Anti-Cancer Natural Products and Their Bioactive Compounds Inducing ER Stress-Mediated Apoptosis: A Review



Abstract
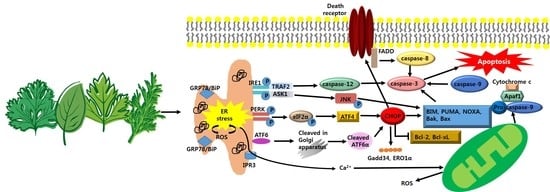
1. Introduction
Overview of ER Stress; A Double-Edged Sword-Cell Survival or Death?
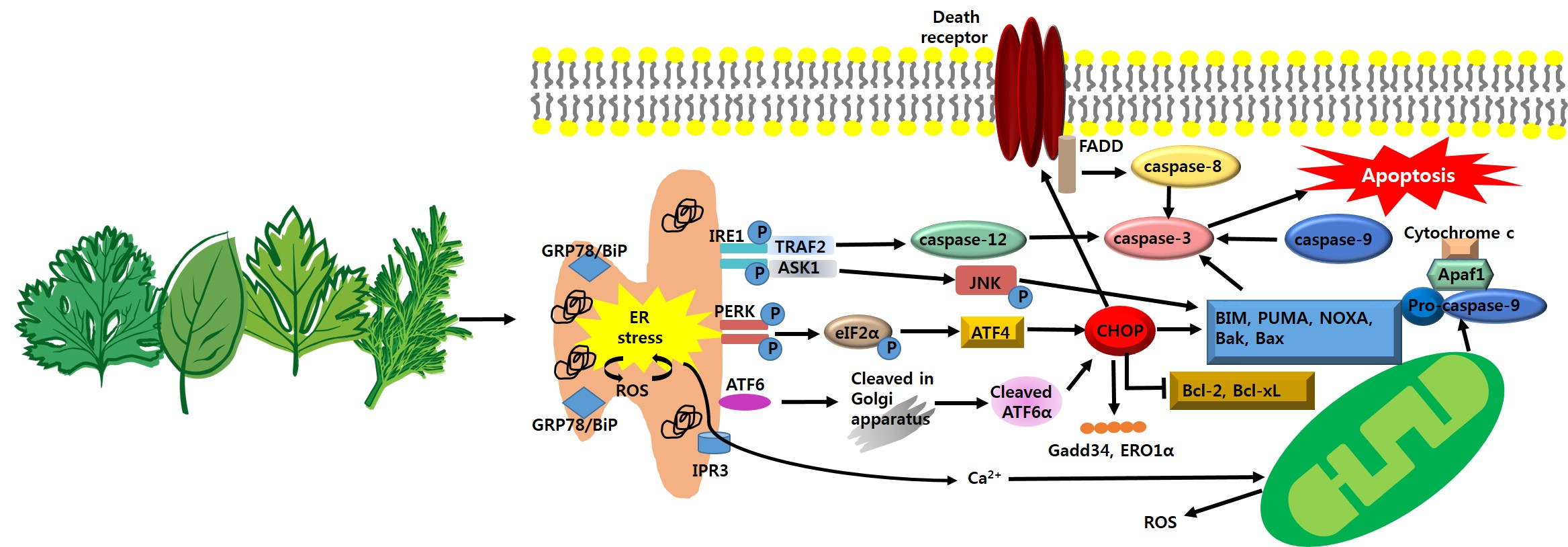
2. Anti-Cancer Strategy Based on Adaptive Pathway of ER Stress in Cancer Growth
2.1. Anti-Cancer Strategy; Targeting ATF6α-GRP78/BiP Signaling
2.2. Anti-Cancer Strategy; Targeting IRE1α-XBP1 Signaling
2.3. Anti-Cancer Strategy; Targeting PERK-eIF2α-ATF4 Signaling
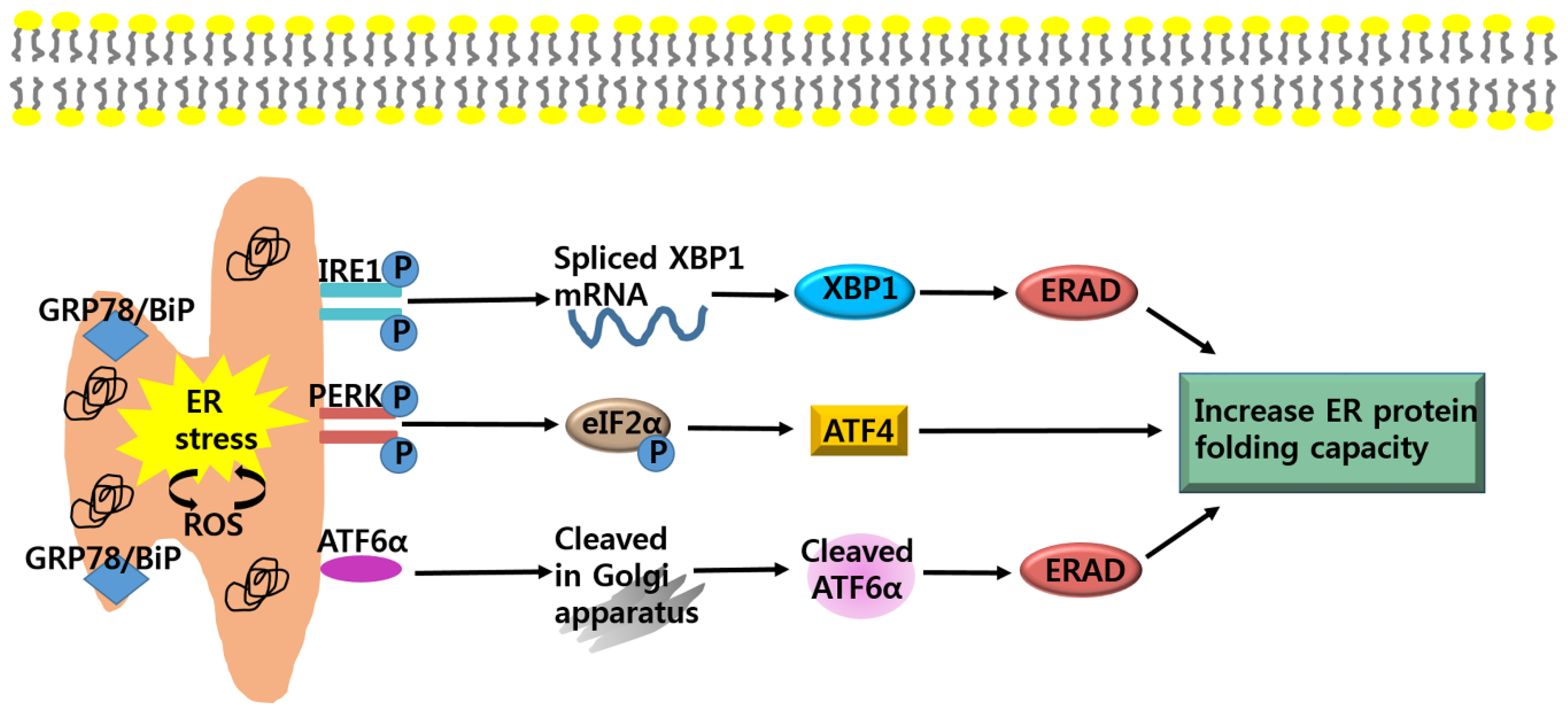
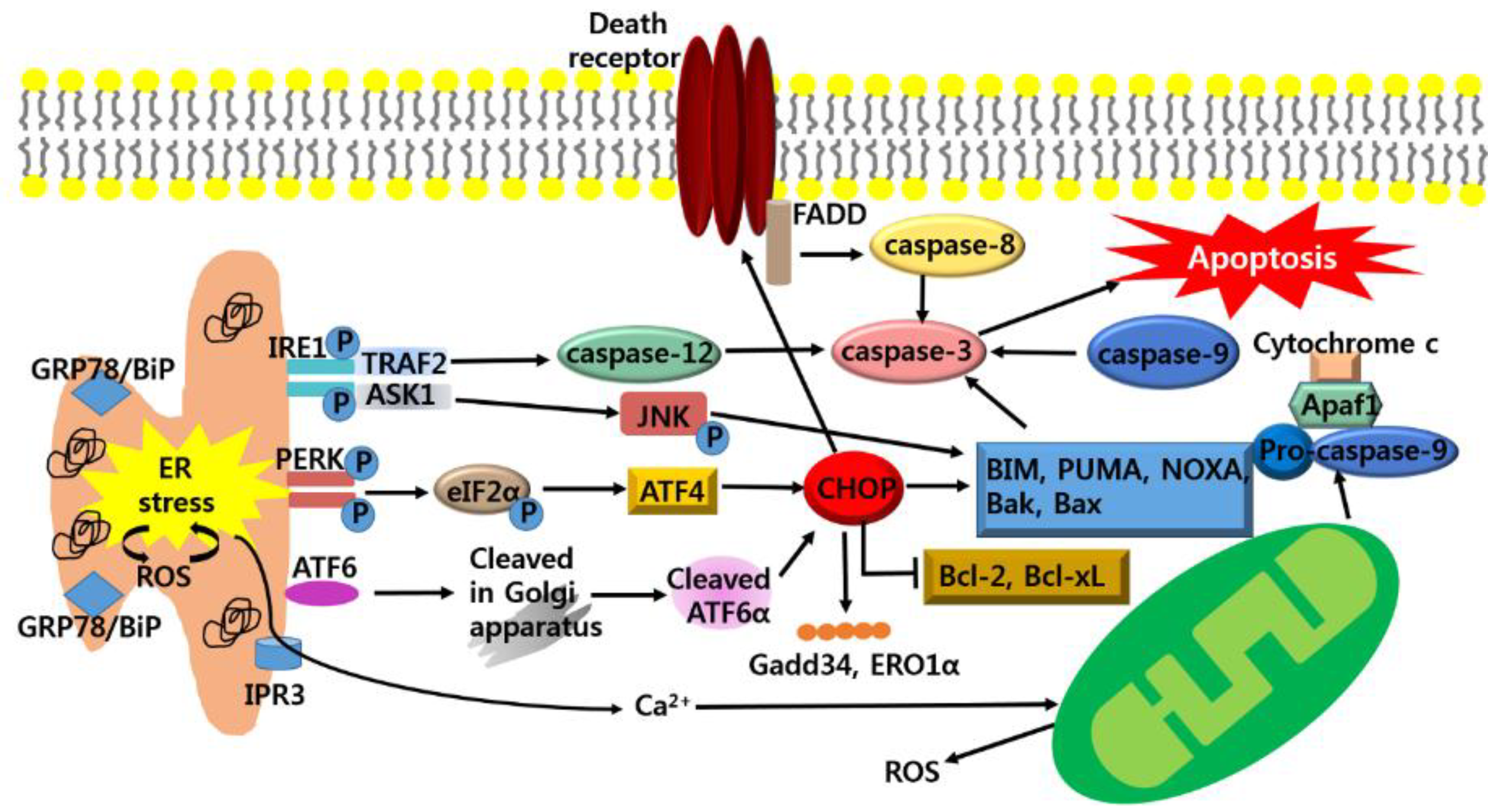
3. Anti-Cancer Strategy Based on Apoptotic Pathway of ER Stress in Cancer Growth
3.1. Anti-Cancer Strategy; Targeting CHOP-Mediated Apoptosis
3.2. Anti-Cancer Strategy; Targeting IRE1-Mediated Apoptosis
3.3. Anti-Cancer Strategy; Targeting Generation of ROS-Mediated Apoptosis
4. Anti-Cancer Effects of Natural Products via ER Stress
4.1. Search Methodology
4.2. Natural Products Targeting ER Stress-Mediated Apoptosis in Lung Cancer
4.3. Natural Products Targeting ER Stress-Mediated Apoptosis in Breast Cancer
4.4. Natural Products Targeting ER Stress-Mediated Apoptosis in Colorectal Cancer
4.5. Natural Product Targeting ER Stress-Mediated Apoptosis in Gastric Cancer
4.6. Natural Products Targeting ER Stress-Mediated Apoptosis in Prostate Cancer
4.7. Natural Product ER Stress-Mediated Apoptosis in Liver Cancer
5. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Hsu, P.P.; Sabatini, D.M. Cancer cell metabolism: Warburg and beyond. Cell 2008, 134, 703–707. [Google Scholar] [CrossRef] [PubMed]
- Stewart, B.W.; Wild, C.P. World Cancer Report 2014; World Health Organization: Lyon, France, 2016. [Google Scholar]
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in globocan 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef] [PubMed]
- Reed, J.C. Dysregulation of apoptosis in cancer. J. Clin. Oncol. 1999, 17, 2941–2953. [Google Scholar] [CrossRef] [PubMed]
- Fisher, D.E. Apoptosis in cancer therapy: Crossing the threshold. Cell 1994, 78, 539–542. [Google Scholar] [CrossRef]
- Brown, J.M.; Attardi, L.D. The role of apoptosis in cancer development and treatment response. Nat. Rev. Cancer 2005, 5, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Ghobrial, I.M.; Witzig, T.E.; Adjei, A.A. Targeting apoptosis pathways in cancer therapy. CA Cancer J. Clin. 2005, 55, 178–194. [Google Scholar] [CrossRef] [PubMed]
- Fulda, S.; Debatin, K.-M. Extrinsic versus intrinsic apoptosis pathways in anticancer chemotherapy. Oncogene 2006, 25, 4798–4811. [Google Scholar] [CrossRef] [PubMed]
- Debatin, K.-M. Apoptosis pathways in cancer and cancer therapy. Cancer Immunol. Immunother. 2004, 53, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.; Xu, W.; Reed, J.C. Cell death and endoplasmic reticulum stress: Disease relevance and therapeutic opportunities. Nat. Rev. Drug Discov. 2008, 7, 1013–1030. [Google Scholar] [CrossRef] [PubMed]
- Schröder, M.; Kaufman, R.J. The mammalian unfolded protein response. Annu. Rev. Biochem. 2005, 74, 739–789. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, R.J. Orchestrating the unfolded protein response in health and disease. J. Clin. Investig. 2002, 110, 1389–1398. [Google Scholar] [CrossRef] [PubMed]
- Hebert, D.N.; Molinari, M. In and out of the ER: Protein folding, quality control, degradation, and related human diseases. Physiol. Rev. 2007, 87, 1377–1408. [Google Scholar] [CrossRef] [PubMed]
- Yadav, R.K.; Chae, S.-W.; Kim, H.-R.; Chae, H.J. Endoplasmic reticulum stress and cancer. J. Cancer Prev. 2014, 19, 75–88. [Google Scholar] [CrossRef] [PubMed]
- Romero-Ramirez, L.; Cao, H.; Nelson, D.; Hammond, E.; Lee, A.-H.; Yoshida, H.; Mori, K.; Glimcher, L.H.; Denko, N.C.; Giaccia, A.J. XBP1 is essential for survival under hypoxic conditions and is required for tumor growth. Cancer Res. 2004, 64, 5943–5947. [Google Scholar] [CrossRef] [PubMed]
- Jamora, C.; Dennert, G.; Lee, A.S. Inhibition of tumor progression by suppression of stress protein GRP78/BIP induction in fibrosarcoma B/C10ME. Proc. Natl. Acad. Sci. USA 1996, 93, 7690–7694. [Google Scholar] [CrossRef] [PubMed]
- Bi, M.; Naczki, C.; Koritzinsky, M.; Fels, D.; Blais, J.; Hu, N.; Harding, H.; Novoa, I.; Varia, M.; Raleigh, J. ER stress-regulated translation increases tolerance to extreme hypoxia and promotes tumor growth. EMBO J. 2005, 24, 3470–3481. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Kaufman, R.J. The impact of the endoplasmic reticulum protein-folding environment on cancer development. Nat. Rev. Cancer 2014, 14, 581–597. [Google Scholar] [CrossRef] [PubMed]
- Urra, H.; Dufey, E.; Lisbona, F.; Rojas-Rivera, D.; Hetz, C. When ER stress reaches a dead end. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2013, 1833, 3507–3517. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Yang, Z.-Q.; Zhang, K. Endoplasmic reticulum stress response in cancer: Molecular mechanism and therapeutic potential. Am. J. Transl. Res. 2010, 2, 65–74. [Google Scholar] [PubMed]
- Sui, X.; Chen, R.; Wang, Z.; Huang, Z.; Kong, N.; Zhang, M.; Han, W.; Lou, F.; Yang, J.; Zhang, Q. Autophagy and chemotherapy resistance: A promising therapeutic target for cancer treatment. Cell Death Dis. 2013, 4, e838. [Google Scholar] [CrossRef] [PubMed]
- Martinon, F. Targeting endoplasmic reticulum signaling pathways in cancer. Acta Oncol. 2012, 51, 822–830. [Google Scholar] [CrossRef] [PubMed]
- Schindler, A.J.; Schekman, R. In vitro reconstitution of er-stress induced atf6 transport in copii vesicles. Proc. Natl. Acad. Sci. USA 2009, 106, 17775–17780. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, H.; Okada, T.; Haze, K.; Yanagi, H.; Yura, T.; Negishi, M.; Mori, K. ATF6 activated by proteolysis binds in the presence of NF-Y (CBF) directly to the cis-acting element responsible for the mammalian unfolded protein response. Mol. Cell. Biol. 2000, 20, 6755–6767. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Rutkowski, D.T.; Dubois, M.; Swathirajan, J.; Saunders, T.; Wang, J.; Song, B.; Yau, G.D.-Y.; Kaufman, R.J. ATF6Α optimizes long-term endoplasmic reticulum function to protect cells from chronic stress. Dev. Cell 2007, 13, 351–364. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.S. GRP78 induction in cancer: Therapeutic and prognostic implications. Cancer Res. 2007, 67, 3496–3499. [Google Scholar] [CrossRef] [PubMed]
- Scriven, P.; Coulson, S.; Haines, R.; Balasubramanian, S.; Cross, S.; Wyld, L. Activation and clinical significance of the unfolded protein response in breast cancer. Br. J. Cancer 2009, 101, 1692–1698. [Google Scholar] [CrossRef] [PubMed]
- Uramoto, H.; Sugio, K.; Oyama, T.; Nakata, S.; Ono, K.; Yoshimastu, T.; Morita, M.; Yasumoto, K. Expression of endoplasmic reticulum molecular chaperone GRP78 in human lung cancer and its clinical significance. Lung Cancer 2005, 49, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Jiang, Y.; Jia, Z.; Li, Q.; Gong, W.; Wang, L.; Wei, D.; Yao, J.; Fang, S.; Xie, K. Association of elevated GRP78 expression with increased lymph node metastasis and poor prognosis in patients with gastric cancer. Clin. Exp. Metast. 2006, 23, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Daneshmand, S.; Quek, M.L.; Lin, E.; Lee, C.; Cote, R.J.; Hawes, D.; Cai, J.; Groshen, S.; Lieskovsky, G.; Skinner, D.G. Glucose-regulated protein GRP78 is up-regulated in prostate cancer and correlates with recurrence and survival. Hum. Pathol. 2007, 38, 1547–1552. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.; Nichols, P.; Spicer, D.; Groshen, S.; Mimi, C.Y.; Lee, A.S. GRP78 as a novel predictor of responsiveness to chemotherapy in breast cancer. Cancer Res. 2006, 66, 7849–7853. [Google Scholar] [CrossRef] [PubMed]
- Abcouwer, S.F.; Marjon, P.L.; Loper, R.K.; Vander Jagt, D.L. Response of VEGF expression to amino acid deprivation and inducers of endoplasmic reticulum stress. Investig. Ophthalmol. Vis. Sci. 2002, 43, 2791–2798. [Google Scholar]
- Backer, J.M.; Krivoshein, A.V.; Hamby, C.V.; Pizzonia, J.; Gilbert, K.S.; Ray, Y.S.; Brand, H.; Paton, A.W.; Paton, J.C.; Backer, M.V. Chaperone-targeting cytotoxin and endoplasmic reticulum stress-inducing drug synergize to kill cancer cells. Neoplasia 2009, 11, 1165–1173. [Google Scholar] [CrossRef] [PubMed]
- Pyrko, P.; Schönthal, A.H.; Hofman, F.M.; Chen, T.C.; Lee, A.S. The unfolded protein response regulator GRP78/BiP as a novel target for increasing chemosensitivity in malignant gliomas. Cancer Res. 2007, 67, 9809–9816. [Google Scholar] [CrossRef] [PubMed]
- Firczuk, M.; Gabrysiak, M.; Barankiewicz, J.; Domagala, A.; Nowis, D.; Kujawa, M.; Jankowska-Steifer, E.; Wachowska, M.; Glodkowska-Mrowka, E.; Korsak, B. GRP78-targeting subtilase cytotoxin sensitizes cancer cells to photodynamic therapy. Cell Death Dis. 2013, 4, e741. [Google Scholar] [CrossRef] [PubMed]
- Wey, S.; Luo, B.; Tseng, C.-C.; Ni, M.; Zhou, H.; Fu, Y.; Bhojwani, D.; Carroll, W.L.; Lee, A.S. Inducible knockout of GRP78/BiP in the hematopoietic system suppresses pten-null leukemogenesis and AKT oncogenic signaling. Blood 2012, 119, 817–825. [Google Scholar] [CrossRef] [PubMed]
- Saito, S.; Furuno, A.; Sakurai, J.; Sakamoto, A.; Park, H.-R.; Shin-ya, K.; Tsuruo, T.; Tomida, A. Chemical genomics identifies the unfolded protein response as a target for selective cancer cell killing during glucose deprivation. Cancer Res. 2009, 69, 4225–4234. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Wey, S.; Wang, M.; Ye, R.; Liao, C.-P.; Roy-Burman, P.; Lee, A.S. Pten null prostate tumorigenesis and AKT activation are blocked by targeted knockout of er chaperone GRP78/BiP in prostate epithelium. Proc. Natl. Acad. Sci. USA 2008, 105, 19444–19449. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Z.; Harding, H.P.; Zhang, Y.; Jolicoeur, E.M.; Kuroda, M.; Ron, D. Cloning of mammalian Ire1 reveals diversity in the ER stress responses. EMBO J. 1998, 17, 5708–5717. [Google Scholar] [CrossRef] [PubMed]
- Tirasophon, W.; Welihinda, A.A.; Kaufman, R.J. A stress response pathway from the endoplasmic reticulum to the nucleus requires a novel bifunctional protein kinase/endoribonuclease (IRE1P) in mammalian cells. Genes Dev. 1998, 12, 1812–1824. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Adachi, M.; Zhao, S.; Hareyama, M.; Koong, A.; Luo, D.; Rando, T.; Imai, K.; Shinomura, Y. Preventing oxidative stress: A new role for XBP1. Cell Death Differ. 2009, 16, 847–857. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.-H.; Iwakoshi, N.N.; Glimcher, L.H. XBP-1 regulates a subset of endoplasmic reticulum resident chaperone genes in the unfolded protein response. Mol. Cell. Biol. 2003, 23, 7448–7459. [Google Scholar] [CrossRef] [PubMed]
- Koong, A.C.; Chauhan, V.; Romero-Ramirez, L. Targeting XBP-1 as a novel anti-cancer strategy. Cancer Biol. Ther. 2006, 5, 756–759. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Feldman, D.E.; Deng, C.; Brown, J.A.; De Giacomo, A.F.; Gaw, A.F.; Shi, G.; Le, Q.T.; Brown, J.M.; Koong, A.C. Identification of mitogen-activated protein kinase signaling pathways that confer resistance to endoplasmic reticulum stress in saccharomyces cerevisiae. Mol. Cancer Res. 2005, 3, 669–677. [Google Scholar] [CrossRef] [PubMed]
- Romero-Ramirez, L.; Cao, H.; Regalado, M.P.; Kambham, N.; Siemann, D.; Kim, J.J.; Le, Q.T.; Koong, A.C. X box-binding protein 1 regulates angiogenesis in human pancreatic adenocarcinomas. Transl. Oncol. 2009, 2, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Acosta-Alvear, D.; Zhou, Y.; Blais, A.; Tsikitis, M.; Lents, N.H.; Arias, C.; Lennon, C.J.; Kluger, Y.; Dynlacht, B.D. XBP1 controls diverse cell type-and condition-specific transcriptional regulatory networks. Mol. Cell 2007, 27, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, X.; Liang, Y.; Yu, H.; Chen, X.; Zheng, T.; Zheng, B.; Wang, L.; Zhao, L.; Shi, C. Targeting x box-binding protein-1 (XBP1) enhances sensitivity of glioma cells to oxidative stress. Neuropathol. Appl. Neurobiol. 2011, 37, 395–405. [Google Scholar] [CrossRef] [PubMed]
- Papandreou, I.; Denko, N.C.; Olson, M.; Van Melckebeke, H.; Lust, S.; Tam, A.; Solow-Cordero, D.E.; Bouley, D.M.; Offner, F.; Niwa, M. Identification of an ire1alpha endonuclease specific inhibitor with cytotoxic activity against human multiple myeloma. Blood 2011, 117, 1311–1314. [Google Scholar] [CrossRef] [PubMed]
- Marciniak, S.J.; Garcia-Bonilla, L.; Hu, J.; Harding, H.P.; Ron, D. Activation-dependent substrate recruitment by the eukaryotic translation initiation factor 2 kinase perk. J Cell Biol. 2006, 172, 201–209. [Google Scholar] [CrossRef] [PubMed]
- DuRose, J.B.; Scheuner, D.; Kaufman, R.J.; Rothblum, L.I.; Niwa, M. Phosphorylation of eukaryotic translation initiation factor 2α coordinates rrna transcription and translation inhibition during endoplasmic reticulum stress. Mol. Cell. Biol. 2009, 29, 4295–4307. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Vattem, K.M.; Sood, R.; An, J.; Liang, J.; Stramm, L.; Wek, R.C. Identification and characterization of pancreatic eukaryotic initiation factor 2 α-subunit kinase, PEK, involved in translational control. Mol. Cell. Biol. 1998, 18, 7499–7509. [Google Scholar] [CrossRef] [PubMed]
- Harding, H.P.; Zhang, Y.; Zeng, H.; Novoa, I.; Lu, P.D.; Calfon, M.; Sadri, N.; Yun, C.; Popko, B.; Paules, R. An integrated stress response regulates amino acid metabolism and resistance to oxidative stress. Mol. Cell 2003, 11, 619–633. [Google Scholar] [CrossRef]
- Verfaillie, T.; Garg, A.D.; Agostinis, P. Targeting er stress induced apoptosis and inflammation in cancer. Cancer Lett. 2013, 332, 249–264. [Google Scholar] [CrossRef] [PubMed]
- Harding, H.P.; Zhang, Y.; Bertolotti, A.; Zeng, H.; Ron, D. Perk is essential for translational regulation and cell survival during the unfolded protein response. Mol. Cell 2000, 5, 897–904. [Google Scholar] [CrossRef]
- Rouschop, K.M.; Dubois, L.J.; Keulers, T.G.; van den Beucken, T.; Lambin, P.; Bussink, J.; van der Kogel, A.J.; Koritzinsky, M.; Wouters, B.G. PERK/eIF2α signaling protects therapy resistant hypoxic cells through induction of glutathione synthesis and protection against ros. Proc. Natl. Acad. Sci. USA 2013, 110, 4622–4627. [Google Scholar] [CrossRef] [PubMed]
- Koumenis, C.; Naczki, C.; Koritzinsky, M.; Rastani, S.; Diehl, A.; Sonenberg, N.; Koromilas, A.; Wouters, B.G. Regulation of protein synthesis by hypoxia via activation of the endoplasmic reticulum kinase perk and phosphorylation of the translation initiation factor eIF2α. Mol. Cell. Biol. 2002, 22, 7405–7416. [Google Scholar] [CrossRef] [PubMed]
- Blais, J.D.; Addison, C.L.; Edge, R.; Falls, T.; Zhao, H.; Wary, K.; Koumenis, C.; Harding, H.P.; Ron, D.; Holcik, M. Perk-dependent translational regulation promotes tumor cell adaptation and angiogenesis in response to hypoxic stress. Mol. Cell. Biol. 2006, 26, 9517–9532. [Google Scholar] [CrossRef] [PubMed]
- Ferri, K.F.; Kroemer, G. Organelle-specific initiation of cell death pathways. Nat. Cell Biol. 2001, 3, E255–E263. [Google Scholar] [CrossRef] [PubMed]
- Tabas, I.; Ron, D. Integrating the mechanisms of apoptosis induced by endoplasmic reticulum stress. Nat. Cell Biol. 2011, 13, 184. [Google Scholar] [CrossRef] [PubMed]
- Cullinan, S.B.; Diehl, J.A. Coordination of er and oxidative stress signaling: The PERK/Nrf2 signaling pathway. Int. J. Biochem. Cell Biol. 2006, 38, 317–332. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.-W.; Zhu, H.-T.; Chen, K.-L.; Dong, X.; Wei, J.; Qiu, C.; Xue, J.-H. Protein kinase RNA-like endoplasmic reticulum kinase (PERK) signaling pathway plays a major role in reactive oxygen species (ROS)-mediated endoplasmic reticulum stress-induced apoptosis in diabetic cardiomyopathy. Cardiovasc. Diabetol. 2013, 12, 158. [Google Scholar] [CrossRef] [PubMed]
- Yokouchi, M.; Hiramatsu, N.; Hayakawa, K.; Okamura, M.; Du, S.; Kasai, A.; Takano, Y.; Shitamura, A.; Shimada, T.; Yao, J. Involvement of selective reactive oxygen species upstream of proapoptotic branches of unfolded protein response. J. Biol. Chem. 2008, 283, 4252–4260. [Google Scholar] [CrossRef] [PubMed]
- Oyadomari, S.; Mori, M. Roles of CHOP/GADD153 in endoplasmic reticulum stress. Cell Death Differ. 2004, 11, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Novoa, I.; Zeng, H.; Harding, H.P.; Ron, D. Feedback inhibition of the unfolded protein response by GADD34-mediated dephosphorylation of eIF2α. J. Cell Biol. 2001, 153, 1011–1022. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Mongillo, M.; Chin, K.-T.; Harding, H.; Ron, D.; Marks, A.R.; Tabas, I. Role of ero1-α–mediated stimulation of inositol 1, 4, 5-triphosphate receptor activity in endoplasmic reticulum stress–induced apoptosis. J. Cell Biol. 2009, 186, 783–792. [Google Scholar] [CrossRef] [PubMed]
- Ozcan, L.; Tabas, I. Pivotal Role of Calcium/Calmodulin-Dependent Protein Kinase II in ER Stress-Induced Apoptosis. Cell cycle 2010, 9, 223–224. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, H.; Wang, H.-G. Chop is involved in endoplasmic reticulum stress-induced apoptosis by enhancing DR5 expression in human carcinoma cells. J. Biol. Chem. 2004, 279, 45495–45502. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhu, H.; Xu, C.-J.; Yuan, J. Cleavage of bid by caspase 8 mediates the mitochondrial damage in the fas pathway of apoptosis. Cell 1998, 94, 491–501. [Google Scholar] [CrossRef]
- Puthalakath, H.; O’Reilly, L.A.; Gunn, P.; Lee, L.; Kelly, P.N.; Huntington, N.D.; Hughes, P.D.; Michalak, E.M.; McKimm-Breschkin, J.; Motoyama, N. ER stress triggers apoptosis by activating BH3-only protein Bim. Cell 2007, 129, 1337–1349. [Google Scholar] [CrossRef] [PubMed]
- Reimertz, C.; Kögel, D.; Rami, A.; Chittenden, T.; Prehn, J.H. Gene expression during ER stress–induced apoptosis in neurons: Induction of the BH3-only protein Bbc3/PUMA and activation of the mitochondrial apoptosis pathway. J.Cell Biol. 2003, 162, 587–597. [Google Scholar] [CrossRef] [PubMed]
- Huber, A.-L.; Lebeau, J.; Guillaumot, P.; Pétrilli, V.; Malek, M.; Chilloux, J.; Fauvet, F.; Payen, L.; Kfoury, A.; Renno, T. P58 IPK-mediated attenuation of the proapoptotic perk-chop pathway allows malignant progression upon low glucose. Mol. Cell 2013, 49, 1049–1059. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, H.; Umemura, A.; Taniguchi, K.; Font-Burgada, J.; Dhar, D.; Ogata, H.; Zhong, Z.; Valasek, M.A.; Seki, E.; Hidalgo, J. ER stress cooperates with hypernutrition to trigger TNF-dependent spontaneous hcc development. Cancer Cell 2014, 26, 331–343. [Google Scholar] [CrossRef] [PubMed]
- Matsukawa, J.; Matsuzawa, A.; Takeda, K.; Ichijo, H. The ASK1-MAP kinase cascades in mammalian stress response. J. Biochem. 2004, 136, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Fischer, H.; Koenig, U.; Eckhart, L.; Tschachler, E. Human caspase 12 has acquired deleterious mutations. Biochem. Biophy. Res. Commun. 2002, 293, 722–726. [Google Scholar] [CrossRef]
- Dhanasekaran, D.N.; Reddy, E.P. JNK signaling in apoptosis. Oncogene 2008, 27, 6245. [Google Scholar] [CrossRef] [PubMed]
- Yoneda, T.; Imaizumi, K.; Oono, K.; Yui, D.; Gomi, F.; Katayama, T.; Tohyama, M. Activation of caspase-12, an endoplastic reticulum (ER) resident caspase, through tumor necrosis factor receptor-associated factor 2-dependent mechanism in response to the er stress. J. Biol. Chem. 2001, 276, 13935–13940. [Google Scholar] [CrossRef] [PubMed]
- Nishitoh, H.; Matsuzawa, A.; Tobiume, K.; Saegusa, K.; Takeda, K.; Inoue, K.; Hori, S.; Kakizuka, A.; Ichijo, H. ASK1 is essential for endoplasmic reticulum stress-induced neuronal cell death triggered by expanded polyglutamine repeats. Genes Dev. 2002, 16, 1345–1355. [Google Scholar] [CrossRef] [PubMed]
- Harris, C.A.; Johnson, E.M. BH3-only Bcl-2 family members are coordinately regulated by the JNK pathway and require Bax to induce apoptosis in neurons. J. Biol. Chem. 2001, 276, 37754–37760. [Google Scholar] [PubMed]
- Yu, C.; Minemoto, Y.; Zhang, J.; Liu, J.; Tang, F.; Bui, T.N.; Xiang, J.; Lin, A. JNK suppresses apoptosis via phosphorylation of the proapoptotic Bcl-2 family protein bad. Mol. Cell 2004, 13, 329–340. [Google Scholar] [CrossRef]
- Urano, F.; Wang, X.; Bertolotti, A.; Zhang, Y.; Chung, P.; Harding, H.P.; Ron, D. Coupling of stress in the ER to activation of JNK protein kinases by transmembrane protein kinase IRE1. Science 2000, 287, 664–666. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ni, M.; Lee, B.; Barron, E.; Hinton, D.; Lee, A. The unfolded protein response regulator GRP78/BiP is required for endoplasmic reticulum integrity and stress-induced autophagy in mammalian cells. Cell Death Differ. 2008, 15, 1460–1471. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Guo, S.-H.; Shang, X.-J.; Yu, L.-S.; Zhu, J.-W.; Zhao, A.; Zhou, Y.-F.; An, G.-H.; Zhang, Q.; Ma, B. Triptolide induces sertoli cell apoptosis in mice via ROS/JNK-dependent activation of the mitochondrial pathway and inhibition of Nrf2-mediated antioxidant response. Acta Pharmacol. Sin. 2018, 39, 311–327. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, F. Reactive oxygen species (ROS), troublemakers between nuclear factor-κb (NF-κb) and c-Jun NH2-terminal kinase (JNK). Cancer Res. 2004, 64, 1902–1905. [Google Scholar] [CrossRef] [PubMed]
- Kowaltowski, A.J.; Naia-da-Silva, E.S.; Castilho, R.F.; Vercesi, A.E. Ca2+-stimulated mitochondrial reactive oxygen species generation and permeability transition are inhibited by dibucaine or Mg2+. Arch. Biochem. Biophys. 1998, 359, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Dourdin, N.; Wu, C.; De Veyra, T.; Elce, J.S.; Greer, P.A. Ubiquitous calpains promote caspase-12 and JNK activation during endoplasmic reticulum stress-induced apoptosis. J. Biol. Chem. 2006, 281, 16016–16024. [Google Scholar] [CrossRef] [PubMed]
- Duchen, M.R. Contributions of mitochondria to animal physiology: From homeostatic sensor to calcium signalling and cell death. J. Physiol. 1999, 516, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Kuwana, T.; Bouchier-Hayes, L.; Chipuk, J.E.; Bonzon, C.; Sullivan, B.A.; Green, D.R.; Newmeyer, D.D. BH3 domains of BH3-only proteins differentially regulate Bax-mediated mitochondrial membrane permeabilization both directly and indirectly. Mol. Cell 2005, 17, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Hao, Z.; Duncan, G.S.; Chang, C.-C.; Elia, A.; Fang, M.; Wakeham, A.; Okada, H.; Calzascia, T.; Jang, Y.; You-Ten, A. Specific ablation of the apoptotic functions of cytochrome C reveals a differential requirement for cytochrome C and Apaf-1 in apoptosis. Cell 2005, 121, 579–591. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Nijhawan, D.; Budihardjo, I.; Srinivasula, S.M.; Ahmad, M.; Alnemri, E.S.; Wang, X. Cytochrome c and datp-dependent formation of Apaf-1/caspase-9 complex initiates an apoptotic protease cascade. Cell 1997, 91, 479–489. [Google Scholar] [CrossRef]
- Li, L.Y.; Luo, X.; Wang, X. Endonuclease G is an apoptotic dnase when released from mitochondria. Nature 2001, 412, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Bobrovnikova-Marjon, E.; Grigoriadou, C.; Pytel, D.; Zhang, F.; Ye, J.; Koumenis, C.; Cavener, D.; Diehl, J.A. PERK promotes cancer cell proliferation and tumor growth by limiting oxidative DNA damage. Oncogene 2010, 29, 3881–3895. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Xu, Y.; Huang, X.; Chen, Y.; Fu, J.; Xi, M.; Wang, L. Berberine-induced apoptosis in human breast cancer cells is mediated by reactive oxygen species generation and mitochondrial-related apoptotic pathway. Tumor Biol. 2015, 36, 1279–1288. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.-C.; Chen, J.-Y. Proteomic analysis reveals that pardaxin triggers apoptotic signaling pathways in human cervical carcinoma hela cells: Cross talk among the UPR, c-Jun and ROS. Carcinogenesis 2013, 34, 1833–1842. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Rodríguez, A.G.; Juárez-Portilla, C.; Olivares-Bañuelos, T.; Zepeda, R.C. Anticancer activity of seaweeds. Drug Discov. Today 2018, 23, 434–437. [Google Scholar] [CrossRef] [PubMed]
- Cragg, G.M.; Grothaus, P.G.; Newman, D.J. Impact of natural products on developing new anti-cancer agents. Chem. Rev. 2009, 109, 3012–3043. [Google Scholar] [CrossRef] [PubMed]
- Kelloff, G.J.; Crowell, J.A.; Steele, V.E.; Lubet, R.A.; Malone, W.A.; Boone, C.W.; Kopelovich, L.; Hawk, E.T.; Lieberman, R.; Lawrence, J.A. Progress in cancer chemoprevention: Development of diet-derived chemopreventive agents. J. Nutr. 2000, 130, 467S–471S. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Stutzman, J.D.; Kelloff, G.J.; Steele, V.E. Screening of potential chemopreventive agents using biochemical markers of carcinogenesis. Cancer Res. 1994, 54, 5848–5855. [Google Scholar] [PubMed]
- Mann, J. Natural products in cancer chemotherapy: Past, present and future. Nat. Rev. Cancer. 2002, 2, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.; Song, H.S.; Park, H.; Kim, B. Activation of ER stress-dependent miR-216b has a critical role in salviamiltiorrhiza ethanol-extract-induced apoptosis in U266 and U937 cells. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Cha, J.A.; Song, H.-S.; Kang, B.; Park, M.N.; Park, K.S.; Kim, S.-H.; Shim, B.-S.; Kim, B. Mir-211 plays a critical role in cnidium officinale makino extract-induced, ROS/ER stress-mediated apoptosis in U937 and U266 cells. Int. J. Mol. Sci. 2018, 19, 865. [Google Scholar] [CrossRef] [PubMed]
- Park, S.K.; Sanders, B.G.; Kline, K. Tocotrienols induce apoptosis in breast cancer cell lines via an endoplasmic reticulum stress-dependent increase in extrinsic death receptor signaling. Breast Cancer Res. Treat. 2010, 124, 361–375. [Google Scholar] [CrossRef] [PubMed]
- Al Husaini, H.; Wheatley-Price, P.; Clemons, M.; Shepherd, F.A. Prevention and management of bone metastases in lung cancer: A review. J. Thorac. Oncol. 2009, 4, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Qazi, A.K.; Siddiqui, J.A.; Jahan, R.; Chaudhary, S.; Walker, L.A.; Sayed, Z.; Jones, D.T.; Batra, S.K.; Macha, M.A. Emerging therapeutic potential of graviola and its constituents in cancers. Carcinogenesis 2018, 39, 522–533. [Google Scholar] [CrossRef] [PubMed]
- Marciniak, S.J. Endoplasmic reticulum stress in lung disease. Eur. Respir. Rev. 2017, 26. [Google Scholar] [CrossRef] [PubMed]
- Siu, F.M.; Ma, D.L.; Cheung, Y.W.; Lok, C.N.; Yan, K.; Yang, Z.; Yang, M.; Xu, S.; Ko, B.C.; He, Q.Y.; et al. Proteomic and transcriptomic study on the action of a cytotoxic saponin (polyphyllin D): Induction of endoplasmic reticulum stress and mitochondria-mediated apoptotic pathways. Proteomics 2008, 8, 3105–3117. [Google Scholar] [CrossRef] [PubMed]
- Hung, J.Y.; Hsu, Y.L.; Ni, W.C.; Tsai, Y.M.; Yang, C.J.; Kuo, P.L.; Huang, M.S. Oxidative and endoplasmic reticulum stress signaling are involved in dehydrocostuslactone-mediated apoptosis in human non-small cell lung cancer cells. Lung Cancer 2010, 68, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.L.; Kim, S.Y.; Shin, S.W.; Park, J.-W. Regulation of brefeldin A-induced ER stress and apoptosis by mitochondrial NADP+-dependent isocitrate dehydrogenase. Biochem. Biophys. Res. Commun. 2012, 417, 760–764. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.M.; Kim, B.M.; Park, J.B. Omega-hydroxyundec-9-enoic acid induces apoptosis through ROS-mediated endoplasmic reticulum stress in non-small cell lung cancer cells. Biochem. Biophys. Res. Commun. 2014, 448, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.-H.; Hang, L.-W.; Yang, J.-S.; Chen, H.-Y.; Lin, H.-Y.; Chiang, J.-H.; Lu, C.-C.; Yang, J.-L.; Lai, T.-Y.; Ko, Y.-C. Curcumin induces apoptosis in human non-small cell lung cancer NCI-h460 cells through er stress and caspase cascade-and mitochondria-dependent pathways. Anticancer Res. 2010, 30, 2125–2133. [Google Scholar] [PubMed]
- Hsia, T.C.; Yu, C.C.; Hsu, S.C.; Tang, N.Y.; Lu, H.F.; Huang, Y.P.; Wu, S.H.; Lin, J.G.; Chung, J.G. Cantharidin induces apoptosis of h460 human lung cancer cells through mitochondria-dependent pathways. Int. J. Oncol. 2014, 45, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.S.; Dang, Y.Y.; Guo, J.J.; Wu, G.S.; Lu, J.J.; Chen, X.P.; Wang, Y.T. Furanodiene induces endoplasmic reticulum stress and presents antiproliferative activities in lung cancer cells. Evid.-Based Complement. Altern. Med. ECAM. 2012, 2012, 426521. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Liu, X.; Su, L. Parthenolide induces apoptosis via tnfrsf10b and pmaip1 pathways in human lung cancer cells. J. Exp. Clin. Cancer Res. 2014, 33, 3. [Google Scholar] [CrossRef] [PubMed]
- Seong, Y.A.; Shin, P.G.; Yoon, J.S.; Yadunandam, A.K.; Kim, G.D. Induction of the endoplasmic reticulum stress and autophagy in human lung carcinoma a549 cells by anacardic acid. Cell Biochem. Biophys. 2014, 68, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Clarke, R.; Cook, K.L.; Hu, R.; Facey, C.O.; Tavassoly, I.; Schwartz, J.L.; Baumann, W.T.; Tyson, J.J.; Xuan, J.; Wang, Y.; et al. Endoplasmic reticulum stress, the unfolded protein response, autophagy, and the integrated regulation of breast cancer cell fate. Cancer Res. 2012, 72, 1321–1331. [Google Scholar] [CrossRef] [PubMed]
- Ko, E.-Y.; Moon, A. Natural products for chemoprevention of breast cancer. J. Cancer Prev. 2015, 20, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Tseng, C.N.; Hong, Y.R.; Chang, H.W.; Yu, T.J.; Hung, T.W.; Hou, M.F.; Yuan, S.S.; Cho, C.L.; Liu, C.T.; Chiu, C.C.; et al. Brefeldin a reduces anchorage-independent survival, cancer stem cell potential and migration of MDA-MB-231 human breast cancer cells. Molecules 2014, 19, 17464–17477. [Google Scholar] [CrossRef] [PubMed]
- Park, I.J.; Kim, M.J.; Park, O.J.; Choe, W.; Kang, I.; Kim, S.S.; Ha, J. Cryptotanshinone induces ER stress-mediated apoptosis in HEPG2 and MCF7 cells. Apoptosis 2012, 17, 248–257. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.M.; Bai, L.L.; Zhang, D.M.; Yiu, A.; Yin, Z.Q.; Han, W.L.; Liu, J.S.; Li, Y.; Fu, D.Y.; Ye, W.C. Saxifragifolin D induces the interplay between apoptosis and autophagy in breast cancer cells through ROS-dependent endoplasmic reticulum stress. Biochem. Pharmacol. 2013, 85, 913–926. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.Y.; Shen, Y.C.; Lu, C.H.; Yang, S.Y.; Ho, T.F.; Peng, Y.T.; Chang, C.C. Prodigiosin activates endoplasmic reticulum stress cell death pathway in human breast carcinoma cell lines. Toxicol. Appl. Pharmacol. 2012, 265, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhao, Y.; Zhang, Y.; Zhang, D. Fucoidan induces cancer cell apoptosis by modulating the endoplasmic reticulum stress cascades. PLoS ONE 2014, 9, e108157. [Google Scholar] [CrossRef] [PubMed]
- Kamiya, T.; Nishihara, H.; Hara, H.; Adachi, T. Ethanol extract of brazilian red propolis induces apoptosis in human breast cancer MCF-7 cells through endoplasmic reticulum stress. J. Agric. Food Chem. 2012, 60, 11065–11070. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Shu, F.; Liang, X.; Chang, H.; Shi, L.; Peng, X.; Zhu, J.; Mi, M. Ampelopsin induces cell growth inhibition and apoptosis in breast cancer cells through ROS generation and endoplasmic reticulum stress pathway. PLoS ONE 2014, 9, e89021. [Google Scholar] [CrossRef] [PubMed]
- Qazi, A.K.; Hussain, A.; Aga, M.A.; Ali, S.; Taneja, S.C.; Sharma, P.R.; Saxena, A.K.; Mondhe, D.M.; Hamid, A. Cell specific apoptosis by RLX is mediated by NFκB in human colon carcinoma HCT-116 cells. BMC Cell Biol. 2014, 15, 36. [Google Scholar] [CrossRef] [PubMed]
- Wielenga, M.C.B.; Colak, S.; Heijmans, J.; van Lidth de Jeude, J.F.; Rodermond, H.M.; Paton, J.C.; Paton, A.W.; Vermeulen, L.; Medema, J.P.; van den Brink, G.R. ER-stress-induced differentiation sensitizes colon cancer stem cells to chemotherapy. Cell Rep. 2015, 13, 489–494. [Google Scholar] [CrossRef] [PubMed]
- Cao, A.; Li, Q.; Yin, P.; Dong, Y.; Shi, H.; Wang, L.; Ji, G.; Xie, J.; Wu, D. Curcumin induces apoptosis in human gastric carcinoma ags cells and colon carcinoma HT-29 cells through mitochondrial dysfunction and endoplasmic reticulum stress. Apoptosis 2013, 18, 1391–1402. [Google Scholar] [CrossRef] [PubMed]
- Guichard, C.; Pedruzzi, E.; Fay, M.; Marie, J.C.; Braut-Boucher, F.; Daniel, F.; Grodet, A.; Gougerot-Pocidalo, M.A.; Chastre, E.; Kotelevets, L.; et al. Dihydroxyphenylethanol induces apoptosis by activating serine/threonine protein phosphatase PP2A and promotes the endoplasmic reticulum stress response in human colon carcinoma cells. Carcinogenesis 2006, 27, 1812–1827. [Google Scholar] [CrossRef] [PubMed]
- Tseng, C.N.; Huang, C.F.; Cho, C.L.; Chang, H.W.; Huang, C.W.; Chiu, C.C.; Chang, Y.F. Brefeldin a effectively inhibits cancer stem cell-like properties and mmp-9 activity in human colorectal cancer colo 205 cells. Molecules 2013, 18, 10242–10253. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-W.; Woo, K.J.; Lee, J.-T.; Lim, J.H.; Lee, T.-J.; Kim, S.H.; Choi, Y.H.; Kwon, T.K. Resveratrol induces pro-apoptotic endoplasmic reticulum stress in human colon cancer cells. Oncol. Rep. 2007, 18, 1269–1273. [Google Scholar] [CrossRef] [PubMed]
- Edagawa, M.; Kawauchi, J.; Hirata, M.; Goshima, H.; Inoue, M.; Okamoto, T.; Murakami, A.; Maehara, Y.; Kitajima, S. Role of activating transcription factor 3 (ATF3) in endoplasmic reticulum (ER) stress-induced sensitization of p53-deficient human colon cancer cells to tumor necrosis factor (TNF)-related apoptosis-inducing ligand (TRAIL)-mediated apoptosis through up-regulation of death receptor 5 (DR5) by zerumbone and celecoxib. J. Biol. Chem. 2014, 289, 21544–21561. [Google Scholar] [PubMed]
- Protiva, P.; Hopkins, M.E.; Baggett, S.; Yang, H.; Lipkin, M.; Holt, P.R.; Kennelly, E.J.; Bernard, W.I. Growth inhibition of colon cancer cells by polyisoprenylated benzophenones is associated with induction of the endoplasmic reticulum response. Int. J. Cancer 2008, 123, 687–694. [Google Scholar] [CrossRef] [PubMed]
- Yaffe, P.B.; Power Coombs, M.R.; Doucette, C.D.; Walsh, M.; Hoskin, D.W. Piperine, an alkaloid from black pepper, inhibits growth of human colon cancer cells via G1 arrest and apoptosis triggered by endoplasmic reticulum stress. Mol. Carcinogenesis 2015, 54, 1070–1085. [Google Scholar] [CrossRef] [PubMed]
- Kuo, Y.F.; Su, Y.Z.; Tseng, Y.H.; Wang, S.Y.; Wang, H.M.; Chueh, P.J. Flavokawain B, a novel chalcone from alpinia pricei Hayata with potent apoptotic activity: Involvement of ros and GADD153 upstream of mitochondria-dependent apoptosis in HCT116 cells. Free Radic. Biol. Med. 2010, 49, 214–226. [Google Scholar] [CrossRef] [PubMed]
- Mullen, J.T.; Ryan, D.P. Neoadjuvant chemotherapy for gastric cancer: What are we trying to accomplish? Ann. Surg. Oncol. 2014, 21, 13–15. [Google Scholar] [CrossRef] [PubMed]
- Boelens, J.; Lust, S.; Offner, F.; Bracke, M.E.; Vanhoecke, B.W. The endoplasmic reticulum: A target for new anticancer drugs. In Vivo 2007, 21, 215–226. [Google Scholar] [PubMed]
- Ahn, J.; Lee, J.S.; Yang, K.M. Ultrafine particles of Ulmus davidiana var. japonica induce apoptosis of gastric cancer cells via activation of caspase and endoplasmic reticulum stress. Arch. Pharm. Res. 2014, 37, 783–792. [Google Scholar] [CrossRef] [PubMed]
- Sheu, M.L.; Liu, S.H.; Lan, K.H. Honokiol induces calpain-mediated glucose-regulated protein-94 cleavage and apoptosis in human gastric cancer cells and reduces tumor growth. PLoS ONE 2007, 2, e1096. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Zhang, Z.; Jia, L.; Zhao, Y.; Zhang, X.; Wu, K. Endoplasmic reticulum stress contributes to vitamin E succinate-induced apoptosis in human gastric cancer SGC-7901 cells. Cancer Lett. 2010, 296, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Li, L.; Zhang, L.; Zhang, Z.; Wang, X.; Zhang, X.; Hou, L.; Wu, K. Crosstalk between endoplasmic reticulum stress and oxidative stress in apoptosis induced by alpha-tocopheryl succinate in human gastric carcinoma cells. Br. J. Nutr. 2013, 109, 727–735. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Tian, L.; Long, L.; Quan, M.; Liu, F.; Cao, J. Casticin potentiates trail-induced apoptosis of gastric cancer cells through endoplasmic reticulum stress. PLoS ONE 2013, 8, e58855. [Google Scholar] [CrossRef] [PubMed]
- Zou, P.; Zhang, J.; Xia, Y.; Kanchana, K.; Guo, G.; Chen, W.; Huang, Y.; Wang, Z.; Yang, S.; Liang, G. ROS generation mediates the anti-cancer effects of WZ35 via activating JNK and ER stress apoptotic pathways in gastric cancer. Oncotarget 2015, 6, 5860. [Google Scholar] [CrossRef] [PubMed]
- Kallifatidis, G.; Hoy, J.J.; Lokeshwar, B.L. Bioactive Natural products for chemoprevention and treatment of castration-resistant prostate cancer. Semin. Cancer Biol. 2016, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Miyake, H.; Hara, I.; Arakawa, S.; Kamidono, S. Stress protein GRP78 prevents apoptosis induced by calcium ionophore, ionomycin, but not by glycosylation inhibitor, tunicamycin, in human prostate cancer cells. J. Cell. Biochem. 2000, 77, 396–408. [Google Scholar] [CrossRef]
- Yu, C.C.; Wu, P.J.; Hsu, J.L.; Ho, Y.F.; Hsu, L.C.; Chang, Y.J.; Chang, H.S.; Chen, I.S.; Guh, J.H. Ardisianone, a natural benzoquinone, efficiently induces apoptosis in human hormone-refractory prostate cancers through mitochondrial damage stress and survivin downregulation. Prostate 2013, 73, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Rizzi, F.; Naponelli, V.; Silva, A.; Modernelli, A.; Ramazzina, I.; Bonacini, M.; Tardito, S.; Gatti, R.; Uggeri, J.; Bettuzzi, S. Polyphenon E(R), a standardized green tea extract, induces endoplasmic reticulum stress, leading to death of immortalized pnt1a cells by anoikis and tumorigenic PC3 by necroptosis. Carcinogenesis 2014, 35, 828–839. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Petiwala, S.M.; Pierce, D.R.; Nonn, L.; Johnson, J.J. Selective modulation of endoplasmic reticulum stress markers in prostate cancer cells by a standardized mangosteen fruit extract. PLoS ONE 2013, 8, e81572. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Sun, J.; Xu, Q.; Liu, Y.; Wei, J.; Young, C.Y.; Yuan, H.; Lou, H. Marchantin M: A novel inhibitor of proteasome induces autophagic cell death in prostate cancer cells. Cell Death Dis. 2013, 4, e761. [Google Scholar] [CrossRef] [PubMed]
- Chiu, H.W.; Fang, W.H.; Chen, Y.L.; Wu, M.D.; Yuan, G.F.; Ho, S.Y.; Wang, Y.J. Monascuspiloin enhances the radiation sensitivity of human prostate cancer cells by stimulating endoplasmic reticulum stress and inducing autophagy. PLoS ONE 2012, 7, e40462. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.C.; Yen, C.Y.; Wu, R.S.; Yang, J.S.; Lu, H.F.; Lu, K.W.; Lo, C.; Chen, H.Y.; Tang, N.Y.; Wu, C.C.; et al. The roles of endoplasmic reticulum stress and mitochondrial apoptotic signaling pathway in quercetin-mediated cell death of human prostate cancer PC-3 cells. Environ. Toxicol. 2014, 29, 428–439. [Google Scholar] [CrossRef] [PubMed]
- Chan, M.L.; Liang, J.W.; Hsu, L.C.; Chang, W.L.; Lee, S.S.; Guh, J.H. Zerumbone, a ginger sesquiterpene, induces apoptosis and autophagy in human hormone-refractory prostate cancers through tubulin binding and crosstalk between endoplasmic reticulum stress and mitochondrial insult. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2015, 388, 1223–1236. [Google Scholar] [CrossRef] [PubMed]
- Spangenberg, H.C.; Thimme, R.; Blum, H.E. Targeted therapy for hepatocellular carcinoma. Nature Reviews Gastroenterol. Hepatol. 2009, 6, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Carr, B.I. Hepatocellular carcinoma: Current management and future trends. Gastroenterology 2004, 127, S218–S224. [Google Scholar] [CrossRef] [PubMed]
- Malhi, H.; Kaufman, R.J. Endoplasmic reticulum stress in liver disease. J. Hepatol. 2011, 54, 795–809. [Google Scholar] [CrossRef] [PubMed]
- Choi, A.Y.; Choi, J.H.; Hwang, K.Y.; Jeong, Y.J.; Choe, W.; Yoon, K.S.; Ha, J.; Kim, S.S.; Youn, J.H.; Yeo, E.J.; et al. Licochalcone a induces apoptosis through endoplasmic reticulum stress via a phospholipase cgamma1-, Ca(2+)-, and reactive oxygen species-dependent pathway in HepG2 human hepatocellular carcinoma cells. Apoptosis 2014, 19, 682–697. [Google Scholar] [CrossRef] [PubMed]
- Moon, D.O.; Park, S.Y.; Choi, Y.H.; Ahn, J.S.; Kim, G.Y. Guggulsterone sensitizes hepatoma cells to trail-induced apoptosis through the induction of chop-dependent DR5: Involvement of ros-dependent er-stress. Biochem. Pharmacol. 2011, 82, 1641–1650. [Google Scholar] [CrossRef] [PubMed]
- Moon, D.O.; Asami, Y.; Long, H.; Jang, J.H.; Bae, E.Y.; Kim, B.Y.; Choi, Y.H.; Kang, C.H.; Ahn, J.S.; Kim, G.Y. Verrucarin a sensitizes trail-induced apoptosis via the upregulation of DR5 in an eIF2alpha/CHOP-dependent manner. Toxicol. In Vitro 2013, 27, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.F.; Cao, J.G.; Tian, L.; Liu, F. 5, 7-dimethoxyflavone sensitizes trail-induced apoptosis through DR5 upregulation in hepatocellular carcinoma cells. Cancer Chemother. Pharmacol. 2012, 69, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.S.; Kim, H.M.; Yadunandam, A.K.; Kim, N.H.; Jung, H.A.; Choi, J.S.; Kim, C.Y.; Kim, G.D. Neferine isolated from nelumbo nucifera enhances anti-cancer activities in Hep3B cells: Molecular mechanisms of cell cycle arrest, ER stress induced apoptosis and anti-angiogenic response. Phytomedicine 2013, 20, 1013–1022. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Song, B.; Sun, G.; Ma, T.; Zhong, F.; Wei, W. Endoplasmic reticulum stress–induced resistance to doxorubicin is reversed by paeonol treatment in human hepatocellular carcinoma cells. PLoS ONE 2013, 8, e62627. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.; Zhou, P.; Peng, Y.-B.; Xu, X.; Ma, J.; Liu, Q.; Zhang, L.; Wen, X.-D.; Qi, L.-W.; Gao, N. 6-shogaol induces apoptosis in human hepatocellular carcinoma cells and exhibits anti-tumor activity in vivo through endoplasmic reticulum stress. PLoS ONE 2012, 7, e39664. [Google Scholar] [CrossRef] [PubMed]
- Yeh, T.C.; Chiang, P.C.; Li, T.K.; Hsu, J.L.; Lin, C.J.; Wang, S.W.; Peng, C.Y.; Guh, J.H. Genistein induces apoptosis in human hepatocellular carcinomas via interaction of endoplasmic reticulum stress and mitochondrial insult. Biochem. Pharmacol. 2007, 73, 782–792. [Google Scholar] [CrossRef] [PubMed]
- Richardson, J.L.; Marks, G.; Levine, A. The influence of symptoms of disease and side effects of treatment on compliance with cancer therapy. J. Clin. Oncol. 1988, 6, 1746–1752. [Google Scholar] [CrossRef] [PubMed]
- Coates, A.; Abraham, S.; Kaye, S.B.; Sowerbutts, T.; Frewin, C.; Fox, R.; Tattersall, M. On the receiving end—Patient perception of the side-effects of cancer chemotherapy. Eur. J. Cancer Clin. Oncol. 1983, 19, 203–208. [Google Scholar] [CrossRef]
- Tsuruo, T.; Naito, M.; Tomida, A.; Fujita, N.; Mashima, T.; Sakamoto, H.; Haga, N. Molecular targeting therapy of cancer: Drug resistance, apoptosis and survival signal. Cancer Sci. 2003, 94, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Shuda, M.; Kondoh, N.; Imazeki, N.; Tanaka, K.; Okada, T.; Mori, K.; Hada, A.; Arai, M.; Wakatsuki, T.; Matsubara, O. Activation of the ATF6, XBP1 and GRP78 genes in human hepatocellular carcinoma: A possible involvement of the ER stress pathway in hepatocarcinogenesis. J. Hepatol. 2003, 38, 605–614. [Google Scholar] [CrossRef]
- Corazzari, M.; Lovat, P.E.; Armstrong, J.L.; Fimia, G.M.; Hill, D.S.; Birch-Machin, M.; Redfern, C.P.F.; Piacentini, M. Targeting homeostatic mechanisms of endoplasmic reticulum stress to increase susceptibility of cancer cells to fenretinide-induced apoptosis: The role of stress proteins ERDJ5 and ERP57. Br. J. Cancer. 2007, 96, 1062–1071. [Google Scholar] [CrossRef] [PubMed]
- Gray, M.D.; Mann, M.; Nitiss, J.L.; Hendershot, L.M. Activation of the unfolded protein response is necessary and sufficient for reducing topoisomerase IIΑ protein levels and decreasing sensitivity to topoisomerase-targeted drugs. Mol. Pharmacol. 2005, 68, 1699–1707. [Google Scholar] [PubMed]
- Malhotra, J.D.; Kaufman, R.J. Endoplasmic reticulum stress and oxidative stress: A vicious cycle or a double-edged sword? Antioxid. Redox Signal. 2007, 9, 2277–2294. [Google Scholar] [CrossRef] [PubMed]
- Da Rocha, A.B.; Lopes, R.M.; Schwartsmann, G. Natural products in anticancer therapy. Curr. Opin. Pharmacol. 2001, 1, 364–369. [Google Scholar] [CrossRef]
- Gordaliza, M. Natural products as leads to anticancer drugs. Clin. Transl. Oncol. 2007, 9, 767–776. [Google Scholar] [CrossRef] [PubMed]
- López-Antón, N.; Rudy, A.; Barth, N.; Schmitz, L.M.; Pettit, G.R.; Schulze-Osthoff, K.; Dirsch, V.M.; Vollmar, A.M. The marine product cephalostatin 1 activates an endoplasmic reticulum stress-specific and apoptosome-independent apoptotic signaling pathway. J. Biol. Chem. 2006, 281, 33078–33086. [Google Scholar] [CrossRef] [PubMed]
- Law, B.Y.; Wang, M.; Ma, D.-L.; Al-Mousa, F.; Michelangeli, F.; Cheng, S.-H.; Ng, M.H.; To, K.-F.; Mok, A.Y.; Ko, R.Y. Alisol B, a novel inhibitor of the sarcoplasmic/endoplasmic reticulum Ca2+ atpase pump, induces autophagy, endoplasmic reticulum stress, and apoptosis. Mol. Cancer Ther. 2010, 9, 718–730. [Google Scholar] [CrossRef] [PubMed]
- Carew, J.S.; Nawrocki, S.T.; Krupnik, Y.V.; Dunner, K.; McConkey, D.J.; Keating, M.J.; Huang, P. Targeting endoplasmic reticulum protein transport: A novel strategy to kill malignant B cells and overcome fludarabine resistance in CLL. Blood 2006, 107, 222–231. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.C.; Wang, W.; Golden, E.B.; Thomas, S.; Sivakumar, W.; Hofman, F.M.; Louie, S.G.; Schonthal, A.H. Green tea epigallocatechin gallate enhances therapeutic efficacy of temozolomide in orthotopic mouse glioblastoma models. Cancer Lett. 2011, 302, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.; Gu, M.; Zhao, Y.; Zheng, X.; Xing, C. Autophagy contributes to falcarindiol-induced cell death in breast cancer cells with enhanced endoplasmic reticulum stress. PLoS ONE 2017, 12, e0176348. [Google Scholar] [CrossRef] [PubMed]
- Dolan, L.C.; Matulka, R.A.; Burdock, G.A. Naturally occurring food toxins. Toxins 2010, 2, 2289–2332. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.Y.; Abdul, A.; Ibrahim, T.A.T.; Abdelwahab, S.I.; Elhassan, M.M.; Syam, M. Evaluation of acute toxicity and the effect of single injected doses of zerumbone on the kidney and liver functions in sprague dawley rats. Afr. J. Biotechnol. 2010, 9, 4442–4450. [Google Scholar]
- Chang, P.Y.; Mirsalis, J.; Riccio, E.S.; Bakke, J.P.; Lee, P.S.; Shimon, J.; Phillips, S.; Fairchild, D.; Hara, Y.; Crowell, J.A. Genotoxicity and toxicity of the potential cancer-preventive agent polyphenon e. Environ. Mol. Mutagenesis 2003, 41, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, H. ER stress and diseases. FEBS J. 2007, 274, 630–658. [Google Scholar] [CrossRef] [PubMed]
- Salminen, A.; Kauppinen, A.; Suuronen, T.; Kaarniranta, K.; Ojala, J. ER stress in Alzheimer’s disease: A novel neuronal trigger for inflammation and alzheimer’s pathology. J. Neuroinflamm. 2009, 6, 41. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Family Name | Compound | Cell Line | Duration/Dosage | Mechanism | Reference |
|---|---|---|---|---|---|
| Paris polyphylla | Polyphyllin D | NCI-H460 | 1.8 µM, 8 h | CHOP ↑, GRP78/BiP ↑, PDI ↑ | [105] |
| cleavage of caspase-3,-4,-9,-12 ↑, Bax ↑, Bcl-2 ↓ | |||||
| Saussurea lappa and Aucklandia lappa | Dehydrocostuslactone | NCI-H460 | 15 μg/mL, 1 h | release of intracellular Ca2+ levels ↑, Δψm ↓, p-PERK ↑, GRP78/BiP ↑, IRE1 ↑, CHOP ↑, XBP-1 ↑ | [106] |
| Cleavage of caspase-4 ↑, JNK ↑, MAPK ↑ | |||||
| ROS ↑ | |||||
| A549 | 15 μg/mL, 1 h | release of intracellular Ca2+ ↑, Δψm ↓,p-PERK ↑, GRP78/BiP ↑, IRE1 ↑, CHOP ↑, XBP1 ↑ | |||
| JNK ↑, MAPK ↑, cleavage of caspase-4 ↑ | |||||
| ROS ↑ | |||||
| BrefeldinA | A549 | 1 µM, 36 h | p-PERK ↑, IRE1α ↑, ATF4 ↑, ATF6 ↑, CHOP ↑, GRP78/BiP ↑ | [107] | |
| cleavage of PARP ↑, cleavage of caspase-2 ↑, Bid ↑ | |||||
| Oryza officinalis | ω-Hydroxyundec-9-enoic acid | H1299 | 500 µM, 24 h | p-eIF2α ↑, CHOP ↑ | [108] |
| cleavage of caspase-6,-9 ↑, cleavage of PARP ↑ | |||||
| ROS ↑ | |||||
| Curcuma longa | Curcumin | NCI-H460 | 25 µM, 24 h | release of intracellular Ca2+ ↑, Δψm ↓, CHOP ↑, GRP78/BiP ↑ | [109] |
| Bcl-2 ↓, Bcl-xL ↓, cytochrome c ↑, cleavage of caspase-3,-8,-9 ↑, BAX ↑, BAD ↑ | |||||
| ROS ↑ | |||||
| Mylabris phalerata Pallas | Cantharidin | H460 | 10 µM, 24 h | release of intracellular Ca2+ ↑, Δψm ↓, GRP78/BiP ↑, IRE1α ↑, IRE1β ↑, ATF6α ↑, XBP1 ↑, calpain ↑, | [110] |
| Bcl-xL ↓, cleavage of caspase-3,-8,-4 ↑ cytochrome c ↑ BAX ↑, AIF ↑, Endo G ↑ | |||||
| ROS ↑ | |||||
| Curcumae Rhizoma | Furanodiene | A549, 95-D | 80 µM, 24 h | CHOP ↑, BIP ↑ | [111] |
| Tanacetum parthenium L. | Parthenolide | A549, Calu-1, H1299, H1792 | 20 µM, 24 h | ATF4 ↑, p-eIF2a ↑, eIF2α ↑ | [112] |
| cleavage of caspase-3,-8,-9 ↑, cleavage of PARP ↑ | |||||
| Anacardium occidentale | Anacardic acid | A549 | 3.0 μg/mL, 18 h | release of intracellular Ca2+ ↑, Δψm ↓, GRP78/BiP ↑, CHOP ↑, IRE1α ↑ ATF6 ↑, p-PERK ↓, p-eIF2α ↓ | [113] |
| cleavage caspase-12 ↑ |
| Family Name | Compound | Cell Line | Duration/Dosage | Mechanism | Reference |
|---|---|---|---|---|---|
| Eupenicillium brefeldianum | Brefeldin A | MDA-MB-231 | 1 μg/mL, 24 h | IRE1α ↑, PERK ↑, CHOP ↑, calnexin ↓ | [116] |
| BIM ↑, cleavage of PARP ↑, Bcl-2 ↓, | |||||
| Salvia miltiorrhiza Bunge | Cryptotanshinone | MCF7 | 10 µM, 24 h | p-eIF2α ↑, GRP94 ↑, GRP78 ↑, CHOP ↑ | [117] |
| cleavage of PARP ↑, cleavage of caspase3 ↓ | |||||
| ROS ↑ | |||||
| Androsace umbellata | Saxifragifolin | MDA-MB-231 | 5 µM, 24 h | IRE1α ↑, calnexin ↑, calpain ↑, XBP1 ↑, CHOP ↑, GRP78/BiP ↑, | [118] |
| cleavage of PARP ↑, cleavage of caspase-3,-9 ↑, Bax ↑, cytochrome C ↑, p-JNK ↑ | |||||
| ROS ↑ | |||||
| MCF7 | 5 µM, 24 h | IRE1α ↑, Calnexin ↑, calpain ↑, XBP-1 ↑, GRP78/BiP ↑, CHOP ↑ | |||
| cytochrome c ↑ | |||||
| Serratia marcescens. | Prodigiosin | MCF7 MDA-MB-231T-47D | 100 µM, 24 h | GRP78 ↑, CHOP ↑, p-IRE1 ↑, IRE1 ↑, p-eIF2a ↑, eIF2a ↑, ATF6 ↑ | [119] |
| cleavage of PARP ↑, p-JNK ↑, JNK ↑, BCL-2 ↓ | |||||
| Fucus vesiculosus | Fucoidan | MDA-MB-231 | 100 μg/mL, 72 h | CHOP ↑, ATF4 ↑, p-eIF2α ↑, GRP78/BiP ↓, p-IRE1 ↓, XBP1 ↓ | [120] |
| Bax ↑, CaMK II ↑, cleavage of caspase-3,-12 ↑, cleavage of PARP ↑ | |||||
| γ-tocotrienol | MDA-MB-231 | 40 µM, 16 h | CHOP ↑, ATF4 ↑, GRP78/BiP ↑, XBP1 ↑ | [101] | |
| cleavage of PARP ↑, cleavage of caspase-3,-8,-9 ↑, DR5 ↑, JNK ↑, p-JNK, C-Jun ↑, p38 MAPK ↑ | |||||
| MCF-7 | 40 µM, 16 h | CHOP ↑, GRP78/BiP ↑, XBP1 ↑ | |||
| cleavage of PARP ↑, cleavage of caspase-8,-9 ↑, JNK ↑, p-JNK ↑, C-Jun ↑, DR5 ↑, p38 MAPK ↑ | |||||
| Brazilian Red propolis | Ethanol extract of Brazilian Red propolis | MCF-7 | 20 μg/mL, 24 h | CHOP ↑ | [121] |
| cleavage of caspase-3 ↑, Bax ↑, BcL-xL ↓, BcL-2 ↓ | |||||
| Ampelopsis grossedentata | Ampelopsin | MCF-7 MDA-MB-231 | 60 µM, 24 h | GRP78/BiP ↑, p-PERK ↑, p-eIF2α ↑, ATF6 ↑, CHOP ↑ | [122] |
| ROS ↑ |
| Family Name | Compound | Cell Line | Duration/Dosage | Mechanism | Reference |
|---|---|---|---|---|---|
| Curcuma longa | Curcumin | HT-29 | 40 μM, 24 h | release of intracellular Ca2+, Δψm ↓, CHOP ↑ | [125] |
| cleavage of caspase-3,-7,-8,-9 ↑, Fas ↑, FasL ↑ FADD ↑ Bid ↑, tBid ↑, p-JNK ↑, Bax ↑, cytochrome c ↑, Bcl-2 ↓ | |||||
| Olea europaea | 2-3,4 dihydroxyphenylethanol | HT-29 | 400 μM, 16 h, | release of intracellular Ca2+ ↑, Δψm ↓, IRE1 ↑, XBP1 ↑, GRP78/BiP ↑, PERK ↑, eIF2α ↑, CHOP ↑, | [126] |
| Bax ↑, Bak ↑, Bad ↑, cytochrome c ↑, cleavage of caspase 3 ↑, TRAF2 ↑, ASK ↑, JNK ↑, AP-1 ↑, p-JNK ↑ C-jun ↑, PI3K/Akt ↓, Bcl-2 ↓ | |||||
| Eupenicillium brefeldianum | Brefeldin A (BFA) | Colo 205 | 15 ng/mL, 24 h | GRP78 ↑, XBP1 ↑, CHOP ↑ | [127] |
| Resveratrol | HT29 | 50 μM, 24 h | GRP78/BiP ↑, CHOP ↑, XBP1 ↑, eIF2α ↑ | [128] | |
| cleavage of caspase-4 ↑, cleavage of PARP ↑ | |||||
| Zingiber zerumbet Smith | Zerumbone | HCT116-p53null | 20 μM, 24 h | ATF4 ↑, CHOP ↑, GRP78/BiP ↑, p-PERK ↑, PERK ↑ eIF2α ↑, p-eIF2α ↑ | [129] |
| DR5 ↑ | |||||
| ROS ↑ | |||||
| SW480 | 20 μM, 24 h | ATF4 ↑, CHOP ↑, GRP78/BiP ↑, p-PERK ↑, PERK ↑ eIF2α ↑, p-eIF2α ↑ | |||
| DR5 ↑ | |||||
| ROS ↑ | |||||
| Garcinia xanthochymus | Guttiferone H | HCT116 | 10 μg/mL, 24 h | ATF4 ↑, XBP1 ↑, CHOP ↑ | [130] |
| Cleavage of caspases-3,-7 ↑ | |||||
| Cladosiphon okamuranus and Fucus evanescens | Fucoidan | HCT116 | 100 μg/mL, 72 h | GRP78↑, p-CaMKII ↑, eIF2a ↑, p-eIF2α ↑, CHOP ↑, IRE1 ↓, XBP1 ↓ | [120] |
| Cleavage of PARP ↑ | |||||
| Piper nigrum Linn and Piper longum Linn | Piperine | HT-29 | 100 μg/mL, 72 h | IRE1α ↑, CHOP ↑, GPR78/BiP ↑ | [131] |
| cleavage of PARP ↑, cytochrome c ↑, JNK ↑, MAPK ↑, PI3K/Akt ↓ | |||||
| ROS ↑ | |||||
| Alpinia pricei Hayata | Flavokawain B | HCT116 | 50 μM, 8 h | CHOP ↑, ATF4 ↑ | [132] |
| Bcl-2 ↓, MAPK ↑, PARP ↑, cytochrome c ↑, BIM ↑, Bak ↑ | |||||
| ROS ↑ |
| Family Name | Compound | Cell Line | Duration/Dosage | Mechanism | Reference |
|---|---|---|---|---|---|
| Curcuma longa | Curcumin | AGS | 20 µM, 24 h | Release of intracellular Ca2+ ↑, Δψm ↓, CHOP ↑ | [125] |
| cleavage of caspase-3,-7,-8,-9 ↑, cytochrome c ↑ | |||||
| Ulmus davidiana var. japonica | Ultrafine | SNU-1 | 200 μg/mL, 24 h | GRP78/BiP ↑, p-eIF2α ↑ | [135] |
| cleavage caspase-3,-6,-9 ↑, cleavage of PARP ↑ | |||||
| SNU-484 | 300 μg/mL, 24 h | p-eIF2α ↑, GRP78/BiP ↑ | |||
| cleavage of caspase-3,-6,-9 ↑, cleavage of PARP ↑, Bcl-2 ↓, Bcl-xL ↓ | |||||
| Magnolia officinalis | Honokiol | MKN45 | 40 µM, 8 h | GRP94 ↓, CHOP ↑, calpain ↑ | [136] |
| cleavage of caspase-7,-12 ↑, cleavage of PARP ↑ | |||||
| SCM-1 | 40 µM, 24 h | GRP94 ↓, CHOP ↑, calpain ↑ | |||
| cleavage of PARP ↑, | |||||
| vitamin E | Vitamin E succinate | SGC-7901 | 20 μg/mL, 24 h | release of Intracellular Ca2+ ↑, Δψm ↓ GRP78/BiP ↑, GRP94 ↓, PERK ↑, ATF4 ↑, ATF6 ↑, XBP1 ↑, CHOP ↑ | [137] |
| cleavage of caspase-4,-7,-12 ↑, p-JNK ↑, JNK ↑, cleavage of PARP ↑ | |||||
| vitamin E | A-tocopheryl succinate | SGC-7901 | 20 μg/mL, 24 h | GRP78/BiP ↑, CHOP ↑ | [138] |
| cleavage of caspase-4 ↑ | |||||
| ROS ↑ | |||||
| Fructus viticis | Casticin | BGC-823 | 1 μmol/mL, 24 h | CHOP ↑, p-eIF2α ↑, eIF2α ↑, GRP78/BiP ↑ | [139] |
| DR5 ↑, Bax ↑, Bid ↑, cleavage of caspase-3,-8,-9 ↑ | |||||
| ROS ↑ | |||||
| Curcuma longa | 1-(4-hydroxy-3-methoxyphenyl)-5-(2-nitrophenyl)penta-1,4-dien-3-one (WZ35) | SGC-7901 | 10 μM, 12 h | ATF6 ↑, ATF4 ↑, XBP1 ↑, CHOP ↑ | [140] |
| P-JNK ↑, Bax ↑, cleavage of caspase-3 ↑ Bcl-2 ↓ | |||||
| ROS ↑ | |||||
| SGC-7901xenograft in athymic BALB/Ca-nu/nu female mice | WZ35 (orally, 50 mg/kg) for 28 days | CHOP ↑ | |||
| cleavage of caspase-3 ↑ |
| Family Name | Compound | Cell Line | Duration/Dosage | Mechanism | Reference |
|---|---|---|---|---|---|
| Ardisia virens Kurz | Ardisianone | PC-3 | 10 μg/mL, 24 h | GRP78/BiP ↑ | [143] |
| cleavage ofcaspases-3,-7,-8,-9 ↑, cleavage of PARP ↑, Bcl-2 ↑, Bcl-xL ↑, Bak ↑, Bax ↑, Bid ↑, PI3K/Akt ↓, cytochrome c ↑, AIF ↑ | |||||
| ROS ↑ | |||||
| Camellia sinensis | Polyphenon E | PNT1a | 35 μg/mL, 12 h | ATF4 ↑, PERK ↑, p-eIF2α ↑, eIF2α ↑, GRP78/BiP ↑, CHOP ↑, XBP1 ↑ | [144] |
| cleavage of caspase-3,-7,-9 ↑, Bak ↑, Puma ↑, cleavage of PARP ↑ | |||||
| ROS ↑ | |||||
| PC3 | 145 μg/mL, 12 h | ATF4 ↑, PERK ↑, p-eIF2α ↑, eIF2α ↑, CHOP ↑, XBP1 ↑, | |||
| cleavage of caspase-3,-7,-9 ↑, Puma ↑, Bak ↑, Bax ↑, PARP ↑ AIF ↑ | |||||
| ROS ↑ | |||||
| Garcinia mangostana | Mangosteen Fruit Extract | LNCaP | 15 μg/mL, 24 h | PERK ↑, IRE1 ↑, CHOP ↑, GRP78/BiP ↑, Ero1 ↑, ER chaperone ↑, PDI ↑, XBP1 ↑, calnexin ↑ | [145] |
| Cleavage of caspase-3,-4 ↑, Bax ↑ | |||||
| 22Rv1 cells | 15 μg/mL, 24 h | PERK ↑, IRE1 ↑, CHOP ↑, GRP78/BiP ↑, Ero1 ↑, ER chaperone ↑, PDI ↑, XBP1 ↑, calnexin ↑ | |||
| Cleavage of caspase-3,-4 ↑, Bax ↑ | |||||
| Asterella angusta | Marchantin M | PC-3 | 10 μm, 48 h | GRP78/BiP ↑, CHOP ↑, XBP-1 ↑, p-eIF2α ↑,eIF2α ↑,ATF4 ↑, ATF6 ↑, ERAD ↓ | [146] |
| Cleavage of caspase-3,-4 ↑ | |||||
| DU145 | 10 μM, 48 h | GRP78/BiP ↑, CHOP ↑, XBP1 ↑, p-eIF2α ↑, eIF2α ↑, ATF4 ↑, ATF6 ↑, ERAD ↓ | |||
| Cleavage of caspase-3,-4 ↑ | |||||
| LNCaP | 10 μM, 48 h | GRP78/BiP, CHOP ↑, XBP1 ↑, p-eIF2α ↑, eIF2α ↑, ATF4 ↑, ATF6 ↑, ERAD ↓ | |||
| Cleavage of caspase-3,-4 ↑ | |||||
| Monascus pilosus | Monascuspiloin | PC-3 | 25 μM, 48 h | IRE1α ↑, p-eIF2α ↑, eIF2α ↑ | [147] |
| Quercetin | PC-3 | 150 μM, 48 h | Release of intracellular Ca2+ ↑, Δψm ↓, GRP78/BiP ↑, ATF4 ↑, IRE1α ↑ ATF6 ↑ | [148] | |
| Bid ↓, Bcl-2 ↓, cleavage of caspase-12 ↓Bax ↑, PARP ↑, cytochrome c ↑, AIF ↑, Endo G ↑, cleavage of caspase-3,-8,-9 ↑ | |||||
| Zingiber zerumbet Smith | Zerumbone | PC-3 | 30 μM, 24 h | Release of intracellular Ca2+ ↑, Δψm ↓, calpain ↑, GRP78, CHOP ↑ | [149] |
| cleavage of caspase-3,-7,-9 ↑, cleavage of PARP ↑, Bid ↑, Bcl-2 ↓ |
| Family Name | Compound | Cell Line | Duration/Dosage | Mechanism | Reference |
|---|---|---|---|---|---|
| Glycyrrhiza inflate | Licochalcone A | HepG2 | 10 μM, 24 h | release of intracellular Ca2+ ↑, Δψm ↓, ATF6 ↑, eIF2α ↑, IRE1α ↑, CHOP ↑, GRP94 ↑, XBP1 ↑, GRP78/BiP ↑ | [153] |
| Cleavage of caspases-3,-4,-9 ↑, cleavage of PARP ↑ | |||||
| ROS ↑ | |||||
| Commiphora mukul | Guggulsterone | Hep3B | 30 μM, 12 h | Release of intracellular Ca2+ ↑, Δψm ↓, IRE1 ↑, JNK ↑, GRP78/BiP, PERK ↑, eIF2α ↑, ATF4 ↑, CHOP ↑ | [154] |
| DR5 ↑, cleavage of caspase-3 ↑, cleavage of PARP ↑ | |||||
| ROS ↑ | |||||
| Verrucarin A | Hep3B | 1 μM, 12 h | GRP78/BiP ↑, p-PERK ↑, p-eIF2α ↑, CHOP ↑ | [155] | |
| DR5 ↑, cleavage of caspase-3,-8 ↑, cleavage of PARP ↑ | |||||
| ROS ↑ | |||||
| HepG2 | 1 μM, 12 h | Chop ↑ | |||
| DR 5 ↑ | |||||
| 7-dimethoxyflavone | Hep3B | 5 μmol, 24 h | CHOP ↑, GPR78/BiP ↑, ATF4 ↑ | [156] | |
| DR5 ↑, cleavage of caspase-3,-8,-9 ↑ | |||||
| ROS ↑ | |||||
| Nelumbo nucifera Gaertn | Neferine | Hep3B | 20 μmol, 24 h | GRP78/BiP ↑, calnexin ↑, PDI ↑, calpain ↑ | [157] |
| cleavage of caspase-3,-6,-7,-8,-12, cleavage of PARP ↑, Puma ↑, BIM ↑, Bid ↑ | |||||
| Pycnostelma paniculatum K Schum | Paeonol | HepG2 | 31.25 mg/mL, 24 h | GRP78 ↑, CHOP ↑ | [158] |
| cleavage of caspase-3 ↑, PI3K/AKT ↓ | |||||
| Salvia miltiorrhiza Bunge | Cryptotanshinone | HepG2 | 10 μM, 24 h | eIF2α ↑, GRP94 ↑, GRP78/BiP ↑, cisplatin ↑, CHOP ↑ | [117] |
| cleavage of PARP ↑, JNK ↑, MAPK ↑ | |||||
| ROS ↑ | |||||
| Zingiber officinale Rosc | 6-Shogaol | SMMC-7721 | 20 μM, 6 h | GRP94 ↑, GRP78/BiP ↑, CHOP ↑, p-PERK ↑, PERK ↑, eIF2α ↑, p-eIF2α ↑ | [159] |
| cleavage of PARP ↑, cleavage of caspase-3 ↑ | |||||
| SMMC-7721 xenograft in Male SCID mice | 6-shogaol (orally, 10 mg/kg), 28 days | p-PERK ↓, eIF2α ↓, p-eIF2α ↓ | |||
| cleavage of caspase-3 ↑ | |||||
| Genistein | Hep3B | 100 μM, 48 h | release of intracellular Ca2+ ↑, Δψm ↓, calpain ↑, CHOP ↑, GRP78/BiP ↑ | [160] | |
| cleavage of caspase-2,-3,-7,-12 ↑, cleavage of PARP ↑, Apaf-1 ↑ cytochrome c ↑, Bad ↑ | |||||
| ROS ↑ |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, C.; Kim, B. Anti-Cancer Natural Products and Their Bioactive Compounds Inducing ER Stress-Mediated Apoptosis: A Review. Nutrients 2018, 10, 1021. https://doi.org/10.3390/nu10081021
Kim C, Kim B. Anti-Cancer Natural Products and Their Bioactive Compounds Inducing ER Stress-Mediated Apoptosis: A Review. Nutrients. 2018; 10(8):1021. https://doi.org/10.3390/nu10081021
Chicago/Turabian StyleKim, Changmin, and Bonglee Kim. 2018. "Anti-Cancer Natural Products and Their Bioactive Compounds Inducing ER Stress-Mediated Apoptosis: A Review" Nutrients 10, no. 8: 1021. https://doi.org/10.3390/nu10081021
APA StyleKim, C., & Kim, B. (2018). Anti-Cancer Natural Products and Their Bioactive Compounds Inducing ER Stress-Mediated Apoptosis: A Review. Nutrients, 10(8), 1021. https://doi.org/10.3390/nu10081021