Beneficial Effect of Intestinal Fermentation of Natural Polysaccharides




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Intestinal Microbiota
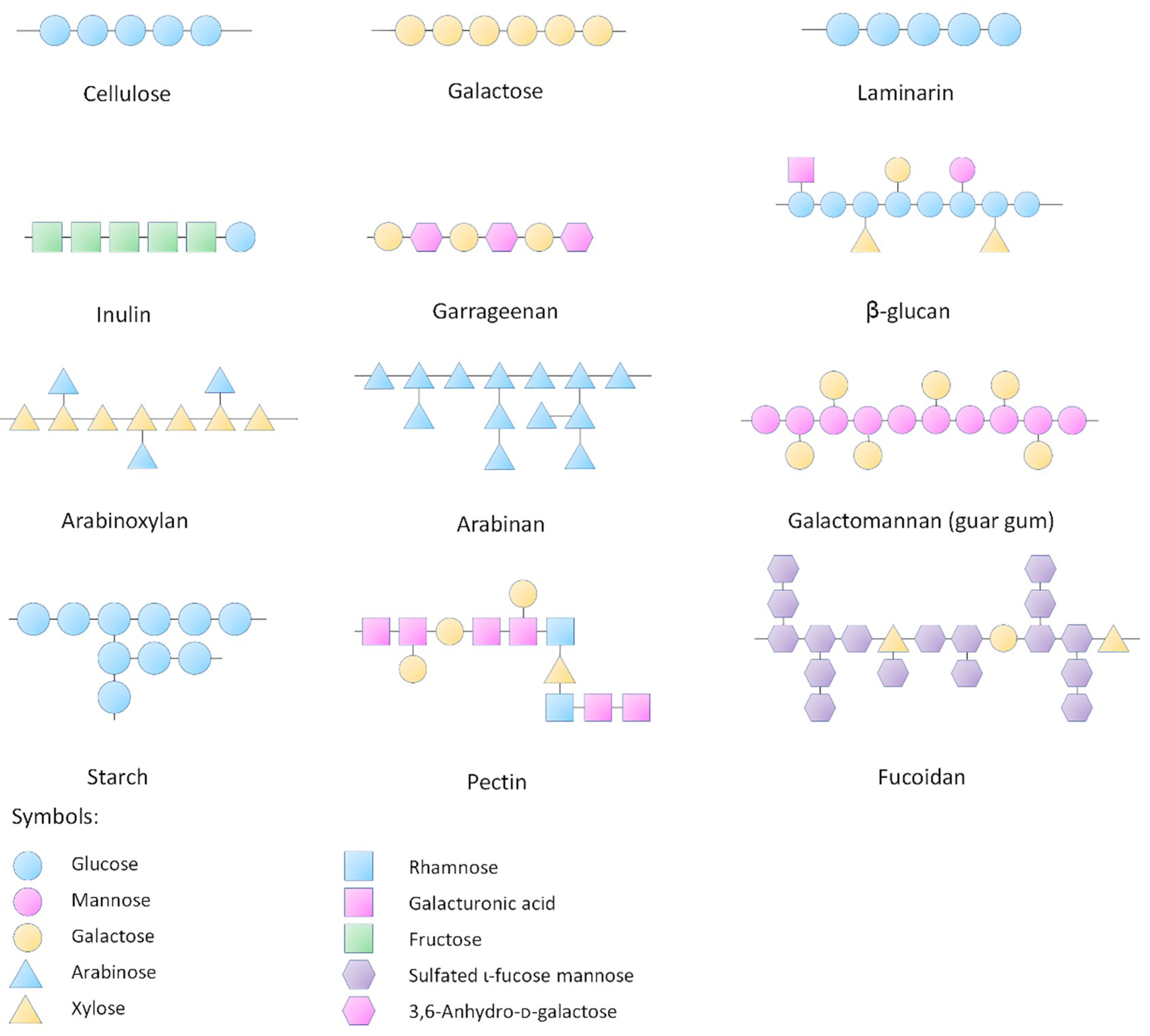
3. Influences of Natural Polysaccharides on Intestinal Microbiota
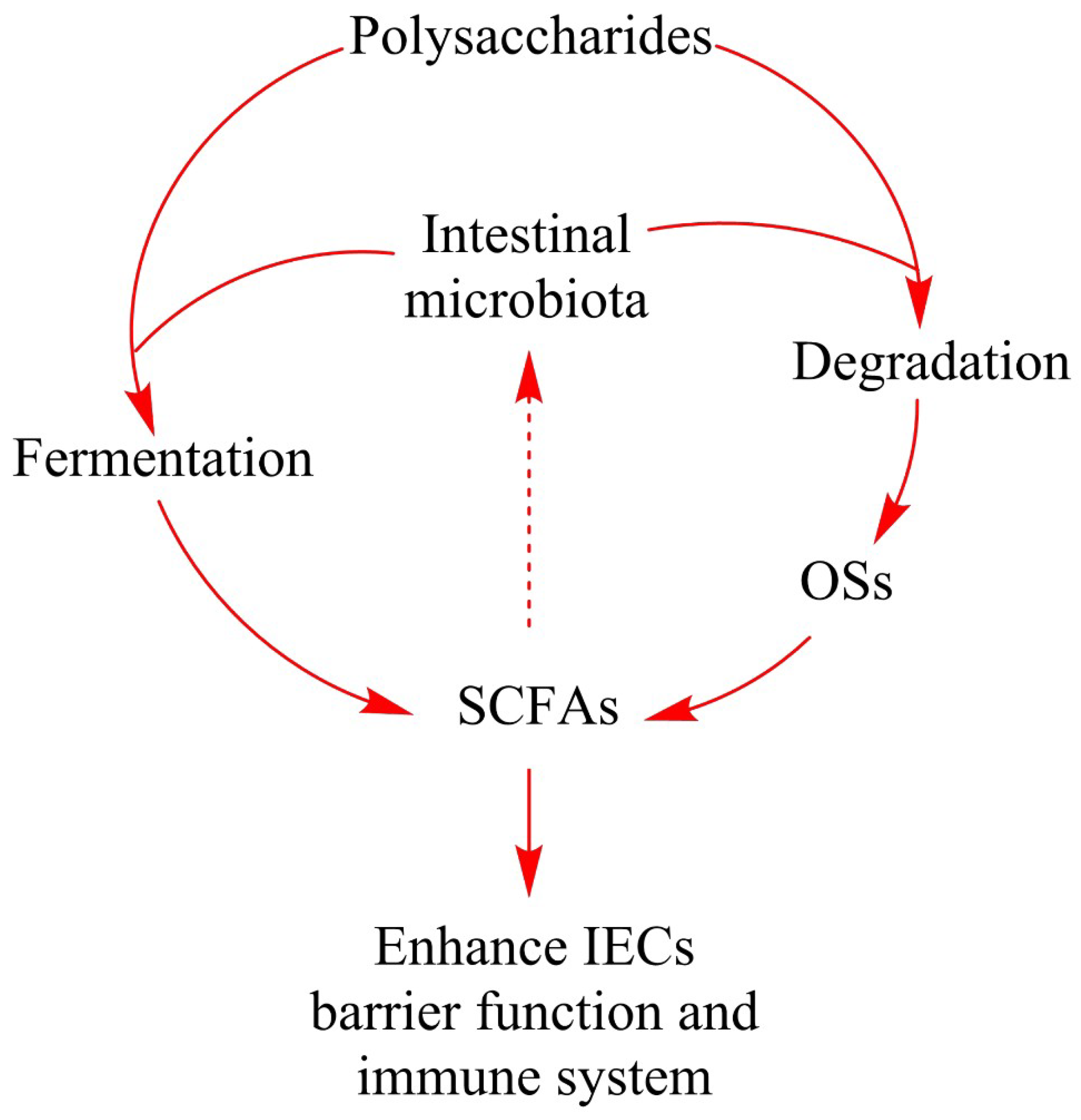
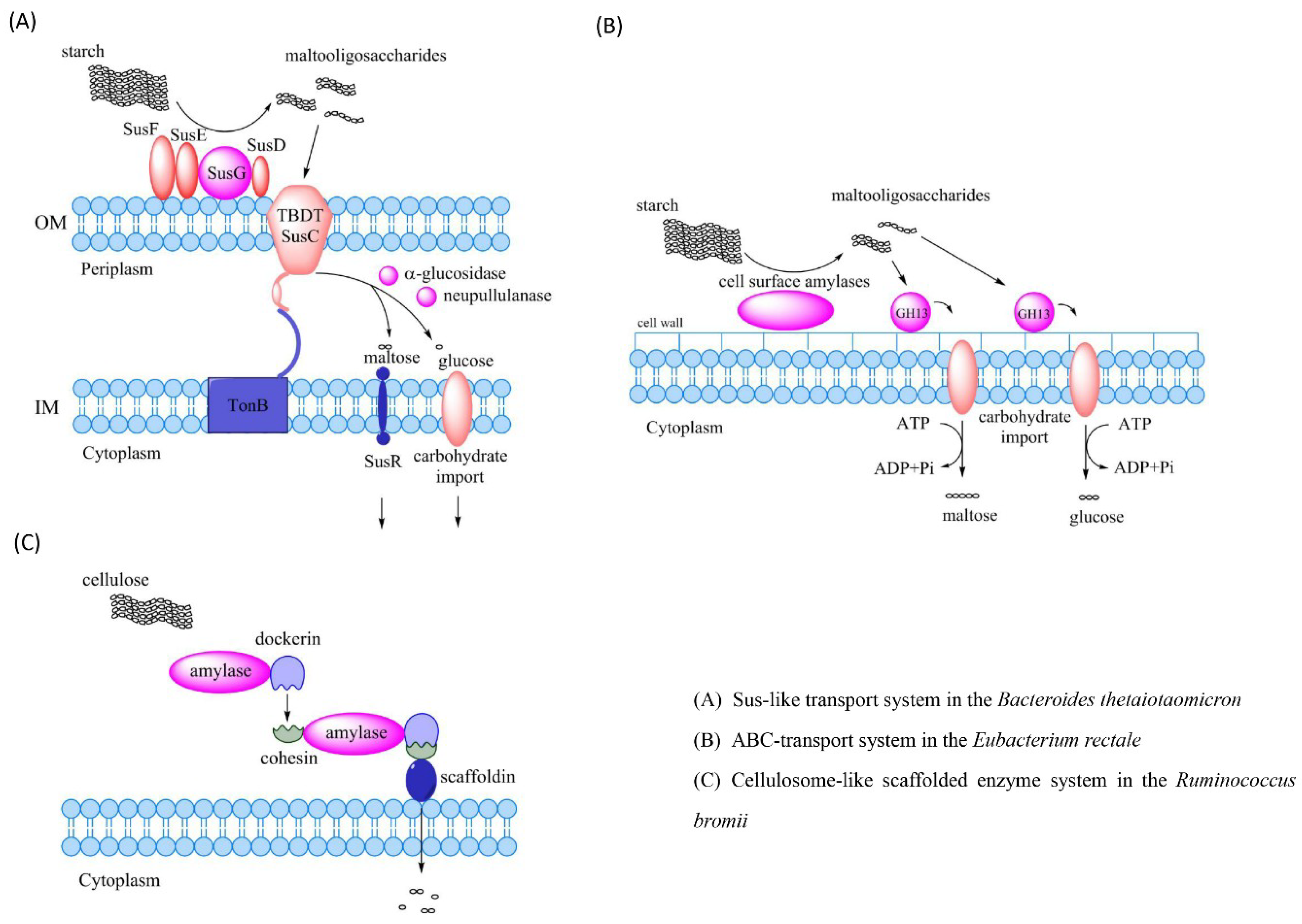
4. Polysaccharide Degradation by the Intestinal Microbiota
5. Production of Short-Chain Fatty Acids during Intestinal Fermentation
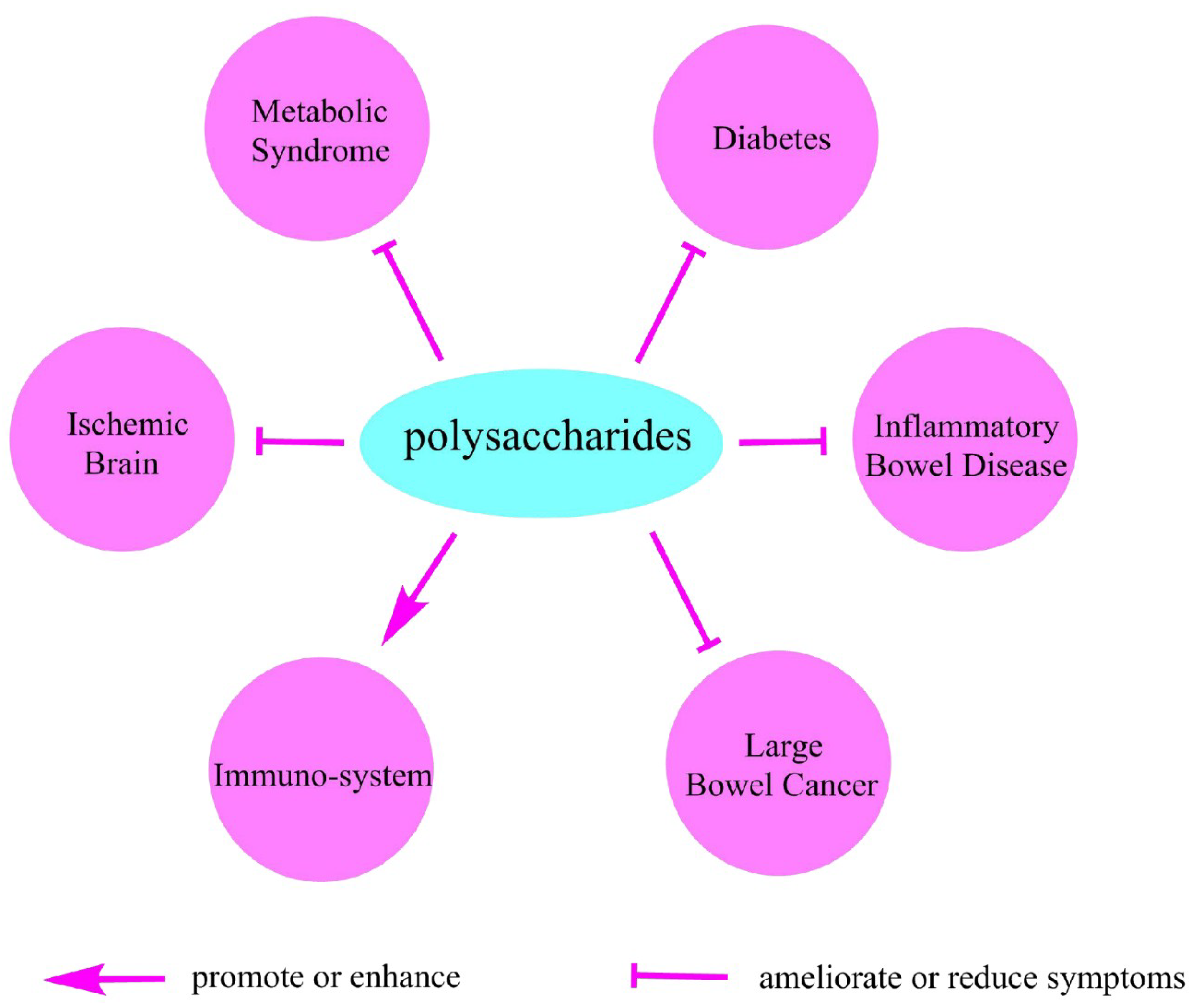
6. Polysaccharides and Health
6.1. Metabolic Syndrome
6.2. Diabetes
6.3. Inflammatory Bowel Disease
6.4. Large Bowel Cancer
6.5. Polysaccharides Regulate Immuno-System
6.6. Ischemic Brain
7. Conclusions
Funding
Conflicts of Interest
References
- Gilbert, J.A.; Quinn, R.A.; Debelius, J.; Xu, Z.Z.; Morton, J.; Garg, N.; Jansson, J.K.; Dorrestein, P.C.; Knight, R. Microbiome-wide association studies link dynamic microbial consortia to disease. Nature 2016, 535, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Hornung, B.; Santos, V.A.P.M.D.; Smidt, H.; Schaap, P.J. Studying microbial functionality within the gut ecosystem by systems biology. Genes Nutr. 2018, 13, 5. [Google Scholar] [CrossRef] [PubMed]
- Partida-Rodríguez, O.; Serrano-Vázquez, A.; Nieves-Ramírez, M.E.; Moran, P.; Rojas, L.; Portillo, T.; González, E.; Hernández, E.; Finlay, B.B.; Ximenez, C. Human intestinal microbiota: Interaction between parasites and the host immune response. Arch. Med. Res. 2017, 48, 690–700. [Google Scholar] [CrossRef] [PubMed]
- Shanahan, F. The host-microbe interface within the gut. Best Pract. Res. Clin. Gastroenterol. 2002, 16, 915–931. [Google Scholar] [CrossRef] [PubMed]
- Turroni, F.; Milani, C.; Duranti, S.; Mancabelli, L.; Mangifesta, M.; Viappiani, A.; Lugli, G.A.; Ferrario, C.; Gioiosa, L.; Ferrarini, A. Deciphering bifidobacterial-mediated metabolic interactions and their impact on gut microbiota by a multi-omics approach. ISME J. 2016, 10, 1656–1668. [Google Scholar] [CrossRef] [PubMed]
- Rivière, A.; Gagnon, M.; Weckx, S.; Roy, D.; De, V.L. Mutual cross-feeding interactions between Bifidobacterium longum subsp. Longum NCC2705 and Eubacterium rectale ATCC 33656 explain the bifidogenic and butyrogenic effects of arabinoxylan oligosaccharides. Appl. Environ. Microbiol. 2015, 81, 7767. [Google Scholar] [CrossRef] [PubMed]
- Dwidar, M.; Yokobayashi, Y. Controlling bdellovibrio bacteriovorus gene expression and predation using synthetic riboswitches. ACS Synth. Biol. 2017, 6, 2035–2041. [Google Scholar] [CrossRef] [PubMed]
- Atterbury, R.J.; Hobley, L.; Till, R.; Lambert, C.; Capeness, M.J.; Lerner, T.R.; Fenton, A.K.; Barrow, P.; Sockett, R.E. Effects of orally administered bdellovibrio bacteriovorus on the well-being and salmonella colonization of young chicks. Appl. Environ. Microbiol. 2011, 77, 5794–5803. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, stability and resilience of the human gut microbiota. Nature 2012, 489, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Bouwknegt, M.; Van, D.S.; Kuijper, E. Burden of clostridium difficile infection in the united states. N. Engl. J. Med. 2015, 372, 825–834. [Google Scholar]
- Stein, R.R.; Bucci, V.; Toussaint, N.C.; Buffie, C.G.; Rätsch, G.; Pamer, E.G.; Sander, C.; Xavier, J.B. Ecological modeling from time-series inference: Insight into dynamics and stability of intestinal microbiota. PLoS Comput. Biol. 2013, 9, e1003388. [Google Scholar] [CrossRef] [PubMed]
- Voth, D.E.; Ballard, J.D. Clostridium difficile toxins: Mechanism of action and role in disease. Clin. Microbiol. Rev. 2005, 18, 247–263. [Google Scholar] [CrossRef] [PubMed]
- Chung, W.S.F.; Walker, A.W.; Louis, P.; Parkhill, J.; Vermeiren, J.; Bosscher, D.; Duncan, S.H.; Flint, H.J. Modulation of the human gut microbiota by dietary fibres occurs at the species level. BMC Biol. 2016, 14, 3. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Gao, X.; Wu, C.; Tian, F.; Lei, Q.; Bi, J.; Xie, B.; Wang, H.Y.; Chen, S.; Wang, X. Apple-derived pectin modulates gut microbiota, improves gut barrier function, and attenuates metabolic endotoxemia in rats with diet-induced obesity. Nutrients 2016, 8, 126. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.Y.; Hou, R.; Yang, G.Q.; Zhao, F.; Dong, W.G. In vitro effects of inulin and soya bean oligosaccharide on skatole production and the intestinal microbiota in broilers. J. Anim. Physiol. Anim. Nutr. 2017, 102, 706–716. [Google Scholar] [CrossRef] [PubMed]
- Sarikaya, H.; Aslim, B.; Yuksekdag, Z. Assessment of anti-biofilm activity and bifidogenic growth stimulator (BGS) effect of lyophilized exopolysaccharides (L-EPSs) from lactobacilli strains. Int. J. Food Prop. 2016, 20, 362–371. [Google Scholar] [CrossRef]
- Lyte, M.; Chapel, A.; Lyte, J.M.; Ai, Y.; Proctor, A.; Jane, J.L.; Phillips, G.J. Resistant starch alters the microbiota-gut brain axis: Implications for dietary modulation of behavior. PLoS ONE 2016, 11, e0146406. [Google Scholar] [CrossRef] [PubMed]
- Ordiz, M.I.; May, T.D.; Mihindukulasuriya, K.; Martin, J.; Crowley, J.; Tarr, P.I.; Ryan, K.; Mortimer, E.; Gopalsamy, G.; Maleta, K. The effect of dietary resistant starch type 2 on the microbiota and markers of gut inflammation in rural malawi children. Microbiome 2015, 3, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Barouei, J.; Bendiks, Z.; Martinic, A.; Mishchuk, D.; Heeney, D.; Hsieh, Y.H.; Kieffer, D.; Zaragoza, J.; Martin, R.; Slupsky, C. Microbiota, metabolome, and immune alterations in obese mice fed a high-fat diet containing type 2 resistant starch. Mol. Nutr. Food Res. 2017, 61, 1700184. [Google Scholar] [CrossRef] [PubMed]
- Shtriker, M.G.; Hahn, M.; Taieb, E.; Nyska, A.; Moallem, U.; Tirosh, O.; Madar, Z. Fenugreek galactomannan and citrus pectin improve several parameters associated with glucose metabolism, and modulate gut microbiota in mice. Nutrition 2017, 46, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Garciamazcorro, J.F.; Barcenaswalls, J.R.; Suchodolski, J.S.; Steiner, J.M. Molecular assessment of the fecal microbiota in healthy cats and dogs before and during supplementation with fructo-oligosaccharides (FOS) and inulin using high-throughput 454-pyrosequencing. PeerJ 2017, 5, e3184. [Google Scholar] [CrossRef] [PubMed]
- Lamia, L.; Philippe, P.; Hichem Ben, S.; Mouledi, E.F.; Moncef, M.; Fabien, M.; Irène, M.; Antonia, S.; Ester, P.; Mohamed, H. Effect of dietary fibre of barley variety ‘rihane’ on azoxymethane-induced aberrant crypt foci development and on colonic microbiota diversity in rats. Br. J. Nutr. 2012, 108, 2034–2042. [Google Scholar]
- Cheng, W.; Lu, J.; Li, B.; Lin, W.; Zhang, Z.; Wei, X.; Sun, C.; Chi, M.; Bi, W.; Yang, B.; et al. Effect of functional oligosaccharides and ordinary dietary fiber on intestinal microbiota diversity. Front. Microbiol. 2017, 8, 1750. [Google Scholar] [CrossRef] [PubMed]
- Tannaz, G.; Nittaya, M.; Frida, F.; Margareta, N. Molecular properties of guar gum and pectin modify cecal bile acids, microbiota, and plasma lipopolysaccharide-binding protein in rats. PLoS ONE 2016, 11, e0157427. [Google Scholar]
- Zhu, L.; Qin, S.; Zhai, S.; Gao, Y.; Li, L. Inulin with different degrees of polymerization modulates composition of intestinal microbiota in mice. FEMS Microbiol. Lett. 2017, 364. [Google Scholar] [CrossRef] [PubMed]
- Shang, Q.; Jiang, H.; Cai, C.; Hao, J.; Li, G.; Yu, G. Gut microbiota fermentation of marine polysaccharides and its effects on intestinal ecology: An overview. Carbohydr. Polym. 2018, 179, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R. Linking long-term dietary patterns with gut microbial enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Kaoutari, A.E.; Armougom, F.; Gordon, J.I.; Raoult, D.; Henrissat, B. The abundance and variety of carbohydrate-active enzymes in the human gut microbiota. Nat. Rev. Microbiol. 2013, 11, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Ridaura, V.K.; Faith, J.J.; Rey, F.E.; Knight, R.; Gordon, J.I. The effect of diet on the human gut microbiome: A metagenomic analysis in humanized gnotobiotic mice. Sci. Transl. Med. 2009, 1, 6ra14. [Google Scholar] [CrossRef] [PubMed]
- Salyers, A.A.; Vercellotti, J.R.; West, S.E.; Wilkins, T.D. Fermentation of mucin and plant polysaccharides by strains of bacteroides from the human colon. Appl. Environ. Microbiol 1977, 34, 529–533. [Google Scholar] [PubMed]
- Noah, V.; Anne, K.; Weir, T.L. Diet and the development of the human intestinal microbiome. Front. Microbiol. 2014, 5, 494. [Google Scholar]
- Xu, J.; Bjursell, M.K.; Himrod, J.; Deng, S.; Carmichael, L.K.; Chiang, H.C.; Hooper, L.V.; Gordon, J.I. A genomic view of the human-bacteroides thetaiotaomicron symbiosis. Science 2003, 299, 2074. [Google Scholar] [CrossRef] [PubMed]
- Hehemann, J.H.; Kelly, A.G.; Pudlo, N.A.; Martens, E.C.; Boraston, A.B. Bacteria of the human gut microbiome catabolize red seaweed glycans with carbohydrate-active enzyme updates from extrinsic microbes. Proc. Natl. Acad. Sci. USA 2012, 109, 19786–19791. [Google Scholar] [CrossRef] [PubMed]
- Foley, M.H.; Cockburn, D.W.; Koropatkin, N.M. The sus operon: A model system for starch uptake by the human gut bacteroidetes. Cell. Mol. Life Sci. 2016, 73, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Cockburn, D.W.; Orlovsky, N.I.; Foley, M.H.; Kwiatkowski, K.J.; Bahr, C.M.; Maynard, M.; Demeler, B.; Koropatkin, N.M. Molecular details of a starch utilization pathway in the human gut symbiont eubacterium rectale. Mol. Microbiol. 2015, 95, 209–230. [Google Scholar] [CrossRef] [PubMed]
- David, Y.B.; Dassa, B.; Borovok, I.; Lamed, R.; Koropatkin, N.M.; Martens, E.C.; White, B.A.; Bernalierdonadille, A.; Duncan, S.H.; Flint, H.J. Ruminococcal cellulosome systems from rumen to human. Environ. Microbiol. 2015, 17, 3407–3426. [Google Scholar] [CrossRef] [PubMed]
- Anderson, K.L.; Salyers, A.A. Genetic evidence that outer membrane binding of starch is required for starch utilization by bacteroides thetaiotaomicron. J. Bacteriol. 1989, 171, 3199–3204. [Google Scholar] [CrossRef] [PubMed]
- Sonnenburg, J.L.; Xu, J.; Leip, D.D.; Chen, C.H.; Westover, B.P.; Weatherford, J.; Buhler, J.D.; Gordon, J.I. Glycan foraging in vivo by an intestine-adapted bacterial symbiont. Science 2005, 307, 1955–1959. [Google Scholar] [CrossRef] [PubMed]
- Reeves, A.R.; Wang, G.R.; Salyers, A.A. Characterization of four outer membrane proteins that play a role in utilization of starch by bacteroides thetaiotaomicron. J. Bacteriol. 1997, 179, 643. [Google Scholar] [CrossRef] [PubMed]
- Martens, E.C.; Lowe, E.C.; Chiang, H.; Pudlo, N.A.; Wu, M.; Mcnulty, N.P.; Abbott, D.W.; Henrissat, B.; Gilbert, H.J.; Bolam, D.N. Recognition and degradation of plant cell wall polysaccharides by two human gut symbionts. PLoS Biol. 2011, 9, e1001221. [Google Scholar] [CrossRef] [PubMed]
- Terrapon, N.; Lombard, V.; Gilbert, H.J.; Henrissat, B. Automatic prediction of polysaccharide utilization loci in bacteroidetes species. Bioinformatics 2015, 31, 647–655. [Google Scholar] [CrossRef] [PubMed]
- Tancula, E.; Feldhaus, M.J.; Bedzyk, L.A.; Salyers, A.A. Location and characterization of genes involved in binding of starch to the surface of bacteroides thetaiotaomicron. J. Bacteriol. 1992, 174, 5609–5616. [Google Scholar] [CrossRef] [PubMed]
- Ejby, M.; Fredslund, F.; Andersen, J.M.; Henriksen, J.R.; Andersen, T.L.; Svensson, B.; Slotboom, D.J.; Hachem, M.A. An atp binding cassette transporter mediates the uptake of α-(1,6)-linked dietary oligosaccharides in bifidobacterium and correlates with competitive growth on these substrates. J. Biol. Chem. 2016, 291, 20220. [Google Scholar] [CrossRef] [PubMed]
- Garrido, D.; Kim, J.H.; German, J.B.; Raybould, H.E.; Mills, D.A. Oligosaccharide binding proteins from bifidobacterium longum subsp. Infantis reveal a preference for host glycans. PLoS ONE 2011, 6, e17315. [Google Scholar] [CrossRef] [PubMed]
- Ramsay, A.G.; Scott, K.P.; Martin, J.C.; Rincon, M.T.; Flint, H.J. Cell-associated alpha-amylases of butyrate-producing firmicute bacteria from the human colon. Microbiology 2006, 152, 3281–3290. [Google Scholar] [CrossRef] [PubMed]
- Chassard, C.; Delmas, E.; Robert, C.; Lawson, P.A.; Bernalier-Donadille, A. Ruminococcus champanellensis sp. Nov., a cellulose-degrading bacterium from human gut microbiota. Int. J. Syst. Evol. Microbiol. 2012, 62, 138. [Google Scholar] [CrossRef] [PubMed]
- Bayer, E.A.; Lamed, R.; White, B.A.; Flint, H.J. From cellulosomes to cellulosomics. Chem. Rec. 2010, 8, 364–377. [Google Scholar] [CrossRef] [PubMed]
- Bayer, E.A.; Belaich, J.P.; Shoham, Y.; Lamed, R. The cellulosomes: Multienzyme machines for degradation of plant cell wall polysaccharides. Ann. Rev. Microbiol. 2004, 58, 521–554. [Google Scholar] [CrossRef] [PubMed]
- Fierobe, H.P.; Bayer, E.A.; Tardif, C.; Czjzek, M.; Mechaly, A.; Bélaïch, A.; Lamed, R.; Shoham, Y.; Bélaïch, J.P. Degradation of cellulose substrates by cellulosome chimeras. Substrate targeting versus proximity of enzyme components. J. Biol. Chem. 2002, 277, 49621. [Google Scholar] [CrossRef] [PubMed]
- Pryde, S.E.; Duncan, S.H.; Hold, G.L.; Stewart, C.S.; Flint, H.J. The microbiology of butyrate formation in the human colon. FEMS Microbiol. Lett. 2002, 217, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Pajak, B.; Orzechowski, A.; Gajkowska, B. Molecular basis of sodium butyrate-dependent proapoptotic activity in cancer cells. Adv. Med. Sci. 2007, 52, 83. [Google Scholar] [PubMed]
- Hamer, H.M.; Jonkers, D.; Venema, K.; Vanhoutvin, S.; Troost, F.J.; Brummer, R.J. Review article: The role of butyrate on colonic function. Aliment Pharmacol. Ther. 2008, 27, 104–119. [Google Scholar] [CrossRef] [PubMed]
- Maslowski, K.M.; Mackay, C.R. Diet, gut microbiota and immune responses. Nat. Immunol. 2011, 12, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Csordas, A. Butyrate, aspirin and colorectal cancer. Eur. J. Cancer Prev. 1996, 5, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Wächtershäuser, A.; Stein, J. Rationale for the luminal provision of butyrate in intestinal diseases. Eur. J. Nutr. 2000, 39, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Topping, D.L.; Clifton, P.M. Short-chain fatty acids and human colonic function: Roles of resistant starch and nonstarch polysaccharides. Physiol. Rev. 2001, 81, 1031–1064. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Macfarlane, G.T. Human Colonic Bacteria: Role in Nutrition, Physiology, and Pathology; CRC Press: Boca Raton, FL, USA, 1995. [Google Scholar]
- Perrin, P.; Pierre, F.; Patry, Y.; Champ, M.; Berreur, M.; Pradal, G.; Bornet, F.; Meflah, K.; Menanteau, J. Only fibres promoting a stable butyrate producing colonic ecosystem decrease the rate of aberrant crypt foci in rats. Gut 2001, 48, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Le, B.G.; Michel, C.; Blottière, H.M.; Cherbut, C. Prolonged intake of fructo-oligosaccharides induces a short-term elevation of lactic acid-producing bacteria and a persistent increase in cecal butyrate in rats. J. Nutr. 1999, 129, 2231–2235. [Google Scholar]
- Goodlad, R.A.; Ratcliffe, B.; Fordham, J.P.; Wright, N.A. Does dietary fibre stimulate intestinal epithelial cell proliferation in germ free rats? Gut 1989, 30, 820. [Google Scholar] [CrossRef] [PubMed]
- Louis, P.; Flint, H.J. Diversity, metabolism and microbial ecology of butyrate-producing bacteria from the human large intestine. FEMS Microbiol. Lett. 2009, 294, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Duncan, S.H.; Holtrop, G.; Lobley, G.E.; Calder, A.G.; Stewart, C.S.; Flint, H.J. Contribution of acetate to butyrate formation by human faecal bacteria. Br. J. Nutr. 2004, 91, 915–923. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, S.; Macfarlane, G.T. Regulation of short-chain fatty acid production. Proc. Nutr. Soc. 2003, 62, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Kishino, F.; Ebihara, K.; Ohya, J. Dietary fibre degradation and fermentation by two xylanolytic bacteria bacteroides xylanisolvens xb1at and Roseburia intestinalis xb6b4 from the human intestine. J. Appl. Microbiol. 2010, 109, 451–460. [Google Scholar]
- Scott, K.P.; Martin, J.C.; Campbell, G.; Mayer, C.D.; Flint, H.J. Whole-genome transcription profiling reveals genes up-regulated by growth on fucose in the human gut bacterium “Roseburia inulinivorans”. J. Bacteriol. 2006, 188, 4340. [Google Scholar] [CrossRef] [PubMed]
- Scott, K.P.; Martin, J.C.; Duncan, S.H.; Flint, H.J. Prebiotic stimulation of human colonic butyrate-producing bacteria and bifidobacteria, in vitro. FEMS Microbiol. Ecol. 2014, 87, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Kusuhara, S.; Yokoi, W.; Ito, M.; Miyazaki, K. Prebiotic potential of l-sorbose and xylitol in promoting the growth and metabolic activity of specific butyrate-producing bacteria in human fecal culture. FEMS Microbiol. Ecol. 2016, 93. [Google Scholar] [CrossRef] [PubMed]
- Ravn, J.L.; Thøgersen, J.C.; Eklöf, J.; Dan, P.; Ducatelle, R.; Immerseel, F.V.; Pedersen, N.R. Gh11 xylanase increases prebiotic oligosaccharides from wheat bran favouring butyrate-producing bacteria in vitro. Anim. Feed Sci. Technol. 2017, 226, 113–123. [Google Scholar] [CrossRef]
- Mu, C.; Zhang, L.; He, X.; Smidt, H.; Zhu, W. Dietary fibres modulate the composition and activity of butyrate-producing bacteria in the large intestine of suckling piglets. Anton. Van Leeuwenhoek 2017, 110, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Li, P.; Chen, M.; Luo, Y.; Prabhakar, M.; Zheng, H.; He, Y.; Qi, Q.; Long, H.; Zhang, Y. Fructooligosaccharide (FOS) and galactooligosaccharide (GOS) increase bifidobacterium but reduce butyrate producing bacteria with adverse glycemic metabolism in healthy young population. Sci. Rep. 2017, 7, 11789. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Li, Y.; Cai, Z.; Li, S.; Zhu, J.; Zhang, F.; Liang, S.; Zhang, W.; Guan, Y.; Shen, D. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 2012, 490, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Cai, G.; Qiu, Y.; Fei, N.; Zhang, M.; Pang, X.; Jia, W.; Cai, S.; Zhao, L. Structural segregation of gut microbiota between colorectal cancer patients and healthy volunteers. ISME J. 2012, 6, 320–329. [Google Scholar] [CrossRef] [PubMed]
- Biagi, E.; Nylund, L.; Candela, M.; Ostan, R.; Bucci, L.; Pini, E.; Nikkïla, J.; Monti, D.; Satokari, R.; Franceschi, C. Through ageing, and beyond: Gut microbiota and inflammatory status in seniors and centenarians. PLoS ONE 2010, 5, e10667. [Google Scholar] [CrossRef]
- Geirnaert, A.; Calatayud, M.; Grootaert, C.; Laukens, D.; Devriese, S.; Smagghe, G.; Vos, M.D.; Boon, N.; Wiele, T.V.D. Butyrate-producing bacteria supplemented in vitro to crohn’s disease patient microbiota increased butyrate production and enhanced intestinal epithelial barrier integrity. Sci. Rep. 2017, 7, 11450. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Nishida, A.; Fujimoto, T.; Fujii, M.; Shioya, M.; Imaeda, H.; Inatomi, O.; Bamba, S.; Andoh, A.; Sugimoto, M. Reduced abundance of butyrate-producing bacteria species in the fecal microbial community in crohn’s disease. Digestion 2016, 93, 59. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Chen, L.; Zhou, R.; Wang, X.; Song, L.; Huang, S.; Wang, G.; Xia, B. Increased proportions of bifidobacterium and the lactobacillus group and loss of butyrate-producing bacteria in inflammatory bowel disease. J. Clin. Microbiol. 2014, 52, 398–406. [Google Scholar] [CrossRef] [PubMed]
- Kumari, R.; Ahuja, V.; Paul, J. Fluctuations in butyrate-producing bacteria in ulcerative colitis patients of north india. World J. Gastroenterol. 2013, 19, 3404–3414. [Google Scholar] [CrossRef] [PubMed]
- Dillon, S.M.; Kibbie, J.; Lee, E.J.; Guo, K.; Santiago, M.L.; Austin, G.L.; Gianella, S.; Landay, A.L.; Donovan, A.M.; Frank, D.N. Low abundance of colonic butyrate-producing bacteria in hiv infection is associated with microbial translocation and immune activation. AIDS 2017, 31, 511. [Google Scholar] [CrossRef] [PubMed]
- Marteau, P. Butyrate-producing bacteria as pharmabiotics for inflammatory bowel disease. Gut 2013, 62, 1673. [Google Scholar] [CrossRef] [PubMed]
- Nylund, L.; Nermes, M.; Isolauri, E.; Salminen, S.; de Vos, W.M.; Satokari, R. Severity of atopic disease inversely correlates with intestinal microbiota diversity and butyrate-producing bacteria. Allergy 2015, 70, 241–244. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Li, D.; Feng, N.; Shamoon, M.; Sun, Z.; Lei, D.; Hao, Z.; Wei, C.; Jia, S.; Yong, Q.C. Anti-diabetic effects ofclostridium butyricumcgmcc0313.1 through promoting the growth of gut butyrate-producing bacteria in type 2 diabetic mice. Sci. Rep. 2017, 7, 7046. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, W.; Fabricius, E.M.; Schneeweiss, U. The tumour-clostridium phenomenon: 50 years of developmental research (review). Int. J. Oncol. 2006, 29, 1479–1492. [Google Scholar] [CrossRef] [PubMed]
- Minton, N.P. Clostridia in cancer therapy. Nat. Rev. Microbiol. 2003, 1, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Ohkawara, S.; Furuya, H.; Nagashima, K.; Asanuma, N.; Hino, T. Oral administration of butyrivibrio fibrisolvens, a butyrate-producing bacterium, decreases the formation of aberrant crypt foci in the colon and rectum of mice. J. Nutr. 2005, 135, 2878–2883. [Google Scholar] [CrossRef] [PubMed]
- Cummings, J.H.; Edmond, L.M.; Magee, E.A. Dietary carbohydrates and health: Do we still need the fibre concept? Clin. Nutr. Suppl. 2004, 1, 5–17. [Google Scholar] [CrossRef]
- Minamida, K.; Nishimura, M.; Miwa, K.; Nishihira, J. Effects of dietary fiber with Bacillus coagulans lilac-01 on bowel movement and fecal properties of healthy volunteers with a tendency for constipation. Biosci. Biotechnol. Biochem. 2014, 79, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Scott, K.P.; Duncan, S.H.; Flint, H.J. Dietary fibre and the gut microbiota. Nutr. Bull. 2008, 33, 201–211. [Google Scholar] [CrossRef]
- Ishizuka, S.; Tanaka, S.; Xu, H.; Hara, H. Fermentable dietary fiber potentiates the localization of immune cells in the rat large intestinal crypts. Exp. Biol. Med. 2004, 229, 876–884. [Google Scholar] [CrossRef]
- Cario, E. Bacterial interactions with cells of the intestinal mucosa: Toll-like receptors and nod2. Gut 2005, 54, 1182–1193. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Bibiloni, R.; Knauf, C.; Waget, A.; Neyrinck, A.M.; Delzenne, N.M.; Burcelin, R. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet–induced obesity and diabetes in mice. Diabetes 2008, 57, 1470–1481. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Tao, H.; Na, L.; Zang, B.; Wu, X. Soluble dietary fiber improves energy homeostasis in obese mice by remodeling the gut microbiota. Biochem. Biophys. Res. Commun. 2018, 498, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.; Kale, M.; Kim, D.H.; Kim, H.S.; Chon, J.W.; Seo, K.H.; Lee, H.G.; Yokoyama, W.; Kim, H. Anti-obesity effect of exopolysaccharides isolated from kefir grains. J. Agricult. Food Chem. 2017, 65, 10011–10019. [Google Scholar] [CrossRef] [PubMed]
- Goldsmith, F.; Guice, J.; Page, R.; Welsh, D.A.; Taylor, C.M.; Blanchard, E.E.; Meng, L.; Raggio, A.M.; Stout, R.W.; Carvajalaldaz, D. Obese zdf rats fermented resistant starch with effects on gut microbiota but no reduction in abdominal fat. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef] [PubMed]
- Kral, T.V.; Roe, L.S.; Rolls, B.J. Combined effects of energy density and portion size on energy intake in women. Am. J. Clin. Nutr. 2004, 79, 962–968. [Google Scholar] [CrossRef] [PubMed]
- Rolls, B.; Bell, E. Intake of fat and carbohydrate: Role of energy density. Eur. J. Clin. Nutr. 1999, 53, S166–S173. [Google Scholar] [CrossRef] [PubMed]
- Blundell, J.; Burley, V. Satiation, satiety and the action of fibre on food intake. Int. J. Obes. 1986, 11, 9–25. [Google Scholar]
- Burton-Freeman, B. Dietary fiber and energy regulation. J. Nutr. 2000, 130, S272–S275. [Google Scholar] [CrossRef] [PubMed]
- Konturek, S.; Konturek, P.; Pawlik, T.; Brzozowski, T. Brain-gut axis and its role in the control of food intake. J. Physiol. Pharmacol. 2004, 55, 137–154. [Google Scholar] [PubMed]
- Lin, Y.; Huybrechts, I.; Vereecken, C.; Mouratidou, T.; Valtueña, J.; Kersting, M.; González-Gross, M.; Bolca, S.; Wärnberg, J.; Cuenca-García, M. Dietary fiber intake and its association with indicators of adiposity and serum biomarkers in european adolescents: The helena study. Eur. J. Nutr. 2005, 54, 771–782. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Qiu, H.; Zhao, G.; Zhou, Y.; Zhang, Z.; Zhang, H.; Jiang, Q.; Sun, Q.; Wu, H.; Yang, L. Dietary fiber intake is associated with hba1c level among prevalent patients with type 2 diabetes in pudong new area of shanghai, china. PLoS ONE 2012, 7, e46552. [Google Scholar] [CrossRef] [PubMed]
- Brauchla, M.; Juan, W.; Story, J.; Kranz, S. Sources of dietary fiber and the association of fiber intake with childhood obesity risk (in 2–18 year olds) and diabetes risk of adolescents 12–18 year olds: Nhanes 2003–2006. J. Nutr. Metab. 2012. [Google Scholar] [CrossRef] [PubMed]
- Nelson, R.; Ihle, S.L.; Lewis, L.; Salisbury, S.; Miller, T.; Bergdall, V.; Bottoms, G. Effects of dietary fiber supplementation on glycemic control in dogs with alloxan-induced diabetes mellitus. Am. J. Veter. Res. 1991, 52, 2060–2066. [Google Scholar]
- Würsch, P.; Pi-Sunyer, F.X. The role of viscous soluble fiber in the metabolic control of diabetes: A review with special emphasis on cereals rich in β-glucan. Diabetes Care 1997, 20, 1774–1780. [Google Scholar] [CrossRef] [PubMed]
- Flammang, A.M.; Kendall, D.M.; Baumgartner, C.J.; Slagle, T.D.; Choe, Y.S. Effect of a viscous fiber bar on postprandial glycemia in subjects with type 2 diabetes. J. Am. Coll. Nutr. 2006, 25, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Riccardi, G.; Rivellese, A.A. Effects of dietary fiber and carbohydrate on glucose and lipoprotein metabolism in diabetic patients. Diabetes Care 1991, 14, 1115–1125. [Google Scholar] [CrossRef] [PubMed]
- Uebelhack, R.; Busch, R.; Alt, F.; Beah, Z.-M.; Chong, P.-W. Effects of cactus fiber on the excretion of dietary fat in healthy subjects: A double blind, randomized, placebo-controlled, crossover clinical investigation. Curr. Ther. Res. 2014, 76, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Story, J.A.; Kritchevsky, D. Bile acid metabolism and fiber. Am. J. Clin. Nutr. 1978, 31, S199–S202. [Google Scholar] [CrossRef] [PubMed]
- Torcello-Gómez, A.; Foster, T.J. Interactions between cellulose ethers and a bile salt in the control of lipid digestion of lipid-based systems. Carbohydr. Polym. 2014, 113, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.J.; He, Q.Y.; Luo, A.S.; Wang, M.Y.; Luo, A.X. Characterization and antihyperglycemic activity of a polysaccharide from dioscorea opposita thunb roots. Int. J. Mol. Sci. 2015, 16, 6391–6401. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Gao, H.; Wang, S.; Wen, S.; Qin, S. Hypolipidemic and antioxidant properties of a polysaccharide fraction from enteromorpha prolifera. Int. J. Biol. Macromol. 2013, 58, 186–189. [Google Scholar] [CrossRef] [PubMed]
- Raish, M. Momordica charantia polysaccharides ameliorate oxidative stress, hyperlipidemia, inflammation, and apoptosis during myocardial infarction by inhibiting the Nf-κb signaling pathway. Int. J. Biol. Macromol. 2017, 97, 544–551. [Google Scholar] [CrossRef] [PubMed]
- Ji, W.C.; Synytsya, A.; Capek, P.; Bleha, R.; Pohl, R.; Yong, I.P. Structural analysis and anti-obesity effect of a pectic polysaccharide isolated from korean mulberry fruit Oddi (Morus alba L.). Carbohydr. Polym. 2016, 146, 187–196. [Google Scholar]
- Kolsi, R.B.A.; Gara, A.B.; Chaaben, R.; Feki, A.E.; Patti, F.P.; Feki, L.E.; Belghith, K. Anti-obesity and lipid lowering effects of cymodocea nodosa sulphated polysaccharide on high cholesterol-fed-rats. Arch. Int. Physiol. 2015, 121, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Kolsi, R.B.A.; Jardak, N.; Hajkacem, F.; Chaaben, R.; Jribi, I.; Feki, A.E.; Rebai, T.; Jamoussi, K.; Fki, L.; Belghith, H.; et al. Anti-obesity effect and protection of liver-kidney functions by codium fragile sulphated polysaccharide on high fat diet induced obese rats. Int. J. Biol. Macromol. 2017, 102, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.; Wei, B.; Teng, J.; Xia, N.; Zhao, M.; Huang, L.; Ye, Y. Polysaccharides from chinese liupao dark tea and their protective effect against hyperlipidemia. Int. J. Food Sci. Technol. 2017, 53, 599–607. [Google Scholar] [CrossRef]
- Wang, Y.; Zhu, Y.; Ruan, K.; Wei, H.; Feng, Y. Mdg-1, a polysaccharide from Ophiopogon japonicus, prevents high fat diet-induced obesity and increases energy expenditure in mice. Carbohydr. Polym. 2014, 114, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Wang, J.; Wang, Y.; Feng, Y. Mdg-1, an ophiopogon polysaccharide, alleviates hyperlipidemia in mice based on metabolic profile of bile acids. Carbohydr. Polym. 2016, 150, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Wild, S.; Roglic, G.; Green, A.; Sicree, R.; King, H. Global prevalence of diabetes: Estimates for the year 2000 and projections for 2030. Diab. Care 2004, 27, 1047–1053. [Google Scholar] [CrossRef]
- Amos, A.F.; McCarty, D.J.; Zimmet, P. The rising global burden of diabetes and its complications: Estimates and projections to the year 2010. Diab. Med. 1997, 14, S7–S85. [Google Scholar] [CrossRef]
- Arathuzik, G.G.; Goebel-Fabbri, A.E. Nutrition therapy and the management of obesity and diabetes: An update. Curr. Diabetes Rep. 2011, 11, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Rahati, S.; Shahraki, M.; Arjomand, G.; Shahraki, T. Food pattern, lifestyle and diabetes mellitus. Int. J. High Risk Behav. Addict 2014, 3, e8725. [Google Scholar] [CrossRef] [PubMed]
- Marcason, W. What is the role of carbohydrates in the management of diabetes? J. Acad. Nutr. Diet. 2014, 114, 1696. [Google Scholar] [CrossRef] [PubMed]
- Kaline, K.; Bornstein, S.; Bergmann, A.; Hauner, H.; Schwarz, P. The importance and effect of dietary fiber in diabetes prevention with particular consideration of whole grain products. Horm. Metab. Res. 2007, 39, 687–693. [Google Scholar] [CrossRef] [PubMed]
- Weickert, M.; Pfeiffer, A. preventing type 2 diabetes: What does dietary fiber achieve? MMW Fortschr. Med. 2005, 147, 28–30. [Google Scholar] [PubMed]
- Narayan, S.; Lakshmipriya, N.; Vaidya, R.; Bai, M.R.; Sudha, V.; Krishnaswamy, K.; Unnikrishnan, R.; Anjana, R.M.; Mohan, V. Association of dietary fiber intake with serum total cholesterol and low density lipoprotein cholesterol levels in urban asian-indian adults with type 2 diabetes. Indian J. Endocrinol. Metab. 2014, 18, 624–630. [Google Scholar] [PubMed]
- Maghsoudi, Z.; Azadbakht, L. How dietary patterns could have a role in prevention, progression, or management of diabetes mellitus? Review on the current evidence. J. Res. Med. Sci. 2012, 17, 694–709. [Google Scholar] [PubMed]
- Pcsolyar, N.S.; De Jonghe, B.C. Examining the use of dietary fiber in reducing the risk of type 2 diabetes mellitus in latino youth. J. Transc. Nurs. 2013, 25, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.S.; Qi, L.; Fahey, G.C.; Klurfeld, D.M. Consumption of cereal fiber, mixtures of whole grains and bran, and whole grains and risk reduction in type 2 diabetes, obesity, and cardiovascular disease. Am. J. Clin. Nutr. 2013, 98, 594–619. [Google Scholar] [CrossRef] [PubMed]
- Wannamethee, S.G.; Whincup, P.H.; Thomas, M.C.; Sattar, N. Associations between dietary fiber and inflammation, hepatic function, and risk of type 2 diabetes in older men potential mechanisms for the benefits of fiber on diabetes risk. Diabetes Care 2009, 32, 1823–1825. [Google Scholar] [CrossRef] [PubMed]
- Mahalle, N.; Kulkarni, M.V.; Naik, S.S.; Garg, M.K. Association of dietary factors with insulin resistance and inflammatory markers in subjects with diabetes mellitus and coronary artery disease in indian population. J. Diabetes Comp. 2014, 28, 536–541. [Google Scholar] [CrossRef] [PubMed]
- Clemens, R.; Kranz, S.; Mobley, A.R.; Nicklas, T.A.; Raimondi, M.P.; Rodriguez, J.C.; Slavin, J.L.; Warshaw, H. Filling america’s fiber intake gap: Summary of a roundtable to probe realistic solutions with a focus on grain-based foods. J. Nutr. 2012, 142, S1390–S1401. [Google Scholar] [CrossRef] [PubMed]
- Larsen, N.; Vogensen, F.K.; van den Berg, F.W.; Nielsen, D.S.; Andreasen, A.S.; Pedersen, B.K.; Al-Soud, W.A.; Sørensen, S.J.; Hansen, L.H.; Jakobsen, M. Gut microbiota in human adults with type 2 diabetes differs from non-diabetic adults. PLoS ONE 2010, 5, e9085. [Google Scholar] [CrossRef] [PubMed]
- Bindels, L.B.; Munoz, R.R.S.; Gomesneto, J.C.; Mutemberezi, V.; Martínez, I.; Salazar, N.; Cody, E.A.; Quinterovillegas, M.I.; Kittana, H.; Reyesgavilán, C.G.D.L. Resistant starch can improve insulin sensitivity independently of the gut microbiota. Microbiome 2017, 5, 12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Yu, H.; Xiao, X.; Hu, L.; Xin, F.; Yu, X. Inulin-type fructan improves diabetic phenotype and gut microbiota profiles in rats. PeerJ 2018, 6, e4446. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Lan, Y.; Tian, C.; Zhu, Y.; Liu, H.; Wang, W.; Liu, W.; Zhang, J.; Jia, L. The characterization, renoprotection and antioxidation of enzymatic and acidic exopolysaccharides from hypsizigus marmoreus. Sci. Rep. 2018, 8, 2048. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Cao, D.; Liu, X.; Mai, W.; Lan, H.; Huo, W.; Zheng, Q. The herbal medicine cordyceps sinensis protects pancreatic beta cells from streptozotocin-induced endoplasmic reticulum stress. Can. J. Diabetes 2016, 40, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Olatunji, O.J.; Chen, H.; Tola, A.J.; Oluwaniyi, O.O. Evaluation of the antidiabetic activity of polysaccharide from cordyceps cicadae in experimental diabetic rats. Chem. Biodivers. 2018. [Google Scholar] [CrossRef] [PubMed]
- Tang, T.; Duan, X.; Ke, Y.; Zhang, L.; Shen, Y.; Hu, B.; Liu, A.; Chen, H.; Li, C.; Wu, W. Antidiabetic activities of polysaccharides from anoectochilus roxburghii and anoectochilus formosanus in stz-induced diabetic mice. Int. J. Biol. Macromol. 2018, 112, 882–888. [Google Scholar] [CrossRef] [PubMed]
- El-Salhy, M. The possible role of the gut neuroendocrine system in diabetes gastroenteropathy. Histol. Histopathol. 2002, 17, 1153–1161. [Google Scholar] [PubMed]
- Kerr, K.; Morris, C.; Burke, S.; Swanson, K. Influence of dietary fiber type and amount on energy and nutrient digestibility, fecal characteristics, and fecal fermentative end-product concentrations in captive exotic felids fed a raw beef-based diet. J. Anim. Sci. 2013, 91, 2199–2210. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.; Tian, H.; Ding, C.; Gu, L.; Wei, Y.; Gong, J.; Zhu, W.; Li, N.; Li, J. Fecal microbiota transplantation in combination with soluble dietary fiber for treatment of slow transit constipation: A pilot study. Arch. Med. Res. 2016, 47, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Asghar, S.; Yang, L.; Li, H.; Wang, Z.; Ping, Q.; Xiao, Y. Lactoferrin-coated polysaccharide nanoparticles based on chitosan hydrochloride/hyaluronic acid/peg for treating brain glioma. Carbohydr. Polym. 2017, 157, 419–428. [Google Scholar] [CrossRef] [PubMed]
- Elamin, S.; Alkhawaja, M.J.; Bukhamsin, A.Y.; Mas, I.; Abdelrahman, M.M.; Abutaleb, N.K.; Housawi, A.A. Gum arabic reduces c-reactive protein in chronic kidney disease patients without affecting urea or indoxyl sulfate levels. Int. J. Nephrol. 2017, 2017, 9501470. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, F.; Yun, H.; Zhang, H.; Zhang, Q. Effect and mechanism of fucoidan derivatives from laminaria japonica in experimental adenine-induced chronic kidney disease. J. Ethnopharmacol. 2012, 139, 807–813. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, Q.; Jin, W.; Niu, X.; Zhang, H. Effects and mechanism of low molecular weight fucoidan in mitigating the peroxidative and renal damage induced by adenine. Carbohydr. Polym. 2011, 84, 417–423. [Google Scholar] [CrossRef]
- Kemp, R.; Dunn, E.; Schultz, M. Immunomodulators in inflammatory bowel disease: An emerging role for biologic agents. Biodrugs 2013, 27, 585–590. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, G.C.; Chong, C.A.; Chong, R.Y. National estimates of the burden of inflammatory bowel disease among racial and ethnic groups in the united states. J. Crohrane Colitis 2014, 8, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Molodecky, N.A.; Soon, I.S.; Rabi, D.M.; Ghali, W.A.; Ferris, M.; Chernoff, G.; Benchimol, E.I.; Panaccione, R.; Ghosh, S.; Barkema, H.W. Increasing incidence and prevalence of the inflammatory bowel diseases with time, based on systematic review. Gastroenterology 2012, 142, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Lim, W.C.; Hanauer, S. Aminosalicylates for induction of remission or response in crohn’s disease. Cochrane Diabetes Syst. Rev. 2010, 12, CD008870. [Google Scholar]
- Sullivan, P.W.; Ghushchyan, V.H.; Globe, G.; Schatz, M. Oral corticosteroid exposure and adverse effects in asthma. Retour Au Numéro 2017, 141, 110–116.e7. [Google Scholar]
- Yue, Y.; Wu, S.; Li, Z.; Li, J.; Li, X.; Xiang, J.; Ding, H. Wild jujube polysaccharides protect against experimental inflammatory bowel disease by enabling enhanced intestinal barrier function. Food Funct. 2015, 6, 2568–2577. [Google Scholar] [CrossRef] [PubMed]
- Hung, T.V.; Suzuki, T. Dietary fermentable fiber reduces intestinal barrier defects and inflammation in colitic mice. J. Nutr. 2016, 146, 1970–1979. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Lin, Q.; Yang, T.; Nie, Y.; Li, X.; Liu, B.; Shen, J.; Liang, Y.; Tang, Y.; Luo, F. Oral administration of lentinus edodes β-glucans ameliorates dss-induced ulcerative colitis in mice via mapk-elk-1 and mapk-pparγ pathways. Food Funct. 2016, 7, 4614–4627. [Google Scholar] [CrossRef] [PubMed]
- Segarra, S.; Martínezsubiela, S.; Cerdàcuéllar, M.; Martínezpuig, D.; Muñozprieto, A.; Rodríguezfranco, F.; Rodríguezbertos, A.; Allenspach, K.; Velasco, A.; Cerón, J. Oral chondroitin sulfate and prebiotics for the treatment of canine inflammatory bowel disease: A randomized, controlled clinical trial. BMC Vet. Res. 2016, 12, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Leu, R.K.L.; Christophersen, C.T.; Somashekar, R.; Conlon, M.A.; Meng, X.Q.; Winter, J.M.; Woodman, R.J.; Mckinnon, R.; Young, G.P. Manipulation of the gut microbiota using resistant starch is associated with protection against colitis-associated colorectal cancer in rats. Carcinogenesis 2016, 37, 366–375. [Google Scholar] [CrossRef] [PubMed]
- Panebianco, C.; Adamberg, K.; Adamberg, S.; Saracino, C.; Jaagura, M.; Kolk, K.; Chio, A.G.D.; Graziano, P.; Vilu, R.; Pazienza, V. Engineered resistant-starch (ers) diet shapes colon microbiota profile in parallel with the retardation of tumor growth in in vitro and in vivo pancreatic cancer models. Nutrients 2017, 9, 331. [Google Scholar] [CrossRef] [PubMed]
- Moen, B.; Henjum, K.; Måge, I.; Knutsen, S.H.; Rud, I.; Hetland, R.B.; Paulsen, J.E. Effect of dietary fibers on cecal microbiota and intestinal tumorigenesis in azoxymethane treated a/j min/+ mice. PLoS ONE 2016, 11, e0155402. [Google Scholar] [CrossRef] [PubMed]
- Bishehsari, F.; Engen, P.A.; Preite, N.Z. Dietary fiber treatment corrects the composition of gut microbiota, promotes scfa production, and suppresses colon carcinogenesis. Genes 2018, 9, 102. [Google Scholar] [CrossRef] [PubMed]
- Desai, M.S.; Seekatz, A.M.; Koropatkin, N.M.; Kamada, N.; Hickey, C.A.; Wolter, M.; Pudlo, N.A.; Kitamoto, S.; Terrapon, N.; Muller, A. A dietary fiber-deprived gut microbiota degrades the colonic mucus barrier and enhances pathogen susceptibility. Cell 2016, 167, 1339–1353. [Google Scholar] [CrossRef] [PubMed]
- Ishiguro, S.; Uppalapati, D.; Goldsmith, Z.; Robertson, D.; Hodge, J.; Holt, H.; Nakashima, A.; Turner, K.; Tamura, M. Exopolysaccharides extracted from parachlorella kessleri inhibit colon carcinoma growth in mice via stimulation of host antitumor immune responses. PLoS ONE 2017, 12, e0175064. [Google Scholar] [CrossRef] [PubMed]
- Di, W.; Zhang, L.; Wang, S.; Yi, H.; Han, X.; Fan, R.; Zhang, Y. Physicochemical characterization and antitumour activity of exopolysaccharides produced by Lactobacillus casei sb27 from yak milk. Carbohydr. Polym. 2017, 171, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.; Yu, G.; Nie, W.; Jing, J.; Chen, L.; Chen, X. Antitumor activity and underlying mechanism of sargassum fusiforme polysaccharides in cne-bearing mice. Int. J. Biol. Macromol. 2018, 112, 516–522. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Liang, H.; Luo, L. Antitumor polysaccharides from mushrooms: A review on the structural characteristics, antitumor mechanisms and immunomodulating activities. Carbohydr. Res. 2016, 424, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Jiang, X.; Xie, H.; Li, X.; Shi, L. Structural characterization and antitumor activity of a polysaccharide from ramulus mori. Carbohydr. Polym. 2018, 190, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Moutschen, M.; Scheen, A.; Lefebvre, P. Impaired immune responses in diabetes mellitus: Analysis of the factors and mechanisms involved. Relevance to the increased susceptibility of diabetic patients to specific infections. Diabetes Métab. 1992, 18, 187–201. [Google Scholar]
- Pozzilli, P.; Leslie, R.D.G. Infections and diabetes: Mechanisms and prospects for prevention. Diabetes Med. 1994, 11, 935–941. [Google Scholar] [CrossRef]
- Geerlings, S.E.; Hoepelman, A.I. Immune dysfunction in patients with diabetes mellitus (dm). FEMS Immunol. Med. Microbiol. 1999, 26, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Koirala, J.; Khardori, R.; Khardori, N. Infections in diabetes mellitus and hyperglycemia. Infect. Dis. Clin. N. Am. 2007, 21, 617–638. [Google Scholar] [CrossRef] [PubMed]
- Graves, D.T.; Kayal, R.A. Diabetic complications and dysregulated innate immunity. Front. Biosci. A J. Virtual Libr. 2008, 13, 1227. [Google Scholar] [CrossRef]
- Geerlings, S.E.; Brouwer, E.C.; Van Kessel, K.C.; Gaastra, W.; Stolk, R.P.; Hoepelman, A.I. Cytokine secretion is impaired in women with diabetes mellitus. Eur. J. Clin. Investig. 2000, 30, 995–1001. [Google Scholar] [CrossRef]
- Lim, B.O.; Yamada, K.; Nonaka, M.; Kuramoto, Y.; Hung, P.; Sugano, M. Dietary fibers modulate indices of intestinal immune function in rats. J. Nutr. 1997, 127, 663–667. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Tang, D.; Wang, Y.; Li, X.; Hong, L.; Sun, C. Characteristics and immune-enhancing activity of pectic polysaccharides from sweet cherry (Prunus avium). Food Chem. 2018, 254, 4754. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Wang, Y.; Chen, F.; Xiao, T.; Zhang, L. Polysaccharides from diaphragma juglandis fructus: Extraction optimization, antitumor, and immune-enhancement effects. Int. J. Biol. Macromol. 2018, 115, 835–845. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Liang, Y.; Zhang, C.; Wang, K.; Tu, Y.; Chen, M.; Li, P.; Wan, J.-B.; He, C. Polysaccharide prm3 from Rhynchosia minima root enhances immune function through tlr4-nf-κb pathway. Biochim. Biophys. Acta Gen. Sub. 2018, 1862, 1751–1759. [Google Scholar] [CrossRef] [PubMed]
- Kanmani, P.; Albarracin, L.; Kobayashi, H.; Iida, H.; Komatsu, R.; Akm, H.K.; Ikedaohtsubo, W.; Suda, Y.; Aso, H.; Makino, S. Exopolysaccharides from lactobacillus delbrueckii oll1073r-1 modulate innate antiviral immune response in porcine intestinal epithelial cells. Mol. Immunol. 2017, 93, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zeng, Y.; Men, Y.; Zhang, J.; Liu, H.; Sun, Y. Structural characterization and immunomodulatory activity of exopolysaccharides from submerged culture of auricularia auricula-judae. Int. J. Biol. Macromol. 2018, 115, 978–984. [Google Scholar] [CrossRef] [PubMed]
- Yarahmadi, P.; Miandare, H.K.; Farahmand, H.; Mirvaghefi, A.; Hoseinifar, S.H. Dietary fermentable fiber upregulated immune related genes expression, increased innate immune response and resistance of rainbow trout (Oncorhynchus mykiss) against aeromonas hydrophila. Fish Shell. Immunol. 2014, 41, 326–331. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.X.; Liu, Q.Y.; Zhang, M.; Yang, Z.; Pei, X.; Wu, X.; Chen, X.; Hong, J.; Xu, K.Z. Polysaccharides from bupleurum induce immune reversal in late sepsis. Shock 2018, 49, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Pan, D.; Li, H.; Sun, Y.; Zeng, X.; Yan, B. Antioxidant and immunomodulatory activity of selenium exopolysaccharide produced by Lactococcus lactis subsp. Lactis. Food Agric. Immunol. 2013, 138, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Clarke, T.B.; Davis, K.M.; Lysenko, E.S.; Zhou, A.Y.; Yu, Y.; Weiser, J.N. Recognition of peptidoglycan from the microbiota by nod1 enhances systemic innate immunity. Nat. Med. 2010, 16, 228–231. [Google Scholar] [CrossRef] [PubMed]
- Mithieux, G.; Gautier-Stein, A. Intestinal glucose metabolism revisited. Diabetes Res. Clin. Pract. 2014, 105, 295–301. [Google Scholar] [CrossRef] [PubMed]
- De Vadder, F.; Kovatcheva-Datchary, P.; Goncalves, D.; Vinera, J.; Zitoun, C.; Duchampt, A.; Bäckhed, F.; Mithieux, G. Microbiota-generated metabolites promote metabolic benefits via gut-brain neural circuits. Cell 2014, 156, 84–96. [Google Scholar] [CrossRef] [PubMed]
- Kimura, I. Host energy regulation via scfas receptors, as dietary nutrition sensors, by gut microbiota. J. Pharm. Soc. Jpn. 2013, 134, 1037–1042. [Google Scholar] [CrossRef]
- Li, H.; Wu, F.; Miao, H.; Xiong, K. Effects of polysaccharide of gastrodia elata blume and electro-acupuncture on expressions of brain-derived neurotrophic factor and stem cell factor protein in caudate putamen of focal cerebral ischemia rats. Med. Sci. Monit. Basic Res. 2016, 22, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Su, D.; Li, S.; Zhang, W.; Wang, J.; Lv, M. Corrigendum to structural elucidation of a polysaccharide from Lonicera japonica flowers, and its neuroprotective effect on cerebral ischemia-reperfusion injury in rat. Int. J. Biol. Macromol. 2017, 99, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.J.; Jiang, H.F.; Rehman, F.U.; Zhang, J.W.; Chang, Y.; Jing, L.; Zhang, J.Z. Lycium barbarum polysaccharides decrease hyperglycemia-aggravated ischemic brain injury through maintaining mitochondrial fission and fusion balance. Int. J. Biol. Sci. 2017, 13, 901–910. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Zhu, L.; Li, T.; Tang, X.; Xiang, Y.; Han, X.; Xia, L.; Zeng, L.; Nie, J.; Huang, Y. Neuroprotective mechanisms oflycium barbarumpolysaccharides against ischemic insults by regulating nr2b and nr2a containing nmda receptor signaling pathways. Front. Cell. Neurosci. 2017, 11, 288. [Google Scholar] [CrossRef] [PubMed]
- Eckburg, P.B.; Bik, E.M.; Bernstein, C.N.; Purdom, E.; Dethlefsen, L.; Sargent, M.; Gill, S.R.; Nelson, K.E.; Relman, D.A. Diversity of the human intestinal microbial flora. Science 2005, 308, 1635–1638. [Google Scholar] [CrossRef] [PubMed]
- Pinna, C.; Stefanelli, C.; Biagi, G. In vitro effect of dietary protein level and non-digestible oligosaccharides on feline fecal microbiota. J. Anim. Sci. 2014, 92, 5593–5602. [Google Scholar] [CrossRef] [PubMed]
- Lamichhane, S.; Yde, C.C.; Forssten, S.; Ouwehand, A.; Saarinen, M.; Jensen, H.M.; Gibson, G.R.; Rastall, R.A.; Fava, F.; Bertram, H.C. Impact of dietary polydextrose fiber on the human gut metabolome. J. Agric. Food Chem. 2014, 62, 9944–9951. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.-J.L.; Anderson, J.W.; Jennings, D. Propionate may mediate the hypocholesterolemic effects of certain soluble plant fibers in cholesterol-fed rats. Exp. Biol. Med. 1984, 175, 215–218. [Google Scholar] [CrossRef]
- Mazmanian, S.K.; Liu, C.H.; Tzianabos, A.O.; Kasper, D.L. An immunomodulatory molecule of symbiotic bacteria directs maturation of the host immune system. Cell 2005, 122, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Bengmark, S. Immunonutrition: Role of biosurfactants, fiber, and probiotic bacteria. Nutrition 1998, 14, 585–594. [Google Scholar] [CrossRef]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, T.; Yang, Y.; Liang, Y.; Jiao, X.; Zhao, C. Beneficial Effect of Intestinal Fermentation of Natural Polysaccharides. Nutrients 2018, 10, 1055. https://doi.org/10.3390/nu10081055
Zhang T, Yang Y, Liang Y, Jiao X, Zhao C. Beneficial Effect of Intestinal Fermentation of Natural Polysaccharides. Nutrients. 2018; 10(8):1055. https://doi.org/10.3390/nu10081055
Chicago/Turabian StyleZhang, Tiehua, Yang Yang, Yuan Liang, Xu Jiao, and Changhui Zhao. 2018. "Beneficial Effect of Intestinal Fermentation of Natural Polysaccharides" Nutrients 10, no. 8: 1055. https://doi.org/10.3390/nu10081055
APA StyleZhang, T., Yang, Y., Liang, Y., Jiao, X., & Zhao, C. (2018). Beneficial Effect of Intestinal Fermentation of Natural Polysaccharides. Nutrients, 10(8), 1055. https://doi.org/10.3390/nu10081055