Inhibitory Effects of Helianthus tuberosus Ethanol Extract on Dermatophagoides farina body-induced Atopic Dermatitis Mouse Model and Human Keratinocytes


Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
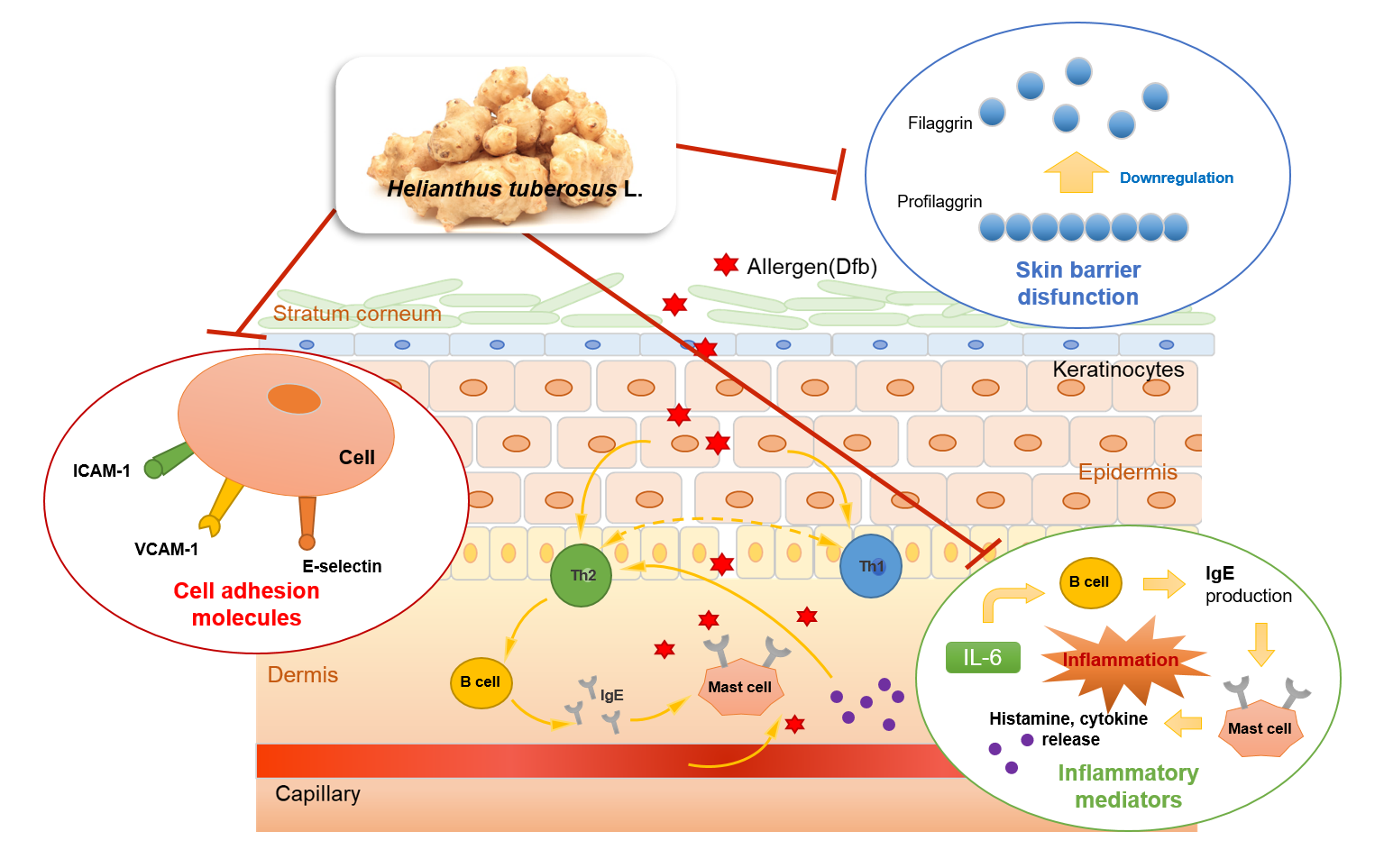
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Sample Preparation
2.3. Dfb-Induced AD Model
2.4. Scoring of Dermatitis Severity
2.5. Cytokine Assays
2.6. Histamine and IgE Assay
2.7. Histopathological and Immunohistochemical (IHC) Analysis
2.8. Cell Culture and Sample Treatment
2.9. Cell Viability Assay
2.10. Western Blot Analysis
2.11. Statistical Analysis
3. Results
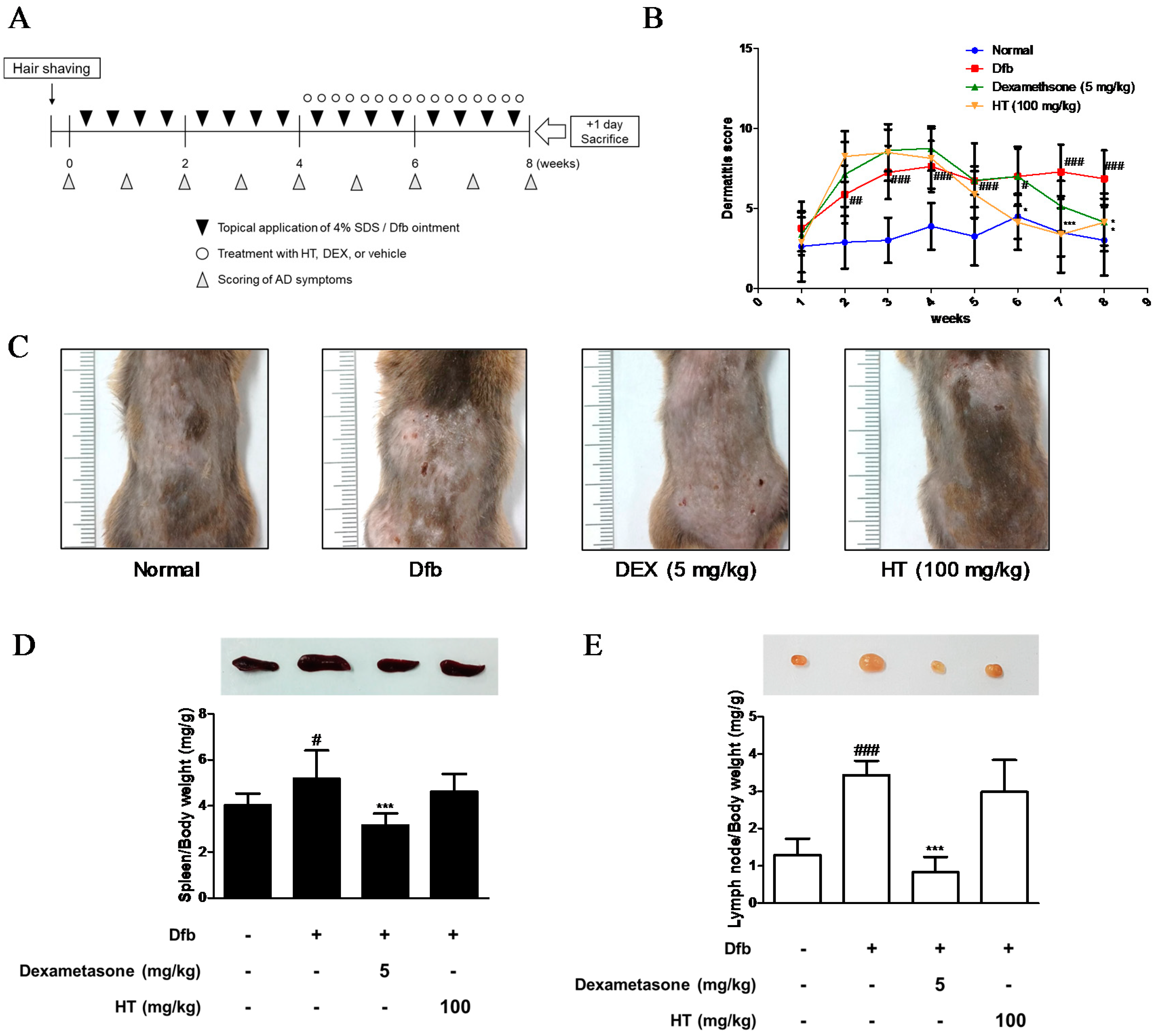
3.1. HT Attenuated Clinical Severity of AD Skin Symptoms in Dfb-Induced AD Mice
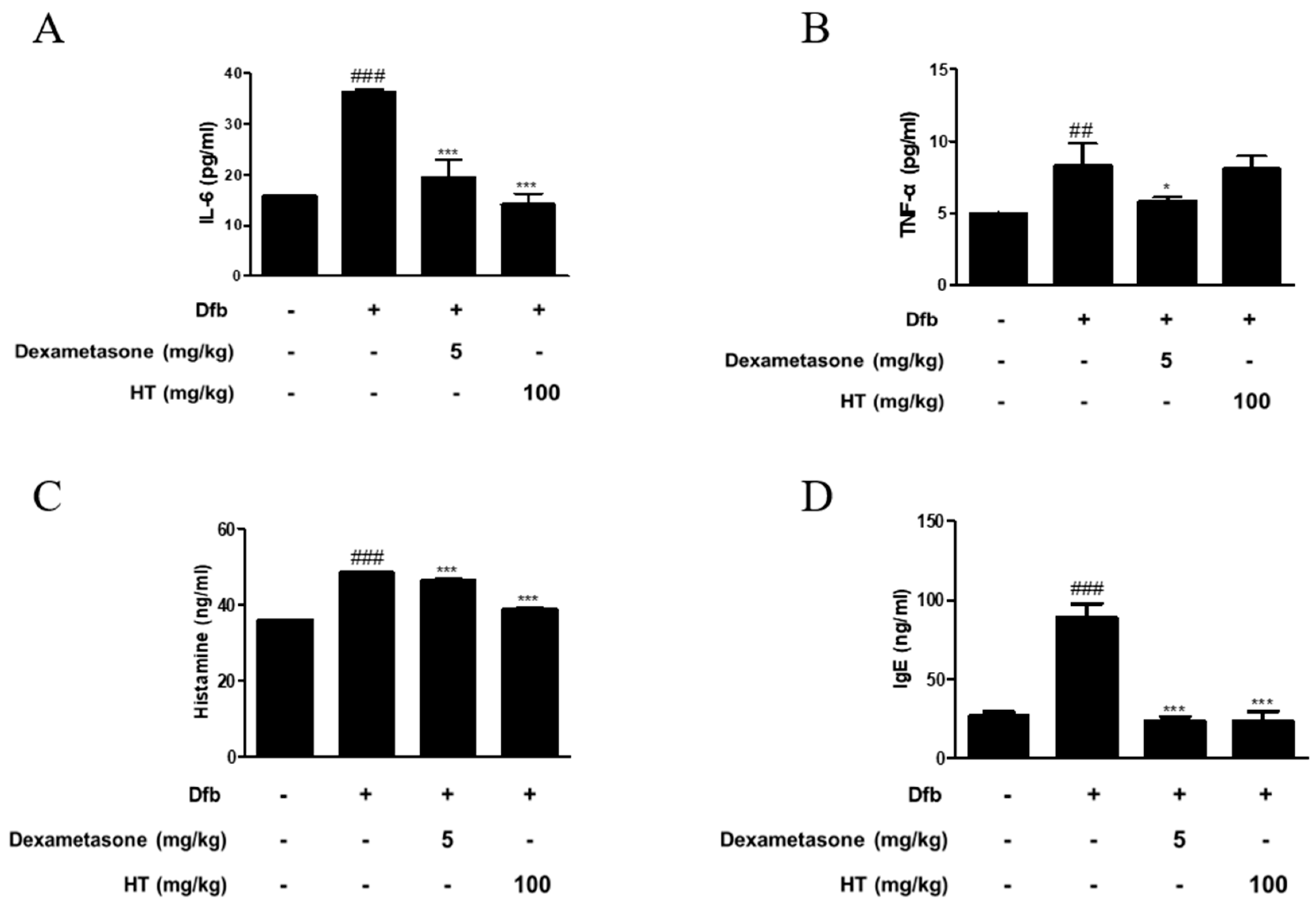
3.2. HT Inhibited the Production of Inflammatory Mediators in Dfb-Induced AD Mice
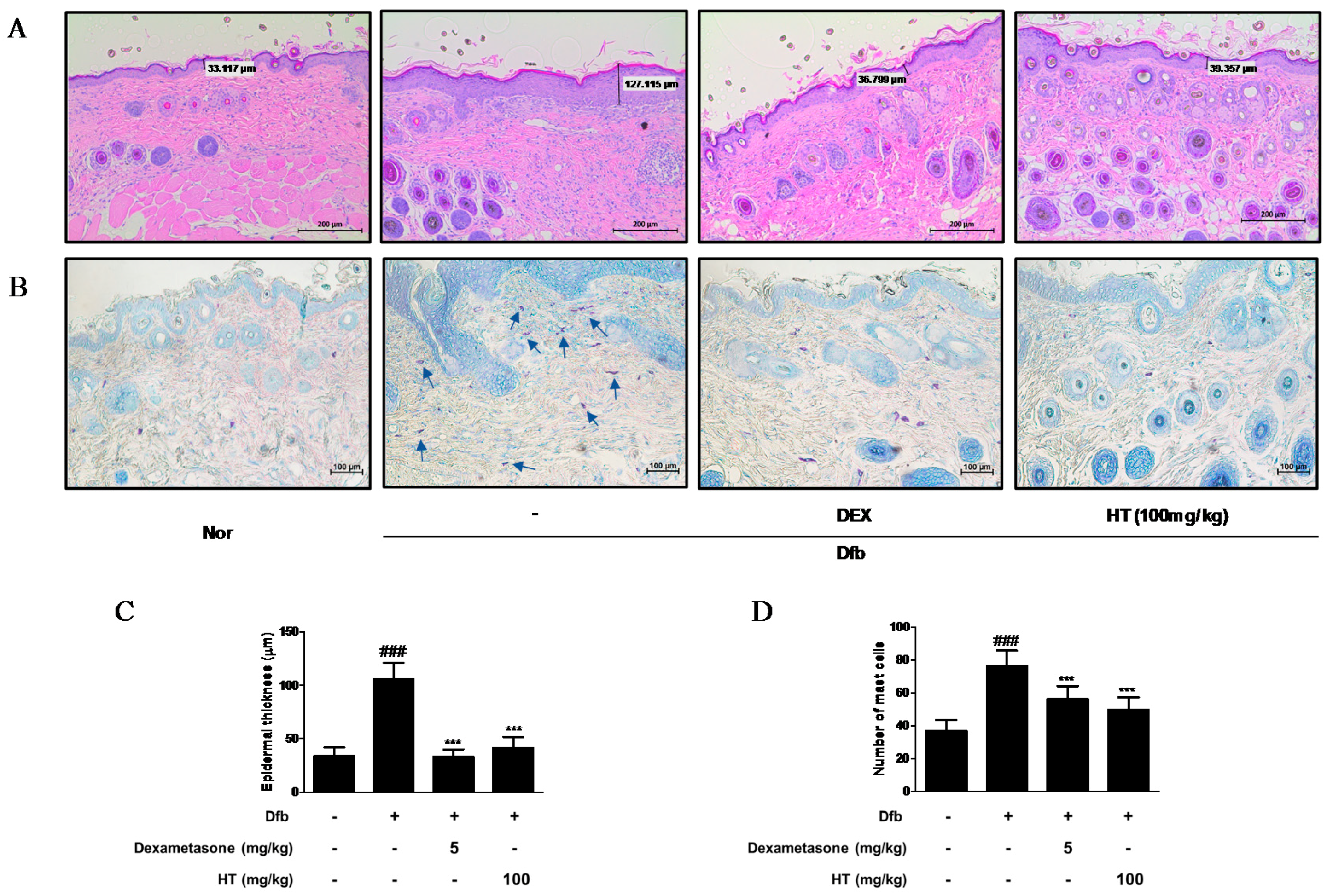
3.3. HT Reduced Epidermal Thickness and Mast Cell Infiltration in Dfb-Induced AD Mice Skin
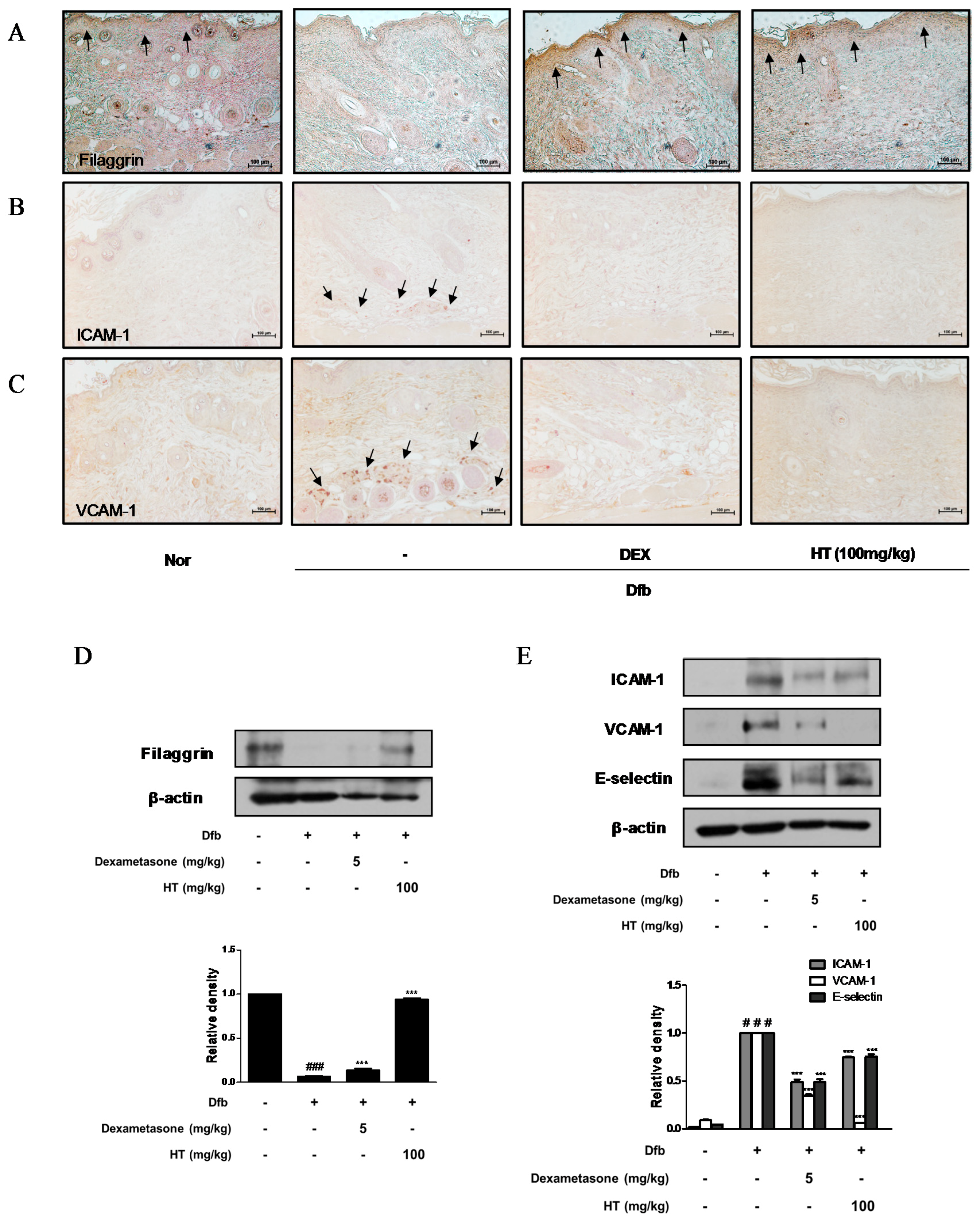
3.4. HT Restored Filaggrin Level and Inhibited the Expression of Adhesion Molecules in Dfb-Induced AD Mice Skin
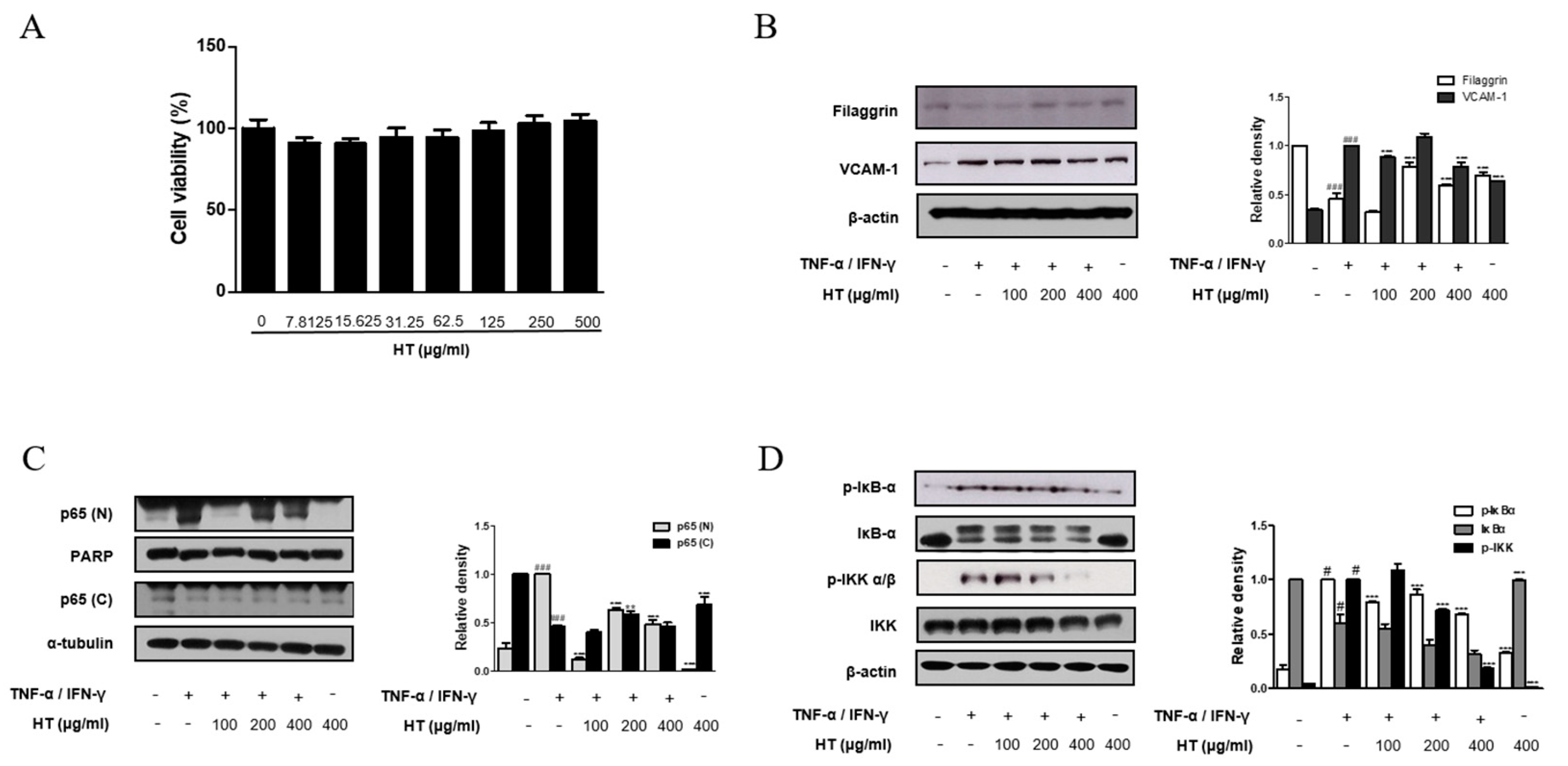
3.5. HT Regulated the Expression of Filaggrin and VCAM-1 in TNF-α/IFN-γ-Stimulated HaCaT Keratinocytes
3.6. HT Suppressed the Activation of the NF-κB Signaling Pathway in TNF-α/IFN-γ-Stimulated HaCaT Keratinocytes
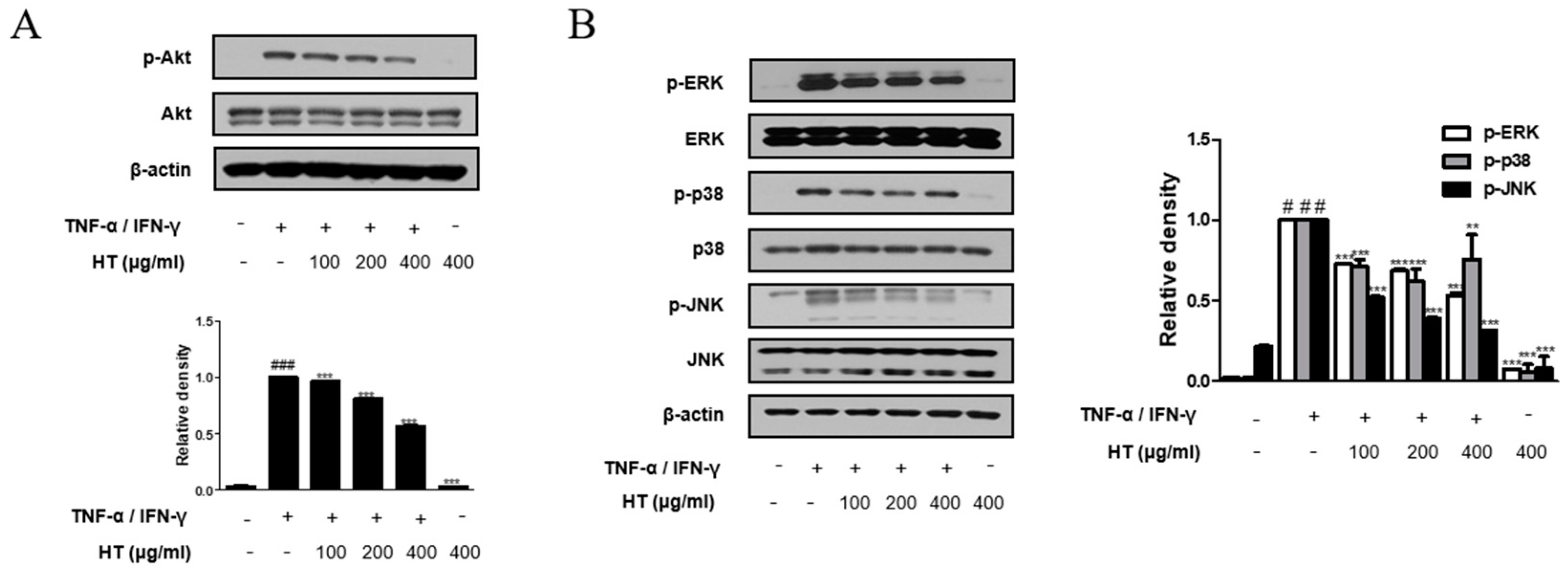
3.7. HT Suppressed the Phosphorylation of the Akt and MAPK Signaling Pathways in TNF-α/IFN-γ-Stimulated HaCaT Keratinocytes
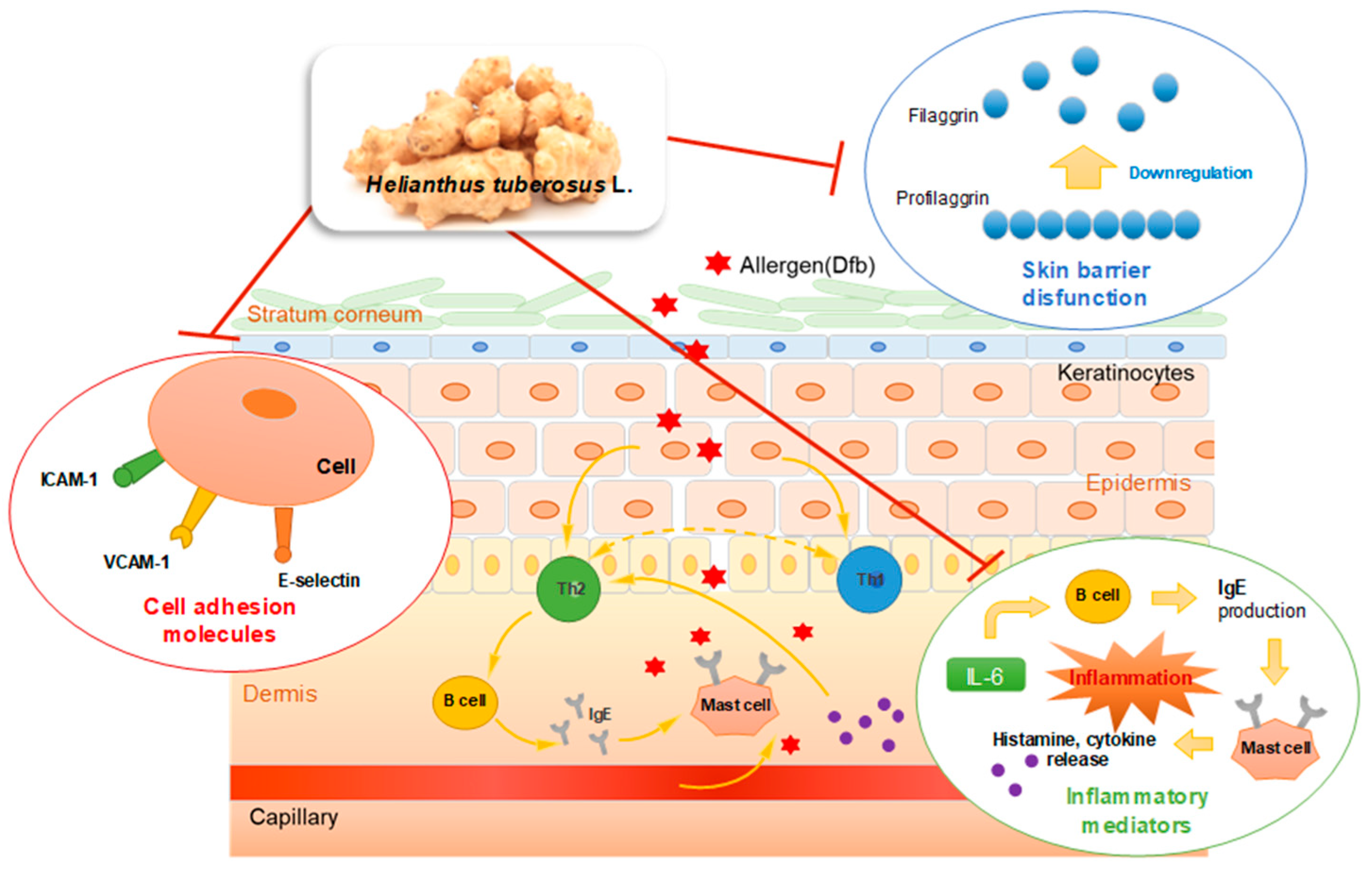
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jin, M.; Choi, J.K.; Choi, Y.A.; Kim, Y.Y.; Baek, M.C.; Lee, B.H.; Jang, Y.H.; Lee, W.J.; Lee, S.J.; Kim do, W.; et al. 1,2,4,5-Tetramethoxybenzene suppresses house dust mite-induced allergic inflammation in BALB/c mice. Int. Arch. Allergy Immunol. 2016, 170, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.K.; Oh, H.M.; Lee, S.; Kwon, T.K.; Shin, T.Y.; Rho, M.C.; Kim, S.H. Salvia plebeia suppresses atopic dermatitis-like skin lesions. Am. J. Chin. Med. 2014, 42, 967–985. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.K.; Nakamura, N.; Fuseda, K.; Tanaka, H.; Inagaki, N.; Nagai, H. Establishment of allergic dermatitis in NC/Nga mice as a model for severe atopic dermatitis. Biol. Pharm Bull. 2004, 27, 1376–1381. [Google Scholar] [CrossRef] [PubMed]
- Cabanillas, B.; Novak, N. Atopic dermatitis and filaggrin. Curr. Opin. Immunol. 2016, 42, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.W.; Lin, Z.C.; Hu, S.C.; Chiang, Y.C.; Hsu, L.F.; Lin, Y.C.; Lee, I.T.; Tsai, M.H.; Fang, J.Y. Urban particulate matter down-regulates filaggrin via COX2 expression/PGE2 production leading to skin barrier dysfunction. Sci. Rep. 2016, 6, 27995. [Google Scholar] [CrossRef] [PubMed]
- Lugovic, L.; Cupic, H.; Lipozencic, J.; Jakic-Razumovic, J. The role of adhesion molecules in atopic dermatitis. Acta Dermatovenerol. Croat. 2006, 14, 2–7. [Google Scholar] [PubMed]
- Jung, K.; Linse, F.; Heller, R.; Moths, C.; Goebel, R.; Neumann, C. Adhesion molecules in atopic dermatitis: VCAM-1 and ICAM-1 expression is increased in healthy-appearing skin. Allergy 1996, 51, 452–460. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.C.; Jia, H.; Aw, W.; Saito, K.; Hasegawa, S.; Kato, H. Beneficial effects of soluble dietary Jerusalem artichoke (Helianthus tuberosus) in the prevention of the onset of type 2 diabetes and non-alcoholic fatty liver disease in high-fructose diet-fed rats. Br. J. Nutr. 2014, 112, 709–717. [Google Scholar] [CrossRef] [PubMed]
- Radulovic, N.S.; Dordevic, M.R. Chemical composition of the tuber essential oil from Helianthus tuberosus L. (Asteraceae). Chem. Biodivers. 2014, 11, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Long, X.; Liu, Z.; Shao, H.; Liu, L. Analysis of phenolic acids of Jerusalem artichoke (Helianthus tuberosus L.) responding to salt-stress by liquid chromatography/tandem mass spectrometry. Sci. World J. 2014, 2014, 568043. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.M.; Zhao, J.Q.; Yang, J.L.; Idong, P.T.; Mei, L.J.; Tao, Y.D.; Shi, Y.P. Antioxidant and alpha-glucosidase inhibitory ingredients identified from Jerusalem artichoke flowers. Nat. Prod. Res. 2017, 1–5. [Google Scholar] [CrossRef]
- Yang, L.; He, Q.S.; Corscadden, K.; Udenigwe, C.C. The prospects of Jerusalem artichoke in functional food ingredients and bioenergy production. Biotechnol. Rep. (Amst.) 2015, 5, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Cheon, S.Y.; Chung, K.S.; Lee, S.J.; Hong, C.H.; Lee, K.T.; Jang, D.S.; Jeong, J.C.; Kwon, O.K.; Nam, J.H.; et al. Solanum tuberosum L. cv jayoung epidermis extract inhibits mite antigen-induced atopic dermatitis in NC/Nga mice by regulating the Th1/Th2 balance and expression of filaggrin. J. Med. Food 2015, 18, 1013–1021. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Choi, E.J.; Lee, K.S.; Kim, H.R.; Na, B.R.; Kwon, M.S.; Jeong, G.S.; Choi, H.G.; Choi, E.Y.; Jun, C.D. Oral administration of p-hydroxycinnamic acid attenuates atopic dermatitis by downregulating Th1 and Th2 cytokine production and keratinocyte activation. PLoS ONE 2016, 11, e0150952. [Google Scholar] [CrossRef] [PubMed]
- Arts, J.H.; Droge, S.C.; Bloksma, N.; Kuper, C.F. Local lymph node activation in rats after dermal application of the sensitizers 2,4-dinitrochlorobenzene and trimellitic anhydride. Food Chem. Toxicol. 1996, 34, 55–62. [Google Scholar] [CrossRef]
- Purushothaman, B.; Arumugam, P.; Song, J.M. A novel catecholopyrimidine based small molecule PDE4B inhibitor suppresses inflammatory cytokines in atopic mice. Front. Pharmacol. 2018, 9, 485. [Google Scholar] [CrossRef] [PubMed]
- Limandjaja, G.C.; van den Broek, L.J.; Waaijman, T.; van Veen, H.A.; Everts, V.; Monstrey, S.; Scheper, R.J.; Niessen, F.B.; Gibbs, S. Increased epidermal thickness and abnormal epidermal differentiation in keloid scars. Br. J. Dermatol. 2017, 176, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Irvine, A.D.; McLean, W.H.; Leung, D.Y. Filaggrin mutations associated with skin and allergic diseases. N. Engl. J. Med. 2011, 365, 1315–1327. [Google Scholar] [CrossRef] [PubMed]
- Yang, I.J.; Lee, D.U.; Shin, H.M. Inhibitory effect of valencene on the development of atopic dermatitis-like skin lesions in NC/Nga mice. Evid. Based Complement. Altern. Med. 2016, 2016, 9370893. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Jeon, Y.D.; Lee, Y.M.; Kim, D.K. The suppressive effect of puerarin on atopic dermatitis-like skin lesions through regulation of inflammatory mediators in vitro and in vivo. Biochem. Biophys. Res. Commun. 2018, 498, 707–714. [Google Scholar] [CrossRef] [PubMed]
- Seo, W.Y.; Youn, G.S.; Choi, S.Y.; Park, J. Butein, a tetrahydroxychalcone, suppresses pro-inflammatory responses in HaCaT keratinocytes. BMB Rep. 2015, 48, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Segrelles, C.; Ruiz, S.; Santos, M.; Martinez-Palacio, J.; Lara, M.F.; Paramio, J.M. Akt mediates an angiogenic switch in transformed keratinocytes. Carcinogenesis 2004, 25, 1137–1147. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.M.; Lee, B.; Min, J.H.; Kim, E.Y.; Kim, J.H.; Hong, S.; Kim, J.J.; Sohn, Y.; Jung, H.S. Effect of peiminine on DNCB-induced atopic dermatitis by inhibiting inflammatory cytokine expression in vivo and in vitro. Int. Immunopharmacol. 2018, 56, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Cole, C.; Kroboth, K.; Schurch, N.J.; Sandilands, A.; Sherstnev, A.; O’Regan, G.M.; Watson, R.M.; McLean, W.H.; Barton, G.J.; Irvine, A.D.; et al. Filaggrin-stratified transcriptomic analysis of pediatric skin identifies mechanistic pathways in patients with atopic dermatitis. J. Allergy Clin. Immunol. 2014, 134, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Arlian, L.G.; Morgan, M.S.; Peterson, K.T. House dust and storage mite extracts influence skin keratinocyte and fibroblast function. Int. Arch. Allergy Immunol. 2008, 145, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, K.; Mizutani, H. The role of cytokines/chemokines in the pathogenesis of atopic dermatitis. Curr. Probl. Dermatol. 2011, 41, 80–92. [Google Scholar] [CrossRef] [PubMed]
- Kwon, T.R.; Mun, S.K.; Oh, C.T.; Hong, H.; Choi, Y.S.; Kim, B.J.; Kim, B.J. Therapeutic effects of full spectrum light on the development of atopic dermatitis-like lesions in NC/Nga mice. Photochem. Photobiol. 2014, 90, 1160–1169. [Google Scholar] [CrossRef] [PubMed]
- Pendaries, V.; Malaisse, J.; Pellerin, L.; Le Lamer, M.; Nachat, R.; Kezic, S.; Schmitt, A.M.; Paul, C.; Poumay, Y.; Serre, G.; et al. Knockdown of filaggrin in a three-dimensional reconstructed human epidermis impairs keratinocyte differentiation. J. Investig. Dermatol. 2014, 134, 2938–2946. [Google Scholar] [CrossRef] [PubMed]
- O’Regan, G.M.; Sandilands, A.; McLean, W.H.; Irvine, A.D. Filaggrin in atopic dermatitis. J. Allergy Clin. Immunol. 2008, 122, 689–693. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.H.; Choi, D.; Chun, Y.J.; Noh, M. Keratinocyte-derived IL-24 plays a role in the positive feedback regulation of epidermal inflammation in response to environmental and endogenous toxic stressors. Toxicol. Appl. Pharmacol. 2014, 280, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Howell, M.D.; Kim, B.E.; Gao, P.; Grant, A.V.; Boguniewicz, M.; DeBenedetto, A.; Schneider, L.; Beck, L.A.; Barnes, K.C.; Leung, D.Y. Cytokine modulation of atopic dermatitis filaggrin skin expression. J. Allergy Clin. Immunol. 2009, 124, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Sawamura, D.; Meng, X.; Ina, S.; Sato, M.; Tamai, K.; Hanada, K.; Hashimoto, I. Induction of keratinocyte proliferation and lymphocytic infiltration by in vivo introduction of the IL-6 gene into keratinocytes and possibility of keratinocyte gene therapy for inflammatory skin diseases using IL-6 mutant genes. J. Immunol. 1998, 161, 5633–5639. [Google Scholar] [PubMed]
- Hanel, K.H.; Cornelissen, C.; Luscher, B.; Baron, J.M. Cytokines and the skin barrier. Int. J. Mol. Sci. 2013, 14, 6720–6745. [Google Scholar] [CrossRef] [PubMed]
- Noh, M.; Yeo, H.; Ko, J.; Kim, H.K.; Lee, C.H. MAP17 is associated with the T-helper cell cytokine-induced down-regulation of filaggrin transcription in human keratinocytes. Exp. Dermatol. 2010, 19, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Son, E.D.; Kim, H.J.; Park, T.; Shin, K.; Bae, I.H.; Lim, K.M.; Cho, E.G.; Lee, T.R. Staphylococcus aureus inhibits terminal differentiation of normal human keratinocytes by stimulating interleukin-6 secretion. J. Dermatol. Sci. 2014, 74, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Golias, C.; Tsoutsi, E.; Matziridis, A.; Makridis, P.; Batistatou, A.; Charalabopoulos, K. Review. Leukocyte and endothelial cell adhesion molecules in inflammation focusing on inflammatory heart disease. In Vivo 2007, 21, 757–769. [Google Scholar] [PubMed]
- Yoneshige, A.; Hagiyama, M.; Fujita, M.; Ito, A. Pathogenic actions of cell adhesion molecule 1 in pulmonary emphysema and atopic dermatitis. Front. Cell. Dev. Biol. 2015, 3, 75. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.J.; Ding, P.J.; Yang, L.L.; He, X.F.; Chen, M.J.; Wang, D.M.; Tian, Y.X.; Zhang, H.M. Oxymatrine sensitizes the HaCaT cells to the IFN-gamma pathway and downregulates MDC, ICAM-1, and SOCS1 by activating p38, JNK, and Akt. Inflammation 2018, 41, 606–613. [Google Scholar] [CrossRef] [PubMed]
- Sung, Y.Y.; Yoon, T.; Jang, S.; Kim, H.K. Forsythia suspensa suppresses house dust mite extract-induced atopic dermatitis in NC/Nga mice. PLoS ONE 2016, 11, e0167687. [Google Scholar] [CrossRef] [PubMed]
- Waters, C.E.; Shi-Wen, X.; Denton, C.P.; Abraham, D.J.; Pearson, J.D. Signaling pathways regulating intercellular adhesion molecule 1 expression by endothelin 1: Comparison with interleukin-1beta in normal and scleroderma dermal fibroblasts. Arthritis Rheum. 2006, 54, 649–660. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Kimura, K.; Yanai, R.; Chikama, T.; Nishida, T. Cytokine, chemokine, and adhesion molecule expression mediated by MAPKs in human corneal fibroblasts exposed to poly(I:C). Investig. Ophthalmol. Vis. Sci. 2008, 49, 3336–3344. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Xia, Y.Y.; Wang, L.; Liu, R.; Khoo, K.S.; Feng, Z.W. Neural cell adhesion molecule modulates mesenchymal stromal cell migration via activation of MAPK/ERK signaling. Exp. Cell. Res. 2012, 318, 2257–2267. [Google Scholar] [CrossRef] [PubMed]
- Homey, B.; Steinhoff, M.; Ruzicka, T.; Leung, D.Y. Cytokines and chemokines orchestrate atopic skin inflammation. J. Allergy Clin. Immunol. 2006, 118, 178–189. [Google Scholar] [CrossRef] [PubMed]
- Signorelli, S.S.; Mazzarino, M.C.; Di Pino, L.; Malaponte, G.; Porto, C.; Pennisi, G.; Marchese, G.; Costa, M.P.; Digrandi, D.; Celotta, G.; et al. High circulating levels of cytokines (IL-6 and TNFalpha), adhesion molecules (VCAM-1 and ICAM-1) and selectins in patients with peripheral arterial disease at rest and after a treadmill test. Vasc. Med. 2003, 8, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; He, Q.; Ren, Z.; Li, F.; Chen, W.; Lin, X.; Zhang, H.; Tai, G. Association of serum levels of intercellular adhesion molecule-1 and interleukin-6 with migraine. Neurol. Sci. 2015, 36, 535–540. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.; Li, X.K. Oxidative stress in atopic dermatitis. Oxid. Med. Cell. Longev. 2016, 2016, 2721469. [Google Scholar] [CrossRef] [PubMed]
- van den Bogaard, E.H.; Bergboer, J.G.; Vonk-Bergers, M.; van Vlijmen-Willems, I.M.; Hato, S.V.; van der Valk, P.G.; Schroder, J.M.; Joosten, I.; Zeeuwen, P.L.; Schalkwijk, J. Coal tar induces AHR-dependent skin barrier repair in atopic dermatitis. J. Clin. Investig. 2013, 123, 917–927. [Google Scholar] [CrossRef] [PubMed]







© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, Y.-M.; Lee, K.-Y.; An, H.-J. Inhibitory Effects of Helianthus tuberosus Ethanol Extract on Dermatophagoides farina body-induced Atopic Dermatitis Mouse Model and Human Keratinocytes. Nutrients 2018, 10, 1657. https://doi.org/10.3390/nu10111657
Kang Y-M, Lee K-Y, An H-J. Inhibitory Effects of Helianthus tuberosus Ethanol Extract on Dermatophagoides farina body-induced Atopic Dermatitis Mouse Model and Human Keratinocytes. Nutrients. 2018; 10(11):1657. https://doi.org/10.3390/nu10111657
Chicago/Turabian StyleKang, Yun-Mi, Kyou-Young Lee, and Hyo-Jin An. 2018. "Inhibitory Effects of Helianthus tuberosus Ethanol Extract on Dermatophagoides farina body-induced Atopic Dermatitis Mouse Model and Human Keratinocytes" Nutrients 10, no. 11: 1657. https://doi.org/10.3390/nu10111657
APA StyleKang, Y.-M., Lee, K.-Y., & An, H.-J. (2018). Inhibitory Effects of Helianthus tuberosus Ethanol Extract on Dermatophagoides farina body-induced Atopic Dermatitis Mouse Model and Human Keratinocytes. Nutrients, 10(11), 1657. https://doi.org/10.3390/nu10111657