Longitudinal Variation of Amino Acid Levels in Human Milk and Their Associations with Infant Gender

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants and Study Design
2.2. Sample Collection
2.3. Amino Acid Analyses
2.4. Statistical Analysis
2.5. Ethics
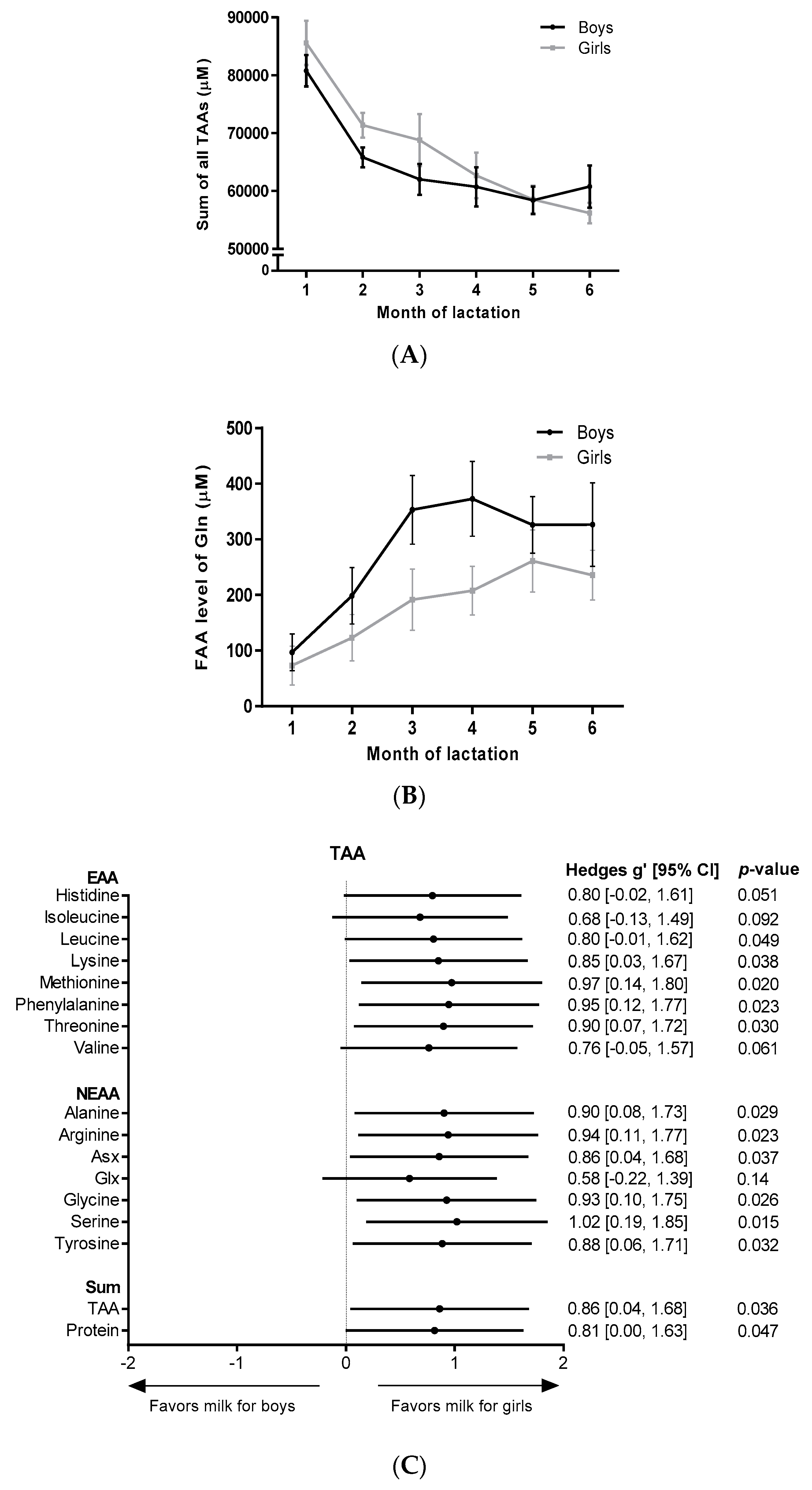
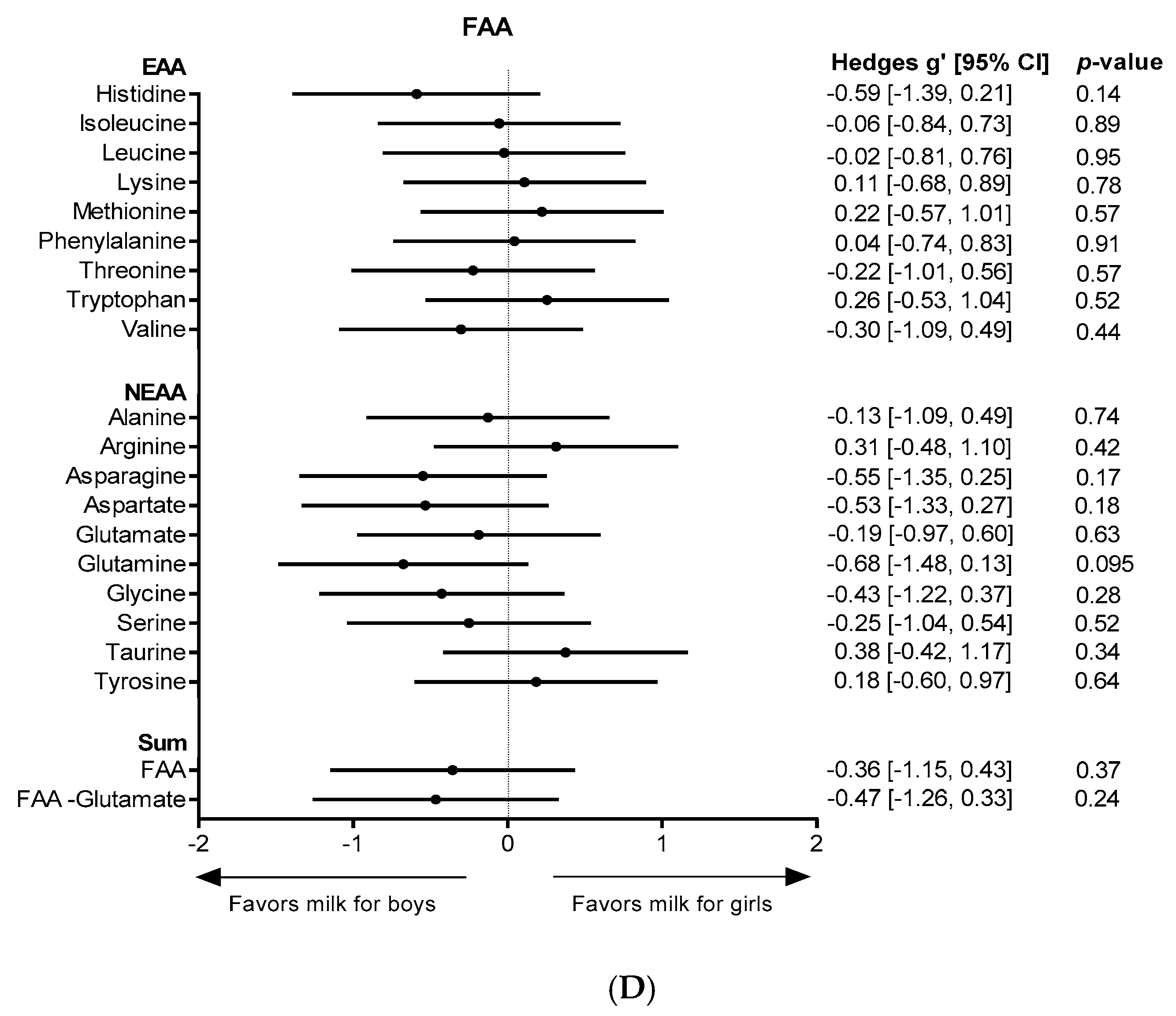
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jakaitis, B.M.; Denning, P.W. Human breast milk and the gastrointestinal innate immune system. Clin. Perinatol. 2014, 41, 423–435. [Google Scholar] [CrossRef] [PubMed]
- Yellis, M.B. Human breast milk and facilitation of gastrointestinal development and maturation. Gastroenterol. Nurs. 1995, 18, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Horta, B.L.; Loret de Mola, C.; Victora, C.G. Long-term consequences of breastfeeding on cholesterol, obesity, systolic blood pressure and type 2 diabetes: A systematic review and meta-analysis. Acta Paediatr. 2015, 104, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Lodge, C.J.; Tan, D.J.; Lau, M.X.; Dai, X.; Tham, R.; Lowe, A.J.; Bowatte, G.; Allen, K.J.; Dharmage, S.C. Breastfeeding and asthma and allergies: A systematic review and meta-analysis. Acta. Paediatr. 2015, 104, 38–53. [Google Scholar] [CrossRef] [PubMed]
- Meijer, C.R.; Discepolo, V.; Troncone, R.; Mearin, M.L. Does infant feeding modulate the manifestation of celiac disease and type 1 diabetes? Curr. Opin. Clin. Nutr. Metab. Care 2017, 20, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Sarwar, G.; Botting, H.G.; Davis, T.A.; Darling, P.; Pencharz, P.B. Free amino acids in milks of human subjects, other primates and non-primates. Br. J. Nutr. 1998, 79, 129–131. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Adelman, A.S.; Rai, D.; Boettcher, J.; Lonnerdal, B. Amino acid profiles in term and preterm human milk through lactation: A systematic review. Nutrients 2013, 5, 4800–4821. [Google Scholar] [CrossRef] [PubMed]
- Agostoni, C.; Carratu, B.; Boniglia, C.; Lammardo, A.M.; Riva, E.; Sanzini, E. Free glutamine and glutamic acid increase in human milk through a three-month lactation period. J. Pediatr. Gastroenterol. Nutr. 2000, 31, 508–512. [Google Scholar] [CrossRef] [PubMed]
- San Gabriel, A.; Uneyama, H. Amino acid sensing in the gastrointestinal tract. Amino Acids 2013, 45, 451–461. [Google Scholar] [CrossRef] [PubMed]
- Burrin, D.G.; Janeczko, M.J.; Stoll, B. Emerging aspects of dietary glutamate metabolism in the developing gut. Asia. Pac. J. Clin. Nutr. 2008, 17, 368–371. [Google Scholar] [PubMed]
- Munblit, D.; Peroni, D.G.; Boix-Amoros, A.; Hsu, P.S.; Van’t Land, B.; Gay, M.C.L.; Kolotilina, A.; Skevaki, C.; Boyle, R.J.; Collado, M.C.; et al. Human milk and allergic diseases: An unsolved puzzle. Nutrients 2017, 9, 894. [Google Scholar] [CrossRef] [PubMed]
- Almqvist, C.; Worm, M.; Leynaert, B. Impact of gender on asthma in childhood and adolescence: A ga2len review. Allergy 2008, 63, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Kelly, C.; Gangur, V. Sex disparity in food allergy: Evidence from the pubmed database. J.Allergy 2009, 2009, 159845. [Google Scholar] [CrossRef] [PubMed]
- Mandhane, P.J.; Greene, J.M.; Cowan, J.O.; Taylor, D.R.; Sears, M.R. Sex differences in factors associated with childhood- and adolescent-onset wheeze. Am. J. Respir. Crit. Care Med. 2005, 172, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Vriezinga, S.L.; Auricchio, R.; Bravi, E.; Castillejo, G.; Chmielewska, A.; Crespo Escobar, P.; Kolacek, S.; Koletzko, S.; Korponay-Szabo, I.R.; Mummert, E.; et al. Randomized feeding intervention in infants at high risk for celiac disease. New Engl. J. Med. 2014, 371, 1304–1315. [Google Scholar] [CrossRef] [PubMed]
- Thakkar, S.K.; Giuffrida, F.; Cristina, C.H.; De Castro, C.A.; Mukherjee, R.; Tran, L.A.; Steenhout, P.; Lee, L.Y.; Destaillats, F. Dynamics of human milk nutrient composition of women from singapore with a special focus on lipids. Am. J. Hum. Biol. 2013, 25, 770–779. [Google Scholar] [CrossRef] [PubMed]
- Powe, C.E.; Knott, C.D.; Conklin-Brittain, N. Infant sex predicts breast milk energy content. Am. J. Hum. Biol. 2010, 22, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Terrlink, T.; van Leeuwen, P.A.; Houdijk, A. Plasma amino acids determined by liquid chromatography within 17 minutes. Clin. Chem. 1994, 40, 245–249. [Google Scholar] [PubMed]
- Mariotti, F.; Tome, D.; Mirand, P.P. Converting nitrogen into protein—Beyond 6.25 and jones’ factors. Crit. Rev. Food Sci. Nutr. 2008, 48, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Wiles, P.G.; Gray, I.K.; Kissling, R.C. Routine analysis of proteins by kjeldahl and dumas methods: Review and interlaboratory study using dairy products. J. AOAC Int. 1998, 81, 620–632. [Google Scholar] [PubMed]
- Van de Heijning, B.J.M.; Schaart, M.W.; van der Beek, E.M.; Rings, E.H.H.M.; Luisa Mearin, M.L. Fatty acid and amino acid content and composition of human milk in the course of lactation. Adv. Pediatr. Res. 2017. [Google Scholar] [CrossRef]
- Da Costa, T.H.; Haisma, H.; Wells, J.C.; Mander, A.P.; Whitehead, R.G.; Bluck, L.J. How much human milk do infants consume? Data from 12 countries using a standardized stable isotope methodology. J. Nutr. 2010, 140, 2227–2232. [Google Scholar] [CrossRef] [PubMed]
- Dupont, C. Protein requirements during the first year of life. Am. J. Clin. Nutr. 2003, 77, 1544S–1549S. [Google Scholar] [CrossRef] [PubMed]
- Carratù, B.; Boniglia, C.; Scalise, F.; Ambruzzi, A.M.; Sanzini, E. Nitrogenous components of human milk: Non-protein nitrogen, true protein and free amino acids. Food Chem. 2003, 81, 357–362. [Google Scholar] [CrossRef]
- Garcia-Rodenas, C.L.; Affolter, M.; Vinyes-Pares, G.; De Castro, C.A.; Karagounis, L.G.; Zhang, Y.; Wang, P.; Thakkar, S.K. Amino acid composition of breast milk from urban chinese mothers. Nutrients 2016, 8, 606. [Google Scholar] [CrossRef] [PubMed]
- Pamblanco, M.; Portoles, M.; Paredes, C.; Ten, A.; Comin, J. Free amino acids in preterm and term milk from mothers delivering appropriate- or small-for-gestational-age infants. Am. J. Clin. Nutr. 1989, 50, 778–781. [Google Scholar] [CrossRef] [PubMed]
- Schanler, R.J.; Garza, C. Plasma amino acid differences in very low birth weight infants fed either human milk or whey-dominant cow milk formula. Pediatr. Res. 1987, 21, 301–305. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Knabe, D.A.; Kim, S.W.; Lynch, C.J.; Hutson, S.M.; Wu, G. Lactating porcine mammary tissue catabolizes branched-chain amino acids for glutamine and aspartate synthesis. J. Nutr. 2009, 139, 1502–1509. [Google Scholar] [CrossRef] [PubMed]
- Ruth, M.R.; Field, C.J. The immune modifying effects of amino acids on gut-associated lymphoid tissue. J. Anim. Sci. Biotechnol. 2013, 4, 27. [Google Scholar] [CrossRef] [PubMed]
- Sevastiadou, S.; Malamitsi-Puchner, A.; Costalos, C.; Skouroliakou, M.; Briana, D.D.; Antsaklis, A.; Roma-Giannikou, E. The impact of oral glutamine supplementation on the intestinal permeability and incidence of necrotizing enterocolitis/septicemia in premature neonates. J. Matern. Fetal Neonatal. Med. 2011, 24, 1294–1300. [Google Scholar] [CrossRef] [PubMed]
- Blachier, F.; Boutry, C.; Bos, C.; Tome, D. Metabolism and functions of l-glutamate in the epithelial cells of the small and large intestines. Am. J. Clin. Nutr. 2009, 90, 814S–821S. [Google Scholar] [CrossRef] [PubMed]
- Jiao, N.; Wu, Z.; Ji, Y.; Wang, B.; Dai, Z.; Wu, G. L-glutamate enhances barrier and antioxidative functions in intestinal porcine epithelial cells. J. Nutr. 2015, 145, 2258–2264. [Google Scholar] [CrossRef] [PubMed]
- Rao, R.; Samak, G. Role of glutamine in protection of intestinal epithelial tight junctions. J. Epithel. Biol. Pharmacol. 2012, 5, 47–54. [Google Scholar] [PubMed]
- Rook, D.; Te Braake, F.W.; Schierbeek, H.; Longini, M.; Buonocore, G.; Van Goudoever, J.B. Glutathione synthesis rates in early postnatal life. Pediatr. Res. 2010, 67, 407–411. [Google Scholar] [CrossRef] [PubMed]
- Newsholme, P. Why is l-glutamine metabolism important to cells of the immune system in health, postinjury, surgery or infection? J. Nutr. 2001, 131, 2515S–2522S. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, R.; Oliva, H.; Martinez-Navio, J.M.; Climent, N.; Ciruela, F.; Gatell, J.M.; Gallart, T.; Mallol, J.; Lluis, C.; Franco, R. Glutamate released by dendritic cells as a novel modulator of t cell activation. J. Immunol. 2006, 177, 6695–6704. [Google Scholar] [CrossRef] [PubMed]
- Lechowski, S.; Feilhauer, K.; Staib, L.; Coeffier, M.; Bischoff, S.C.; Lorentz, A. Combined arginine and glutamine decrease release of de novo synthesized leukotrienes and expression of proinflammatory cytokines in activated human intestinal mast cells. Eur. J. Nutr. 2013, 52, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Malek, T.R.; Bayer, A.L. Tolerance, not immunity, crucially depends on IL-2. Nat. Rev. Immunol. 2004, 4, 665–674. [Google Scholar] [CrossRef] [PubMed]
- Sikalidis, A.K. Amino acids and immune response: A role for cysteine, glutamine, phenylalanine, tryptophan and arginine in T-cell function and cancer? Pathol. Oncol. Res. 2015, 21, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, R.; Ciruela, F.; Casado, V.; Mallol, J.; Gallart, T.; Lluis, C.; Franco, R. Group I metabotropic glutamate receptors mediate a dual role of glutamate in T cell activation. J. Biol. Chem. 2004, 279, 33352–33358. [Google Scholar] [CrossRef] [PubMed]
- Thakkar, S.K.; Giuffrida, F.; Bertschy, E.; De Castro, A.; Destaillats, F.; Lee le, Y. Protein evolution of human milk. Nestle Nutr. Inst. Workshop Ser. 2016, 86, 77–85. [Google Scholar] [PubMed]
- Larnkjaer, A.; Bruun, S.; Pedersen, D.; Zachariassen, G.; Barkholt, V.; Agostoni, C.; Mlgaard, C.; Husby, S.; Michaelsen, K.F. Free amino acids in human milk and associations with maternal anthropometry and infant growth. J. Pediatr. Gastroenterol. Nutr. 2016, 63, 374–378. [Google Scholar] [CrossRef] [PubMed]
- Hill, I.D.; Dirks, M.H.; Liptak, G.S.; Colletti, R.B.; Fasano, A.; Guandalini, S.; Hoffenberg, E.J.; Horvath, K.; Murray, J.A.; Pivor, M.; et al. Guideline for the diagnosis and treatment of celiac disease in children: Recommendations of the north american society for pediatric gastroenterology, hepatology and nutrition. J. Pediatr. Gastroenterol. Nutr. 2005, 40, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, C.L.; Cubero, J.; Sánchez, J.; Franco, L.; Rodríguez, A.B.; Rivero, M.; Barriga, C. Evolution of the circadian profile of human milk amino acids during breastfeeding. J. Appl. Biomed. 2013, 11, 59–70. [Google Scholar] [CrossRef]
- Feng, P.; Gao, M.; Burgher, A.; Zhou, T.H.; Pramuk, K. A nine-country study of the protein content and amino acid composition of mature human milk. Food Nutr. Res. 2016, 60, 31042. [Google Scholar] [CrossRef] [PubMed]
- Lonnerdal, B. Effects of maternal dietary intake on human milk composition. J. Nutr. 1986, 116, 499–513. [Google Scholar] [CrossRef] [PubMed]
- Ding, M.; Li, W.; Zhang, Y.; Wang, X.; Zhao, A.; Zhao, X.; Wang, P.; Sheng, Q.H. Amino acid composition of lactating mothers’ milk and confinement diet in rural north china. Asia Pac. J. Clin. Nutr. 2010, 19, 344–349. [Google Scholar] [PubMed]



{kind=link}
{kind=link}
| Male (n = 12) | Female (n = 13) | Statistical Analysis of the Difference | |
|---|---|---|---|
| Weight (g) | |||
| Birth | 3511.3 ± 144.3 (12) | 3655.0 ± 104.1 (13) | 0.4223 |
| Δ 0–2 months | 1996.7 ± 202.4 (12) | 1406.5 ± 122.6 (10) | * 0.0279 |
| Δ 2–4 months | 1375.5 ± 187.5 (10) | 1107.8 ± 78.8 (9) | 0.2232 |
| Δ 4–6 months | 1033.6 ± 140.9 (7) | 798.4 ± 97.6 (9) | 0.1785 |
| Δ 0–6 months | 4156.7 ± 279.7 (9) | 3392.8 ± 273.1 (9) | o 0.0684 |
| Length (cm) | |||
| Birth | 50.0 ± 0.7 (4) | 50.8 ± 0.4 (9) | 0.3327 |
| Δ 0–2 months | 6.6 ± 1.0 (4) | 7.7 ± 0.6 (6) | 0.3876 |
| Δ 2–4 months | 5.9 ± 0.5 (9) | 4.4 ± 0.6 (8) | o 0.0556 |
| Δ 4–6 months | 4.0 ± 0.6 (6) | 3.6 ± 0.5 (8) | 0.6312 |
| Δ 0–6 months | 17.2 ± 1.8 (4) | 14.2 ± 0.9 (6) | 0.1335 |
| FAA (µmol/L) | Month 1 (n = 23) | Month 2 (n = 25) | Month 3 (n = 24) | Month 4 (n = 18) | Month 5 (n = 23) | Month 6 (n = 22) | Statistical Analysis of the Changes |
|---|---|---|---|---|---|---|---|
| EAA | |||||||
| Histidine | 23.3 ± 1.4 | 21.8 ± 1.2 | 20.9 ± 1.1 | 19.2 ± 1.2 | 17.7 ± 0.7 | 18.5 ± 1.1 | |
| Isoleucine | 12.5 ± 2.2 | 12.4 ± 1.4 | 10.4 ± 1.1 | 12.3 ± 1.9 | 10.4 ± 0.8 | 14.4 ± 2.7 | |
| Leucine | 35.4 ± 5.1 | 36.0 ± 3.4 | 31.8 ± 2.2 | 40.8 ± 7.0 | 31.5 ± 2.3 | 40.1 ± 7.3 | |
| Lysine | 31.2 ± 4.1 | 28.5 ± 4.9 | 28.9 ± 3.1 | 28.7 ± 7.6 | 24.5 ± 2.0 | 26.9 ± 3.3 | |
| Methionine | 2.3 ± 0.5 | 3.0 ± 0.4 | 2.6 ± 0.2 | 3.2 ± 0.5 | 3.4 ± 0.3 | 3.9 ± 0.7 | |
| Phenylalanine | 18.8 ± 2.5 | 19.3 ± 1.6 | 17.4 ± 1.9 | 18.2 ± 1.8 | 15.5 ± 0.8 | 21.0 ± 3.9 | |
| Threonine | 66.6 ± 4.7 | 80.2 ± 6.0 | 75.4 ± 3.9 | 81.7 ± 6.4 | 85.1 ± 4.4 | 80.5 ± 5.2 | a *, c * |
| Tryptophan | 23.8 ± 1.3 | 23.3 ± 1.4 | 21.8 ± 1.5 | 22.3 ± 1.8 | 20.8 ± 1.2 | 24.1 ± 2.6 | |
| Valine | 54.2 ± 4.6 | 57.9 ± 3.5 | 53.3 ± 3.3 | 56.3 ± 3.9 | 45.6 ± 2.4 | 49.6 ± 3.7 | |
| NEAA | |||||||
| Alanine | 251.5 ± 22.0 | 236.2 ± 13.6 | 241.7 ± 13.2 | 256.2 ± 15.8 | 245.3 ± 11.9 | 227.7 ± 16.4 | |
| Arginine | 16.4 ± 1.6 | 15.9 ± 2.5 | 13.4 ± 1.2 | 20.6 ± 5.9 | 15.3 ± 1.8 | 16.8 ± 2.0 | |
| Asparagine | 9.1 ± 1.9 | 9.5 ± 1.9 | 11.4 ± 2.4 | 12.9 ± 3.4 | 13.2 ± 2.3 | 10.1 ± 2.2 | |
| Aspartate | 39.8 ± 3.8 | 50.4 ± 4.0 | 61.8 ± 7.4 | 63.6 ± 5.9 | 74.4 ± 7.7 | 67.0± 7.0 | a **, c ** |
| Glutamate | 1267.4 ± 84.3 | 1634.3 ± 62.1 | 1713.4 ± 54.6 | 1825.4 ± 66.1 | 1776.5 ± 60.4 | 1769.2 ± 70.9 | a **, c ** |
| Glutamine | 84.1 ± 23.9 | 159.3 ± 32.9 | 265.6 ± 43.6 | 290.1 ± 43.8 | 293.4 ± 37.6 | 281.0 ± 43.3 | a **, c ** |
| Glycine | 82.7 ± 7.9 | 94.7 ± 6.6 | 102.5 ± 6.4 | 104.1 ± 5.4 | 105.2 ± 5.2 | 111.5 ± 7.3 | a **, c ** |
| Serine | 79.6 ± 6.7 | 97.4 ± 6.1 | 107.0 ± 7.1 | 107.3 ± 5.8 | 104.76 ± 6.4 | 100.0 ± 6.2 | a *, c ** |
| Taurine | 244.1 ± 14.2 | 240.1 ± 16.6 | 250.0 ± 20.7 | 232.1 ± 19.6 | 218.0 ± 12.8 | 245.9 ± 26.9 | |
| Tyrosine | 16.8 ± 1.9 | 15.8 ± 1.5 | 13.8 ± 1.0 | 14.7 ± 1.7 | 14.0 ± 1.2 | 17.3 ± 1.8 | |
| SUM | |||||||
| EAA | 284.7 ± 21.5 | 298.2 ± 20.0 | 268.9 ± 12.2 | 304.1 ± 29.8 | 269.7 ±11.3 | 295.7 ± 25.7 | |
| NEAA | 2075.0 ±129.1 | 2537.9±102.6 | 2767.2± 94.1 | 2906.3 ±107.4 | 2844.8 ±81.7 | 2829.8 ±118.6 | a **, c ** |
| FAA | 2359.7 ±140.2 | 2836.1±111.5 | 3036.1 ±100.9 | 3210.4 ±117.8 | 3114.5 ± 88.1 | 3125.5 ±133.6 | a **, c ** |
| FAA without Glutamate | 1092.3 ± 64.9 | 1201.8 ± 66.7 | 1322.7 ± 60.5 | 1385.1 ± 70.2 | 1338.0 ± 54.8 | 1356.2 ± 81.6 | a **, c ** |
| TAA (µmol/L) | Month 1 (n = 23) | Month 2 (n = 25) | Month 3 (n = 24) | Month 4 (n = 18) | Month 5 (n = 23) | Month 6 (n = 22) | Statistical Analysis of the Changes |
|---|---|---|---|---|---|---|---|
| EAA | |||||||
| Histidine | 1872.9 ± 53.9 | 1530.9 ± 31.6 | 1462.9 ± 62.2 | 1368.1 ± 58.6 | 1278.0 ± 40.0 | 1259.7 ± 38.8 | b **, d ** |
| Isoleucine | 5112.8 ±145.9 | 4235.2 ± 98.1 | 4071.5 ± 162.7 | 3825.6 ± 167.7 | 3544.6 ± 109.6 | 3339.8 ± 79.2 | b **, d ** |
| Leucine | 9774.1 ±276.8 | 8094.0 ± 190.7 | 7748.7 ± 314.5 | 7299.3 ± 318.2 | 6813.3 ± 208.4 | 6667.9 ± 191.1 | b **, d ** |
| Lysine | 6281.0 ±169.4 | 5154.2 ± 123.1 | 4902.8 ± 202.9 | 4578.6 ± 195.9 | 4281.1 ± 128.7 | 4259.6 ± 136.7 | b **, d ** |
| Methionine | 1352.4 ±41.6 | 1068.5 ± 25.8 | 1028.2 ± 52.1 | 944.2 ± 43.2 | 879.8 ±3 1.0 | 832.6 ± 25.3 | b **, d ** |
| Phenylalanine | 3021.2±91.9 | 2452.1 ± 61.3 | 2309.4 ± 107.9 | 2154.0 ± 93.3 | 2050.6 ± 62.3 | 2089.5 ± 89.3 | b **, d ** |
| Threonine | 4800.2 ±142.3 | 3932.4 ± 94.4 | 3717.7 ± 166.6 | 3470.8 ± 144.1 | 3317.5 ± 96.5 | 3393.9 ± 146.2 | b **, d ** |
| Valine | 5727.5 ±181.5 | 4691.9 ± 103.8 | 4546.9 ± 204.6 | 4289.8 ± 187.3 | 4072.3 ± 121.2 | 4041.1 ± 141.5 | b **, d ** |
| NEAA | |||||||
| Alanine | 5573.0 ± 173.6 | 4539.4 ±108.2 | 4280.9 ± 200.3 | 4015.8 ± 169.5 | 3891.3 ± 111.7 | 4068.7 ± 205.9 | b **, d ** |
| Arginine | 2730.2 ± 104.5 | 2127.9 ± 64.6 | 2007.9 ± 117.4 | 1847.8 ± 87.7 | 1840.2 ± 64.1 | 1993.2 ± 140.6 | b **, d ** |
| Asx | 9174.8 ± 278.4 | 7383.2 ± 165.1 | 6932.7 ± 315.1 | 6475.7 ±2 74.2 | 6171.5 ± 182.2 | 6189.0 ± 244.3 | b **, d ** |
| Glx | 15,044.7 ± 397.7 | 13,123.7 ± 244.1 | 12,885.6 ± 433.4 | 12,331.7 ± 443.1 | 11,573.0 ± 294.3 | 11,274.9 ± 238.1 | b **, d ** |
| Glycine | 4115.1 ± 130.9 | 3331.5 ± 95.2 | 3105.4 ± 161.5 | 2886.6 ± 121.0 | 2808.1 ± 87.1 | 3023.6 ± 180.1 | b **, d ** |
| Serine | 5870.4 ± 180.6 | 4769.1 ± 111.4 | 4480.4 ± 202.7 | 4177.7 ± 172.4 | 3990.0 ± 118.2 | 4122.6 ± 200.5 | b **, d ** |
| Tyrosine | 2848.9 ± 89.2 | 2285.1 ± 54.1 | 2204.0 ± 98.7 | 2044.1 ± 92.7 | 1949.8 ± 59.9 | 1929.7 ± 72.7 | b **, d ** |
| SUM | |||||||
| TAA (mmol/L) | 83.3 ± 2.4 | 68.7 ± 1.5 | 65.7 ±2 .8 | 61.7 ± 2.5 | 58.5 ± 1.7 | 58.5 ± 2.0 | b **, d ** |
| Protein (g/L) | 14.4 ± 0.4 | 11.8 ± 0.2 | 11.3 ± 0.4 | 10.5 ± 0.4 | 10.0 ± 0.3 | 9.9 ± 0.3 | b **, d ** |
| FAA/TAA Ratio (%) | Month 1 (n = 22) | Month 2 (n = 25) | Month 3 (n = 24) | Month 4 (n = 18) | Month 5 (n = 23) | Month 6 (n = 22) | Statistical Analysis of the Changes |
|---|---|---|---|---|---|---|---|
| EAA | |||||||
| Histidine | 1.26 ± 0.083 | 1.45 ± 0.093 | 1.47 ± 0.095 | 1.41 ± 0.074 | 1.39 ± 0.058 | 1.50 ± 0.10 | c * |
| Isoleucine | 0.24 ± 0.040 | 0.30 ± 0.038 | 0.25 ± 0.022 | 0.33 ± 0.021 | 0.30 ± 0.025 | 0.36 ± 0.039 | a o |
| Leucine | 0.36 ± 0.047 | 0.46 ± 0.048 | 0.42 ± 0.031 | 0.58 ± 0.10 | 0.47 ± 0.037 | 0.60 ± 0.10 | a * |
| Lysine | 0.50 ± 0.067 | 0.57 ± 0.10 | 0.45 ± 0.041 | 0.63 ± 0.16 | 0.58 ± 0.052 | 0.64 ± 0.082 | a o |
| Methionine | 0.17 ± 0.038 | 0.28 ± 0.041 | 0.26 ± 0.026 | 0.44 ± 0.10 | 0.39 ± 0.038 | 0.40 ± 0.044 | a ** |
| Phenylalanine | 0.61 ± 0.074 | 0.80 ± 0.071 | 0.74 ± 0.050 | 0.87 ± 0.084 | 0.76 ± 0.044 | 0.85 ± 0.066 | a o |
| Threonine | 1.41 ± 0.11 | 2.09 ± 0.17 | 2.13 ± 0.16 | 2.38 ± 0.17 | 2.58 ± 0.16 | 2.50 ± 0.20 | a **, c ** |
| Valine | 0.95 ± 0.076 | 1.27 ± 0.092 | 1.19 ± 0.075 | 1.35 ± 0.11 | 1.12 ± 0.057 | 1.20 ± 0.080 | c * |
| NEAA | |||||||
| Alanine | 4.54 ± 0.41 | 5.28 ± 0.31 | 5.84 ± 0.35 | 6.39 ± 0.32 | 6.35 ± 0.037 | 5.81 ± 0.39 | a *, c ** |
| Arginine | 0.60 ± 0.054 | 0.75 ± 0.12 | 0.68 ± 0.060 | 1.11 ± 0.31 | 0.86 ± 0.12 | 0.84 ± 0.072 | a * |
| Asx | 0.55 ± 0.064 | 0.84 ± 0.079 | 1.14 ± 0.17 | 1.21 ± 0.14 | 1.49 ± 0.17 | 1.33 ± 0.18 | a **, c ** |
| Glx | 9.12 ± 0.77 | 13.81 ± 0.66 | 15.69 ± 0.76 | 17.50 ± 0.3 | 17.97 ± 0.69 | 18.33 ± 0.90 | a **, c ** |
| Glycine | 2.04 ± 0.19 | 2.86 ± 0.18 | 3.37 ± 0.20 | 3.63 ± 0.16 | 3.77 ± 0.20 | 3.88 ± 0.27 | a **, c ** |
| Serine | 1.39 ± 0.14 | 2.08 ± 0.15 | 2.50 ± 0.21 | 2.60 ± 0.12 | 2.59 ± 0.17 | 2.57 ± 0.21 | a **, c ** |
| Tyrosine | 0.59 ± 0.064 | 0.71 ± 0.076 | 0.64 ± 0.047 | 0.72 ± 0.076 | 0.70 ± 0.059 | 0.85 ± 0.070 | a * |
| Total | |||||||
| FAA to TAA | 2.56 ± 0.20 | 3.81 ± 0.19 | 4.36 ± 0.23 | 4.91 ± 0.26 | 4.97 ± 0.20 | 5.04 ± 0.29 | a **, c ** |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van Sadelhoff, J.H.J.; Van de Heijning, B.J.M.; Stahl, B.; Amodio, S.; Rings, E.H.H.M.; Mearin, M.L.; Garssen, J.; Hartog, A. Longitudinal Variation of Amino Acid Levels in Human Milk and Their Associations with Infant Gender. Nutrients 2018, 10, 1233. https://doi.org/10.3390/nu10091233
Van Sadelhoff JHJ, Van de Heijning BJM, Stahl B, Amodio S, Rings EHHM, Mearin ML, Garssen J, Hartog A. Longitudinal Variation of Amino Acid Levels in Human Milk and Their Associations with Infant Gender. Nutrients. 2018; 10(9):1233. https://doi.org/10.3390/nu10091233
Chicago/Turabian StyleVan Sadelhoff, Joris H. J., Bert J. M. Van de Heijning, Bernd Stahl, Sonia Amodio, Edmond H. H. M. Rings, M. Luisa Mearin, Johan Garssen, and Anita Hartog. 2018. "Longitudinal Variation of Amino Acid Levels in Human Milk and Their Associations with Infant Gender" Nutrients 10, no. 9: 1233. https://doi.org/10.3390/nu10091233
APA StyleVan Sadelhoff, J. H. J., Van de Heijning, B. J. M., Stahl, B., Amodio, S., Rings, E. H. H. M., Mearin, M. L., Garssen, J., & Hartog, A. (2018). Longitudinal Variation of Amino Acid Levels in Human Milk and Their Associations with Infant Gender. Nutrients, 10(9), 1233. https://doi.org/10.3390/nu10091233