Immunomodulatory Protein Hydrolysates and Their Application



Abstract
1. Introduction
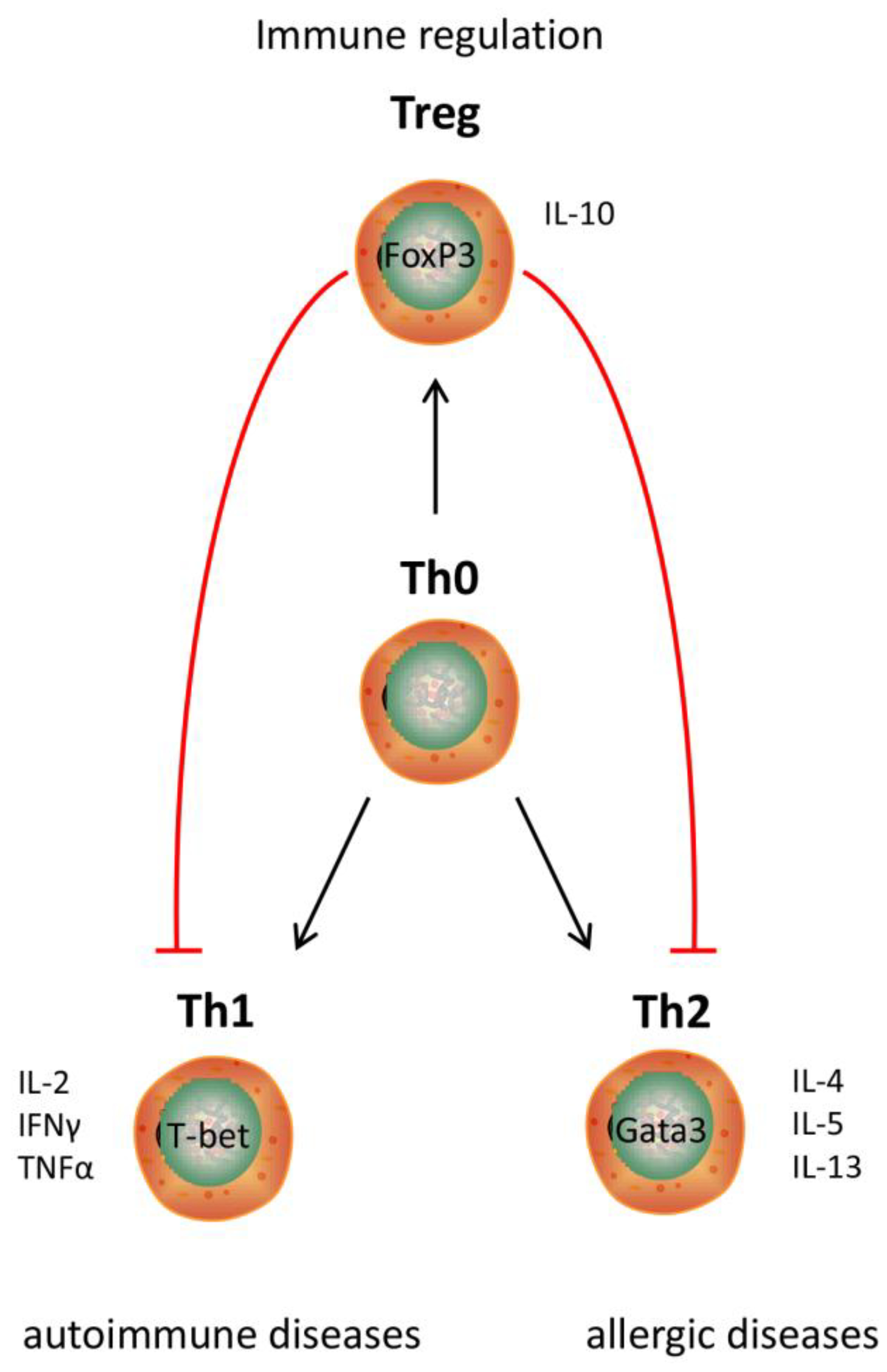
2. Immune Effects of Hydrolysates
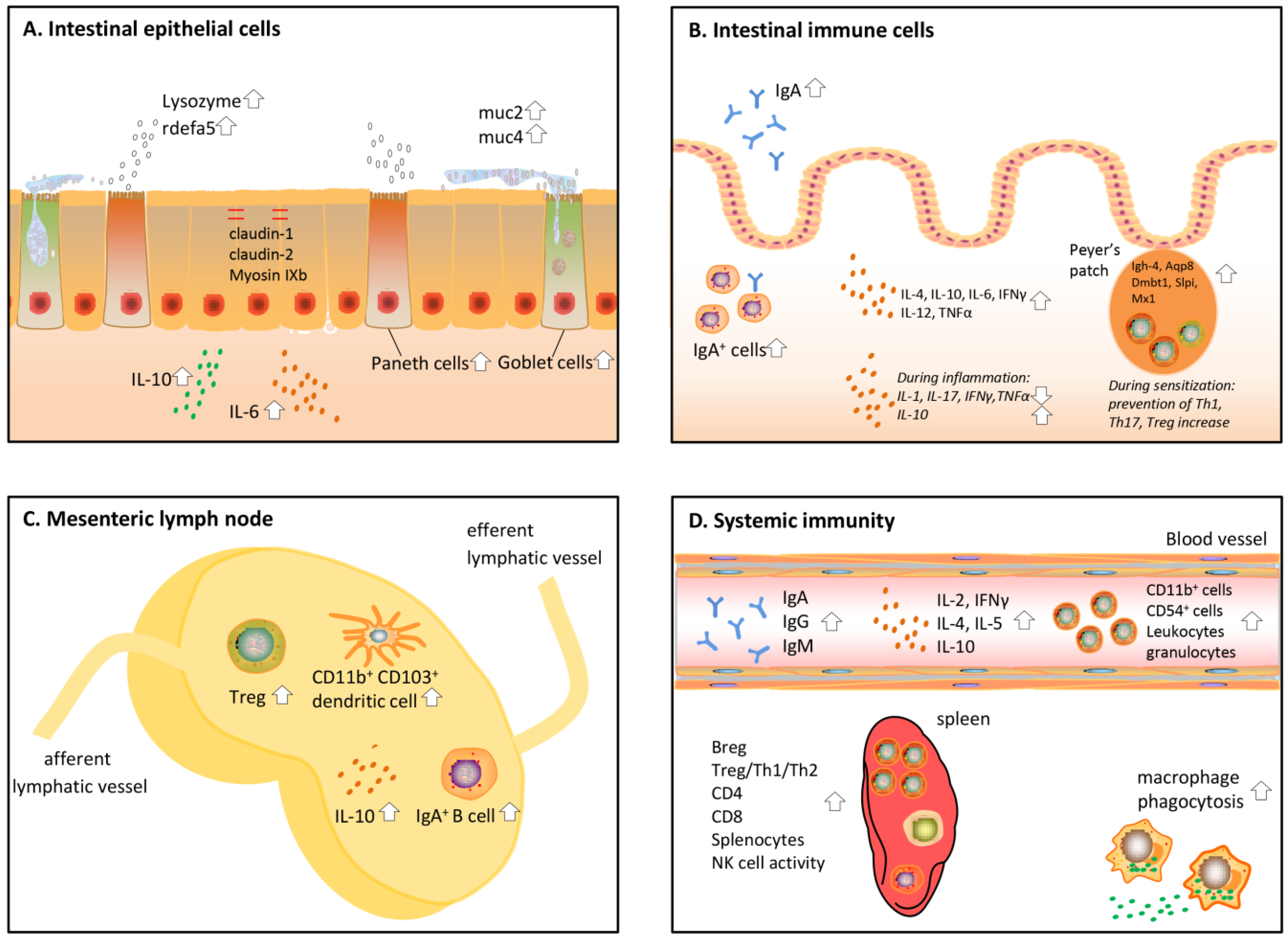
3. Effects of Protein Hydrolysates on the Gut Barrier
4. Effects of Protein Hydrolysates on the Intestinal Immune System
5. Effects of Protein Hydrolysates on the Mesenteric Lymph Nodes (MLN)
6. Effects of Protein Hydrolysates on Systemic Immunity
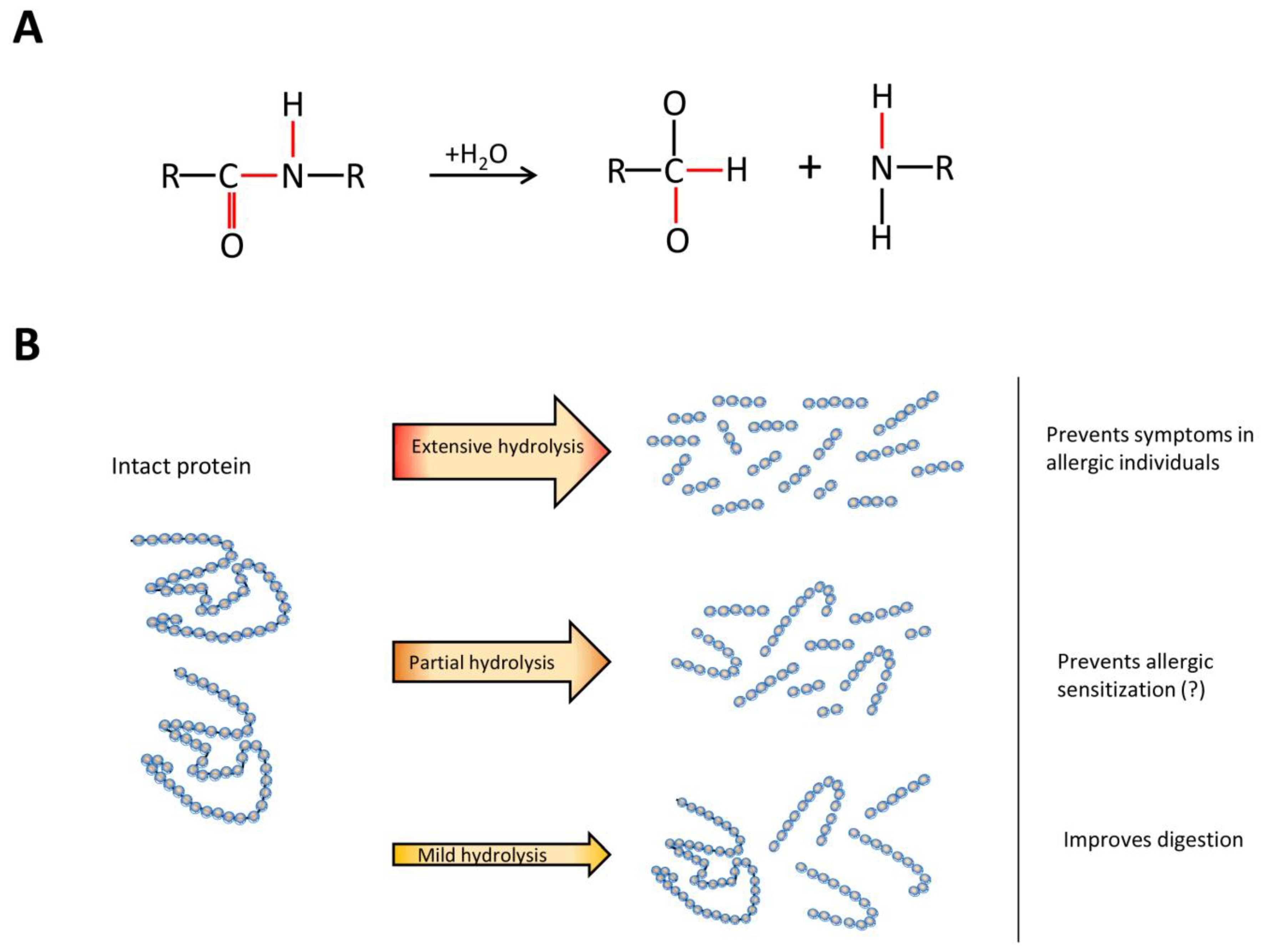
7. Understanding Hydrolysate Compositions
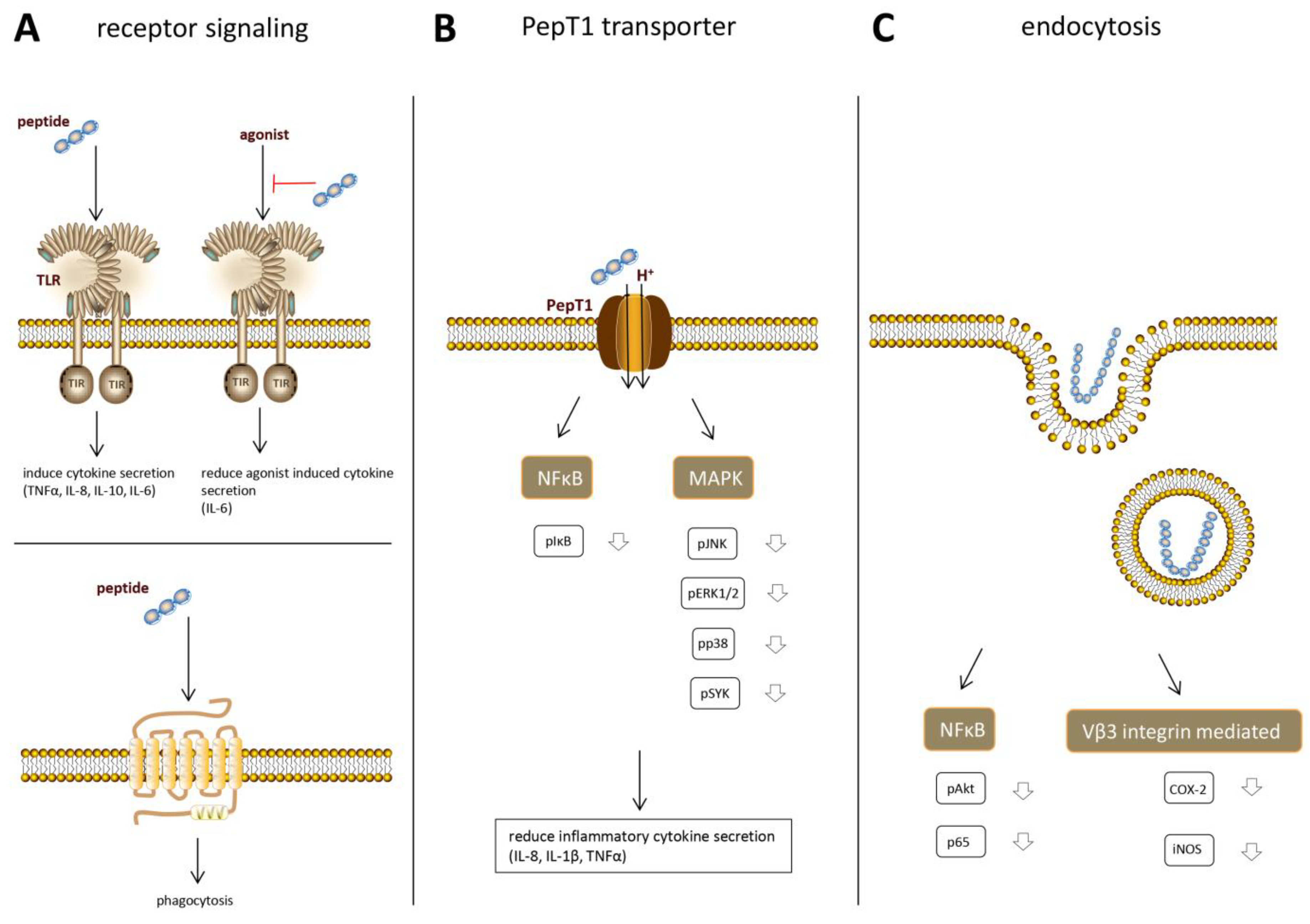
8. Underlying Mechanisms of Immunomodulatory Effects
9. Receptor Binding
10. PepT1 Dependent Intracellular Effects
11. Endocytosis
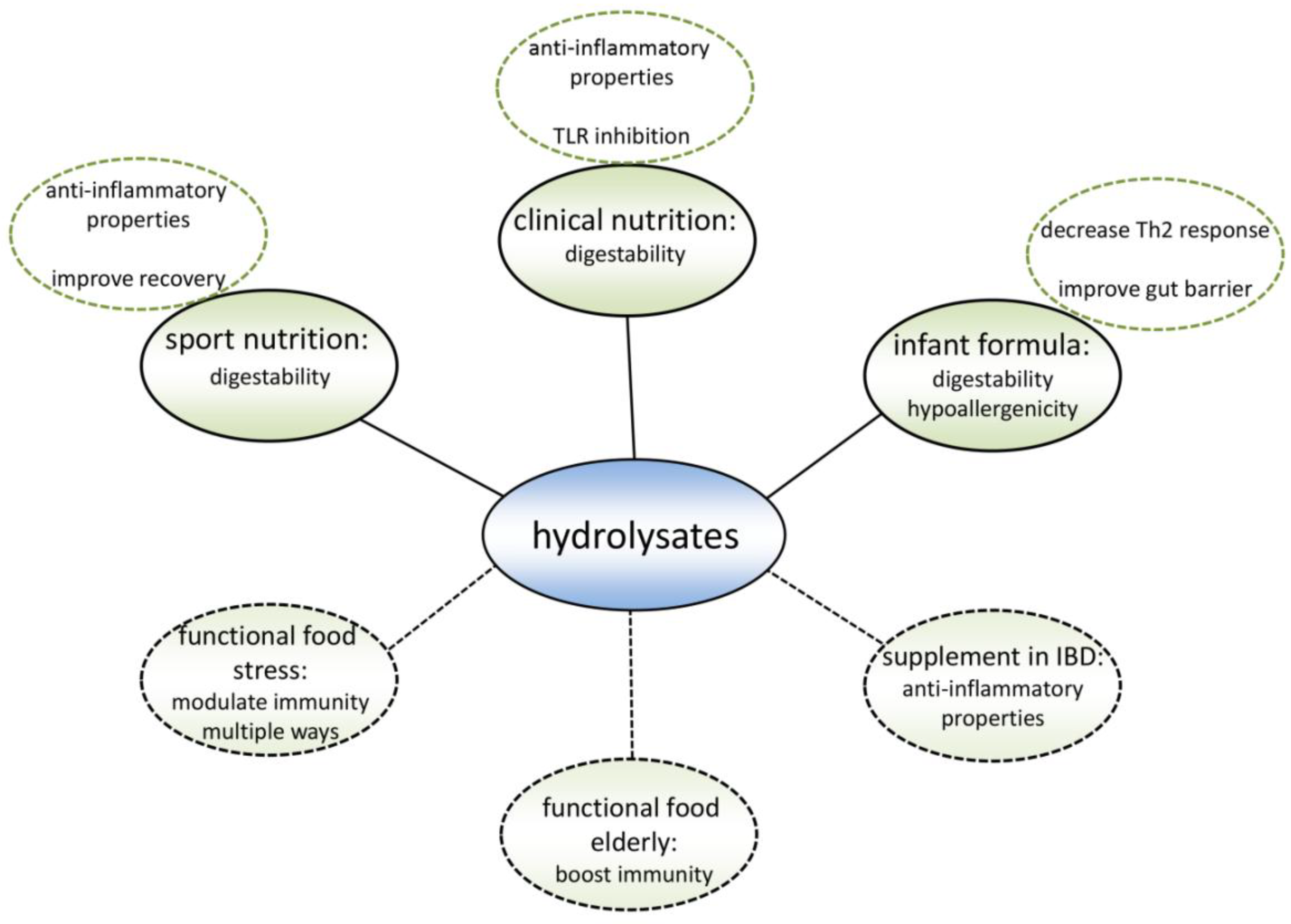
12. Possibilities for Hydrolysate Application
13. Existing Products
14. Target Groups for New Products
15. Conclusions
Author Contributions
Conflicts of Interest
References
- Sánchez, A.; Vázquez, A. Bioactive peptides: A review. Food Qual. Saf. 2017, 1, 29–46. [Google Scholar] [CrossRef]
- Li-Chan, E.C.Y. Bioactive peptides and protein hydrolysates: Research trends and challenges for application as nutraceuticals and functional food ingredients. Curr. Opin. Food Sci. 2015, 1, 28–37. [Google Scholar] [CrossRef]
- Dhaval, A.; Yadav, N.; Purwar, S. Potential Applications of Food Derived Bioactive Peptides in Management of Health. Int. J. Pept. Res. Ther. 2016, 22, 377–398. [Google Scholar] [CrossRef]
- Bouglé, D.; Bouhallab, S. Dietary bioactive peptides: Human studies. Crit. Rev. Food Sci. Nutr. 2015, 57, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Kiewiet, M.B.G.; Gros, M.; van Neerven, R.J.J.; Faas, M.M.; de Vos, P. Immunomodulating properties of protein hydrolysates for application in cow’s milk allergy. Pediatr. Allergy Immunol. 2015, 26, 206–217. [Google Scholar] [CrossRef] [PubMed]
- Parker, F.; Migliore-Samour, D.; Floch, F.; Zerial, A.; Werner, G.H.; Jollès, J.; Casaretto, M.; Zahn, H.; Jollès, P. Immunostimulating hexapeptide from human casein: Amino acid sequence, synthesis and biological properties. Eur. J. Biochem. 1984, 145, 677–682. [Google Scholar] [CrossRef] [PubMed]
- Ashaolu, T.J.; Yanyiam, N.; Yupanqui, C.T. Immunomodulatory effects of pepsin-educed soy protein hydrolysate in rats and murine cells. Funct. Foods Health Dis. 2017, 7, 889–900. [Google Scholar]
- Caron, S.; Samanez, C.H.; Dehondt, H.; Ploton, M.; Briand, O.; Lien, F.; Dorchies, E.; Dumont, J.; Postic, C.; Cariou, B.; et al. Farnesoid X receptor inhibits the transcriptional activity of carbohydrate response element binding protein in human hepatocytes. Mol. Cell. Biol. 2013, 33, 2202–2211. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Guo, M.; Hua, Y.; Cao, D.; Zhang, C. Enzymatic preparation of immunomodulating hydrolysates from soy proteins. Bioresour. Technol. 2008, 99, 8873–8879. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.Y.; Yang, H.Y.; Song, J.P.; Li, Y.H.; Ren, F.Z. Effect of yak milk casein hydrolysate on Th1/Th2 cytokines production by murine spleen lymphocytes in vitro. J. Agric. Food Chem. 2007, 55, 638–642. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Carrio, J.; Fernández, A.; Riera, F.A.; Suárez, A. Immunomodulatory activities of whey β-lactoglobulin tryptic-digested fractions. Int. Dairy J. 2014, 34, 65–73. [Google Scholar] [CrossRef]
- Wu, W.; Zhang, M.; Sun, C.; Brennan, M.; Li, H.; Wang, G.; Lai, F.; Wu, H. Enzymatic preparation of immunomodulatory hydrolysates from defatted wheat germ (Triticum Vulgare) globulin. Int. J. Food Sci. Technol. 2016, 51, 2556–2566. [Google Scholar] [CrossRef]
- He, X.Q.; Cao, W.H.; Pan, G.K.; Yang, L.; Zhang, C.H. Enzymatic hydrolysis optimization of Paphia undulata and lymphocyte proliferation activity of the isolated peptide fractions. J. Sci. Food Agric. 2015, 95, 1544–1553. [Google Scholar] [CrossRef] [PubMed]
- Lozano-Ojalvo, D.; Molina, E.; López-Fandiño, R. Hydrolysates of egg white proteins modulate T- and B-cell responses in mitogen-stimulated murine cells. Food Funct. 2016, 7, 1048–1056. [Google Scholar] [CrossRef] [PubMed]
- Sutas, Y.; Soppi, E.; Korhonen, H.; Syvaoja, E.L.; Saxelin, M.; Rokka, T.; Isolauri, E. Suppression of lymphocyte proliferation in vitro by bovine caseins hydrolyzed with Lactobacillus casei GG-derived enzymes. J. Allergy Clin. Immunol. 1996, 98, 216–224. [Google Scholar] [CrossRef]
- Meram, C.; Wu, J. Anti-inflammatory effects of egg yolk livetins (α, β, and γ-livetin) fraction and its enzymatic hydrolysates in lipopolysaccharide-induced RAW 264.7 macrophages. Food Res. Int. 2017, 100, 449–459. [Google Scholar] [CrossRef] [PubMed]
- Kazlauskaite, J.; Biziulevicius, G.A.; Zukaite, V.; Biziuleviciene, G.; Miliukiene, V.; Siaurys, A. Oral tryptic casein hydrolysate enhances phagocytosis by mouse peritoneal and blood phagocytic cells but fails to prevent induced inflammation. Int. Immunopharmacol. 2005, 5, 1936–1944. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.; Chen, Y.; Zhang, L.; Yu, H.; Xu, Z.; You, H.; Cheng, Y. Rice protein hydrolysates (RPHs) inhibit the LPS-stimulated inflammatory response and phagocytosis in RAW264.7 macrophages by regulating the NF-κB signaling pathway. RSC Adv. 2016, 6, 71295–71304. [Google Scholar] [CrossRef]
- Visser, J.; Lammers, K.; Hoogendijk, A.; Boer, M.; Brugman, S.; Beijer-Liefers, S.; Zandvoort, A.; Harmsen, H.; Welling, G.; Stellaard, F.; et al. Restoration of impaired intestinal barrier function by the hydrolysed casein diet contributes to the prevention of type 1 diabetes in the diabetes-prone BioBreeding rat. Diabetologia 2010, 53, 2621–2628. [Google Scholar] [CrossRef] [PubMed]
- Visser, J.; Bos, N.; Harthoorn, L.; Stellaard, F.; Beijer-Liefers, S.; Rozing, J.; van Tol, E.A.F. Potential mechanisms explaining why hydrolyzed casein-based diets outclass single amino acid-based diets in the prevention of autoimmune diabetes in diabetes-prone BB rats. Diabete Metab. Res. Rev. 2012, 28, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Plaisancié, P.; Claustre, J.; Estienne, M.; Henry, G.; Boutrou, R.; Paquet, A.; Léonil, J. A novel bioactive peptide from yoghurts modulates expression of the gel-forming MUC2 mucin as well as population of goblet cells and Paneth cells along the small intestine. J. Nutr. Biochem. 2013, 24, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Thoreux, K.; Balas, D.; Bouley, C.; Senegas-Balas, F. Diet Supplemented with Yoghurt or Milk Fermented by Lactobacillus casei DN-114 001 Stimulates Growth and Brush-Border Enzyme Activities in Mouse Small Intestine. Digestion 1998, 59, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Vinderola, G.; Matar, C.; Perdigón, G. Milk fermentation products of L. helveticus R389 activate calcineurin as a signal to promote gut mucosal immunity. BMC Immunol. 2007, 8, 19. [Google Scholar] [CrossRef] [PubMed]
- Nelson, R.; Katayama, S.; Mine, Y.; Duarte, J.; Matar, C. Immunomodulating effects of egg yolk low lipid peptic digests in a murine model. Food Agric. Immunol. 2007, 18, 1–15. [Google Scholar] [CrossRef]
- Chalamaiah, M.; Hemalatha, R.; Jyothirmayi, T.; Diwan, P.V.; Bhaskarachary, K.; Vajreswari, A.; Ramesh Kumar, R.; Dinesh Kumar, B. Chemical composition and immunomodulatory effects of enzymatic protein hydrolysates from common carp (Cyprinus carpio) egg. Nutrition 2015, 31, 388–398. [Google Scholar] [CrossRef] [PubMed]
- Ndiaye, F.; Vuong, T.; Duarte, J.; Aluko, R.E.; Matar, C. Anti-oxidant, anti-inflammatory and immunomodulating properties of an enzymatic protein hydrolysate from yellow field pea seeds. Eur. J. Nutr. 2012, 51, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Duarte, J.; Vinderola, G.; Ritz, B.; Perdigón, G.; Matar, C. Immunomodulating capacity of commercial fish protein hydrolysate for diet supplementation. Immunobiology 2006, 211, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Mallet, J.F.; Duarte, J.; Vinderola, G.; Anguenot, R.; Beaulieu, M.; Matar, C. The immunopotentiating effects of shark-derived protein hydrolysate. Nutrition 2014, 30, 706–712. [Google Scholar] [CrossRef] [PubMed]
- LeBlanc, J.; Fliss, I.; Matar, C. Induction of a Humoral Immune Response following an Escherichia coli O157:H7 Infection with an Immunomodulatory Peptidic Fraction Derived from Lactobacillus helveticus-Fermented Milk. Clin. Diagn. Lab. Immunol. 2004, 11, 1171–1181. [Google Scholar] [CrossRef] [PubMed]
- Daddaoua, A.; Puerta, V.; Zarzuelo, A.; Suárez, M.D.; Sánchez de Medina, F.; Martínez-Augustin, O. Bovine Glycomacropeptide Is Anti-Inflammatory in Rats with Hapten-Induced Colitis. J. Nutr. 2005, 135, 1164–1170. [Google Scholar] [CrossRef] [PubMed]
- Espeche Turbay, M.B.; De Leblanc, A.D.M.; Perdigón, G.; Savoy de Giori, G.; Hebert, E.M. β-Casein hydrolysate generated by the cell envelope-associated proteinase of Lactobacillus delbrueckii ssp. lactis CRL 581 protects against trinitrobenzene sulfonic acid-induced colitis in mice. J. Dairy Sci. 2012, 95, 1108–1118. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Gonzalez, M.; Capitan-Canadas, F.; Requena, P.; Ocon, B.; Romero-Calvo, I.; Aranda, C.; Suarez, M.D.; Zarzuelo, A.; de Medina, F.S.; Martinez-Augustin, O. Validation of bovine glycomacropeptide as an intestinal anti-inflammatory nutraceutical in the lymphocyte-transfer model of colitis. Br. J. Nutr. 2014, 111, 1202–1212. [Google Scholar] [CrossRef] [PubMed]
- Requena, P.; Daddaoua, A.; Martínez-Plata, E.; González, M.; Zarzuelo, A.; Suárez, M.D.; Sánchez de Medina, F.; Martínez-Augustin, O. Bovine glycomacropeptide ameliorates experimental rat ileitis by mechanisms involving downregulation of interleukin 17. Br. J. Pharmacol. 2009, 154, 825–832. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Kovacs-Nolan, J.; Archbold, T.; Fan, M.Z.; Juneja, L.R.; Okubo, T.; Mine, Y. Therapeutic potential of hen egg white peptides for the treatment of intestinal inflammation. J. Funct. Foods 2009, 1, 161–169. [Google Scholar] [CrossRef]
- Egusa, S.; Otani, H. Soybean protein fraction digested with neutral protease preparation, “Peptidase R”, produced by Rhizopus oryzae, stimulates innate cellular immune system in mouse. Int. Immunopharmacol. 2009, 9, 931–936. [Google Scholar] [CrossRef] [PubMed]
- Kiewiet, M.; van Esch, B.; Garssen, J.; Faas, M.; de Vos, P. Partially hydrolyzed whey proteins prevent clinical symptoms in a cow’s milk allergy mouse model and enhance regulatory T and B cell frequencies. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef] [PubMed]
- Meulenbroek, L.A.P.M.; van Esch, B.C.A.M.; Hofman, G.A.; den Hartog Jager, C.F.; Nauta, A.J.; Willemsen, L.E.M.; Bruijnzeel-Koomen, C.A.F.M.; Garssen, J.; van Hoffen, E.; Knippels, L.M.J. Oral treatment with beta-lactoglobulin peptides prevents clinical symptoms in a mouse model for cow’s milk allergy. Pediatr. Allergy Immunol. 2013, 24, 656–664. [Google Scholar] [CrossRef] [PubMed]
- Van Esch, B.C.A.M.; Schouten, B.; de Kivit, S.; Hofman, G.A.; Knippels, L.M.J.; Willemsen, L.E.M.; Garssen, J. Oral tolerance induction by partially hydrolyzed whey protein in mice is associated with enhanced numbers of Foxp3(+) regulatory T-cells in the mesenteric lymph nodes. Pediatr. Allergy Immunol. 2011, 22, 820–826. [Google Scholar] [CrossRef] [PubMed]
- Cai, B.; Pan, J.; Wu, Y.; Wan, P.; Sun, H. Immune functional impacts of oyster peptide-based enteral nutrition formula (OPENF) on mice: A pilot study. Chin. J. Oceanol. Limnol. 2013, 31, 813–820. [Google Scholar] [CrossRef]
- Pan, D.D.; Wu, Z.; Liu, J.; Cao, X.Y.; Zeng, X.Q. Immunomodulatory and hypoallergenic properties of milk protein hydrolysates in ICR mice. J. Dairy Sci. 2013, 96, 4958–4964. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Zhang, Z.; Pei, X.; Han, X.; Wang, J.; Wang, L.; Long, Z.; Shen, X.; Li, Y. Immunomodulatory effects of marine oligopeptide preparation from Chum Salmon (Oncorhynchus keta) in mice. Food Chem. 2009, 113, 464–470. [Google Scholar] [CrossRef]
- Kim, M.J.; Kim, K.B.W.R.; Sung, N.Y.; Byun, E.H.; Nam, H.S.; Ahn, D.H. Immune-enhancement effects of tuna cooking drip and its enzymatic hydrolysate in Balb/c mice. Food Sci. Biotechnol. 2018, 27, 131–137. [Google Scholar] [CrossRef]
- Yimit, D.; Hoxur, P.; Amat, N.; Uchikawa, K.; Yamaguchi, N. Effects of soybean peptide on immune function, brain function, and neurochemistry in healthy volunteers. Nutrition 2012, 28, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Horiguchi, N.; Horiguchi, H.; Suzuki, Y. Effect of wheat gluten hydrolysate on the immune system in healthy human subjects. Biosci. Biotechnol. Biochem. 2005, 69, 2445–2449. [Google Scholar] [CrossRef] [PubMed]
- Nesse, K.O.; Nagalakshmi, A.P.; Marimuthu, P.; Singh, M. Efficacy of a Fish Protein Hydrolysate in Malnourished Children. Indian J. Clin. Biochem. 2011, 26, 360–365. [Google Scholar] [CrossRef] [PubMed]
- Farhadi, A.; Banan, A.; Fields, J.; Keshavarzian, A. Intestinal barrier: An interface between health and disease. J. Gastroenterol. Hepatol. 2003, 18, 479–497. [Google Scholar] [CrossRef] [PubMed]
- Peterson, L.W.; Artis, D. Intestinal epithelial cells: Regulators of barrier function and immune homeostasis. Nat. Rev. Immunol. 2014, 14, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Poursine-Laurent, J.; Wu, H.; Link, D.C. Interleukin-6 and the granulocyte colony-stimulating factor receptor are major independent regulators of granulopoiesis in vivo but are not required for lineage commitment or terminal differentiation. Blood 1997, 90, 2583–2590. [Google Scholar] [PubMed]
- Smith, K.A.; Maizels, R.M. IL-6 controls susceptibility to helminth infection by impeding Th2 responsiveness and altering the Treg phenotype in vivo. Eur. J. Immunol. 2014, 44, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Hunter, C.A.; Jones, S.A. IL-6 as a keystone cytokine in health and disease. Nat. Immunol. 2015, 16, 448–457. [Google Scholar] [CrossRef] [PubMed]
- Pabst, O. New concepts in the generation and functions of IgA. Nat. Rev. Immunol. 2012, 12, 821–832. [Google Scholar] [CrossRef] [PubMed]
- Ligtenberg, A.; Veerman, E.; Nieuw Amerongen, A.; Mollenhauer, J. Salivary agglutinin/glycoprotein-340/DMBT1: A single molecule with variable composition and with different functions in infection, inflammation and cancer. Biol. Chem. 2007, 388, 1275–1289. [Google Scholar] [CrossRef] [PubMed]
- Fernie-King, B.; Seilly, D.; Binks, M.; Sriprakash, K.; Lachmann, P. Streptococcal DRS (distantly related to SIC) and SIC inhibit antimicrobial peptides, components of mucosal innate immunity: A comparison of their activities. Microbes Infect. 2007, 9, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Samuel, C.E. Antiviral actions of interferons. Clin. Microbiol. Rev. 2001, 14, 778–809. [Google Scholar] [CrossRef] [PubMed]
- Macpherson, A.J.; Smith, K. Mesenteric lymph nodes at the center of immune anatomy. J. Exp. Med. 2006, 203, 497–500. [Google Scholar] [CrossRef] [PubMed]
- Santiago, A.F.; Fernandes, R.M.; Santos, B.P.; Assis, F.A.; Oliveira, R.P.; Carvalho, C.R.; Faria, A.M.C. Role of mesenteric lymph nodes and aging in secretory IgA production in mice. Cell. Immunol. 2008, 253, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Chabance, B.; Marteau, P.; Rambaud, J.C.; Migliore-Samour, D.; Boynard, M.; Perrotin, P.; Guillet, R.; Jolles, P.; Fiat, A.M. Casein peptide release and passage to the blood in humans during digestion of milk or yogurt. Biochimie 1998, 80, 155–165. [Google Scholar] [CrossRef]
- Dia, V.P.; Torres, S.; De Lumen, B.O.; Erdman, J.W.; Gonzalez De Mejia, E. Presence of lunasin in plasma of men after soy protein consumption. J. Agric. Food Chem. 2009, 57, 1260–1266. [Google Scholar] [CrossRef] [PubMed]
- Rosser, E.C.; Mauri, C. Regulatory B Cells: Origin, Phenotype, and Function. Immunity 2015, 42, 607–612. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.R.; Kim, H.S.W.H.S.; Kim, D.K.; Nam, S.T.; Kim, H.S.W.H.S.; Park, Y.H.; Lee, D.; Lee, M.B.; Lee, J.H.; Kim, B.; et al. Mesenteric IL-10-producing CD5(+) regulatory B cells suppress cow’s milk casein-induced allergic responses in mice. Sci. Rep. 2016, 6, 19685. [Google Scholar] [CrossRef] [PubMed]
- Agyei, D.; Ongkudon, C.M.; Wei, C.Y.; Chan, A.S.; Danquah, M.K. Bioprocess challenges to the isolation and purification of bioactive peptides. Food Bioprod. Process. 2016, 98, 244–256. [Google Scholar] [CrossRef]
- Rutherfurd, S.M. Methodology for determining degree of hydrolysis of proteins in Hydrolysates: A review. J. AOAC Int. 2010, 93, 1515–1522. [Google Scholar] [PubMed]
- Silvestre, M.P.C. Review of methods for the analysis of protein hydrolysates. Food Chem. 1997, 60, 263–271. [Google Scholar] [CrossRef]
- Lemieux, L.; Piot, J.-M.; Guillochon, D.; Amiot, J. Study of the efficiency of a mobile phase used in size-exclusion HPLC for the separation of peptides from a casein hydrolysate according to their hydrodynamic volume. Chromatographia 1991, 32, 499–504. [Google Scholar] [CrossRef]
- Chen, H.-M.; Muramoto, K.; Yamauchi, F. Structural Analysis of Antioxidative Peptides from Soybean .beta.-Conglycinin. J. Agric. Food Chem. 1995, 43, 574–578. [Google Scholar] [CrossRef]
- Kiewiet, M.B.G.; Dekkers, R.; Ulfman, L.H.; Groeneveld, A.; de Vos, P.; Faas, M.M. Immunomodulating protein aggregates in soy and whey hydrolysates and their resistance to digestion in an in vitro infant gastrointestinal model: New insights in the mechanism of immunomodulatory hydrolysates. Food Funct. 2018, 9, 604–613. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhya, A.; Noronha, N.; Bahar, B.; Ryan, M.T.; Murray, B.A.; Kelly, P.M.; O’Loughlin, I.B.; O’Doherty, J.V.; Sweeney, T. The anti-inflammatory potential of a moderately hydrolysed casein and its 5 kDa fraction in in vitro and ex vivo models of the gastrointestinal tract. Food Funct. 2015, 6, 612–621. [Google Scholar] [CrossRef] [PubMed]
- Lafarga, T.; Hayes, M. Bioactive protein hydrolysates in the functional food ingredient industry: Overcoming current challenges. Food Rev. Int. 2017, 33, 217–246. [Google Scholar] [CrossRef]
- Chalamaiah, M.; Yu, W.; Wu, J. Immunomodulatory and anticancer protein hydrolysates (peptides) from food proteins: A review. Food Chem. 2018, 245, 205–222. [Google Scholar] [CrossRef] [PubMed]
- Monahan, F.J.; German, J.B.; Kinsella, J.E. Effect of Ph and Temperature on Protein Unfolding and Thiol-Disulfide Interchange Reactions during Heat-Induced Gelation of Whey Proteins. J. Agric. Food Chem. 1995, 43, 46–52. [Google Scholar] [CrossRef]
- Gordon, S. Pattern recognition receptors: Doubling up for the innate immune response. Cell 2002, 111, 927–930. [Google Scholar] [CrossRef]
- Michallet, M.-C.; Rota, G.; Maslowski, K.; Guarda, G. Innate receptors for adaptive immunity. Curr. Opin. Microbiol. 2013, 16, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Abreu, M.T. Toll-like receptor signalling in the intestinal epithelium: How bacterial recognition shapes intestinal function. Nat. Rev. Immunol. 2010, 10, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Kiewiet, M.B.G.; Dekkers, R.; Gros, M.; van Neerven, R.J.J.; Groeneveld, A.; de Vos, P.; Faas, M.M. Toll-like receptor mediated activation is possibly involved in immunoregulating properties of cow’s milk hydrolysates. PLoS ONE 2017, 12, e0178191. [Google Scholar] [CrossRef] [PubMed]
- Tobita, K.; Kawahara, T.; Otani, H. Bovine beta-casein (1-28), a casein phosphopeptide, enhances proliferation and IL-6 expression of mouse CD19(+) cells via toll-like receptor 4. J. Agric. Food Chem. 2006, 54, 8013–8017. [Google Scholar] [CrossRef] [PubMed]
- Iskandar, M.M.; Lands, L.C.; Sabally, K.; Azadi, B.; Meehan, B.; Mawji, N.; Skinner, C.D.; Kubow, S. High Hydrostatic Pressure Pretreatment of Whey Protein Isolates Improves Their Digestibility and Antioxidant Capacity. Foods 2015, 4, 184–207. [Google Scholar] [CrossRef] [PubMed]
- Tsuruki, T.; Kishi, K.; Takahashi, M.; Tanaka, M.; Matsukawa, T.; Yoshikawa, M. Soymetide, an immunostimulating peptide derived from soybean beta-conglycinin, is an fMLP agonist. FEBS Lett. 2003, 540, 206–210. [Google Scholar] [CrossRef]
- Stefanucci, A.; Mollica, A.; Macedonio, G.; Zengin, G.; Ahmed, A.A.; Novellino, E. Exogenous opioid peptides derived from food proteins and their possible uses as dietary supplements: A critical review. Food Rev. Int. 2018, 34, 70–86. [Google Scholar] [CrossRef]
- Liang, X.; Liu, R.; Chen, C.; Ji, F.; Li, T. Opioid System Modulates the Immune Function: A Review. Transl. Perioper. Pain Med. 2016, 1, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Börner, C.; Lanciotti, S.; Koch, T.; Höllt, V.; Kraus, J. μ opioid receptor agonist-selective regulation of interleukin-4 in T lymphocytes. J. Neuroimmunol. 2013, 263, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Cheido, M.A.; Gevorgyan, M.M.; Zhukova, E.N. Comparative Evaluation of Opioid-Induced Changes in Immune Reactivity of CBA Mice. Bull. Exp. Biol. Med. 2014, 156, 363–365. [Google Scholar] [CrossRef] [PubMed]
- Tomassini, N.; Renaud, F.; Roy, S.; Loh, H.H. Morphine inhibits Fc-mediated phagocytosis through mu and delta opioid receptors. J. Neuroimmunol. 2004, 147, 131–133. [Google Scholar] [CrossRef] [PubMed]
- Dalmasso, G.; Charrier-Hisamuddin, L.; Thu Nguyen, H.T.; Yan, Y.; Sitaraman, S.; Merlin, D. PepT1-Mediated Tripeptide KPV Uptake Reduces Intestinal Inflammation. Gastroenterology 2008, 134, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Kovacs-Nolan, J.; Zhang, H.; Ibuki, M.; Nakamori, T.; Yoshiura, K.; Turner, P.V.; Matsui, T.; Mine, Y. The PepT1-transportable soy tripeptide VPY reduces intestinal inflammation. Biochim. Biophys. Acta Gen. Subj. 2012, 1820, 1753–1763. [Google Scholar] [CrossRef] [PubMed]
- Oyama, M.; Van Hung, T.; Yoda, K.; He, F.; Suzuki, T. A novel whey tetrapeptide IPAV reduces interleukin-8 production induced by TNF-α in human intestinal Caco-2 cells. J. Funct. Foods 2017, 35, 376–383. [Google Scholar] [CrossRef]
- Adibi, S.A. Regulation of expression of the intestinal oligopeptide transporter (Pept-1) in health and disease. Am. J. Physiol. Liver Physiol. 2003, 285, G779–G788. [Google Scholar] [CrossRef] [PubMed]
- Bermudez-Brito, M.; Sahasrabudhe, N.M.; Rösch, C.; Schols, H.A.; Faas, M.M.; de Vos, P. The impact of dietary fibers on dendritic cell responses in vitro is dependent on the differential effects of the fibers on intestinal epithelial cells. Mol. Nutr. Food Res. 2015, 59, 698–710. [Google Scholar] [CrossRef] [PubMed]
- Charrier, L.; Driss, A.; Yan, Y.; Nduati, V.; Klapproth, J.-M.; Sitaraman, S.V.; Merlin, D. hPepT1 mediates bacterial tripeptide fMLP uptake in human monocytes. Lab. Investig. 2006, 86, 490–503. [Google Scholar] [CrossRef] [PubMed]
- Knipp, G.T.; Vander Velde, D.G.; Siahaan, T.J.; Borchardt, R.T. The effect of beta-turn structure on the passive diffusion of peptides across Caco-2 cell monolayers. Pharm. Res. 1997, 14, 1332–1340. [Google Scholar] [CrossRef] [PubMed]
- Regazzo, D.; Molle, D.; Gabai, G.; Tome, D.; Dupont, D.; Leonil, J.; Boutrou, R. The (193-209) 17-residues peptide of bovine beta-casein is transported through Caco-2 monolayer. Mol. Nutr. Food Res. 2010, 54, 1428–1435. [Google Scholar] [CrossRef] [PubMed]
- Cam, A.; Sivaguru, M.; Gonzalez de Mejia, E. Endocytic Mechanism of Internalization of Dietary Peptide Lunasin into Macrophages in Inflammatory Condition Associated with Cardiovascular Disease. PLoS ONE 2013, 8, e72115. [Google Scholar] [CrossRef] [PubMed]
- Kneepkens, C.M.F.; Meijer, Y. Clinical practice. Diagnosis and treatment of cow’s milk allergy. Eur. J. Pediatr. 2009, 168, 891–896. [Google Scholar] [CrossRef] [PubMed]
- Turcanu, V.; Brough, H.A.; Du Toit, G.; Foong, R.-X.; Marrs, T.; Santos, A.F.; Lack, G. Immune mechanisms of food allergy and its prevention by early intervention. Curr. Opin. Immunol. 2017, 48, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Berin, M.C.; Sampson, H.A. Review Mucosal Immunology of Food Allergy. Curr. Biol. 2013, 23, R389–R400. [Google Scholar] [CrossRef] [PubMed]
- Curotto de Lafaille, M.A.; Kutchukhidze, N.; Shen, S.; Ding, Y.; Yee, H.; Lafaille, J.J. Adaptive Foxp3+ Regulatory T Cell-Dependent and -Independent Control of Allergic Inflammation. Immunity 2008, 29, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Morgan, M.E.; Koelink, P.J.; Zheng, B.; den Brok, M.H.M.G.M.; van de Kant, H.J.; Verspaget, H.W.; Folkerts, G.; Adema, G.J.; Kraneveld, A.D. Toll-like receptor 6 stimulation promotes T-helper 1 and 17 responses in gastrointestinal-associated lymphoid tissue and modulates murine experimental colitis. Mucosal Immunol. 2014, 7, 1266–1277. [Google Scholar] [CrossRef] [PubMed]
- Dolina, J.S.; Schoenberger, S.P. Toll-like receptor 9 is required for the maintenance of CD25+FoxP3+CD4+ Treg cells during Listeria monocytogenes infection. J. Immunol. 2017, 198 (Suppl. 1), 151.9. [Google Scholar]
- Deutz, N.E.; Matheson, E.M.; Matarese, L.E.; Luo, M.; Baggs, G.E.; Nelson, J.L.; Hegazi, R.A.; Tappenden, K.A.; Ziegler, T.R.; Grp, N.S. Readmission and mortality in malnourished, older, hospitalized adults treated with a specialized oral nutritional supplement: A randomized clinical trial. Clin. Nutr. 2016, 35, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Elting, L.S.; Cooksley, C.; Chambers, M.; Cantor, S.B. The burdens of cancer therapy—Clinical and economic outcomes of chemotherapy-induced mucositis. Cancer 2003, 98, 1531–1539. [Google Scholar] [CrossRef] [PubMed]
- Kaczmarek, A.; Brinkman, B.M.; Heyndrickx, L.; Vandenabeele, P.; Krysko, D.V. Severity of doxorubicin-induced small intestinal mucositis is regulated by the TLR-2 and TLR-9 pathways. J. Pathol. 2012, 226, 598–608. [Google Scholar] [CrossRef] [PubMed]
- De Koning, B.A.E.; van Dieren, J.M.; Lindenbergh-Kortleve, D.J.; van der Sluis, M.; Matsumoto, T.; Yamaguchi, K.; Einerhand, A.W.; Samsom, J.N.; Pieters, R.; Nieuwenhuis, E.E.S. Contributions of mucosal immune cells to methotrexate-induced mucositis. Int. Immunol. 2006, 18, 941–949. [Google Scholar] [CrossRef] [PubMed]
- Cario, E. Toll-like receptors in the pathogenesis of chemotherapy-induced gastrointestinal toxicity. Curr. Opin. Support. Palliat. Care 2016, 10, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Villa, A.; Sonis, S.T. Mucositis: Pathobiology and management. Curr. Opin. Oncol. 2015, 27, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Sahasrabudhe, N.M.; Beukema, M.; Tian, L.; Troost, B.; Scholte, J.; Bruininx, E.; Bruggeman, G.; van den Berg, M.; Scheurink, A.; Schols, H.A.; et al. Dietary Fiber Pectin Directly Blocks Toll-Like Receptor 2–1 and Prevents Doxorubicin-Induced Ileitis. Front. Immunol. 2018, 9, 383. [Google Scholar] [CrossRef] [PubMed]
- Crittenden, R.; Buckley, J.; Cameron-Smith, D.; Brown, A.; Thomas, K.; Davey, S.; Hobman, P. Functional dairy protein supplements for elite athletes. Aust. J. Dairy Technol. 2009, 64, 133–138. [Google Scholar]
- Foltz, M.; Ansems, P.; Schwarz, J.; Tasker, M.C.; Lourbakos, A.; Gerhardt, C.C. Protein hydrolysates induce CCK release from enteroendocrine cells and act as partial agonists of the CCK1receptor. J. Agric. Food Chem. 2008, 56, 837–843. [Google Scholar] [CrossRef] [PubMed]
- Kuehl, K.S.; Perrier, E.T.; Elliot, D.L.; Chesnutt, J.C. Efficacy of tart cherry juice in reducing muscle pain during running: A randomized controlled trial. J. Int. Soc. Sports Nutr. 2010, 7, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Howatson, G.; McHugh, M.P.; Hill, J.A.; Brouner, J.; Jewell, A.P.; Van Someren, K.A.; Shave, R.E.; Howatson, S.A. Influence of tart cherry juice on indices of recovery following marathon running. Scand. J. Med. Sci. Sport. 2010, 20, 843–852. [Google Scholar] [CrossRef] [PubMed]
- Koikawa, N.; Nakamura, A.; Ngaoka, I.; Aoki, K.; Sawaki, K.; Suzuki, Y. Delayed-onset muscle injury and its modification by wheat gluten hydrolysate. Nutrition 2009, 25, 493–498. [Google Scholar] [CrossRef] [PubMed]
- Hansen, M.; Bangsbo, J.; Bibby, B.M.; Madsen, K. Effect of whey protein hydrolysate on performance and recovery of top-clas orienteering runners. Int. J. Sport Nutr. Exerc. Metab. 2014, 25, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Buckley, J.D.; Thomson, R.L.; Coates, A.M.; Howe, P.R.C.; DeNichilo, M.O.; Rowney, M.K. Supplementation with a whey protein hydrolysate enhances recovery of muscle force-generating capacity following eccentric exercise. J. Sci. Med. Sport 2010, 13, 178–181. [Google Scholar] [CrossRef] [PubMed]
- Cruzat, V.F.; Krause, M.; Newsholme, P. Amino acid supplementation and impact on immune function in the context of exercise. J. Int. Soc. Sports Nutr. 2014, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Okada, H.; Kuhn, C.; Feillet, H.; Bach, J.-F. The “hygiene hypothesis” for autoimmune and allergic diseases: An update. Clin. Exp. Immunol. 2010, 160, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Dabelea, D.; Mayer-Davis, E.J.; Saydah, S.; Imperatore, G.; Linder, B.; Divers, J.; Bell, R.; Badaru, A.; Talton, J.W.; Crume, T.; et al. Prevalence of Type 1 and Type 2 Diabetes Among Children and Adolescents From 2001 to 2009. JAMA 2014, 311, 1778. [Google Scholar] [CrossRef] [PubMed]
- Aw, D.; Silva, A.B.; Palmer, D.B. Immunosenescence: Emerging challenges for an ageing population. Immunology 2007, 120, 435–446. [Google Scholar] [CrossRef] [PubMed]
- Plowden, J.; Renshaw-Hoelscher, M.; Engleman, C.; Katz, J.; Sambhara, S. Innate immunity in aging: Impact on macrophage function. Aging Cell 2004, 3, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Lloberas, J.; Celada, A. Effect of aging on macrophage function. Exp. Gerontol. 2002, 37, 1325–1331. [Google Scholar] [CrossRef]
- Morey, J.N.; Boggero, I.A.; Scott, A.B.; Segerstrom, S.C. Current directions in stress and human immune function. Curr. Opin. Psychol. 2015, 5, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Organization, W.H. Mental health: Facing the challenges, building solutions. In Proceedings of the First WHO European Ministerial Conference on Mental Health, Helsinki, Finland, 12–15 January 2005. [Google Scholar]
- Corazon, S.; Nyed, P.; Sidenius, U.; Poulsen, D.; Stigsdotter, U. A Long-Term Follow-Up of the Efficacy of Nature-Based Therapy for Adults Suffering from Stress-Related Illnesses on Levels of Healthcare Consumption and Sick-Leave Absence: A Randomized Controlled Trial. Int. J. Environ. Res. Public Health 2018, 15, 137. [Google Scholar] [CrossRef] [PubMed]
- Dhabhar, F.S. Effects of stress on immune function: The good, the bad, and the beautiful. Immunol. Res. 2014, 58, 193–210. [Google Scholar] [CrossRef] [PubMed]
- Godbout, J.P.; Glaser, R. Stress-induced immune dysregulation: Implications for wound healing, infectious disease and cancer. J. Neuroimmune Pharmacol. 2006, 1, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Steptoe, A.; Hamer, M.; Chida, Y. The effects of acute psychological stress on circulating inflammatory factors in humans: A review and meta-analysis. Brain Behav. Immun. 2007, 21, 901–912. [Google Scholar] [CrossRef] [PubMed]
- Glaser, R.; MacCallum, R.C.; Laskowski, B.F.; Malarkey, W.B.; Sheridan, J.F.; Kiecolt-Glaser, J.K. Evidence for a shift in the Th-1 to Th-2 cytokine response associated with chronic stress and aging. J. Gerontol. A. Biol. Sci. Med. Sci. 2001, 56, M477–M482. [Google Scholar] [CrossRef] [PubMed]
- Veru, F.; Dancause, K.; Laplante, D.P.; King, S.; Luheshi, G. Prenatal maternal stress predicts reductions in CD4+ lymphocytes, increases in innate-derived cytokines, and a Th2 shift in adolescents: Project Ice Storm. Physiol. Behav. 2015, 144, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Dhabhar, F.S.; Malarkey, W.B.; Neri, E.; McEwen, B.S. Stress-induced redistribution of immune cells—From barracks to boulevards to battlefields: A tale of three hormones—Curt Richter Award Winner. Psychoneuroendocrinology 2012, 37, 1345–1368. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hydrolysate/Peptide | Enzyme/Treatment | Immune Effects | Species | Reference |
|---|---|---|---|---|
| Casein hydrolysate diet (200 g/kg casein, TD99482, Harlan-Teklad Custom Research, Madison, WI, USA | Not applicable (NA) | Reduction of autoimmune diabetes by 50%, decreased lactulose/mannitol ratio, decreased serum zonulin levels, increased ileal TEER, altered ileal mRNA expression of Myo9b, claudin-1, and claudin-2. | Diabetes prone BB rat | [19] |
| Casein hydrolysate (20% of diet Pancase STM (Sensient Flavours, Strassbourg, France) or NutramigenTM (Mead Johnson Nutrition, Zeeland, MI, USA)) | NA | Reduction of autoimmune diabetes, decreased lactulose/mannitol ratio, increased ileal IL-10 levels, beneficial gut microbiota changes (increased Lactobacilli and reduced Bacteroides spp. levels) | Diabetes prone BB rat | [20] |
| β-CN(94-123) from commercial yoghurt | NA | Enhanced numbers of goblet and Paneth cells in the small intestine, increased expression of Muc2, Muc4, lysozyme, and rdefa5. | rat | [21] |
| Yoghurt or Milk Fermented by Lactobacillus casei DN-114 001 | Lactobacillus casei DN-114 001 | Increased cell proliferation and villous area in the proximal intestine, hypertrophy and hyperplasia of Paneth and goblet cells. | mouse | [22] |
| Milk fermentation products of L. Helveticus R389 | L. Helveticus R389 | Enhanced expression of calcineurin in the small intestine, upregulated IL-2 and TNF production, increased number of mucosal mast cells and goblet cells | mouse | [23] |
| Egg yolk digests | Pepsin | Increase of the IL-6 secretion by small intestinal epithelial cells, increase in IgA+ cells, orchestrating the Th1/Th2 response. | mouse | [24] |
| Common carp egg hydrolysate | Pepsin, alcalase | Increase of secretory immunoglobulin A in the gut. Pepsin hydrolysate increased the splenic NK cell cytotoxicity, macrophage phagocytosis and level of serum immunoglobulin A (IgA). S-IgA in the gut was significantly enhanced by pepsin and alcalase hydrolysates. Trypsin hydrolysate increased the percentages of CD4+ and CD8+ cells in the spleen. | mouse | [25] |
| Yellow field pea hydrolysate | Thermolysin | Increased number of IgA+ cells in the small intestine lamina propria, accompanied by an increase in the number of IL-4+, IL-10+, and IFNγ + cells. | mouse | [26] |
| Fermented pacific whiting protein | Yeast | Enhanced phagocytic activity of peritoneal macrophages, increased number of IgA+ cells, and increased IL-4, IL-6, IL-10, IFNγ, and TNFα levels in the small intestine lamina propria | mouse | [27] |
| Shark protein hydrolysate PeptibalTM (innoVactiv Inc) | Trypsin and chymotrypsin | Increase of small intestinal immunoglobulin A-producing cells and intestinal IL-6, TNFα, TGFβ, and IL-10 | mouse | [28] |
| Peptide fraction from Lactobacillus helveticus-Fermented Milk | Lactobacillus Helveticus | Increased intestinal and serum IgA levels, increase in the number of IgA-secreting B lymphocytes in the intestinal lamina propria, stimulation of Th2 response (IL-4 vs. IFNγ) | mouse | [29] |
| κ-casein–derived glycomacropeptide | NA | Decreased body weight loss, decreased anorexia, colonic damage, a reduction in colonic alkaline phosphatase activity, IL-1, trefoil factor 3, and iNOS mRNA levels. | Rat (TNBS induced colitis) | [30] |
| β-Casein hydrolysate | Cell envelope-associated proteinase of Lactobacillus delbrueckii ssp. lactis CRL 581 | Decreased mortality rates, faster recovery of initial body weight loss, less microbial translocation to the liver, decreased β-glucuronidase and myeloperoxidase activities in the gut, decreased colonic macroscopic and microscopic damage, increased IL-10 and decreased IFNγ. | Mouse (TNBS induced colitis) | [31] |
| κ-casein–derived glycomacropeptide | NA | Rag1-/-:increased body-weight gain, decreased colonic damage score and myeloperoxidase (MPO) activity, reduced percentage of CD4+ interferon IFNγ+ cells and increased IL-6 in MLN. Increased colonic expression of TNFα and IFNγ and increased IL-10 in MLN, by MLN. DSS: decreased MPO activity, increased IL-10 production in MLN. | Mouse (DSS induced colitis and Rag1-/-) | [32] |
| bovine glycomacropeptide | NA | Decrease of inflammatory injury, as assessed by lower extension of necrosis and damage score, myeloperoxidase, alkaline phosphatase, inducible nitric oxide synthase, IL- 1β, TNFα, and IL-17. | Rat (TNBS induced colitis) | [33] |
| Egg white hydrolysate | Aminopeptidase | Attenuated DSS-induced clinical symptoms, including weight loss, mucosal and submucosal inflammation, crypt distortion, and colon muscle thickening, and decreased intestinal permeability and increased mucin gene expression, reduced intestinal expression of pro-inflammatory cytokines TNFα, IL-6, IL-1β, IFNγ, IL-8, and IL-17. | Pig (DSS induced colitis) | [34] |
| Soybean protein hydrolysate | Rhizopus oryzae neutral protease preparation | Increased number of IL-12+CD11b+ in spleens, increased cytotoxic activity of spleen cells, increased Igh-4, Aqp8, Dmbt1, Slpi, and Mx1 in Peyer’s patch cells. | Mouse | [35] |
| Partially hydrolyzed whey protein | NA | Increased Breg and Treg in the spleen, increased IgA+ B-cells in the MLN, increased Th1, activated Treg and activated Th17 cells in the Peyer’s patches | Mouse | [36] |
| LLDAQSAPLRVYVEELKP (from whey) | NA | Reduced acute allergic skin response, decreased whey-specific antibody levels, increased the percentages of CD11b+CD103+ dendritic cells and CD25+Foxp3+ T cells in the MLN. | Mouse | [37] |
| Partial whey hydrolysate | NA | Reduced acute allergic skin response and mast cell degranulation after whey challenge, increased Foxp3+ regulatory T-cell numbers in the MLN. | Mouse | [38] |
| oyster peptide-based enteral nutrition formula | Bromelain, pepsin, trypsin | Enhanced spleen lymphocyte proliferation and of NK cell activity | Mouse | [39] |
| Casein hydrolysate | Trypsin | Phagocytosing capacity of phagocytic cells was increased | Mouse | [17] |
| Milk protein hydrolysate | Improved the level of hemolysin in serum, and enhanced phagocytosis of macrophages. In ovalbumin-sensitized mice, the milk protein hydrolysates reduced IgE levels, reduced IL-4 in serum, reduced the release of histamine and bicarbonate in peritoneal mast cells, and enhanced TGFβ levels. | ICR mouse | [40] | |
| Chum salmon oligopeptide preparation | Complex protease | Enhanced lymphocyte proliferation capacity increased number of plaque-forming cells, increased NK cell activity, increased percentage of CD4+ T helper (Th) cells in spleen and secretion of Th1 (IL-2, IFNγ) and Th2 (IL-5, IL-6) type cell cytokines. | ICR mouse | [41] |
| Tuna cooking drip hydrolysate | Enzyme A and B | Increased weight of the spleen and thymus and enhanced the proliferation of splenocytes. Increased production of IL-10 and IL-2. Increased serum IgG1 and IgG2a levels. | Mouse | [42] |
| Soy protein hydrolysate | Pepsin | Increased serum IgA and IgG levels | Rat | [7] |
| Soy protein hydrolysate | Theroase, bioprase, Sumizyme FP | Total lymphocyte and granulocyte numbers were altered, and the numbers of CD11b+ cells and CD56+ cells increased. | Human | [43] |
| Wheat gluten hydrolysate | NA | NK cell activity increased significantly | Human | [44] |
| Fish protein hydrolysate (Amizate) | NA | No effects observed | Human | [45] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kiewiet, M.B.G.; Faas, M.M.; De Vos, P. Immunomodulatory Protein Hydrolysates and Their Application. Nutrients 2018, 10, 904. https://doi.org/10.3390/nu10070904
Kiewiet MBG, Faas MM, De Vos P. Immunomodulatory Protein Hydrolysates and Their Application. Nutrients. 2018; 10(7):904. https://doi.org/10.3390/nu10070904
Chicago/Turabian StyleKiewiet, Mensiena B. G., Marijke M. Faas, and Paul De Vos. 2018. "Immunomodulatory Protein Hydrolysates and Their Application" Nutrients 10, no. 7: 904. https://doi.org/10.3390/nu10070904
APA StyleKiewiet, M. B. G., Faas, M. M., & De Vos, P. (2018). Immunomodulatory Protein Hydrolysates and Their Application. Nutrients, 10(7), 904. https://doi.org/10.3390/nu10070904