Bee Collected Pollen with Enhanced Health Benefits, Produced by Fermentation with a Kombucha Consortium

 ,
,  , ,
, , 
Abstract
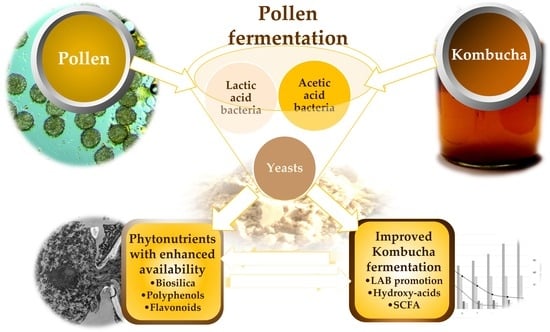
1. Introduction
2. Materials and Methods
2.1. Biological Material
2.2. Pollen Fermentation with the Kombucha Consortium
2.3. Pollen Biosilica Solubilization
2.4. Determination of the Total Phenolic and Flavonoid Compounds
2.5. Antioxidant Activity
2.6. Dynamics of Lactic Acid Bacteria Population During Product Fermentation
2.6.1. Real Time PCR (qPCR) Technique
2.6.2. On-plate LAB Counting
2.7. Analysis of Organic Acids
2.7.1. Analysis of Hydroxy-Acids
2.7.2. Analysis of Short Chains Fatty Acids
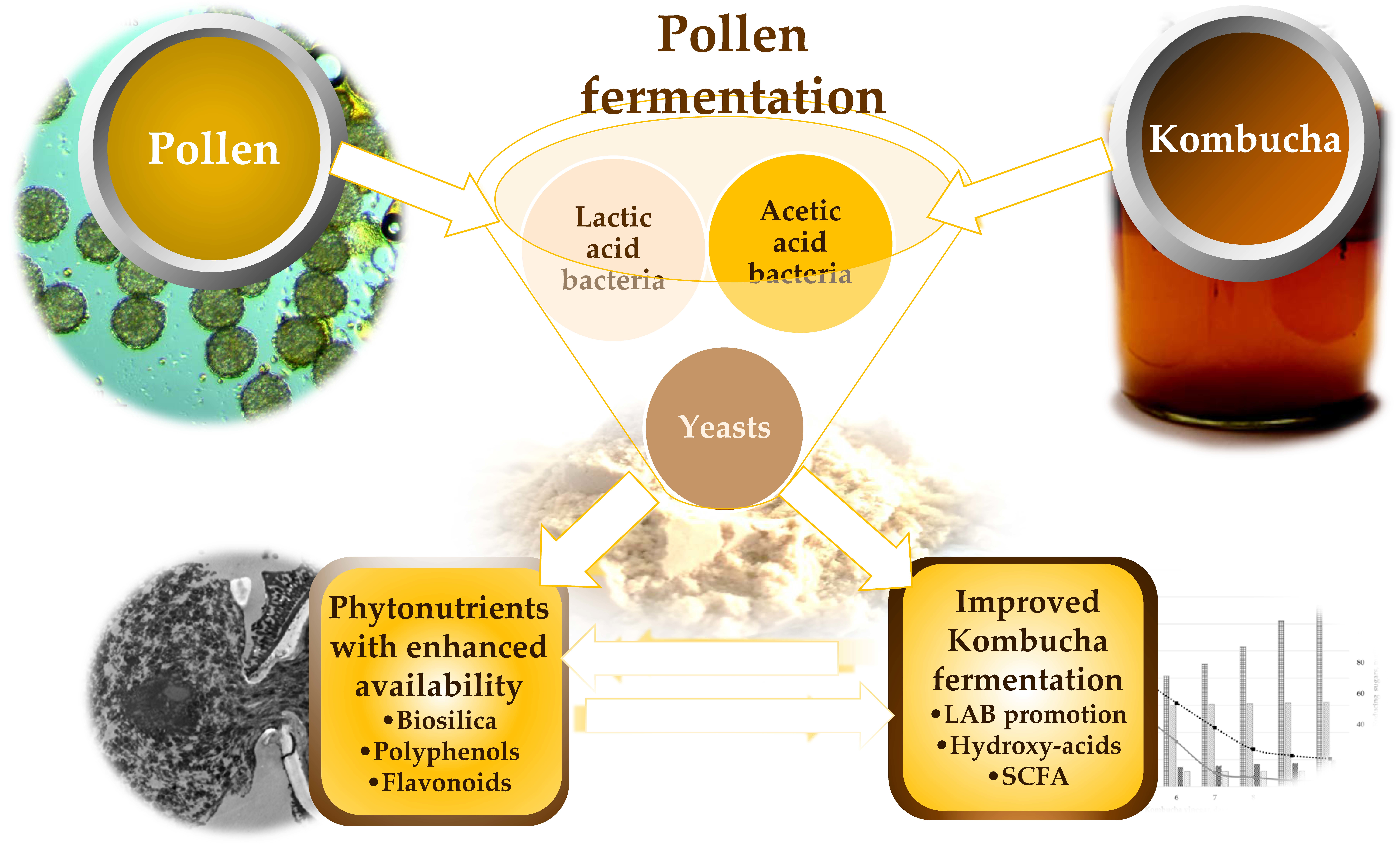
2.8. Ultrastructural and Morphological Analysis
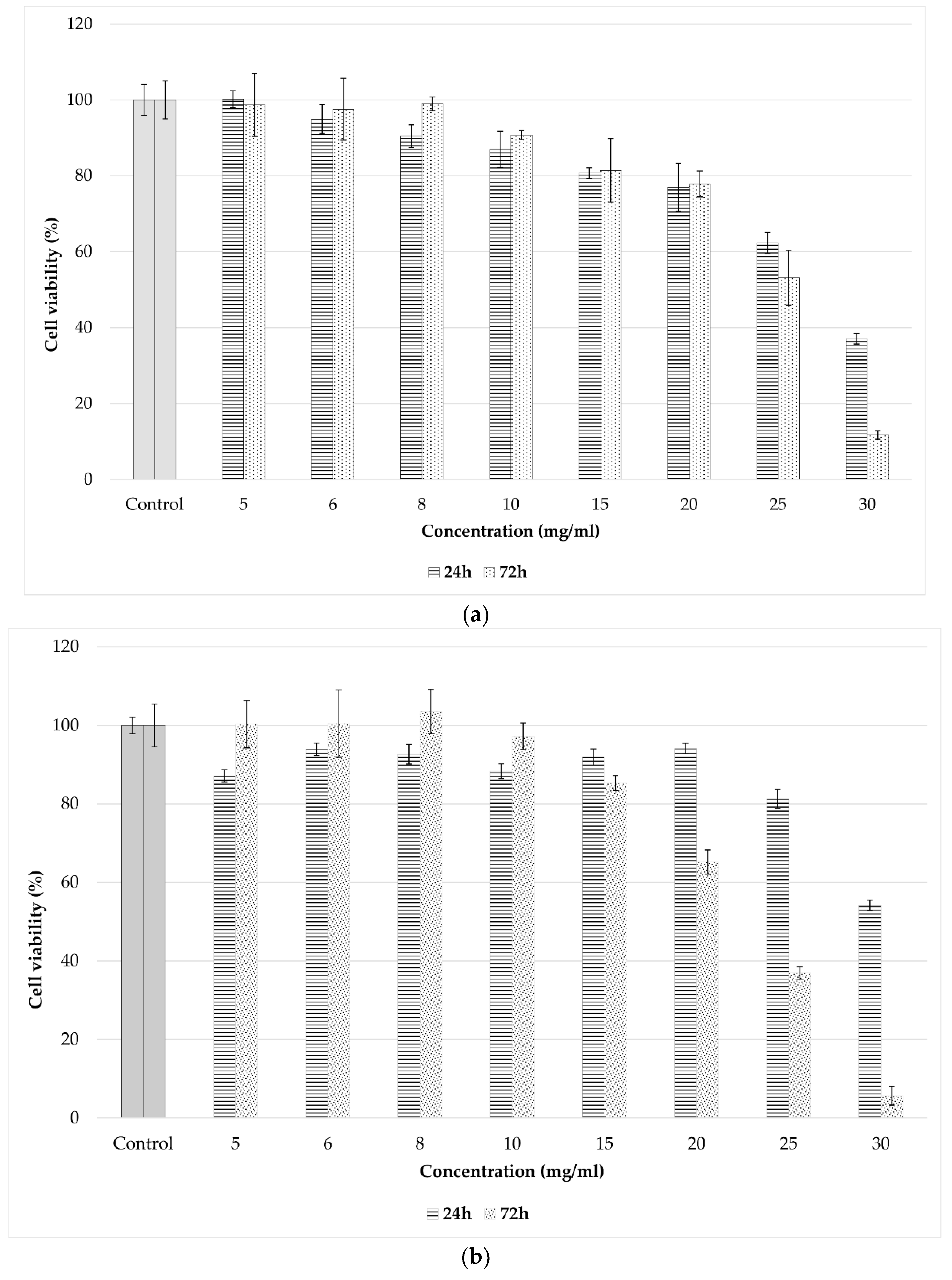
2.9. Biocompatibility and Antitumoral Activity
2.9.1. Cell Viability
2.9.2. Cell Morphology
2.10. Statistical Analysis
3. Results
3.1. Release of Pollen Biosilica, Polyphenols and Flavonoids During Fermentation with Kombucha
3.2. Dynamic of Lactic Acid Bacteria Population During Product Fermentation
3.3. Dynamics of Organic Acids During Product Fermentation
3.4. Morphological and Structural Analysis of Fermented Pollen
3.5. Cytotoxic and Antitumoral Effects of Spray-Dried Product
Cell Morphology
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Denisow, B.; Denisow-Pietrzyk, M. Biological and therapeutic properties of bee pollen: A review. J. Sci. Food Agric. 2016, 96, 4303–4309. [Google Scholar] [CrossRef] [PubMed]
- Pascoal, A.; Rodrigues, S.; Teixeira, A.; Feás, X.; Estevinho, L.M. Biological activities of commercial bee pollens: Antimicrobial, antimutagenic, antioxidant and anti-inflammatory. Food Chem. Toxicol. 2014, 63, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Cornara, L.; Biagi, M.; Xiao, J.; Burlando, B. Therapeutic properties of bioactive compounds from different honeybee products. Front. Pharmacol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Yan, Y.; Mi, J.; Zhang, H.; Lu, L.; Luo, Q.; Li, X.; Zeng, X.; Cao, Y. Simulated digestion and fermentation in vitro by human gut microbiota of polysaccharides from bee collected pollen of chinese wolfberry. J. Agric. Food Chem. 2018, 66, 898–907. [Google Scholar] [CrossRef] [PubMed]
- Zuluaga, C.M.; Serrato, J.C.; Quicazan, M.C. Bee-pollen structure modification by physical and biotechnological processing: Influence on the availability of nutrients and bioactive compounds. In Proceedings of the 12th International Conference on Chemical & Process Engineering (ICHEAP12), Milano, Italy, 19–22 May 2015; Aidic Servizi Srl: Milano, Italy, 2015; Volume 43, pp. 79–84. [Google Scholar]
- Mackenzie, G.; Boa, A.N.; Diego-Taboada, A.; Atkin, S.L.; Sathyapalan, T. Sporopollenin, the least known yet toughest natural biopolymer. Front. Mater. 2015, 2. [Google Scholar] [CrossRef]
- Roulston, T.A.H.; Cane, J.H. Pollen nutritional content and digestibility for animals. Plant Syst. Evol. 2000, 222, 187–209. [Google Scholar] [CrossRef]
- Feng, Z.-F.; Chen, X.-F.; Di, D.-L. Online extraction and isolation of highly polar chemical constituents from brassica napus l. Pollen by high shear technique coupled with high-performance counter-current chromatography. J. Sep. Sci. 2012, 35, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.-S.; Suh, H.-J.; Chung, I.K. Enhanced extraction of bioactive compounds from bee pollen by wet-grinding technology. J. Life Sci. 2016, 26, 651–656. [Google Scholar] [CrossRef]
- Liu, X.-D.; Zhang, F.-B.; Zhou, B.; Shan, H.; Chen, P.-Y. Effect of sonication on different quality parameters of pinus massoniana pollen. Ultrason. Sonochem. 2015, 22, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Wang, K.; Qiao, J.; Dong, J.; Li, Z.; Zhang, H. Improving nutrient release of wall-disrupted bee pollen with a combination of ultrasonication and high shear technique. J. Sci. Food Agric. 2018. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Gao, K.; Wang, K.; Xu, X.; Zhang, H. Cell wall disruption of rape bee pollen treated with combination of protamex hydrolysis and ultrasonication. Food Res. Int. 2015, 75, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Sun, L.; Dong, J.; Zhang, H. Breaking the cells of rape bee pollen and consecutive extraction of functional oil with supercritical carbon dioxide. Innov. Food Sci. Emerg. Technol. 2009, 10, 42–46. [Google Scholar] [CrossRef]
- Kaskoniene, V.; Katileviciute, A.; Kaskonas, P.; Maruska, A. The impact of solid-state fermentation on bee pollen phenolic compounds and radical scavenging capacity. Chem. Pap. 2018, 72, 2115–2120. [Google Scholar] [CrossRef]
- Yang, S.Y.; Wu, Y.Y.; Luo, C.B.; Di, Z.H.; Wu, Y.P.; Zhang, J.; Xu, Y.M. A Bacillus coagulans and its application in pine pollen fermentation. Food Sci. Biotechnol. 2015, 24, 2129–2135. [Google Scholar] [CrossRef]
- Yang, S.Y.; Li, H.L. Optimization of pine pollen fermentation conditions using lactobacillus paracasei. Food Sci. Biotechnol. 2015, 24, 155–160. [Google Scholar] [CrossRef]
- Zhang, Z.; Cao, H.G.; Chen, C.; Chen, X.; Wei, Q.; Zhao, F.Y. Effects of fermentation by ganoderma lucidum and saccharomyces cerevisiae on rape pollen morphology and its wall. J. Food Sci. Technol. 2017, 54, 4026–4034. [Google Scholar] [CrossRef] [PubMed]
- Salazar-Gonzalez, C.; Diaz-Moreno, C. The nutritional and bioactive aptitude of bee pollen for a solid-state fermentation process. J. Apic. Res. 2016, 55, 161–175. [Google Scholar] [CrossRef]
- Tomás, A.; Falcão, S.I.; Russo-Almeida, P.; Vilas-Boas, M. Potentialities of beebread as a food supplement and source of nutraceuticals: Botanical origin, nutritional composition and antioxidant activity. J. Apic. Res. 2017, 56, 219–230. [Google Scholar] [CrossRef]
- Kieliszek, M.; Piwowarek, K.; Kot, A.M.; Błażejak, S.; Chlebowska-Śmigiel, A.; Wolska, I. Pollen and bee bread as new health-oriented products: A review. Trends Food Sci. Technol. 2018, 71, 170–180. [Google Scholar] [CrossRef]
- Vasquez, A.; Olofsson, T.C. The lactic acid bacteria involved in the production of bee pollen and bee bread. J. Apic. Res. 2009, 48, 189–195. [Google Scholar] [CrossRef]
- Gilliam, M. Microbiology of pollen and bee bread: The yeasts. Apidologie 1979, 10, 43–53. [Google Scholar] [CrossRef]
- Gilliam, M. Microbiology of pollen and bee bread: The genus bacillus. Apidologie 1979, 10, 269–274. [Google Scholar] [CrossRef]
- Amores-Arrocha, A.; Roldan, A.; Jimenez-Cantizano, A.; Caro, I.; Palacios, V. Effect on white grape must of multiflora bee pollen addition during the alcoholic fermentation process. Molecules 2018, 23, 1321. [Google Scholar] [CrossRef] [PubMed]
- Zlatev, Z.; Taneva, I.; Baycheva, S.; Petev, M. A comparative analysis of physico-chemical indicators and sensory characteristics of yogurt with added honey and bee pollen. Bulg. J. Agric. Sci. 2018, 24, 132–144. [Google Scholar]
- Dima, S.-O.; Panaitescu, D.-M.; Orban, C.; Ghiurea, M.; Doncea, S.-M.; Fierascu, R.C.; Nistor, C.L.; Alexandrescu, E.; Nicolae, C.-A.; Trică, B.; et al. Bacterial nanocellulose from side-streams of kombucha beverages production: Preparation and physical-chemical properties. Polymers 2017, 9, 374. [Google Scholar] [CrossRef]
- Lončar, E.; Djurić, M.; Malbaša, R.; Kolarov, L.J.; Klašnja, M. Influence of working conditions upon kombucha conducted fermentation of black tea. Food Bioprod. Process. 2006, 84, 186–192. [Google Scholar] [CrossRef]
- Jayabalan, R.; Malbaša, R.V.; Lončar, E.S.; Vitas, J.S.; Sathishkumar, M. A review on kombucha tea-microbiology, composition, fermentation, beneficial effects, toxicity, and tea fungus. Compr. Rev. Food Sci. Food Saf. 2014, 13, 538–550. [Google Scholar] [CrossRef]
- Vīna, I.; Semjonovs, P.; Linde, R.; Deniņa, I. Current evidence on physiological activity and expected health effects of kombucha fermented beverage. J. Med. Food 2014, 17, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Martínez Leal, J.; Valenzuela Suárez, L.; Jayabalan, R.; Huerta Oros, J.; Escalante-Aburto, A. A review on health benefits of kombucha nutritional compounds and metabolites. CyTA J. Food 2018, 16, 390–399. [Google Scholar] [CrossRef]
- Marsh, A.J.; O’Sullivan, O.; Hill, C.; Ross, R.P.; Cotter, P.D. Sequence-based analysis of the bacterial and fungal compositions of multiple kombucha (tea fungus) samples. Food Microbiol. 2014, 38, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Matei, B.; Salzat, J.; Diguta, F.C.; Cornea, P.C.; Luta, G.; Utoiu, R.E.; Matei, F. Lactic acid bacteria strains isolated from kombucha with potential probiotic effect. Romanian Biotechnol. Lett. 2018, 23, 13592–13598. [Google Scholar]
- Chakravorty, S.; Bhattacharya, S.; Chatzinotas, A.; Chakraborty, W.; Bhattacharya, D.; Gachhui, R. Kombucha tea fermentation: Microbial and biochemical dynamics. Int. J. Food Microbiol. 2016, 220, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Kozyrovska, N.O.; Reva, O.M.; Goginyan, V.B.; Vera, J.-P.D. Kombucha microbiome as a probiotic: A view from the perspective of post-genomics and synthetic ecology. Biopolym. Cell 2012, 28, 103–113. [Google Scholar] [CrossRef]
- Aguilar-Toalá, J.E.; Garcia-Varela, R.; Garcia, H.S.; Mata-Haro, V.; González-Córdova, A.F.; Vallejo-Cordoba, B.; Hernández-Mendoza, A. Postbiotics: An evolving term within the functional foods field. Trends Food Sci. Technol. 2018, 75, 105–114. [Google Scholar] [CrossRef]
- Gill, P.A.; van Zelm, M.C.; Muir, J.G.; Gibson, P.R. Short chain fatty acids as potential therapeutic agents in human gastrointestinal and inflammatory disorders. Aliment. Pharmacol. Ther. 2018, 48, 15–34. [Google Scholar] [CrossRef] [PubMed]
- Crang, R.E.; May, G. Evidence for silicon as a prevalent elemental component in pollen wall structure. Can. J. Bot. 1974, 52, 2171–2174. [Google Scholar] [CrossRef]
- Exley, C. A possible mechanism of biological silicification in plants. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Guerriero, G.; Hausman, J.-F.; Legay, S. Silicon and the plant extracellular matrix. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Jugdaohsingh, R. Silicon and bone health. J. Nutr. Health Aging 2007, 11, 99–110. [Google Scholar] [PubMed]
- Price, C.T.; Koval, K.J.; Langford, J.R. Silicon: A review of its potential role in the prevention and treatment of postmenopausal osteoporosis. Int. J. Endocrinol. 2013. [Google Scholar] [CrossRef] [PubMed]
- Rodella, L.F.; Bonazza, V.; Labanca, M.; Lonati, C.; Rezzani, R. A review of the effects of dietary silicon intake on bone homeostasis and regeneration. J. Nutr. Health Aging 2014, 18, 820–826. [Google Scholar] [CrossRef] [PubMed]
- Jurkic, L.M.; Cepanec, I.; Pavelic, S.K.; Pavelic, K. Biological and therapeutic effects of ortho-silicic acid and some ortho-silicic acid-releasing compounds: New perspectives for therapy. Nutr. Metab. 2013, 10. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, F.H. Update on the possible nutritional importance of silicon. J. Trace Elements Med. Biol. 2014, 28, 379–382. [Google Scholar] [CrossRef] [PubMed]
- Barth, O.M.; Freitas, A.S.; Oliveira, E.S.; Silva, R.A.; Maester, F.M.; Andrella, R.R.S.; Cardozo, G. Evaluation of the botanical origin of commercial dry bee pollen load batches using pollen analysis: A proposal for technical standardization. An. Acad. Bras. Cienc. 2010, 82, 893–902. [Google Scholar] [CrossRef] [PubMed]
- Maurizio, A.; Louveaux, J. Pollens de Plantes Mellifères D’Europe; Union des Groupements Apicoles Français: Paris, France, 1965. [Google Scholar]
- Ricciardelli, D.A.G. Textbook of Melissopalynology; Apimondia Publishing House: Bucharest, Romania, 1997. [Google Scholar]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Hauptkorn, S.; Pavel, J.; Seltner, H. Determination of silicon in biological samples by icp-oes after non-oxidative decomposition under alkaline conditions. Fresenius J. Anal. Chem. 2001, 370, 246–250. [Google Scholar] [CrossRef] [PubMed]
- Coradin, T.; Eglin, D.; Livage, J. The silicomolybdic acid spectrophotometric method and its application to silicate/biopolymer interaction studies. J. Spectrosc. 2004, 18, 567–576. [Google Scholar] [CrossRef]
- Huang, D.; Ou, B.; Prior, R.L. The chemistry behind antioxidant capacity assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef] [PubMed]
- Craciunescu, O.; Constantin, D.; Gaspar, A.; Toma, L.; Utoiu, E.; Moldovan, L. Evaluation of antioxidant and cytoprotective activities of Arnica montana L. and Artemisia absinthium L. ethanolic extracts. Chem. Cent. J. 2012, 6. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-C.; Yang, M.-H.; Wen, H.-M.; Chern, J.-C. Estimation of total flavonoid content in propolis by two complementary colorimetric methods. J. Food Drug Anal. 2002, 10, 178–182. [Google Scholar]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved abts radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Ritchie, L.E.; Burke, K.F.; Garcia-Mazcorro, J.F.; Steiner, J.M.; Suchodolski, J.S. Characterization of fecal microbiota in cats using universal 16s rRNA gene and group-specific primers for Lactobacillus and Bifidobacterium spp. Vet. Microbiol. 2010, 144, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Craciunescu, O.; Gaspar, A.; Trif, M.; Moisei, M.; Oancea, A.; Moldovan, L.; Zarnescu, O. Preparation and characterization of a collagen-liposome-chondroitin sulfate matrix with potential application for inflammatory disorders treatment. J. Nanomater. 2014, 2014, 1–9. [Google Scholar] [CrossRef]
- Spulber, R.; Dogaroglu, M.; Babeanu, N.; Popa, O. Physicochemical characteristics of fresh bee pollen from different botanical origins. Romanian Biotechnol. Lett. 2018, 23, 13357–13365. [Google Scholar]
- Mărghitaş, L.A.; Stanciu, O.G.; Dezmirean, D.S.; Bobiş, O.; Popescu, O.; Bogdanov, S.; Campos, M.G. In vitro antioxidant capacity of honeybee-collected pollen of selected floral origin harvested from Romania. Food Chem. 2009, 115, 878–883. [Google Scholar] [CrossRef]
- Altunatmaz, S.S.; Tarhan, D.; Aksu, F.; Barutçu, U.B.; Or, M.E. Mineral element and heavy metal (cadmium, lead and arsenic) levels of bee pollen in turkey. Food Sci. Technol. 2017, 37, 136–141. [Google Scholar] [CrossRef]
- Kolesnikov, M.; Gins, V. Flavonoids and silicon in certain plant pollen. Chem. Nat. Compd. 1999, 35, 520–523. [Google Scholar] [CrossRef]
- Faizul, C.P.; Abdullah, C.; Fazlul, B. Review of extraction of silica from agricultural wastes using acid leaching treatment. In Advanced Materials Engineering and Technology; Abdullah, M.M.A., Jamaludin, L., Razak, R.A., Yahya, Z., Hussin, K., Eds.; Trans Tech Publications Ltd.: Stafa-Zurich, Switzerland, 2012; Volume 626, pp. 997–1000. [Google Scholar]
- Jayabalan, R.; Marimuthu, S.; Swaminathan, K. Changes in content of organic acids and tea polyphenols during kombucha tea fermentation. Food Chem. 2007, 102, 392–398. [Google Scholar] [CrossRef]
- Sun, T.-Y.; Li, J.-S.; Chen, C. Effects of blending wheatgrass juice on enhancing phenolic compounds and antioxidant activities of traditional kombucha beverage. J. Food Drug Anal. 2015, 23, 709–718. [Google Scholar] [CrossRef] [PubMed]
- Mauriello, G.; Prisco, A.D.; Di Prisco, G.; La Storia, A.; Caprio, E.; Galdiero, M. Microbial characterization of bee pollen from the Vesuvius area collected by using three different traps. PLoS ONE 2017, 12, e0183208. [Google Scholar] [CrossRef] [PubMed]
- Corby-Harris, V.; Maes, P.; Anderson, K.E.; Gerardo, N.M. The bacterial communities associated with honey bee (Apis mellifera) foragers. PLoS ONE 2014, 9, e95056. [Google Scholar] [CrossRef] [PubMed]
- Vásquez, A.; Forsgren, E.; Fries, I.; Paxton, R.J.; Flaberg, E.; Szekely, L.; Olofsson, T.C.; Ahmed, N. Symbionts as major modulators of insect health: Lactic acid bacteria and honeybees. PLoS ONE 2012, 7, e33188. [Google Scholar] [CrossRef]
- Malbaša, R.; Lončar, E.; Djurić, M. Comparison of the products of kombucha fermentation on sucrose and molasses. Food Chem. 2008, 106, 1039–1045. [Google Scholar] [CrossRef]
- Chen, C.; Liu, B.Y. Changes in major components of tea fungus metabolites during prolonged fermentation. J. Appl. Microbiol. 2000, 89, 834–839. [Google Scholar] [CrossRef] [PubMed]
- Currie, H.A.; Perry, C.C. Silica in plants: Biological, biochemical and chemical studies. Ann. Bot. 2007, 100, 1383–1389. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, H.; Demadis, K.D.; Pokrovsky, O.S.; Koutsoukos, P.G. Modern views on desilicification: Biosilica and abiotic silica dissolution in natural and artificial environments. Chem. Rev. 2010, 110, 4656–4689. [Google Scholar] [CrossRef] [PubMed]
- Coton, M.; Pawtowski, A.; Taminiau, B.; Burgaud, G.; Deniel, F.; Coulloumme-Labarthe, L.; Fall, A.; Daube, G.; Coton, E. Unraveling microbial ecology of industrial-scale kombucha fermentations by metabarcoding and culture-based methods. FEMS Microbiol. Ecol. 2017, 93. [Google Scholar] [CrossRef] [PubMed]
- Villarreal-Soto, S.A.; Beaufort, S.; Bouajila, J.; Souchard, J.P.; Taillandier, P. Understanding kombucha tea fermentation: A review. J. Food Sci. 2018, 83, 580–588. [Google Scholar] [CrossRef] [PubMed]
- Cvetković, D.; Markov, S.; Djurić, M.; Savić, D.; Velićanski, A. Specific interfacial area as a key variable in scaling-up kombucha fermentation. J. Food Eng. 2008, 85, 387–392. [Google Scholar] [CrossRef]
- Endo, A.; Futagawa-Endo, Y.; Dicks, L.M. Isolation and characterization of fructophilic lactic acid bacteria from fructose-rich niches. Syst. Appl. Microbiol. 2009, 32, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Reva, O.N.; Zaets, I.E.; Ovcharenko, L.P.; Kukharenko, O.E.; Shpylova, S.P.; Podolich, O.V.; de Vera, J.-P.; Kozyrovska, N.O. Metabarcoding of the kombucha microbial community grown in different microenvironments. AMB Express 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Endo, A.; Maeno, S.; Tanizawa, Y.; Kneifel, W.; Arita, M.; Dicks, L.; Salminen, S. Fructophilic lactic acid bacteria, a unique group of fructose-fermenting microbes. Appl. Environ. Microbiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Filannino, P.; Di Cagno, R.; Addante, R.; Pontonio, E.; Gobbetti, M. Metabolism of fructophilic lactic acid bacteria isolated from Apis mellifera L. Bee-gut: A focus on the phenolic acids as external electron acceptors. Appl. Environ. Microbiol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Vajrabhaya, L.-O.; Korsuwannawong, S. Cytotoxicity evaluation of a thai herb using tetrazolium (MTT) and sulforhodamine b (SRB) assays. J. Anal. Sci. Technol. 2018, 9, 15. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fermentation Time (days) | TEAC (µg Trolox/mL) | DPPH (Inhibition Grade%/mL) | IC50 (mg/mL) |
|---|---|---|---|
| 1 | 8.83 ± 0.17 | 1.35 ± 0.1 | 15.16 |
| 2 | 3.54 ± 0.63 | 1.32 ± 0.13 | 13.76 |
| 3 | 3.9 ± 0.18 | 1.62 ± 0.04 | 14 |
| 4 | 4.21 ± 0.21 | 1.64 ± 0.03 | 12.6 |
| 5 | 5.61 ± 0.22 | 2.16 ± 0.06 | 14.96 |
| 6 | 7.8 ± 0.26 | 2.85 ± 0.07 | 13.76 |
| 7 | 7.23 ± 0.31 | 3.79 ± 0.07 | 11.97 |
| 8 | 16.24 ± 0.34 | 4.87 ± 0.04 | 11.95 |
| 9 | 20.94 ± 0.67 | 4.86 ± 0.11 | 11.68 |
| 10 | 22.95 ± 0.77 | 4.91 ± 0.11 | 10.56 |
| Sample | Day | Hydroxy-Acids | Short-Chain Fatty Acids | ||||
|---|---|---|---|---|---|---|---|
| Citric Acid, g/L | Gluconic Acid, g/L | Lactic Acid, g/L | Acetic Acid, g/L | Propionic Acid, g/L | butyric Acid, g/L | ||
| K Lab | 0 | ND | 0.545 ± 0.006 | 0.38 ± 0.01 | 0.375 ± 0.005 | ND | ND |
| 5 | ND | 0.56 ± 0.006 | 0.36 ± 0.006 | 0.605 ± 0.004 | ND | 0.14 ± 0.011 | |
| 9 | ND | 0.555 ± 0.014 | 0.36 ± 0.006 | 1.32 ± 0.03 | 0.12 ± 0.033 | 0.18 ± 0.014 | |
| 13 | ND | 0.94 ± 0.03 | 0.37 ± 0.006 | 2.32 ± 0.015 | 0.16 ± 0.024 | 0.28 ± 0.017 | |
| 17 | ND | 1.59 ± 0.043 | 0.375 ± 0.003 | 4.66 ± 0.025 | 0.24 ± 0.016 | 0.30 ± 0.021 | |
| KP Lab | 0 | 0.02 ± 0.005 | 2.795 ± 0.015 | 0.46 ± 0.015 | 0.415 ± 0.005 | 0.095 ± 0.012 | 0.12 ± 0.038 |
| 5 | 0.02 ± 0.005 | 3.155 ± 0.016 | 0.44 ± 0.01 | 0.515 ± 0.007 | 0.205 ± 0.034 | 0.44 ± 0.032 | |
| 9 | 0.045 ± 0.005 | 3.59 ± 0.090 | 0.57 ± 0.02 | 2.275 ± 0.013 | 0.31 ± 0.023 | 0.74 ± 0.028 | |
| 13 | 0.04 ± 0.003 | 2.545 ± 0.005 | 0.72 ± 0.01 | 2.25 ± 0.04 | 0.42 ± 0.014 | 1.06 ± 0.063 | |
| 17 | 0.055 ± 0.005 | 2.26 ± 0.02 | 0.77 ± 0.01 | 3.51 ± 0.11 | 0.56 ± 0.041 | 1.78 ± 0.054 | |
| K Pilot | 18 | ND | 2.645 ± 0.14 | 1.75 ± 0.15 | 17.64 ± 0.21 | 0.27 ± 0.032 | 0.84 ± 0.037 |
| KP Pilot | 18 | 0.09 ± 0.001 | 3.975 ± 0.32 | 10.265 ± 0.42 | 19.56 ± 0.18 | 0.66 ± 0.037 | 1.92 ± 0.033 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uțoiu, E.; Matei, F.; Toma, A.; Diguță, C.F.; Ștefan, L.M.; Mănoiu, S.; Vrăjmașu, V.V.; Moraru, I.; Oancea, A.; Israel-Roming, F.; et al. Bee Collected Pollen with Enhanced Health Benefits, Produced by Fermentation with a Kombucha Consortium. Nutrients 2018, 10, 1365. https://doi.org/10.3390/nu10101365
Uțoiu E, Matei F, Toma A, Diguță CF, Ștefan LM, Mănoiu S, Vrăjmașu VV, Moraru I, Oancea A, Israel-Roming F, et al. Bee Collected Pollen with Enhanced Health Benefits, Produced by Fermentation with a Kombucha Consortium. Nutrients. 2018; 10(10):1365. https://doi.org/10.3390/nu10101365
Chicago/Turabian StyleUțoiu, Elena, Florentina Matei, Agnes Toma, Camelia Filofteia Diguță, Laura Mihaela Ștefan, Sorin Mănoiu, Virgil Valeriu Vrăjmașu, Ionuț Moraru, Anca Oancea, Florentina Israel-Roming, and et al. 2018. "Bee Collected Pollen with Enhanced Health Benefits, Produced by Fermentation with a Kombucha Consortium" Nutrients 10, no. 10: 1365. https://doi.org/10.3390/nu10101365
APA StyleUțoiu, E., Matei, F., Toma, A., Diguță, C. F., Ștefan, L. M., Mănoiu, S., Vrăjmașu, V. V., Moraru, I., Oancea, A., Israel-Roming, F., Cornea, C. P., Constantinescu-Aruxandei, D., Moraru, A., & Oancea, F. (2018). Bee Collected Pollen with Enhanced Health Benefits, Produced by Fermentation with a Kombucha Consortium. Nutrients, 10(10), 1365. https://doi.org/10.3390/nu10101365