Effect of Low Dose Docosahexaenoic Acid-Rich Fish Oil on Plasma Lipids and Lipoproteins in Pre-Menopausal Women: A Dose–Response Randomized Placebo-Controlled Trial



Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Study Design
2.3. Sample Collection and Processing
2.4. Clinical Chemistry Methods
2.5. Lipoprotein Analysis
2.6. Lipoprotein Particle Size Analysis
2.7. Fatty Acid Analysis
2.8. Statistical Analysis
3. Results
3.1. Subject Characteristics and Dietary Intake
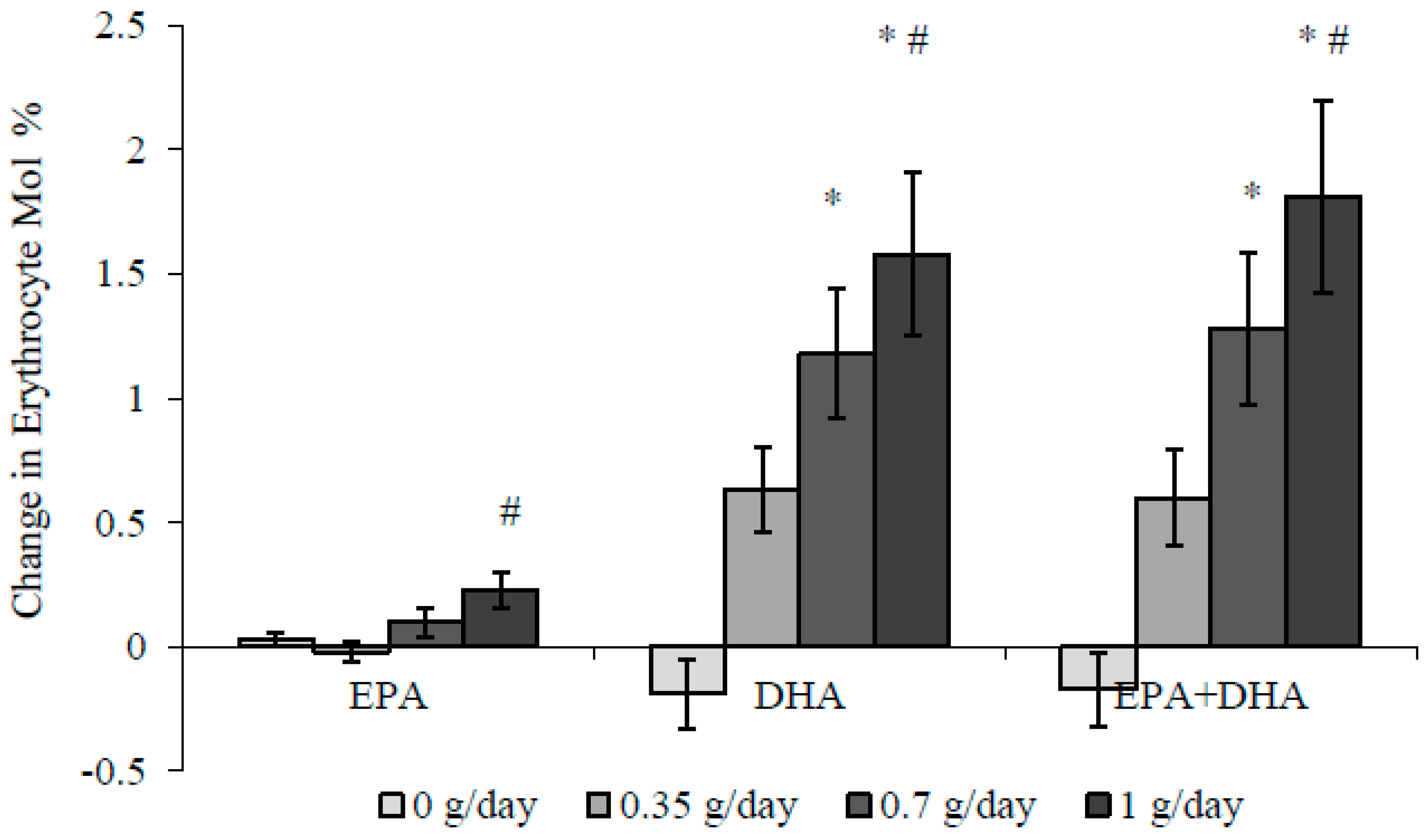
3.2. Blood Pressure and Fatty Acids
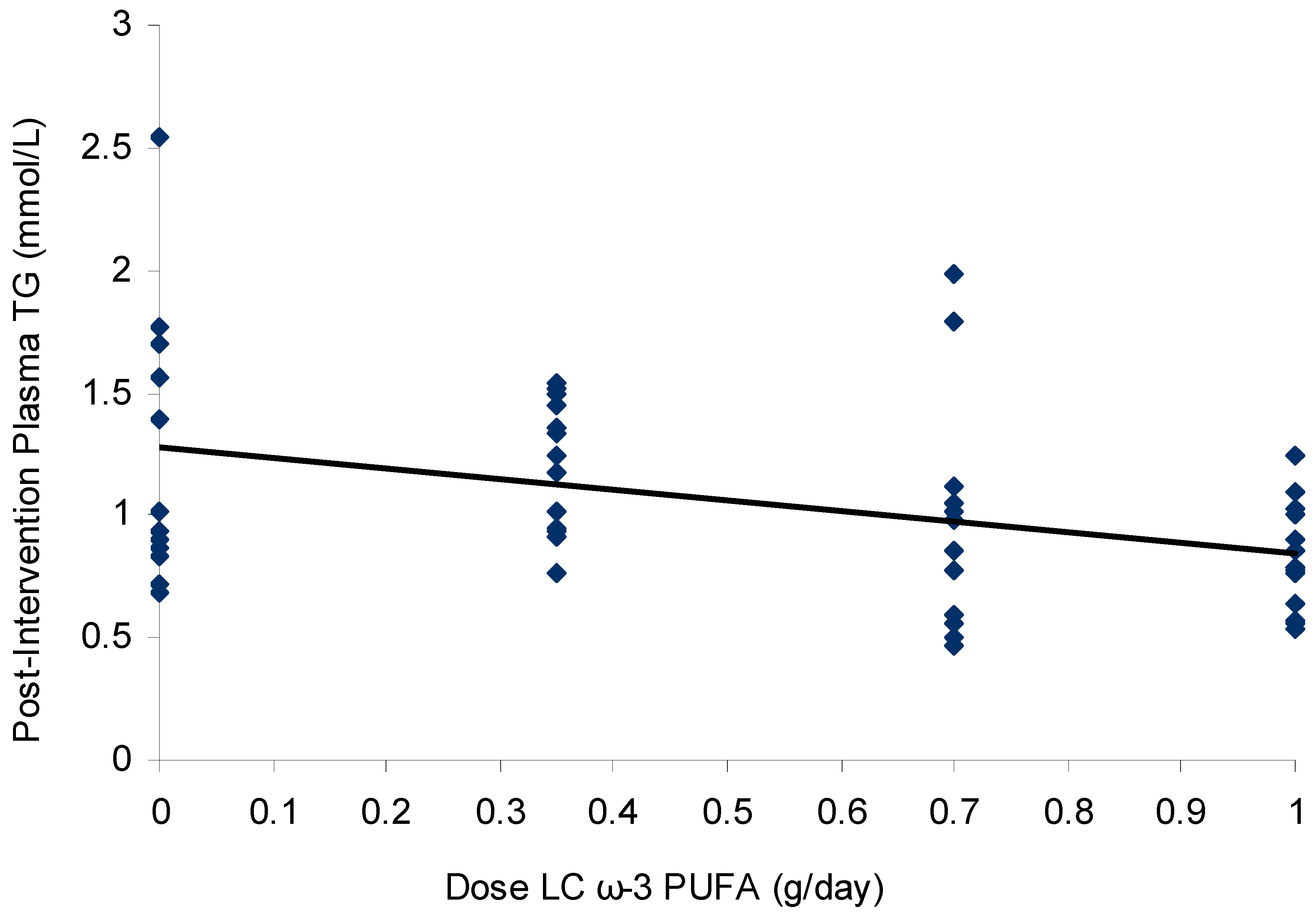
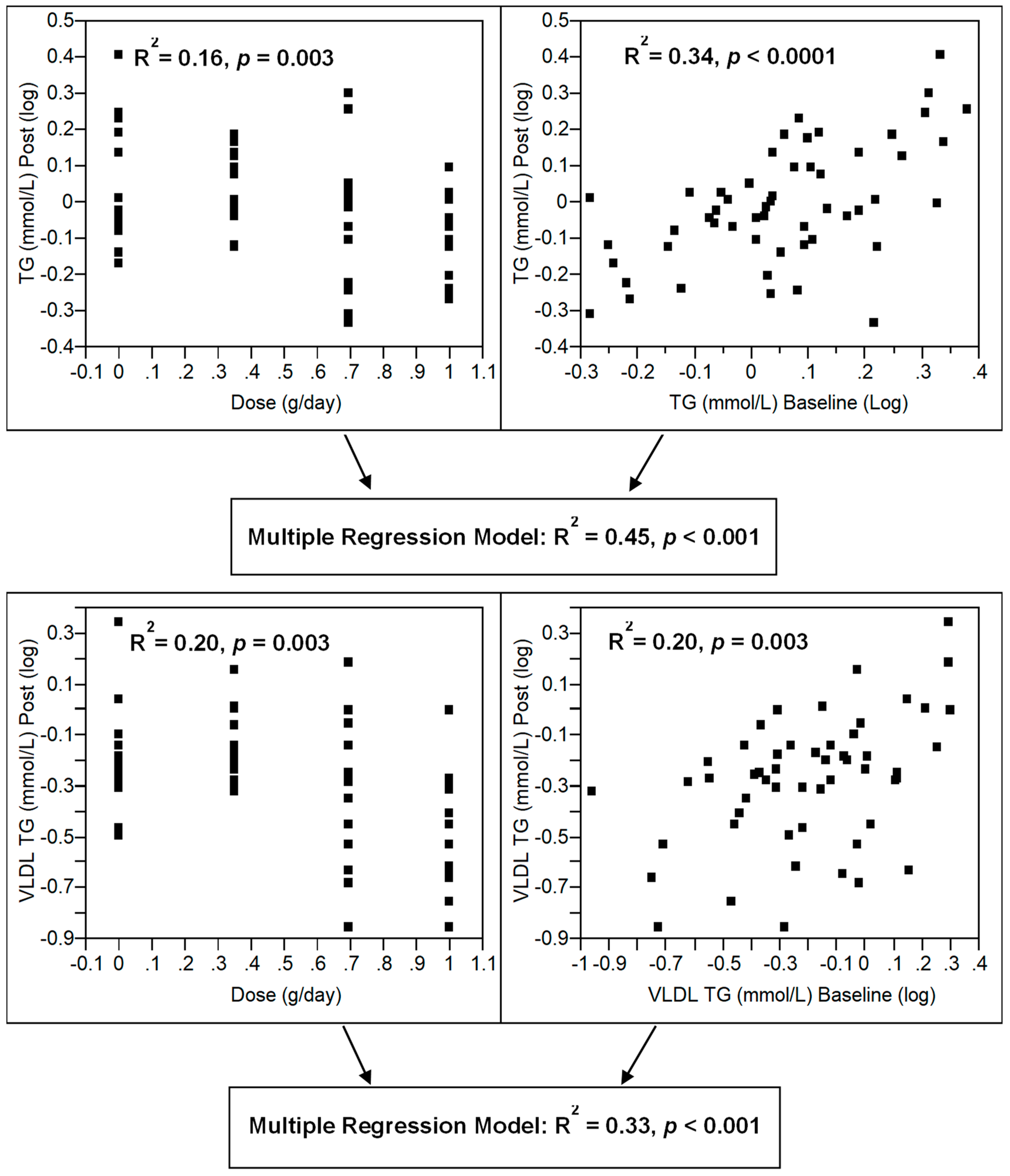
3.3. Effect of Dose–Response n-3 LCPUFA Supplementation on Plasma Lipids
3.4. Lipoprotein Composition and Particle Size
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Harris, W.S. N-3 fatty acids and serum lipoproteins: Human studies. Am. J. Clin. Nutr. 1997, 65, 1645S–1654S. [Google Scholar] [CrossRef] [PubMed]
- Balk, E.M.; Lichtenstein, A.H.; Chung, M.; Kupelnick, B.; Chew, P.; Lau, J. Effects of omega-3 fatty acids on coronary restenosis, intima–media thickness, and exercise tolerance: A systematic review. Atherosclerosis 2006, 184, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Bays, H.E.; Tighe, A.P.; Sadovsky, R.; Davidson, M.H. Prescription omega-3 fatty acids and their lipid effects: Physiologic mechanisms of action and clinical implications. Expert Rev. Cardiovasc. Ther. 2008, 6, 391–409. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, M.; Berneis, K. Low-density lipoprotein size and cardiovascular risk assessment. Q. J. Med. 2006, 99, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.A.; Burke, V.; Puddey, I.B.; Watts, G.F.; O’Neal, D.N.; Best, J.D.; Beilin, L.J. Purified eicosapentaneoic acid and docosahexaneoic acids have differential effects on serum lipids and lipoproteins, LDL particle size, glucose, and insulin in mildly hyperlipidemic men. Am. J. Clin. Nutr. 2000, 71, 1085–1094. [Google Scholar] [CrossRef] [PubMed]
- Maki, K.C.; Van Elswyk, M.E.; McCarthy, D.; Hess, S.P.; Veith, P.E.; Bell, M.; Subbaiah, P.; Davidson, M.H. Lipid responses to a dietary docosahexaenoic acid supplement in men and women with below average levels of high density lipoprotein cholesterol. J. Am. Coll. Nutr. 2005, 24, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Neff, L.M.; Culiner, J.; Cunningham-Rundles, S.; Seidman, C.; Meehan, D.; Maturi, J.; Wittkowski, K.M.; Levine, B.; Breslow, J.L. Algal docosahexaenoic acid affects plasma lipoprotein particle size distribution in overweight and obese adults. J. Nutr. 2010, 141, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Woodman, R.J.; Mori, T.A.; Burke, V.; Puddey, I.B.; Watts, G.F.; Beilin, L.J. Effects of purified eicosapentaenoic and docosahexaenoic acids on glycemic control, blood pressure, and serum lipids in type 2 diabetic patients with treated hypertension. Am. J. Clin. Nutr. 2002, 76, 1007–1015. [Google Scholar] [CrossRef] [PubMed]
- Milte, C.M.; Coates, A.M.; Buckley, J.D.; Hill, A.M.; Howe, P.R. Dose-dependent effects of docosahexaenoic acid-rich fish oil on erythrocyte docosahexaenoic acid and blood lipid levels. Br. J. Nutr. 2008, 99, 1083–1088. [Google Scholar] [CrossRef] [PubMed]
- Stark, K.D.; Holub, B.J. Differential eicosapentaenoic acid elevations and altered cardiovascular disease risk factor responses after supplementation with docosahexaenoic acid in postmenopausal women receiving and not receiving hormone replacement therapy. Am. J. Clin. Nutr. 2004, 79, 765–773. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.H.; Lu, S.C.; Wang, T.F.; Jou, H.J.; Wang, T.A. Effects of docosahexaenoic acid supplementation on blood lipids, estrogen metabolism, and in vivo oxidative stress in postmenopausal vegetarian women. Eur. J. Clin. Nutr. 2005, 60, 386–392. [Google Scholar] [CrossRef] [PubMed]
- Skulas-Ray, A.C.; Alaupovic, P.; Kris-Etherton, P.M.; West, S.G. Dose-response effects of marine omega-3 fatty acids on apolipoproteins, apolipoprotein-defined lipoprotein subclasses, and lp-pla2 in individuals with moderate hypertriglyceridemia. J. Clin. Lipidol. 2015, 9, 360–367. [Google Scholar] [CrossRef] [PubMed]
- Gidez, L.I.; Miller, G.J.; Burstein, M.; Slagle, S.; Eder, H.A. Separation and quantitation of subclasses of human plasma high density lipoproteins by a simple precipitation procedure. J. Lipid Res. 1982, 23, 1206–1223. [Google Scholar] [PubMed]
- Geppert, J.; Kraft, V.; Demmelmair, H.; Koletzko, B. Microalgal docosahexaenoic acid decreases plasma triacylglycerol in normolipidaemic vegetarians: A randomised trial. Br. J. Nutr. 2006, 95, 779–786. [Google Scholar] [CrossRef] [PubMed]
- Schwellenbach, L.J.; Olson, K.L.; McConnell, K.J.; Stolcpart, R.S.; Nash, J.D.; Merenich, J.A.; Clinical Pharmacy Cardiac Risk Service Study Group. The triglyceride-lowering effects of a modest dose of docosahexaenoic acid alone versus in combination with low dose eicosapentaenoic acid in patients with coronary artery disease and elevated triglycerides. J. Am. Coll. Nutr. 2006, 25, 480–485. [Google Scholar] [CrossRef] [PubMed]
- Theobald, H.E.; Chowienczyk, P.J.; Whittall, R.; Humphries, S.E.; Sanders, T.A.B. LDL cholesterol-raising effect of low-dose docosahexaneoic acid in middle-aged men and women. Am. J. Clin. Nutr. 2004, 79, 558–563. [Google Scholar] [CrossRef] [PubMed]
- Barnett, J.B.; Woods, M.N.; Lamon-Fava, S.; Schaefer, E.J.; McNamara, J.R.; Spiegelman, D.; Hertzmark, E.; Goldin, B.; Longcope, C.; Gorbach, S.L. Plasma lipid and lipoprotein levels during the follicular and luteal phases of the menstrual cycle. J. Clin. Endocrinol. Metab. 2004, 89, 776–782. [Google Scholar] [CrossRef] [PubMed]
- Tonolo, G.; Ciccarese, M.; Brizzi, P.; Milia, S.; Dessole, S.; Puddu, L.; Secchi, G.; Maoili, M. Cyclical variation of plasma lipids, apolipoproteins, and lipoprotein (a) during menstrual cycle of normal women. Am. J. Physiol. 1995, 269, E1101–E1105. [Google Scholar] [CrossRef] [PubMed]
- Iso, H.; Naito, Y.; Sato, S.; Kitamura, A.; Okamura, T.; Sankai, T.; Shimamoto, T.; Iida, M.; Komachi, Y. Serum triglycerides and risk of coronary heart disease among Japanese men and women. Am. J. Epidemiol. 2000, 153, 490–499. [Google Scholar] [CrossRef]
- Sharrett, A.R.; Sorlie, P.D.; Chambless, L.E.; Folsom, A.R.; Hutchinson, R.G.; Heiss, G.; Szklo, M. Relative importance of various risk factors for asymptomatic carotid atherosclerosis versus coronary heart disease incidence: The atherosclerosis risk in communities study. Am. J. Epidemiol. 1999, 149, 843–852. [Google Scholar] [CrossRef] [PubMed]
- Geleijnse, J.M.; Giltay, E.J.; Grobbee, D.E.; Donders, A.R.T.; Kok, F.J. Blood pressure response to fish oil supplementation: Metaregression analysis of randomized trials. J. Hypertens. 2002, 20, 1493–1499. [Google Scholar] [CrossRef] [PubMed]
- Meyer, B.J.; Lane, A.E.; Mann, N.J. Comparison of seal oil to tuna oil on plasma lipid levels and blood pressure in hypertriglyceridaemic subjects. Lipids 2009, 44, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [PubMed]
- Meyer, B.J.; Duvillard, L.; Owen, A.; Packard, C.J.; Caslake, M.J. Fractionation of cholesteryl ester rich intermediate density lipoprotein subpopulations by chondroitin sulphate. Atherosclerosis 2007, 195, e28–e34. [Google Scholar] [CrossRef] [PubMed]
- Blanche, P.J.; Gong, E.L.; Forte, T.M.; Nichols, A.V. Characterization of human high-density lipoproteins by gradient gel electrophoresis. Biochim. Biophys. Acta 1981, 665, 408–419. [Google Scholar] [CrossRef]
- Ridges, L.; Sunderland, R.; Moerman, K.; Meyer, B.; Astheimer, L.; Howe, P. Cholesterol lowering benefits of soy and linseed enriched foods. Asia Pac. J. Clin. Nutr. 2001, 10, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Lepage, G.; Roy, C.C. Direct transesterification of all classes of lipids in a one-step reaction. J. Lipid Res. 1986, 27, 114–120. [Google Scholar] [PubMed]
- Vickers, A.J.; Altman, D.G. Analysing controlled trials with baseline and follow-up measurements. BMJ 2001, 323, 1123–1124. [Google Scholar] [CrossRef] [PubMed]
- Tu, Y.-K.; Blance, A.; Clerehugh, V.; Gilthorpe, M.S. Statistical power for analyses of changes in randomized controlled trials. J. Dent. Res. 2005, 84, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Van Breukelen, G.J.P. ANCOVA versus change from baseline had more power in randomized studies and more bias in nonrandomized studies. J. Clin. Epidemiol. 2006, 59, 920–925. [Google Scholar] [CrossRef] [PubMed]
- Vickers, A.J. The use of percentage change from baseline as an outcome in a controlled trial is statistically inefficient: A simulation study. BMC Med. Res. Methodol. 2001, 1. [Google Scholar] [CrossRef]
- Borm, G.F.; Fransen, J.; Lemmens, W.A.J.G. A simple sample size formula for analysis of covariance in randomized clinical trials. J. Clin. Epidemiol. 2007, 60, 1234–1238. [Google Scholar] [CrossRef] [PubMed]
- Conquer, J.A.; Holub, B.J. Supplementation with an algae source of docosahexaenoic acid increases (n-3) fatty acid status and alters selected risk factors for heart disease in vegetarian subjects. J. Nutr. 1996, 126, 3032–3039. [Google Scholar] [CrossRef] [PubMed]
- Grimsgaard, S.; Bonaa, K.H.; Hansen, J.B.; Nordoy, A. Highly purified eicosapentaenoic acid and docosahexaenoic acid in humans have similar triacylglycerol-lowering effects but divergent effects on serum fatty acids. Am. J. Clin. Nutr. 1997, 66, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Kelley, D.S.; Siegel, D.; Vemuri, M.; Mackey, B.E. Docosahexaenoic acid supplementation improves fasting and postprandial lipid profiles in hypertriglyceridemic men. Am. J. Clin. Nutr. 2007, 86, 324–333. [Google Scholar] [CrossRef] [PubMed]
- Meyer, B.J.; Hammervold, T.; Rustan, A.C.; Howe, P.R.C. Dose-dependent effects of docosahexaenoic acid supplementation on blood lipids in statin-treated hyperlipidaemic subjects. Lipids 2007, 42, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Su, T.-C.; Hwang, J.-J.; Huang, K.-C.; Chiang, F.-T.; Chein, K.-L.; Wang, K.-Y.; Charng, M.-J.; Tsai, W.-C.; Lin, L.-Y.; Vige, R.; et al. A randomized, double-blind, placebo-controlled clinical trial to assess the efficacy and safety of ethyl ester omega-3 fatty acid in Taiwanese hypertriglyceridemic patients. J. Atheroscler. Thromb. 2017, 24, 275–289. [Google Scholar] [CrossRef] [PubMed]
- Higgins, S.; Carroll, Y.L.; McCarthy, S.N.; Corridan, B.M.; Roche, H.M.; Wallace, J.M.W.; O’Brien, N.M.; Morrissey, P.A. Susceptibility of ldl to oxidative modification in healthy volunteers supplemented with low doses of n-3 polyunsaturated fatty acids. Br. J. Nutr. 2001, 85, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Minihane, A.M.; Khan, S.; Leigh-Firbank, E.C.; Talmud, P.; Wright, J.W.; Murphy, M.C.; Griffin, B.A.; Williams, C.M. Apoe polymorphism and fish oil supplementation in subjects with an atherogenic lipoprotein phenotype. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 1990–1997. [Google Scholar] [CrossRef] [PubMed]
- Stark, K.D.; Park, E.J.; Maines, V.A.; Holub, B.J. Effect of a fish-oil concentrate on serum lipids in post-menopausal women receiving and not receiving hormone replacement therapy in a placebo-controlled, double-blind trial. Am. J. Clin. Nutr. 2000, 72, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Schuchardt, J.; Neubronner, J.; Kressel, G.; Merkel, M.; von Schacky, C.; Hahn, A. Associations between Omega-3 Index increase and triacylglyceride decrease in subjects with hypertriglyceridemia in response to six month of EPA and DHA supplementation. Prostagland. Leuk. Essent. Fat. Acids 2014, 91, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Madden, J.; Carrero, J.J.; Brunner, A.; Dastur, N.; Shearman, C.P.; Calder, P.C.; Grimble, R.F. Polymorphisms in the cd36 gene modulate the ability of fish oil supplements to lower fasting plasma triacyl glycerol and raise hdl cholesterol concentrations in healthy middle-aged men. Prostagland. Leuk. Essent. Fat. Acids 2008, 78, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Tai, E.S.; Corella, D.; Demissie, S.; Cupples, L.A.; Coltell, O.; Schaefer, E.J.; Tucker, K.L.; Ordovas, J.M. Polyunsaturated fatty acids interact with the PPARA-L162V polymorphism to affect plasma triglyceride and apolipoprotein C-III concentrations in the Framingham Heart Study. J. Nutr. 2005, 135, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Madden, J.; Williams, C.M.; Calder, P.C.; Lietz, G.; Miles, E.A.; Cordell, H.; Mathers, J.C.; Minihane, A.M. The impact of common gene variants on the response of biomarkers of cardiovascular disease (cvd) risk to increased fish oil fatty acids intakes. Annu. Rev. Nutr. 2011, 31, 203–234. [Google Scholar] [CrossRef] [PubMed]
- Ordovas, J.M. Genetic interactions with diet influence the risk of cardiovascular disease. Am. J. Clin. Nutr. 2006, 83, 443S–446S. [Google Scholar] [CrossRef] [PubMed]
- Asset, G.; Bauge, E.; Fruchart, J.C.; Dallongeville, J. Lack of triglyceride-lowering properties of fish oil in apolipoprotein e-deficient mice. Arterioscler. Thromb. Vasc. Biol. 2001, 21, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Caslake, M.J.; Miles, E.A.; Kofler, B.M.; Lietz, G.; Curtis, P.; Armah, C.K.; Kimber, A.C.; Grew, J.P.; Farrell, L.; Stannard, J.; et al. Effect of sex and genotype on cardiovascular biomarker response to fish oils: The FINGEN study. Am. J. Clin. Nutr. 2008, 88, 618–629. [Google Scholar] [CrossRef] [PubMed]
- Olano-Martin, E.; Anil, E.; Caslake, M.J.; Packard, C.J.; Bedford, D.; Stewart, G.; Peiris, D.; Williams, C.M.; Minihane, A.M. Contribution of apolipoprotein e genotype and docosahexaenoic acid to the ldl-cholesterol response to fish oil. Atherosclerosis 2010, 209, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Davidson, M.H. Mechanisms for the hypotriglyceridemic effect of marine omega-3 fatty acids. Am. J. Cardiol. 2006, 98, 27i–33i. [Google Scholar] [CrossRef] [PubMed]
- Deckelbaum, R.J.; Worgall, T.S.; Seo, T. n-3 fatty acids and gene expression. Am. J. Clin. Nutr. 2006, 83, 1520S–1525S. [Google Scholar] [CrossRef] [PubMed]
- Krey, G.; Braissant, O.; L’Horset, F.; Kalkhoven, E.; Perroud, M.; Parker, M.G.; Wahli, W. Fatty acids, eicosanoids, and hypolipidemic agents identified as ligands of peroxisome proliferator-activated receptors by coactivator-dependent receptor ligand assay. Mol. Endocrinol. 1997, 11, 779–791. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Muniz, F.J.; Bastida, S.; Viejo, J.M.; Terpstra, A.H.M. Small supplements of n-3 fatty acids change serum low density lipoprotein composition by decreasing phospholipid and apolipoprotein b concentrations in young adult women. Eur. J. Nutr. 1999, 38, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Skulas-Ray, A.C.; Kris-Etherton, P.M.; Harris, W.S.; Vanden Heuvel, J.P.; Wagner, P.R.; West, S.G. Dose-response effects of omega-3 fatty acids on triglycerides, inflammation, and endothelial function in healthy persons with moderate hypertriglyceridemia. Am. J. Clin. Nutr. 2011, 93, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Finnegan, Y.E.; Minihane, A.M.; Leigh-Firbank, E.C.; Kew, S.; Meijer, G.W.; Muggli, R.; Calder, P.C.; Williams, C.M. Plant- and marine-derived n-3 polyunsaturated fatty acids have differential effects on fasting and postprandial blood lipid concentrations and on the susceptibility of LDL to oxidative modification in moderately hyperlipidemic subjects. Am. J. Clin. Nutr. 2003, 77, 783–795. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.A. Omega-3 fatty acids and hypertension in humans. Clin. Exp. Pharmacol. Physiol. 2006, 33, 842–846. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
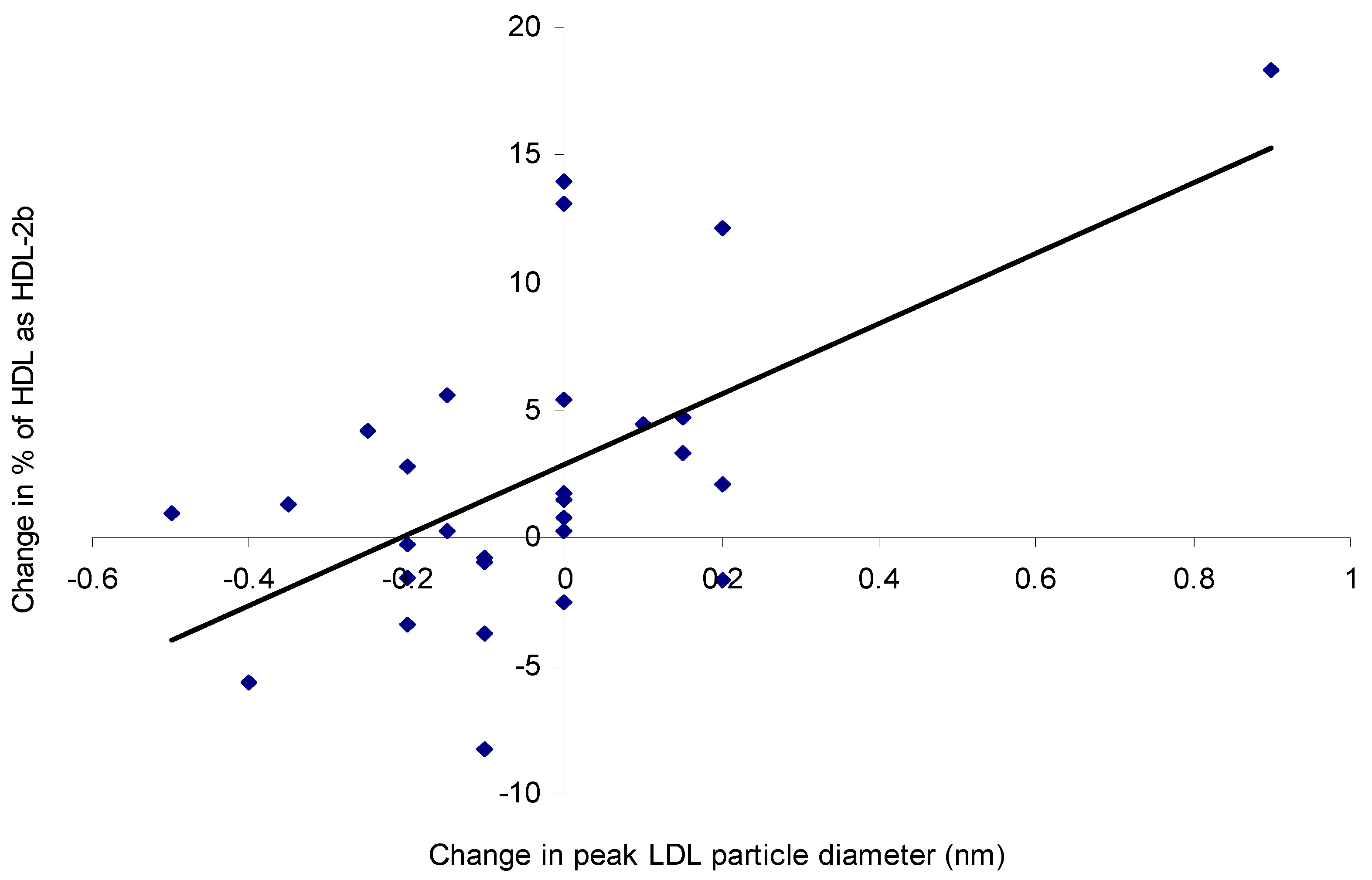{kind=link}
| 0 g/day (n = 12) | 0.35 g/day (n = 15) | 0.7 g/day (n = 13) | 1 g/day (n = 13) | p Value | |
|---|---|---|---|---|---|
| N | 12 | 15 | 13 | 13 | |
| OC/Non-OC | 6/6 | 7/8 | 5/8 | 7/6 | |
| Age (years) | 28 ± 2 | 27 ± 2 | 24 ± 1 | 28 ± 2 | 0.51 |
| BMI (kg/m2) | 23 ± 1 | 26 ± 1 | 26 ± 2 | 24 ± 1 | 0.57 |
| SBP (mm Hg) | 116 (108, 120) | 117 (107, 121) | 115 (110, 125) | 110 (105, 112) | 0.07 |
| DBP (mm Hg) | 69 (65, 75) | 70 (68, 80) | 70 (65, 82) | 69 (66, 72) | 0.58 |
| TC (mmol/L) | 4.60 (4.02, 5.42) | 4.61 (4.13, 4.88) | 4.30 (4.14, 4.95) | 4.60 (3.90, 5.22) | 0.98 |
| Energy (kJ) | 6525 (5696, 7475) | 5871 (5005, 6888) | 6642 (5460, 8081) | 5118 (4115, 6365) | 0.24 |
| Total Fat (g) | 50 (47, 71) | 58 (48, 70) | 60 (49, 75) | 43 (31, 61) | 0.44 |
| SFA (g) | 24 (20, 29) | 23 (18, 28) | 24 (20, 30) | 18 (12, 25) | 0.50 |
| PUFA (g) | 7.1 (5.6, 9.0) | 8.2 (6.4, 11) | 8.6 (6.4, 12) | 5.6 (3.6, 8.6) | 0.34 |
| MUFA (g) | 21 (16, 26) | 22 (18, 26) | 21 (17, 27) | 16 (11, 22) | 0.44 |
| Protein (g) | 81 (69, 94) | 72 (62, 84) | 81 (67, 99) | 70 (60, 82) | 0.44 |
| CHO (g) | 179 (162, 197) | 148 (126, 174) | 177 (142, 221) | 136 (113, 164) | 0.09 |
| n-3 LCPUFA (g/day) | TG (mmol/L) 0 weeks | TG (mmol/L) 8 weeks | TC (mmol/L) 0 weeks | TC (mmol/L) 8 weeks |
|---|---|---|---|---|
| 0.00 (n = 12) | 1.18 (0.77, 1.57) | 0.98 (0.84, 1.66) | 4.60 (4.02, 5.42) | 4.68 (3.89, 5.05) |
| 0.35 (n = 15) | 1.27 (1.07, 1.69) | 1.18 (0.91, 1.45) | 4.61 (4.13, 4.88) | 4.43 (4.04, 4.75) |
| 0.70 (n = 13) | 1.07 (0.91, 1.86) | 0.98 (0.58, 1.08) | 4.30 (4.14, 4.95) | 4.42 (4.07, 4.87) |
| 1.00 (n = 13) | 1.09 (0.78, 1.27) | 0.79 * (0.60, 1.02) | 4.60 (3.90, 5.22) | 4.44 (3.85, 5.41) |
| VLDL | 0 g/day (n = 12) 0 weeks | 0 g/day (n = 12) 8 weeks | 0.35 g/day (n = 15) 0 weeks | 0.35 g/day (n = 15) 8 weeks | 0.7 g/day (n = 13) 0 weeks | 0.7 g/day (n = 13) 8 weeks | 1 g/day (n = 13) 0 weeks | 1 g/day (n = 13) 8 weeks | Dose Effect (R2, p Value) |
|---|---|---|---|---|---|---|---|---|---|
| TG (mmol/L) | 0.81 (0.51, 1.23) | 0.60 (0.50, 0.77) | 0.72 (0.49, 0.94) | 0.66 (0.58, 0.85) | 0.96 (0.31, 1.37) | 0.51 (0.26, 0.80) | 0.50 (0.35, 0.77) | 0.35 # (0.22, 0.52) | R2 = 0.20, p = 0.003 |
| Chol (mmol/L) | 0.36 (0.19, 0.47) | 0.19 (0.16, 0.48) | 0.31 (0.17, 0.40) | 0.24 (0.18, 0.35) | 0.30 (0.14, 0.63) | 0.19 (0.11, 0.29) | 0.21 (0.15, 0.30) | 0.16 (0.09, 0.24) | R2 = 0.14, p = 0.02 |
| PL (mg/dL) | 21 (13, 32) | 15 (12, 29) | 19 (12, 22) | 15 (14, 19) | 21 (9, 41) | 14 (7, 19) | 16 (10, 20) | 11 (6, 17) | R2 = 0.17, p = 0.003 |
| PR (mg/dL) | 14 (10, 17) | 12 (10, 15) | 18 (12, 21) | 12 (9, 17) | 15 (9, 22) | 11 (9, 14) | 10 (9, 15) | 11 (8, 13) | p = 0.33 |
| TG/PR | 5.4 (3.1, 7.5) | 5.0 (3.4, 6.6) | 3.9 (3.3, 5.5) | 6.0 (3.5, 6.7) | 4.0 (2.4, 8.1) | 3.9 (2.6, 5.3) | 4.1 (2.4, 5.4) | 2.4 # (2.2, 4.2) | R2 = 0.17, p = 0.003 |
| apoB (mmol/L) | 6.3 (3.3, 9.0) | 5.2 (3.5, 6.2) | 7.7 (6.5, 9.6) | 5.7 (4.8, 6.5) | 5.5 (4.2, 7.2) | 4.3 (3.2, 6.6) | 5.0 (4.0, 8.0) | 4.7 (2.5, 5.5) | p = 0.49 |
| TG % | 58 (55, 61) | 57 (53, 63) | 57 (49, 60) | 58 (56, 62) | 57 (43, 60) | 55 (50, 59) | 51 (49, 58) | 50 # (48, 55) | R2 = 0.13, p = 0.02 |
| Chol % | 10 (10, 12) | 9 (8, 11) | 9 (8, 11) | 10 (8, 11) | 10 (8, 13) | 9 (8, 11) | 10 (9, 11) | 9 (8, 11) | p = 0.97 |
| PL % | 17 (16, 18) | 16 (16, 19) | 17 (15, 17) | 16 (15, 17) | 16 (16, 19) | 17 (15, 18) | 18 (16, 19) | 18 (16, 19) | p = 0.82 |
| PR % | 11 (8, 18) | 12 (9, 17) | 14 (11, 17) | 10 (9, 16) | 15 (7, 19) | 15 (11, 21) | 14(11, 21) | 21 # (13, 23) | R2 = 0.16, p = 0.002 |
| 0 g/day (n = 12) 0 weeks | 0 g/day (n = 12) 8 weeks | 0.35 g/day (n = 15) 0 weeks | 0.35 g/day (n = 15) 8 weeks | 0.7 g/day (n = 13) 0 weeks | 0.7 g/day (n = 13) 8 weeks | 1 g/day (n = 13) 0 weeks | 1 g/day (n = 13) 8 weeks | Dose Effect (R2, p Value) | |
|---|---|---|---|---|---|---|---|---|---|
| IDL-C (mmol/L) | 0.33 (0.26, 0.40) | 0.37 (0.28, 0.44) | 0.37 (0.29, 0.53) | 0.35 (0.27, 0.50) | 0.37 (0.22, 0.46) | 0.30 (0.20, 0.40) | 0.36 (0.24, 0.49) | 0.41 (0.20, 0.50) | p = 0.17 |
| IDL-TG (mmol/L) | 0.15 (0.10, 0.17) | 0.14 (0.11, 0.16) | 0.15 (0.13, 0.19) | 0.15 (0.12, 0.17) | 0.11 (0.09, 0.14) | 0.11 (0.08, 0.12) | 0.14 (0.11, 0.17) | 0.12 (0.10, 0.18) | p = 0.37 |
| IDL-apoB (mmol/L) | 10 (7.5, 11) | 11 (9.4, 13) | 11 (9.7, 12) | 11 (7.5, 13) | 8.7 (6.7, 11) | 7.8 (6.2, 13) | 11 (7.5, 15) | 14 (7.9, 17) | p = 0.65 |
| LDL-C (mmol/L) | 2.6 (2.0, 2.8) | 2.0 (1.7, 3.0) | 2.5 (1.7, 2.8) | 2.4 (1.9, 2.8) | 2.0 (1.8, 2.3) | 2.2 (2.0, 2.5) | 2.2 (1.8, 3.1) | 2.3 (1.7, 2.8) | p = 0.75 |
| LDL-TG (mmol/L) | 0.17 (0.11, 0.21) | 0.16 (0.13, 0.23) | 0.18 (0.15, 0.20) | 0.17 (0.15, 0.20) | 0.14 (0.11, 0.18) | 0.13 (0.11, 0.16) | 0.17 (0.14, 0.22) | 0.17 (0.14, 0.21) | p = 0.64 |
| LDL-apoB (mmol/L) | 113 (70.3, 121) | 99.5 (61.1, 118) | 94.6 (64.3, 118) | 115 (75.6, 123) | 68.0 (43.9, 98.2) | 77.1 (57.4, 120) | 107 (77.7, 146) | 102 (84.9, 135) | p = 0.92 |
| HDL-C (mmol/L) | 1.2 (0.94, 1.4) | 1.3 (0.98, 1.6) | 1.1 (0.89, 1.2) | 1.2 (0.96, 1.4) | 1.2 (0.94, 1.6) | 1.2 (1.0, 1.5) | 1.2 (1.0, 1.3) | 1.2 (0.99, 1.5) | p = 0.26 |
| HDL2-C (mmol/L) | 0.59 (0.41, 0.62) | 0.59 (0.45, 0.71) | 0.49 (0.35, 0.55) | 0.56 (0.47, 0.71) | 0.61 (0.38, 0.77) | 0.65 (0.40, 0.79) | 0.54 (0.40, 0.75) | 0.66 (0.44, 0.77) | p = 0.98 |
| HDL3-C (mmol/L) | 0.65 (0.51, 0.79) | 0.73 (0.52, 0.83) | 0.62 (0.54, 0.68) | 0.61 * (0.50, 0.68) | 0.63 (0.57, 0.77) | 0.60 * (0.55, 0.71) | 0.61 (0.52, 0.76) | 0.62 (0.49, 0.78) | R2 = 0.01, p = 0.04 |
| 0 g/day (n = 12) 0 weeks | 0 g/day (n = 12) 8 weeks | 0.35 g/day (n = 15) 0 weeks | 0.35 g/day (n = 15) 8 weeks | 0.7 g/day (n = 13) 0 weeks | 0.7 g/day (n = 13) 8 weeks | 1 g/day (n = 13) 0 weeks | 1 g/day (n = 13) 8 weeks | Dose Effect (R2, p Value) | |
|---|---|---|---|---|---|---|---|---|---|
| LDL Radius (nm) | 10.9 (10.8, 11.0) | 10.9 (10.7, 10.9) | 11.0 (10.4, 11.1) | 10.9 (10.4, 11.1) | 10.9 (10.6, 11.1) | 10.7 (10.5, 11.0) | 10.9 (10.7, 11.0) | 10.8 (10.8, 11.3) | p = 0.15 |
| % HDL2b | 37 (32, 43) | 33 (31, 44) | 35 (16, 38) | 35 (23, 38) | 41 (28, 49) | 46 (38, 50) | 32 (28, 47) | 42 (27, 52) | R2 = 0.05, p = 0.04 |
| % HDL2a | 30 (27, 33) | 28 (26, 37) | 24 (22, 31) | 25 (22, 31) | 26 (21, 35) | 26 (21, 32) | 31 (22, 33) | 31 (24, 33) | p = 0.22 |
| % HDL3a | 19 (15, 23) | 20 (15, 23) | 20 (18, 29) | 19 (17, 27) | 19 (15, 25) | 18 (14, 22) | 18 (16, 20) | 17 (13, 19) | p = 0.24 |
| % HDL3b | 12 (7, 14) | 11 (7, 16) | 13 (10, 20) | 13 (10, 18) | 11 (7, 14) | 9 (2, 12) | 17 (11, 18) | 9 (8, 19) | R2 = 0.02, p = 0.07 |
| % HDL3c | 0.8 (0.0, 3.3) | 0.3 (0.0, 3.9) | 6.0 (0.6, 11.5) | 6.1 (0.0, 12.5) | 1.2 (0.0, 8.2) | 1.9 (0.0, 8.2) | 1.2 (0.6, 3.0) | 0.6 (0.0, 2.1) | p = 0.10 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sparkes, C.; Gibson, R.; Sinclair, A.; Else, P.L.; Meyer, B.J. Effect of Low Dose Docosahexaenoic Acid-Rich Fish Oil on Plasma Lipids and Lipoproteins in Pre-Menopausal Women: A Dose–Response Randomized Placebo-Controlled Trial. Nutrients 2018, 10, 1460. https://doi.org/10.3390/nu10101460
Sparkes C, Gibson R, Sinclair A, Else PL, Meyer BJ. Effect of Low Dose Docosahexaenoic Acid-Rich Fish Oil on Plasma Lipids and Lipoproteins in Pre-Menopausal Women: A Dose–Response Randomized Placebo-Controlled Trial. Nutrients. 2018; 10(10):1460. https://doi.org/10.3390/nu10101460
Chicago/Turabian StyleSparkes, Cassandra, Robert Gibson, Andrew Sinclair, Paul L. Else, and Barbara J. Meyer. 2018. "Effect of Low Dose Docosahexaenoic Acid-Rich Fish Oil on Plasma Lipids and Lipoproteins in Pre-Menopausal Women: A Dose–Response Randomized Placebo-Controlled Trial" Nutrients 10, no. 10: 1460. https://doi.org/10.3390/nu10101460
APA StyleSparkes, C., Gibson, R., Sinclair, A., Else, P. L., & Meyer, B. J. (2018). Effect of Low Dose Docosahexaenoic Acid-Rich Fish Oil on Plasma Lipids and Lipoproteins in Pre-Menopausal Women: A Dose–Response Randomized Placebo-Controlled Trial. Nutrients, 10(10), 1460. https://doi.org/10.3390/nu10101460