Comparison of the Incorporation of DHA in Circulatory and Neural Tissue When Provided as Triacylglycerol (TAG), Monoacylglycerol (MAG) or Phospholipids (PL) Provides New Insight into Fatty Acid Bioavailability

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Diets
2.2. Experimental Design
2.3. Sample Preparation
2.4. Fatty acid Methyl Esters Preparation and Analysis by Gas Chromatography
2.5. Statistical Analyses
3. Results
3.1. Food Intake, Body Weight Gain and Body Composition
3.2. Incorporation of DHA in Erythrocytes, Plasma, Retina and Brain Tissues
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Arterburn, L.M.; Hall, E.B.; Oken, H. Distribution, interconversion, and dose response of n-3 fatty acids in humans. Am. J. Clin. Nutr. 2006, 83, 1467S–1476S. [Google Scholar] [CrossRef] [PubMed]
- Gibson, R.A.; Muhlhausler, B.; Makrides, M. Conversion of linoleic acid and alpha-linolenic acid to long-chain polyunsaturated fatty acids (LCPUFAs), with a focus on pregnancy, lactation and the first 2 years of life. Matern. Child Nutr. 2011, 7 (Suppl. 2), 17–26. [Google Scholar] [CrossRef] [PubMed]
- Burdge, G.C.; Wootton, S.A. Conversion of α-linolenic acid to eicosapentaenoic, docosapentaenoic and docosahexaenoic acids in young women. Br. J. Nutr. 2002, 88, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Kris-Etherton, P.M.; Harris, W.S.; Appel, L.J. Fish consumption, fish oil, omega-3 fatty acids, and cardiovascular disease. Circulation 2002, 106, 2747–2757. [Google Scholar] [CrossRef] [PubMed]
- Neuringer, M.; Anderson, G.J.; Connor, W.E. The essentiality of N-3 fatty acids for the development and function of the retina and brain. Annu. Rev. Nutr. 1988, 8, 517–541. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. n-3 fatty acids, inflammation and immunity: New mechanisms to explain old actions. Proc. Nutr. Soc. 2013, 72, 326–336. [Google Scholar] [CrossRef] [PubMed]
- Gil, Á. Polyunsaturated fatty acids and inflammatory diseases. Biomed. Pharmacother. 2002, 56, 388–396. [Google Scholar] [CrossRef]
- Swanson, D.; Block, R.; Mousa, S.A. Omega-3 Fatty Acids EPA and DHA: Health Benefits Throughout Life. Adv. Nutr. 2012, 3, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.; Zirpoli, H.; Qi, K. n-3 fatty acids modulate adipose tissue inflammation and oxidative stress. Curr. Opin. Clin. Nutr. Metab. Care 2013, 16, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. The role of marine omega-3 (n-3) fatty acids in inflammatory processes, atherosclerosis and plaque stability. Mol. Nutr. Food Res. 2012, 56, 1073–1080. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, V.C.; Hassing, M.R.; Lewandowski, P.A. Marine polyunsaturated fatty acids and cancer therapy. Br. J. Cancer 2013, 108, 486–492. [Google Scholar] [CrossRef] [PubMed]
- Lorente-Cebrián, S.; Pérez-Matute, P.; Martínez, J.A.; Marti, A.; Moreno-Aliaga, M.J. Effects of eicosapentaenoic acid (EPA) on adiponectin gene expression and secretion in primary cultured rat adipocytes. J. Physiol. Biochem. 2006, 62, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Querques, G.; Forte, R.; Souied, E.H. Retina and omega-3. J. Nutr. Metab. 2011, 2011, 748361. [Google Scholar] [CrossRef] [PubMed]
- Amate, L.; Ramírez, M.; Gil, A. Positional analysis of triglycerides and phospholipids rich in long-chain polyunsaturated fatty acids. Lipids 1999, 34, 865–871. [Google Scholar] [CrossRef] [PubMed]
- Hayes, K.C. Synthetic and modified glycerides: Effects on plasma lipids. Curr. Opin. Lipidol. 2001, 12, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Anderson, B.M.; Ma, D.W. Are all n-3 polyunsaturated fatty acids created equal? Lipids Health Dis. 2009, 8, 33–53. [Google Scholar] [CrossRef] [PubMed]
- Mu, H.; Høy, C.E. The digestion of dietary triacylglycerols. Prog. Lipid Res. 2004, 43, 105–133. [Google Scholar] [CrossRef]
- Christensen, M.S.; Høy, C.E.; Becker, C.C.; Redgrave, T.G. Intestinal absorption and lymphatic transport of eicosapentaenoic (EPA), docosahexaenoic (DHA), and decanoic acids: Dependence on intramolecular triacylglycerol structure. Am. J. Clin. Nutr. 1995, 61, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Hernandez, C.; Thakkar, S.K.; Moulin, J.; Oliveira, M.; Masserey-Elmelegy, I.; Dionisi, F.; Destaillats, F. Benefits of structured and free monoacylglycerols to deliver eicosapentaenoic (EPA) in a model of lipid malabsorption. Nutrients 2012, 4, 1781–1793. [Google Scholar] [CrossRef] [PubMed]
- Banno, F.; Doisaki, S.; Shimizu, N.; Fujimoto, K. Lymphatic absorption of docosahexaenoic acid given as monoglyceride, diglyceride, triglyceride, and ethyl ester in rats. J. Nutr. Sci. Vitaminol. 2002, 48, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Hansen, J.B.; Grimsgaard, S.; Nilsen, H.; Nordøy, A.; Bønaa, K.H. Effects of highly purified eicosapentaenoic acid and docosahexaenoic acid on fatty acid absorption, incorporation into serum phospholipids and postprandial triglyceridemia. Lipids 1998, 33, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Offman, E.; Marenco, T.; Ferber, S.; Johnson, J.; Kling, D.; Curcio, D.; Davidson, M. Steady-state bioavailability of prescription omega-3 on a low-fat diet is significantly improved with a free fatty acid formulation compared with an ethyl ester formulation: The ECLIPSE II study. Vasc. Health Risk Manag. 2013, 9, 563–573. [Google Scholar] [CrossRef] [PubMed]
- Maki, K.C.; Keane, W.F.; Bobotas, G. Effects of MAT9001, an Omega-3 Fatty Acid Drug, Compared with Eicosapentaenoic Acid Ethyl Esters, on Triglycerides, Lipoprotein Cholesterol and Related Variables in Hypertriglyceridemic Subjects. In Proceedings of the National Lipid Association (NLA) Scientific Sessions, Chicago, IL, USA, 11–14 June 2015. [Google Scholar]
- Hedengran, A.; Szecsi, P.B.; Dyerberg, J.; Harris, W.S.; Stender, S. n-3 PUFA esterified to glycerol or as ethyl esters reduce non-fasting plasma triacylglycerol in subjects with hypertriglyceridemia: A randomized trial. Lipids 2015, 50, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Innis, S.M.; Dyer, R. Dietary triacylglycerols with palmitic acid (16:0) in the 2-position increase 16:0 in the 2-position of plasma and chylomicron triacylglycerols, but reduce phospholipid arachidonic and docosahexaenoic acids, and alter cholesteryl ester metabolism in formula-Fed piglets. J. Nutr. 1997, 127, 1311–1319. [Google Scholar] [PubMed]
- Valenzuela, A.; Valenzuela, V.; Sanhueza, J.; Nieto, S. Effect of supplementation with docosahexaenoic acid ethyl ester and sn-2 docosahexaenyl monoacylglyceride on plasma and erythrocyte fatty acids in rats. Ann. Nutr. Metab. 2005, 49, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Hernandez, C.; Destaillats, F.; Thakkar, S.K.; Goulet, L.; Wynn, E.; Grathwohl, D.; Roessle, C.; de Giorgi, S.; Tappy, L.; Giuffrida, F.; et al. Monoacylglycerol-enriched oil increases EPA/DHA delivery to circulatory system in humans with induced lipid malabsorption conditions. J. Lipid Res. 2016, 57, 2208–2216. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Hernandez, C.; Thakkar, S.K.; Masserey-Elmelegy, I.; Buosi, W.; Fontannaz, P.; Giuffrida, F. Quantification of fatty acids in erythrocytes and plasma by fast gas chromatography. J. Sep. Sci. 2017, 40, 3289–3300. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Hernandez, C.; Oliveira, M.; Pescia, G.; Moulin, J.; Masserey-Elmelegy, I.; Dionisi, F.; Destaillats, F. Lipase inhibitor orlistat decreases incorporation of eicosa-pentaenoic and docosahexaenoic acids in rat tissues. Nutr. Res. 2010, 30, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Destaillats, F.; Cruz-Hernandez, C. Fast analysis by gas-liquid chromatography perspective on the resolution of complex fatty acid compositions. J. Chromatogr. A 2007, 169, 175–178. [Google Scholar] [CrossRef] [PubMed]
- Borgström, B. Importance of phospholipids, pancreatic phospholipase A2, and fatty acid for the digestion of dietary fat. Gastroenterology 1980, 78, 954–962. [Google Scholar] [PubMed]
- Zierenberg, O.; Grundy, S.M. Intestinal absorption of polyenephosphatidylcholine in man. J. Lipid Res. 1982, 23, 1136–1142. [Google Scholar] [PubMed]
- Ghasemifard, S.; Hermon, K.; Turchini, G.M.; Sinclair, A.J. Metabolic fate (absorption, β-oxidation and deposition) of long-chain n-3 fatty acids is affected by sex and by the oil source (krill oil or fish oil) in the rat. Br. J. Nutr. 2015, 114, 684–692. [Google Scholar] [CrossRef] [PubMed]
- Amate, L.; Gil, A.; Ramírez, M. Feeding infant piglets formula with long-chain polyunsaturated fatty acids as triacylglycerols or phospholipids influences the distribution of these fatty acids in plasma lipoprotein fractions. J. Nutr. 2001, 131, 1250–1255. [Google Scholar] [CrossRef] [PubMed]
- Fujita, Y.; Matsuoka, H.; Hirooka, K. Regulation of fatty acid metabolism in bacteria. Mol. Microbiol. 2007, 66, 829–839. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.J.; Yu, J.G.; Lee, I.A.; Liu, M.J.; Shen, Y.F.; Sharma, S.P.; Jamal, M.A.H.M.; Yoo, J.H.; Kim, H.J.; Hong, S.T. Intestinal removal of free fatty acids from hosts by Lactobacilli for the treatment of obesity. FEBS OpenBio 2016, 6, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Ulven, S.M.; Kirkhus, B.; Lamglait, A.; Basu, S.; Elind, E.; Haider, T.; Berge, K.; Vik, H.; Pedersen, J.I. Metabolic effects of krill oil are essentially similar to those of fish oil but at lower dose of EPA and DHA, in healthy volunteers. Lipids 2011, 46, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Ghasemifard, S.; Turchini, G.M.; Sinclair, A.J. Omega-3 long chain fatty acid “bioavailability”: A review of evidence and methodological considerations. Prog. Lipid Res. 2014, 56, 92–108. [Google Scholar] [CrossRef] [PubMed]
- Stevens, L.; Zhang, W.; Peck, L.; Kuczek, T.; Grevstad, N.; Mahon, A.; Zentall, S.S.; Arnold, L.E.; Burgess, J.R. EFA supplementation in children with inattention, hyperactivity, and other disruptive behaviors. Lipids 2003, 38, 1007–1021. [Google Scholar] [CrossRef] [PubMed]
- Hirayama, S.; Hamazaki, T.; Terasawa, K. Effect of docosahexaenoic acid-containing food administration on symptoms of attention-deficit/hyperactivity disorder—A placebo-controlled double-blind study. Eur. J. Clin. Nutr. 2004, 58, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Wijendran, V.; Huang, M.; Diau, G.; Boehm, G.; Nathanielsz, P.W.; Brenna, J.T. Efficacy of dietary arachidonic acid provided as triglyceride or phospholipid as substrates for brain arachidonic acid accretion in baboon neonates. Pediatr. Res. 2002, 51, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Piscitelli, F.; Carta, G.; Bisogno, T.; Murru, E.; Cordeddu, L.; Berge, K.; Tandy, S.; Cohn, J.S.; Griinari, M.; Banni, S.; et al. Effect of dietary krill oil supplementation on the endocannabinoidome of metabolically relevant tissues from high-fat-fed mice. Nutr. Metab. 2011, 8, 51–67. [Google Scholar] [CrossRef] [PubMed]
- Delion, S.; Chalon, S.; Hérault, J.; Guilloteau, D.; Besnard, J.C.; Durand, G. Chronic dietary alpha-linolenic acid deficiency alters dopaminergic and serotoninergic neurotransmission in rats. J. Nutr. 1994, 124, 2466–2476. [Google Scholar] [CrossRef] [PubMed]
- Jumpsen, J.; Lien, E.L.; Goh, Y.K.; Clandinin, M.T. Small changes of dietary (n-6) and (n-3)/fatty acid content ration alter phosphatidylethanolamine and phosphatidylcholine fatty acid composition during development of neuronal and glial cells in rats. J. Nutr. 1997, 127, 724–731. [Google Scholar] [CrossRef] [PubMed]
- Carrié, I.; Clément, M.; de Javel, D.; Francès, H.; Bourre, J.M. Specific phospholipid fatty acid composition of brain regions in mice. Effects of n-3 polyunsaturated fatty acid deficiency and phospholipid supplementation. J. Lipid Res. 2000, 41, 465–472. [Google Scholar] [PubMed]
- Westerberg, A.C.; Schei, R.; Henriksen, C.; Smith, L.; Veierød, M.B.; Drevon, C.A.; Iversen, P.O. Attention among very low birth weight infants following early supplementation with docosahexaenoic and arachidonic acid. Acta Paediatr. 2011, 100, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Sabel, K.G.; Strandvik, B.; Petzold, M.; Lundqvist-Persson, C. Motor, mental and behavioral developments in infancy are associated with fatty acid pattern in breast milk and plasma of premature infants. Prostaglandins Leukot. Essent. Fatty Acids 2012, 86, 183–188. [Google Scholar] [CrossRef] [PubMed]
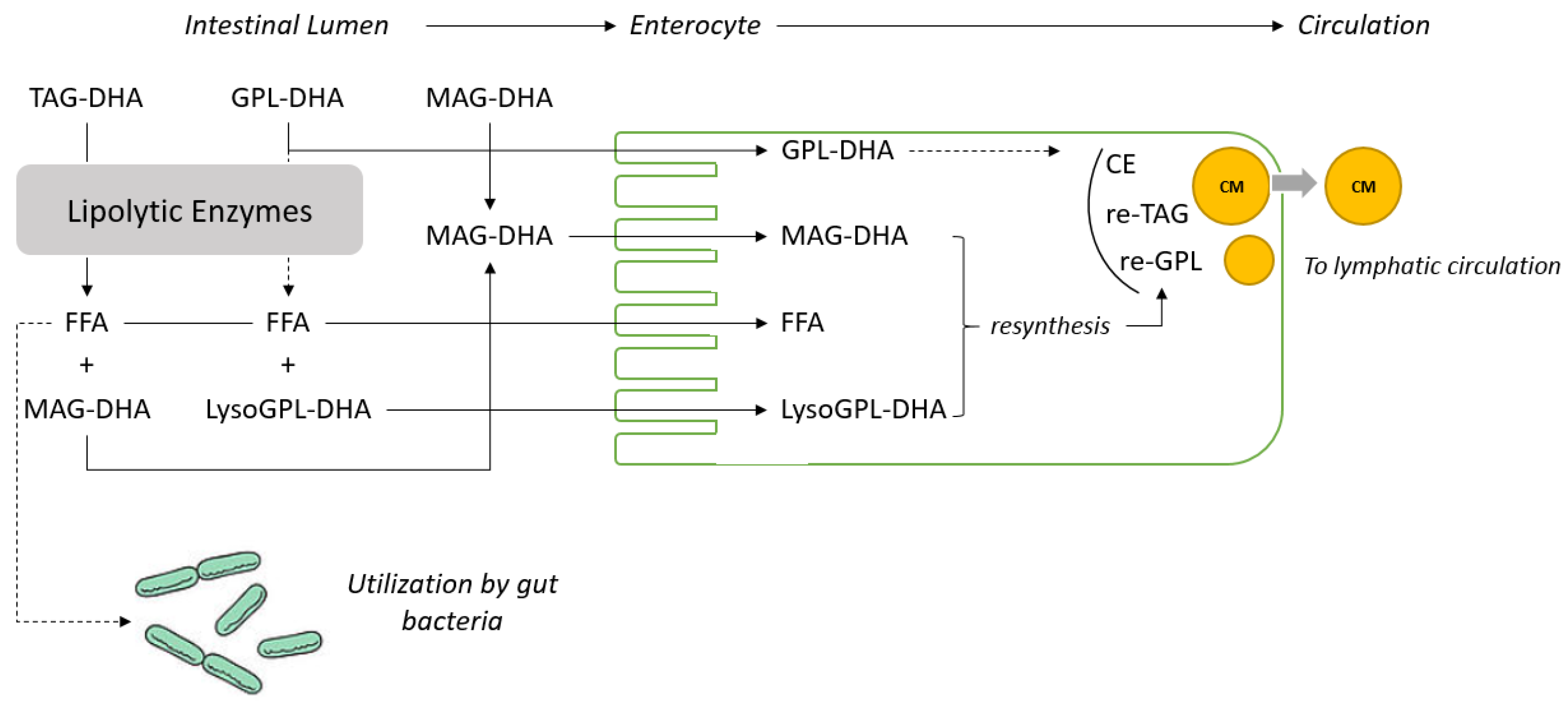
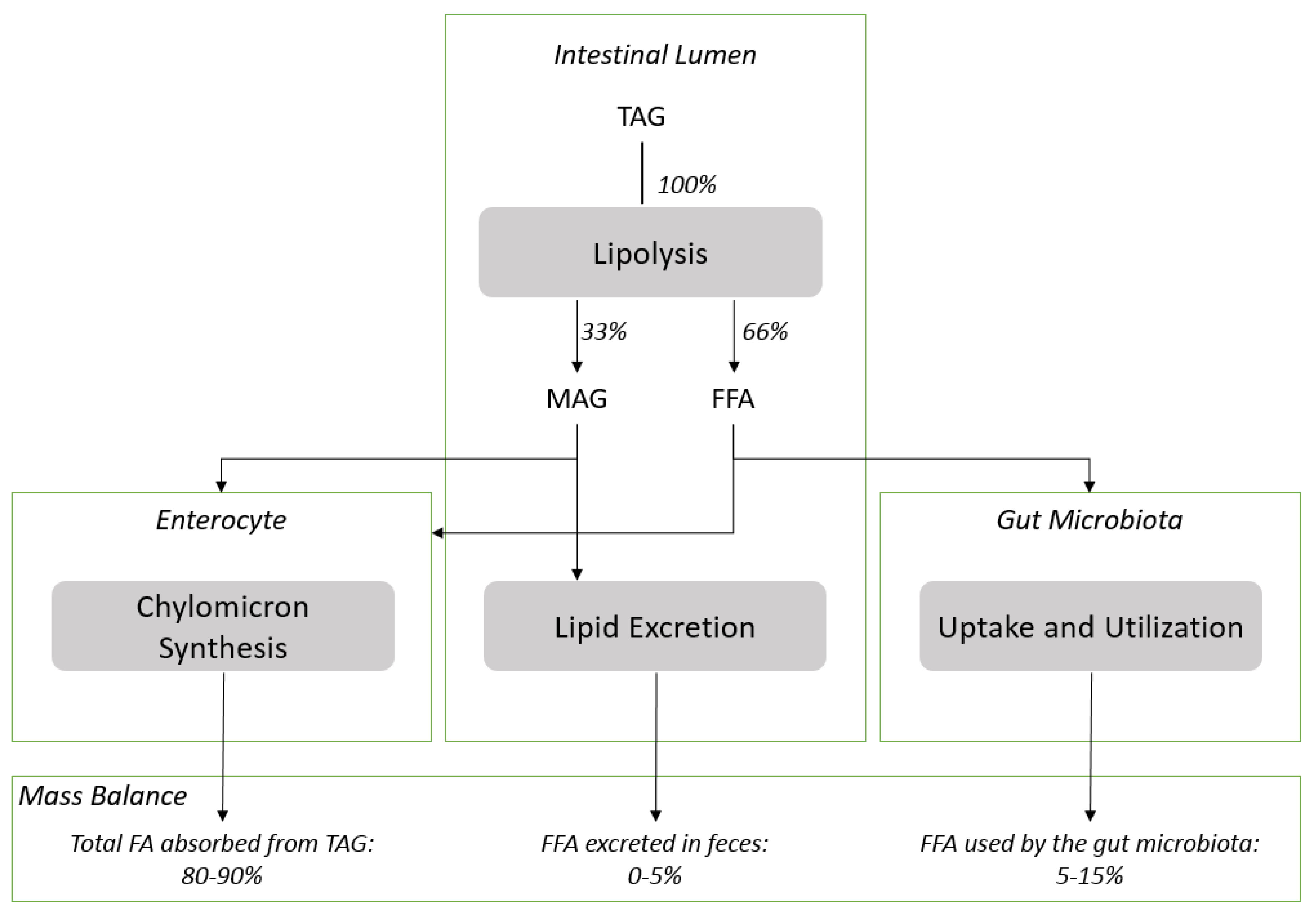

{kind=link}
{kind=link}
| Composition | Control | TAG-DHA | MAG-DHA | PL-DHA |
|---|---|---|---|---|
| Total lipid content | 20.00 | 20.00 | 20.00 | 20.00 |
| Cocoa butter | 3.00 | 1.60 | 2.20 | 1.70 |
| Soybean oil | 11.00 | 11.20 | 11.00 | 10.00 |
| Sunflower oil | 6.00 | 6.46 | 6.00 | 5.20 |
| Oil DHASCO (TAG-DHA) | 0.76 | |||
| sn-1(3)-Monoacylglycerol (MAG-DHA) | 0.74 | |||
| GPL8-DHA (PL-DHA) | 3.10 | |||
| Corn starch | 46.10 | 46.10 | 46.10 | 46.10 |
| κ-Caseinate | 14.00 | 14.00 | 14.00 | 14.00 |
| Sucrose | 10.00 | 10.00 | 10.00 | 10.00 |
| Cellulose | 5.00 | 5.00 | 5.00 | 5.00 |
| Mineral mix AIN-93M | 3.50 | 3.50 | 3.50 | 3.50 |
| Vitamin mix AIN 93VX | 1.00 | 1.00 | 1.00 | 1.00 |
| l-cystine | 0.18 | 0.18 | 0.18 | 0.18 |
| Choline bitartrate | 0.25 | 0.25 | 0.25 | 0.25 |
| Butylhydroxytoluene (BHT) | 0.0008 | 0.0008 | 0.0008 | 0.0008 |
| Composition | Control | TAG-DHA | MAG-DHA | PL-DHA |
|---|---|---|---|---|
| 14:0 | 0.02 | 0.08 | 0.05 | 0.02 |
| 16:0 | 1.81 | 2.02 | 2.09 | 2.16 |
| 16:1 n-7 | 0.02 | 0.03 | 0.03 | 0.04 |
| 17:0 | 0.04 | 0.04 | 0.04 | 0.04 |
| 18:0 | 1.44 | 1.61 | 1.66 | 1.17 |
| 18:1 n-9 | 4.52 | 5.10 | 5.16 | 4.64 |
| 18:2 n-6 | 6.87 | 7.47 | 7.59 | 6.95 |
| 18:3 n-3 | 0.48 | 0.52 | 0.53 | 0.47 |
| 20:0 | 0.04 | 0.08 | 0.08 | 0.06 |
| 20:1 n-9 | 0.03 | 0.04 | 0.03 | 0.04 |
| 20:5 n-3 | 0.08 | 0.09 | 0.09 | 0.08 |
| 22:1 n-9 | 0.00 | 0.00 | 0.00 | 0.00 |
| 22:6 n-3 | 0.00 | 0.29 | 0.31 | 0.34 |
| 24:0 | 0.03 | 0.04 | 0.04 | 0.03 |
| Total n-3 FA | 0.56 | 0.90 | 0.93 | 0.90 |
| Parameter | Experimental Group | |||
|---|---|---|---|---|
| Control | TAG-DHA | MAG-DHA | PL-DHA | |
| Final body weight (g) | 388.1 ± 13.4 | 398.8 ± 28.5 | 395.9 ± 23.6 | 421.04 ± 39.1 |
| Fat Mass d 0 (g) | 21.2 ± 3.6 | 21.8 ± 3.9 | 21.6 ± 3.3 | 22.2 ± 3.3 |
| d 60 (g) | 65.1 ± 12.7 | 77.5 ± 13.6 | 65.5 ± 14.5 | 79.7 ± 28.7 |
| Lean Mass d 0 (g) | 146.3 ± 7.9 | 146.5 ± 5.8 | 146.7 ± 6.9 | 145.8 ± 6.4 |
| d 60 (g) | 272.7 ± 16.7 | 270.6 ± 21.5 | 280.1 ± 13.7 | 288.0 ± 12.7 |
| Daily Food Intake (g/d) | 27.5 ± 10.1 | 25.3 ± 8.8 | 27.5 ± 7.2 | 30.6 ± 10.1 * |
| Group | Feeding Time (days) | ||||
|---|---|---|---|---|---|
| 0 | 14 | 28 | 35 | 49 | |
| Control | 44.2 ± 12.2 | 40.7 ± 6.0 | 51.0 ± 11.4 | 41.4 ± 10.0 | 54.2 ± 30.2 |
| TAG-DHA | 46.1 ± 14.0 | 58.2 ± 12.1 ** | 78.5 ± 13.8 *** | 71.5 ± 15.4 *** | 78.2 ± 14.9 *** |
| PL-DHA | 42.3 ± 15.9 | 77.6 ± 17.4 ***,§,! | 99.8 ± 19.2 ***,! | 103.1 ± 15.5 ***,! | 138.4 ± 71.4 ***,§,! |
| MAG-DHA | 52.3 ± 9.7 | 64.4 ± 11.0 *** | 98.8 ± 22.3 ***,! | 94.1 ± 17.7 ***,! | 105.8 ± 22.3 ***,! |
| Group | Feeding Time (days) | ||||
|---|---|---|---|---|---|
| 0 | 14 | 28 | 35 | 49 | |
| Control | 42.1 ± 21.6 | 34.0 ± 7.9 | 29.7 ± 9.4 | 26.4 ± 8.6 | 32.2 ± 10.3 |
| TAG-DHA | 32.4 ± 15.4 | 54.5 ± 16.5 *** | 48.9 ± 20.3 *** | 59.9 ± 21.0 *** | 75.3 ± 26.8 *** |
| PL-DHA | 36.0 ± 11.8 | 102.2 ± 25.9 ***,§,! | 68.8 ± 35.9 *** | 92.6 ± 14.7 ***,! | 102.7 ± 27.0 ***,§,! |
| MAG-DHA | 48.7 ± 14.6 | 66.5 ± 19.3 *** | 56.7 ± 17.8 *** | 73.6 ± 18.4 *** | 80.1 ± 22.5 *** |
| Tissue | Control | TAG-DHA | MAG-DHA | PL-DHA |
|---|---|---|---|---|
| Retina | 28.92 ± 2.7 | 32.84 ± 1.2 *** | 32.32 ± 1.4 *** | 32.77 ± 1.3 *** |
| Cortex | 12.75 ± 0.6 | 13.56 ± 0.7 *** | 13.41 ± 0.4 *** | 13.62 ± 0.5 *** |
| Hypothalamus | 10.65 ± 0.7 | 11.29 ± 0.8 ** | 11.27 ± 0.4 ** | 10.98 ± 0.5 |
| Hippocampus | 10.96 ± 1.3 | 11.89 ± 1.0 ** | 12.34 ± 0.7 *** | 12.42 ± 0.9 *** |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Destaillats, F.; Oliveira, M.; Bastic Schmid, V.; Masserey-Elmelegy, I.; Giuffrida, F.; Thakkar, S.K.; Dupuis, L.; Gosoniu, M.L.; Cruz-Hernandez, C. Comparison of the Incorporation of DHA in Circulatory and Neural Tissue When Provided as Triacylglycerol (TAG), Monoacylglycerol (MAG) or Phospholipids (PL) Provides New Insight into Fatty Acid Bioavailability. Nutrients 2018, 10, 620. https://doi.org/10.3390/nu10050620
Destaillats F, Oliveira M, Bastic Schmid V, Masserey-Elmelegy I, Giuffrida F, Thakkar SK, Dupuis L, Gosoniu ML, Cruz-Hernandez C. Comparison of the Incorporation of DHA in Circulatory and Neural Tissue When Provided as Triacylglycerol (TAG), Monoacylglycerol (MAG) or Phospholipids (PL) Provides New Insight into Fatty Acid Bioavailability. Nutrients. 2018; 10(5):620. https://doi.org/10.3390/nu10050620
Chicago/Turabian StyleDestaillats, Frédéric, Manuel Oliveira, Viktoria Bastic Schmid, Isabelle Masserey-Elmelegy, Francesca Giuffrida, Sagar K. Thakkar, Lénaïck Dupuis, Maria Laura Gosoniu, and Cristina Cruz-Hernandez. 2018. "Comparison of the Incorporation of DHA in Circulatory and Neural Tissue When Provided as Triacylglycerol (TAG), Monoacylglycerol (MAG) or Phospholipids (PL) Provides New Insight into Fatty Acid Bioavailability" Nutrients 10, no. 5: 620. https://doi.org/10.3390/nu10050620
APA StyleDestaillats, F., Oliveira, M., Bastic Schmid, V., Masserey-Elmelegy, I., Giuffrida, F., Thakkar, S. K., Dupuis, L., Gosoniu, M. L., & Cruz-Hernandez, C. (2018). Comparison of the Incorporation of DHA in Circulatory and Neural Tissue When Provided as Triacylglycerol (TAG), Monoacylglycerol (MAG) or Phospholipids (PL) Provides New Insight into Fatty Acid Bioavailability. Nutrients, 10(5), 620. https://doi.org/10.3390/nu10050620