe oleracea Mart.) counteract UV-A-induced oxidative stress in immortalized fibroblasts. J. Photochem. Photobiol. B Boil. 2017, 172, 42–51. [Google Scholar] [CrossRef]
- Briede, J.J.; De Kok, T.M.; Hogervorst, J.G.; Moonen, E.J.; Op Den Camp, C.L.; Kleinjanst, J.C. Development and application of an electron spin resonance spectrometry method for the determination of oxygen free radical formation by particulate matter. Environ. Sci. Technol. 2005, 39, 8420–8426. [Google Scholar] [CrossRef]
- Drozdz, P.; Seziene, V.; Pyrzynska, K. Phytochemical Properties and Antioxidant Activities of Extracts from Wild Blueberries and Lingonberries. Plant Foods Hum. Nutr. 2017, 72, 360–364. [Google Scholar] [CrossRef] [PubMed]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Blueberry–Apple Juice | Aurora | Bluecrop | Draper | Elliot | |
|---|---|---|---|---|---|
| Cyanidin-3-O-glucoside | 0.6 | 4.7 | 1.4 | 2.7 | 4.7 |
| Peonidin-3-O-glucoside | 1.7 | 4.3 | 1.5 | 2.7 | 2.7 |
| Quercetin-3-β-d-glucoside | 6.9 | 1.1 | 1.2 | 1.2 | 1.1 |
| Vitamin C | 0.6 | <0.5 * | <0.5 * | <0.5 * | <0.5 * |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van Breda, S.G.J.; Briedé, J.J.; De Kok, T.M.C.M. Improved Preventive Effects of Combined Bioactive Compounds Present in Different Blueberry Varieties as Compared to Single Phytochemicals. Nutrients 2019, 11, 61. https://doi.org/10.3390/nu11010061
Van Breda SGJ, Briedé JJ, De Kok TMCM. Improved Preventive Effects of Combined Bioactive Compounds Present in Different Blueberry Varieties as Compared to Single Phytochemicals. Nutrients. 2019; 11(1):61. https://doi.org/10.3390/nu11010061
Chicago/Turabian StyleVan Breda, Simone G. J., Jacob J. Briedé, and Theo M. C. M. De Kok. 2019. "Improved Preventive Effects of Combined Bioactive Compounds Present in Different Blueberry Varieties as Compared to Single Phytochemicals" Nutrients 11, no. 1: 61. https://doi.org/10.3390/nu11010061
APA StyleVan Breda, S. G. J., Briedé, J. J., & De Kok, T. M. C. M. (2019). Improved Preventive Effects of Combined Bioactive Compounds Present in Different Blueberry Varieties as Compared to Single Phytochemicals. Nutrients, 11(1), 61. https://doi.org/10.3390/nu11010061