Associations between Flavonoid Intakes and Gut Microbiota in a Group of Adults with Cystic Fibrosis


Abstract
1. Introduction
2. Materials and Methods
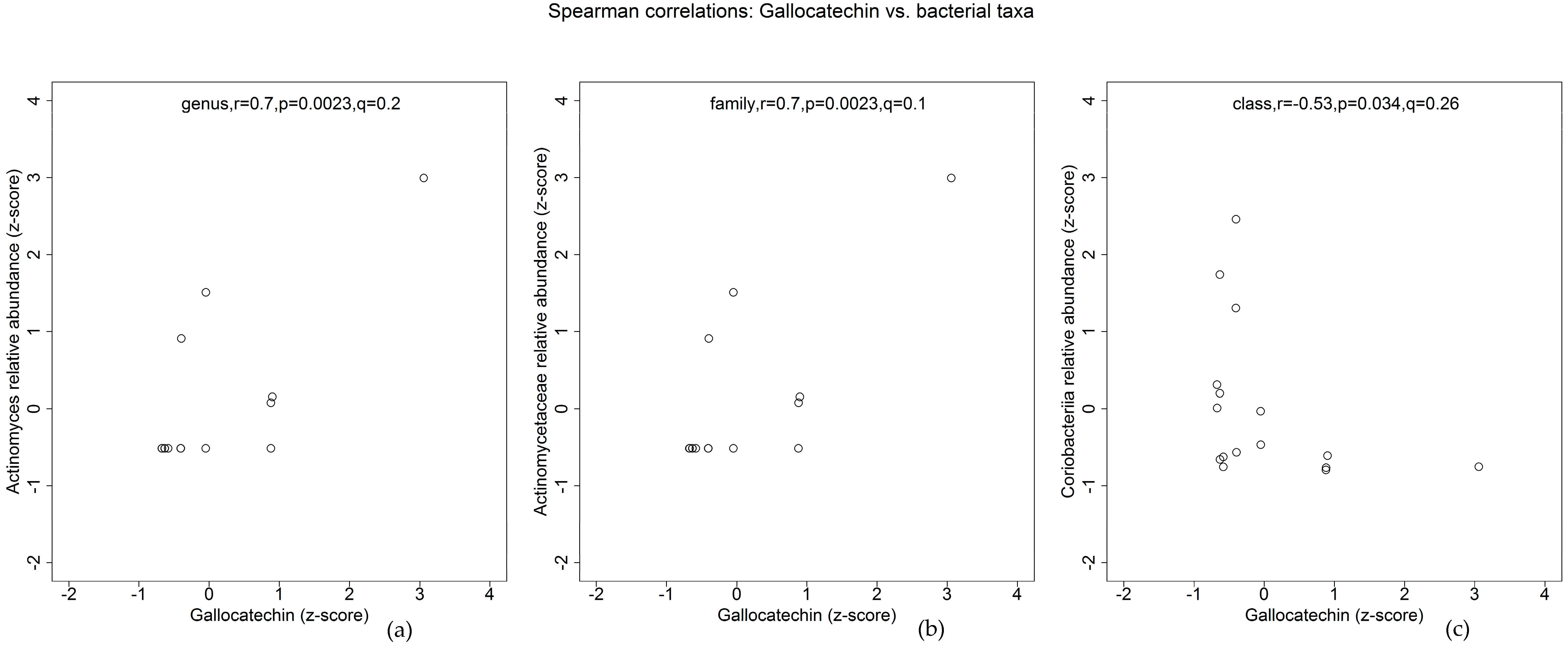
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Parkins, M.D.; Parkins, V.M.; Rendall, J.C.; Elborn, S. Changing epidemiology and clinical issues arising in an ageing cystic fibrosis population. Ther. Adv. Respir. Dis. 2011, 5, 105–119. [Google Scholar] [CrossRef] [PubMed]
- Moran, A.; Becker, D.; Casella, S.J.; Gottlieb, P.A.; Kirkman, M.S.; Marshall, B.C.; Slovis, B.; CFRD Consensus Conference Committee. Epidemiology, pathophysiology, and prognostic implications of cystic fibrosis–related diabetes. Diabetes Care 2010, 33, 2677–2683. [Google Scholar] [CrossRef] [PubMed]
- Gory, I.; Brown, G.; Wilson, J.; Kemp, W.; Paul, E.; Roberts, S.K. Increased risk of colorectal neoplasia in adult patients with cystic fibrosis: A matched case–control study. Scand. J. Gastroenterol. 2014, 49, 1230–1236. [Google Scholar] [CrossRef] [PubMed]
- Atarashi, K.; Tanoue, T.; Oshima, K.; Suda, W.; Nagano, Y.; Nishikawa, H.; Fukuda, S.; Saito, T.; Narushima, S.; Hase, K.; et al. Treg induction by a rationally selected mixture of clostridia strains from the human microbiota. Nature 2013, 500, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Rosser, E.C.; Oleinika, K.; Tonon, S.; Doyle, R.; Bosma, A.; Carter, N.A.; Harris, K.A.; Jones, S.A.; Klein, N.; Mauri, C. Regulatory b cells are induced by gut microbiota-driven interleukin-1[beta] and interleukin-6 production. Nat. Med. 2014, 20, 1334–1339. [Google Scholar] [CrossRef] [PubMed]
- Tolhurst, G.; Heffron, H.; Lam, Y.S.; Parker, H.E.; Habib, A.M.; Diakogiannaki, E.; Cameron, J.; Grosse, J.; Reimann, F.; Gribble, F.M. Short-chain fatty acids stimulate glucagon-like peptide-1 secretion via the g-protein–coupled receptor ffar2. Diabetes 2012, 61, 364–371. [Google Scholar] [CrossRef] [PubMed]
- Vinolo, M.A.R.; Rodrigues, H.G.; Nachbar, R.T.; Curi, R. Regulation of inflammation by short chain fatty acids. Nutrients 2011, 3, 858–876. [Google Scholar] [CrossRef] [PubMed]
- Smyth, A.R.; Bell, S.C.; Bojcin, S.; Bryon, M.; Duff, A.; Flume, P.; Kashirskaya, N.; Munck, A.; Ratjen, F.; Schwarzenberg, S.J.; et al. European cystic fibrosis society standards of care: Best practice guidelines. J. Cyst. Fibros. 2014, 13, S23–S42. [Google Scholar] [CrossRef] [PubMed]
- Louis, P.; Hold, G.L.; Flint, H.J. The gut microbiota, bacterial metabolites and colorectal cancer. Nat. Rev. Microbiol. 2014, 12, 661–672. [Google Scholar] [CrossRef] [PubMed]
- Bruzzese, E.; Callegari, M.L.; Raia, V.; Viscovo, S.; Scotto, R.; Ferrari, S.; Morelli, L.; Buccigrossi, V.; Lo Vecchio, A.; Ruberto, E.; et al. Disrupted intestinal microbiota and intestinal inflammation in children with cystic fibrosis and its restoration with lactobacillus GG: A randomised clinical trial. PLoS ONE 2014, 9, e87796. [Google Scholar] [CrossRef] [PubMed]
- Duytschaever, G.; Huys, G.; Bekaert, M.; Boulanger, L.; De Boeck, K.; Vandamme, P. Dysbiosis of bifidobacteria and clostridium cluster xiva in the cystic fibrosis fecal microbiota. J. Cyst. Fibros. 2013, 12, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Scanlan, P.D.; Buckling, A.; Kong, W.; Wild, Y.; Lynch, S.V.; Harrison, F. Gut dysbiosis in cystic fibrosis. J. Cyst. Fibros. 2012, 11, 454–455. [Google Scholar] [CrossRef] [PubMed]
- Hoen, A.G.; Li, J.; Moulton, L.A.; O’Toole, G.A.; Housman, M.L.; Koestler, D.C.; Guill, M.F.; Moore, J.H.; Hibberd, P.L.; Morrison, H.G.; et al. Associations between gut microbial colonization in early life and respiratory outcomes in cystic fibrosis. J. Pediatr. 2015, 167, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Jafari, S.-A.; Mehdizadeh-Hakkak, A.; Kianifar, H.-R.; Hebrani, P.; Ahanchian, H.; Abbasnejad, E. Effects of probiotics on quality of life in children with cystic fibrosis; a randomized controlled trial. Iran. J. Pediatr. 2013, 23, 669–674. [Google Scholar] [PubMed]
- Weiss, B.; Bujanover, Y.; Yahav, Y.; Vilozni, D.; Fireman, E.; Efrati, O. Probiotic supplementation affects pulmonary exacerbations in patients with cystic fibrosis: A pilot study. Pediatr. Pulmonol. 2010, 45, 536–540. [Google Scholar] [CrossRef] [PubMed]
- Del Campo, R.; Garriga, M.; Pérez-Aragón, A.; Guallarte, P.; Lamas, A.; Máiz, L.; Bayón, C.; Roy, G.; Cantón, R.; Zamora, J.; et al. Improvement of digestive health and reduction in proteobacterial populations in the gut microbiota of cystic fibrosis patients using a lactobacillus reuteri probiotic preparation: A double blind prospective study. J. Cyst. Fibros. 2014, 13, 716–722. [Google Scholar] [CrossRef] [PubMed]
- Van Biervliet, S.; Declercq, D.; Somerset, S. Clinical effects of probiotics in cystic fibrosis patients: A systematic review. Clin. Nutr. ESPEN 2017, 18, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.; Sinha, I.P.; Dwan, K.; Carlos, E.; Schechter, M.; Southern, K.W. Potentiators (specific therapies for class iii and iv mutations) for cystic fibrosis. Cochrane Database Syst. Rev. 2015. [Google Scholar] [CrossRef] [PubMed]
- Wainwright, C.E.; Elborn, J.S.; Ramsey, B.W.; Marigowda, G.; Huang, X.; Cipolli, M.; Colombo, C.; Davies, J.C.; De Boeck, K.; Flume, P.A.; et al. Lumacaftor–ivacaftor in patients with cystic fibrosis homozygous for phe508del cftr. N. Engl. J. Med. 2015, 373, 220–231. [Google Scholar] [CrossRef] [PubMed]
- Sawicki, G.S.; Sellers, D.E.; Robinson, W.M. High treatment burden in adults with cystic fibrosis: Challenges to disease self-management. J. Cyst. Fibros. 2009, 8, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Healey, G.R.; Murphy, R.; Brough, L.; Butts, C.A.; Coad, J. Interindividual variability in gut microbiota and host response to dietary interventions. Nutr. Rev. 2017, 75, 1059–1080. [Google Scholar] [CrossRef] [PubMed]
- Duda-Chodak, A. The inhibitory effect of polyphenols on human gut microbiota. J. Physiol. Pharmacol. 2012, 63, 497–503. [Google Scholar] [PubMed]
- Etxeberria, U.; Fernández-Quintela, A.; Milagro, F.I.; Aguirre, L.; Martínez, J.A.; Portillo, M.P. Impact of polyphenols and polyphenol-rich dietary sources on gut microbiota composition. J. Agric. Food Chem. 2013, 61, 9517–9533. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.-Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking long-term dietary patterns with gut microbial enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Cassidy, A.; Minihane, A.-M. The role of metabolism (and the microbiome) in defining the clinical efficacy of dietary flavonoids. Am. J. Clin. Nutr. 2017, 105, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.-F.; Xiao, Z.-T.; Zhang, H.-Y. Where do health benefits of flavonoids come from? Insights from flavonoid targets and their evolutionary history. Biochem. Biophys. Res. Commun. 2013, 434, 701–704. [Google Scholar] [CrossRef] [PubMed]
- Reverri, E.J.; Morrissey, B.M.; Cross, C.E.; Steinberg, F.M. Inflammation, oxidative stress, and cardiovascular disease risk factors in adults with cystic fibrosis. Free Radic. Biol. Med. 2014, 76, 261–277. [Google Scholar] [CrossRef] [PubMed]
- Richardson, I.; Nyulasi, I.; Cameron, K.; Ball, M.; Wilson, J. Nutritional status of an adult cystic fibrosis population. Nutrition 2000, 16, 255–259. [Google Scholar] [CrossRef]
- Olveira, G.; Dorado, A.; Olveira, C.; Padilla, A.; Rojo-Martínez, G.; García-Escobar, E.; Gaspar, I.; Gonzalo, M.; Soriguer, F. Serum phospholipid fatty acid profile and dietary intake in an adult mediterranean population with cystic fibrosis. Br. J. Nutr. 2006, 96, 343–349. [Google Scholar] [CrossRef] [PubMed]
- CFA. Cystic Fibrosis in Australia 2011: 14th Annual Report from the Australian Cystic Fibrosis Data Registry; Cystic Fibrosis Australia: Sydney, Australia, 2012; p. 42. ISSN 1447–3933. [Google Scholar]
- Li, L.; Krause, L.; Somerset, S. Associations between micronutrient intakes and gut microbiota in a group of adults with cystic fibrosis. Clin. Nutr. 2017, 36, 1097–1104. [Google Scholar] [CrossRef] [PubMed]
- Somerset, S.; Papier, K. A food frequency questionnaire validated for estimating dietary flavonoid intake in an australian population. Nutr. Cancer 2014, 66, 1200–1210. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. Qiime allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.A.; Hamady, M.; Kelley, S.T.; Knight, R. Quantitative and qualitative β diversity measures lead to different insights into factors that structure microbial communities. Appl. Environ. Microbiol. 2007, 73, 1576–1585. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package. R Package Version 2.0-10. 2013. Available online: http://cran.rproject.org/package=vegan (accessed on 9 December 2014).
- Pollard, K.S.; Dudoit, S.; van der Laan, M.J. Multiple testing procedures: R multtest package and applications to genomics. In Bioinformatics and Computational Biology Solutions Using r and Bioconductor; Gentleman, R., Carey, V., Huber, W., Irizarry, R., Dudoit, S., Eds.; Springer: Berlin, Germany, 2005; pp. 251–272. [Google Scholar]
- Lu, W.; Feng, Y.; Jing, F.; Han, Y.; Lyu, N.; Liu, F.; Li, J.; Song, X.; Xie, J.; Qiu, Z.; et al. Association between gut microbiota and cd4 recovery in HIV-1 infected patients. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Zakrzewski, M.; Proietti, C.; Ellis, J.J.; Hasan, S.; Brion, M.-J.; Berger, B.; Krause, L. Calypso: A user-friendly web-server for mining and visualizing microbiome–environment interactions. Bioinformatics 2017, 33, 782–783. [Google Scholar] [CrossRef] [PubMed]
- Maukonen, J.; Saarela, M. Human gut microbiota: Does diet matter? Proc. Nutr. Soc. 2015, 74, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Spor, A.; Koren, O.; Ley, R. Unravelling the effects of the environment and host genotype on the gut microbiome. Nat. Rev. Microbiol. 2011, 9, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Yatsunenko, T.; Rey, F.E.; Manary, M.J.; Trehan, I.; Dominguez-Bello, M.G.; Contreras, M.; Magris, M.; Hidalgo, G.; Baldassano, R.N.; Anokhin, A.P.; et al. Human gut microbiome viewed across age and geography. Nature 2012, 486, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Faith, J.J.; Guruge, J.L.; Charbonneau, M.; Subramanian, S.; Seedorf, H.; Goodman, A.L.; Clemente, J.C.; Knight, R.; Heath, A.C.; Leibel, R.L.; et al. The long-term stability of the human gut microbiota. Science 2013, 341. [Google Scholar] [CrossRef] [PubMed]
- Kutschera, M. Isolation of catechin-converting human intestinal bacteria catechin-converting bacteria. J. Appl. Microbiol. 2011, 111, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Takagaki, A.; Nanjo, F. Biotransformation of (-)-epigallocatechin and (-)-gallocatechin by intestinal bacteria involved in isoflavone metabolism. Biol. Pharm. Bull. 2015, 38, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Hoyles, L.; Clear, J.A.; McCartney, A.L. Use of denaturing gradient gel electrophoresis to detect actinobacteria associated with the human faecal microbiota. Anaerobe 2013, 22, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Claus, S.P.; Ellero, S.L.; Berger, B.; Krause, L.; Bruttin, A.; Molina, J.; Paris, A.; Want, E.J.; de Waziers, I.; Cloarec, O.; et al. Colonization-induced host-gut microbial metabolic interaction. mBio 2011, 2. [Google Scholar] [CrossRef] [PubMed]
- Martinez, I.; Lattimer, J.M.; Hubach, K.L.; Case, J.A.; Yang, J.; Weber, C.G.; Louk, J.A.; Rose, D.J.; Kyureghian, G.; Peterson, D.A.; et al. Gut microbiome composition is linked to whole grain-induced immunological improvements. ISME J. 2013, 7, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.W.; Ince, J.; Duncan, S.H.; Webster, L.M.; Holtrop, G.; Ze, X.; Brown, D.; Stares, M.D.; Scott, P.; Bergerat, A.; et al. Dominant and diet-responsive groups of bacteria within the human colonic microbiota. ISME J. 2011, 5, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Tunney, M.M.; Field, T.R.; Moriarty, T.F.; Patrick, S.; Doering, G.; Muhlebach, M.S.; Wolfgang, M.C.; Boucher, R.; Gilpin, D.F.; McDowell, A.; et al. Detection of anaerobic bacteria in high numbers in sputum from patients with cystic fibrosis. Am. J. Respir. Crit. Care Med. 2008, 177, 995–1001. [Google Scholar] [CrossRef] [PubMed]
- Madan, J.C.; Koestler, D.C.; Stanton, B.A.; Davidson, L.; Moulton, L.A.; Housman, M.L.; Moore, J.H.; Guill, M.F.; Morrison, H.G.; Sogin, M.L.; et al. Serial analysis of the gut and respiratory microbiome in cystic fibrosis in infancy: Interaction between intestinal and respiratory tracts and impact of nutritional exposures. mBio 2012, 3, e00251-12. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Liu, F.; Ling, Z.; Tong, X.; Xiang, C. Human intestinal lumen and mucosa-associated microbiota in patients with colorectal cancer. PLoS ONE 2012, 7, e39743. [Google Scholar] [CrossRef] [PubMed]
- Somerset, S.M.; Johannot, L. Dietary flavonoid sources in australian adults. Nutr. Cancer 2008, 60, 442–449. [Google Scholar] [CrossRef] [PubMed]
- Cuervo, A.; Hevia, A.; López, P.; Suárez, A.; Sánchez, B.; Margolles, A.; González, S. Association of polyphenols from oranges and apples with specific intestinal microorganisms in systemic lupus erythematosus patients. Nutrients 2015, 7, 1301–1317. [Google Scholar] [CrossRef] [PubMed]
- Cardona, F.; Andrés-Lacueva, C.; Tulipani, S.; Tinahones, F.J.; Queipo-Ortuño, M.I. Benefits of polyphenols on gut microbiota and implications in human health. J. Nutr. Biochem. 2013, 24, 1415–1422. [Google Scholar] [CrossRef] [PubMed]
- Kawabata, K.; Sugiyama, Y.; Sakano, T.; Ohigashi, H. Flavonols enhanced production of anti-inflammatory substance(s) by bifidobacterium adolescentis: Prebiotic actions of galangin, quercetin, and fisetin. Biofactors 2013, 39, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Everard, A.; Duparc, T. Gut microbiota, enteroendocrine functions and metabolism. Curr. Opin. Pharm. 2013, 13, 935–940. [Google Scholar] [CrossRef] [PubMed]
- Cohen, T.S.; Prince, A. Cystic fibrosis: A mucosal immunodeficiency syndrome. Nat. Med. 2012, 18, 509–519. [Google Scholar] [CrossRef] [PubMed]
- Porro, C.; Di Gioia, S.; Trotta, T.; Lepore, S.; Panaro, M.A.; Battaglino, A.; Ratclif, L.; Castellani, S.; Bufo, P.; Martinez, M.C.; et al. Pro-inflammatory effect of cystic fibrosis sputum microparticles in the murine lung. J. Cyst. Fibros. 2013, 12, 721–728. [Google Scholar] [CrossRef] [PubMed]
- Constantinescu, A.A.; Gleizes, C.; Alhosin, M.; Yala, E.; Zobairi, F.; Leclercq, A.; Stoian, G.; Mitrea, I.L.; Prévost, G.; Toti, F.; et al. Exocrine cell-derived microparticles in response to lipopolysaccharide promote endocrine dysfunction in cystic fibrosis. J. Cyst. Fibros. 2014, 13, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Eckburg, P.B.; Bik, E.M.; Bernstein, C.N.; Purdom, E.; Dethlefsen, L.; Sargent, M.; Gill, S.R.; Nelson, K.E.; Relman, D.A. Diversity of the human intestinal microbial flora. Science 2005, 308, 1635–1638. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, S.; Dillon, J.F. Microbial biofilms in the human gastrointestinal tract. J. Appl. Microbiol. 2007, 102, 1187–1196. [Google Scholar] [CrossRef] [PubMed]
- Marteau, P.; Pochart, P.; Doré, J.; Béra-Maillet, C.; Bernalier, A.; Corthier, G. Comparative study of bacterial groups within the human cecal and fecal microbiota. Appl. Environ. Microbiol. 2001, 67, 4939–4942. [Google Scholar] [CrossRef] [PubMed]
- Momozawa, Y.; Deffontaine, V.; Louis, E.; Medrano, J.F. Characterization of bacteria in biopsies of colon and stools by high throughput sequencing of the v2 region of bacterial 16s rrna gene in human. PLoS ONE 2011, 6, e16952. [Google Scholar] [CrossRef] [PubMed]
- Zoetendal, E.G.; von Wright, A.; Vilpponen-Salmela, T.; Ben-Amor, K.; Akkermans, A.D.L.; de Vos, W.M. Mucosa-associated bacteria in the human gastrointestinal tract are uniformly distributed along the colon and differ from the community recovered from feces. Appl. Environ. Microbiol. 2002, 68, 3401–3407. [Google Scholar] [CrossRef] [PubMed]
- Guarner, F.; Malagelada, J.-R. Gut flora in health and disease. Lancet 2003, 361, 512–519. [Google Scholar] [CrossRef]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.-M.; et al. Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Koren, O.; Knights, D.; Gonzalez, A.; Waldron, L.; Segata, N.; Knight, R.; Huttenhower, C.; Ley, R.E. A guide to enterotypes across the human body: Meta-analysis of microbial community structures in human microbiome datasets. PLoS Comput. Biol. 2013, 9, e1002863. [Google Scholar] [CrossRef] [PubMed]
- Lam, Y.Y.; Zhang, C.; Zhao, L. Causality in dietary interventions—Building a case for gut microbiota. Genome Med. 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, M.; Martins dos Santos, V.A.P.; Ott, S.J.; Moya, A. Gut microbiota disturbance during antibiotic therapy. Gut Microbes 2013, 5, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Matamouros, S.; Hayden, H.S.; Hager, K.R.; Brittnacher, M.J.; Lachance, K.; Weiss, E.J.; Pope, C.E.; Imhaus, A.-F.; McNally, C.P.; Borenstein, E.; et al. Adaptation of commensal proliferating escherichia coli to the intestinal tract of young children with cystic fibrosis. Proc. Natl. Acad. Sci. USA 2018, 115, 1605–1610. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Variable | p Value 2 | R2 2 | q Value 3 |
|---|---|---|---|
| Gallocatechin | 0.047 | 0.078 | 0.285 |
| Theaflavin digallate | 0.051 | 0.078 | 0.285 |
| Thearubigins | 0.051 | 0.078 | 0.285 |
| Theaflavin | 0.052 | 0.078 | 0.285 |
| Theaflavin-3′-gallate | 0.052 | 0.078 | 0.285 |
| Theaflavin-3-gallate | 0.055 | 0.078 | 0.285 |
| % Anthocyanidins (excluding tea) | 0.056 | 0.077 | 0.285 |
| Petunidin | 0.087 | 0.076 | 0.387 |
| Peonidin | 0.101 | 0.075 | 0.387 |
| Malvidin | 0.108 | 0.076 | 0.387 |
| Eriodictyol | 0.157 | 0.075 | 0.514 |
| Wine flavonoids | 0.186 | 0.074 | 0.514 |
| Cocoa flavonoids | 0.186 | 0.075 | 0.514 |
| Tea flavonoids | 0.265 | 0.070 | 0.654 |
| Kaempferol | 0.273 | 0.070 | 0.654 |
| Delphinidin | 0.314 | 0.069 | 0.685 |
| Total flavonoids | 0.326 | 0.069 | 0.685 |
| Apigenin | 0.343 | 0.069 | 0.685 |
| Epicatechin | 0.376 | 0.068 | 0.712 |
| % Flavones (excluding tea) | 0.411 | 0.068 | 0.740 |
| % Flavan-3-ols (excluding tea) | 0.434 | 0.067 | 0.744 |
| Total flavonoids (excluding tea) | 0.554 | 0.065 | 0.905 |
| Cyanidin | 0.579 | 0.064 | 0.905 |
| Catechin | 0.679 | 0.063 | 0.967 |
| Ouercetin | 0.696 | 0.063 | 0.967 |
| Pelargonidin | 0.702 | 0.062 | 0.967 |
| Naringenin | 0.726 | 0.063 | 0.967 |
| Isorhamnetin | 0.787 | 0.060 | 0.977 |
| Myricetin | 0.791 | 0.061 | 0.977 |
| Luteolin | 0.815 | 0.061 | 0.977 |
| Epicatechin-3-gallate | 0.907 | 0.058 | 0.994 |
| Epigallocatecin | 0.913 | 0.058 | 0.994 |
| Hesperetin | 0.920 | 0.058 | 0.994 |
| % Flavonols (excluding tea) | 0.944 | 0.058 | 0.994 |
| % Flavanones (excluding tea) | 0.986 | 0.055 | 0.994 |
| Epigallocatechin-3-gallate | 0.994 | 0.053 | 0.994 |
| Variable | p Value 2 | R2 2 | q Value 3 |
|---|---|---|---|
| % Flavones (excluding tea) | 0.013 | 0.184 | 0.227 |
| % Flavonols (excluding tea) | 0.016 | 0.151 | 0.227 |
| Apigenin | 0.028 | 0.163 | 0.227 |
| Kaempferol | 0.029 | 0.145 | 0.227 |
| Gallocatechin | 0.053 | 0.131 | 0.227 |
| Theaflavin-3′-gallate | 0.053 | 0.130 | 0.227 |
| Theaflavin-3-gallate | 0.056 | 0.130 | 0.227 |
| Thearubigins | 0.057 | 0.130 | 0.227 |
| Theaflavin digallate | 0.059 | 0.130 | 0.227 |
| Theaflavin | 0.063 | 0.130 | 0.227 |
| Tea flavonoids | 0.087 | 0.113 | 0.285 |
| Total flavonoids | 0.098 | 0.110 | 0.291 |
| Cocoa flavonoids | 0.107 | 0.107 | 0.291 |
| Hesperetin | 0.113 | 0.105 | 0.291 |
| Eriodictyol | 0.127 | 0.103 | 0.305 |
| Pelargonidin | 0.149 | 0.103 | 0.318 |
| Cyanidin | 0.150 | 0.099 | 0.318 |
| Myricetin | 0.165 | 0.096 | 0.324 |
| % Anthocyanidins (excluding tea) | 0.171 | 0.095 | 0.324 |
| Delphinidin | 0.224 | 0.085 | 0.402 |
| % Flavanones (excluding tea) | 0.313 | 0.075 | 0.537 |
| Epigallocatecin | 0.341 | 0.071 | 0.542 |
| Epicatechin-3-gallate | 0.346 | 0.072 | 0.542 |
| Catechin | 0.445 | 0.065 | 0.652 |
| % Flavan-3-ols (excluding tea) | 0.453 | 0.064 | 0.652 |
| Epicatechin | 0.473 | 0.063 | 0.654 |
| Peonidin | 0.495 | 0.060 | 0.659 |
| Petunidin | 0.533 | 0.055 | 0.667 |
| Total flavonoids (excluding tea) | 0.544 | 0.059 | 0.667 |
| Malvidin | 0.559 | 0.053 | 0.667 |
| Ouercetin | 0.574 | 0.054 | 0.667 |
| Wine flavonoids | 0.617 | 0.048 | 0.692 |
| Naringenin | 0.634 | 0.050 | 0.692 |
| Epigallocatechin-3-gallate | 0.704 | 0.044 | 0.745 |
| Isorhamnetin | 0.811 | 0.036 | 0.834 |
| Luteolin | 0.852 | 0.037 | 0.852 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.; Somerset, S. Associations between Flavonoid Intakes and Gut Microbiota in a Group of Adults with Cystic Fibrosis. Nutrients 2018, 10, 1264. https://doi.org/10.3390/nu10091264
Li L, Somerset S. Associations between Flavonoid Intakes and Gut Microbiota in a Group of Adults with Cystic Fibrosis. Nutrients. 2018; 10(9):1264. https://doi.org/10.3390/nu10091264
Chicago/Turabian StyleLi, Li, and Shawn Somerset. 2018. "Associations between Flavonoid Intakes and Gut Microbiota in a Group of Adults with Cystic Fibrosis" Nutrients 10, no. 9: 1264. https://doi.org/10.3390/nu10091264
APA StyleLi, L., & Somerset, S. (2018). Associations between Flavonoid Intakes and Gut Microbiota in a Group of Adults with Cystic Fibrosis. Nutrients, 10(9), 1264. https://doi.org/10.3390/nu10091264