
Assessing the Potential of Sentinel-2 and Pléiades Data for the Detection of Prosopis and Vachellia spp. in Kenya


 ,
,  and
and
Abstract
:
1. Introduction
- (a)
- identify a robust and reliable method for differentiating Prosopis from native vegetation types (Vachellia spp.), including mixed classes;
- (b)
- produce reliable mapping products having good accuracies, validated with independent reference samples;
- (c)
- assess the novel Sentinel-2 sensor for tree species classification and its application in arid and semi-arid environments; and
- (d)
- assess the value of free-of-charge Sentinel-2 data, compared to commercial Pléiades data.
2. Materials and Methods
2.1. Study Area
2.2. Data
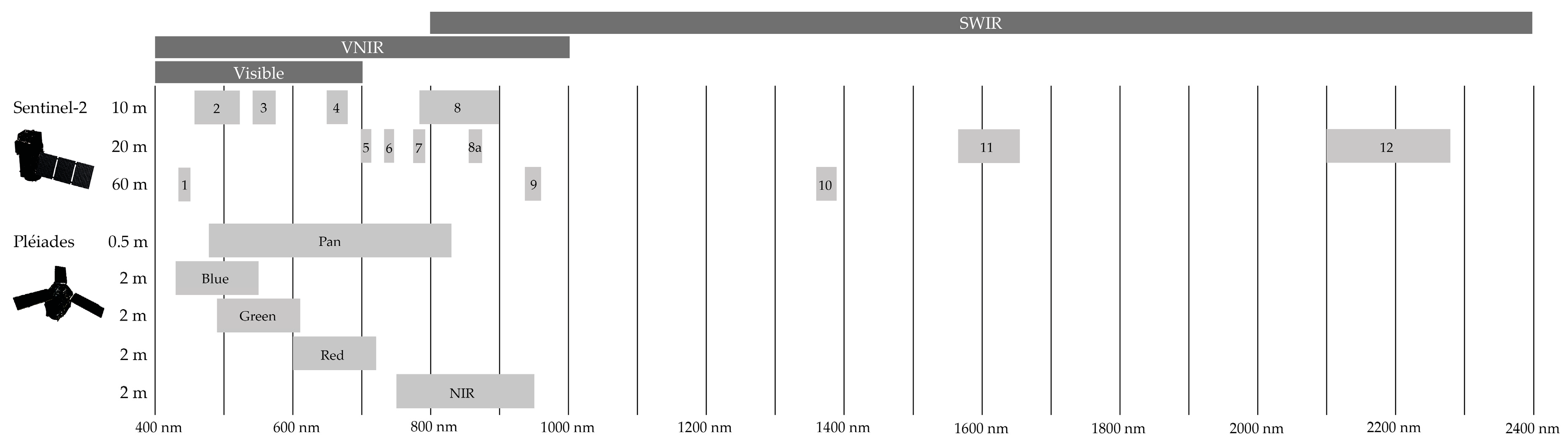
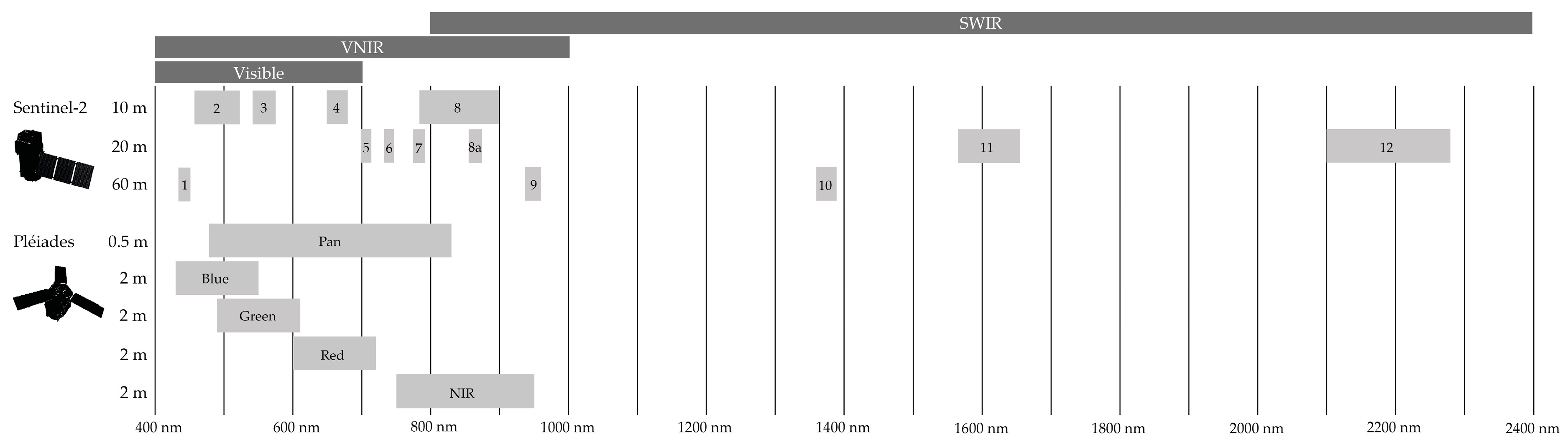
2.2.1. Satellite Data
2.2.2. Reference Data
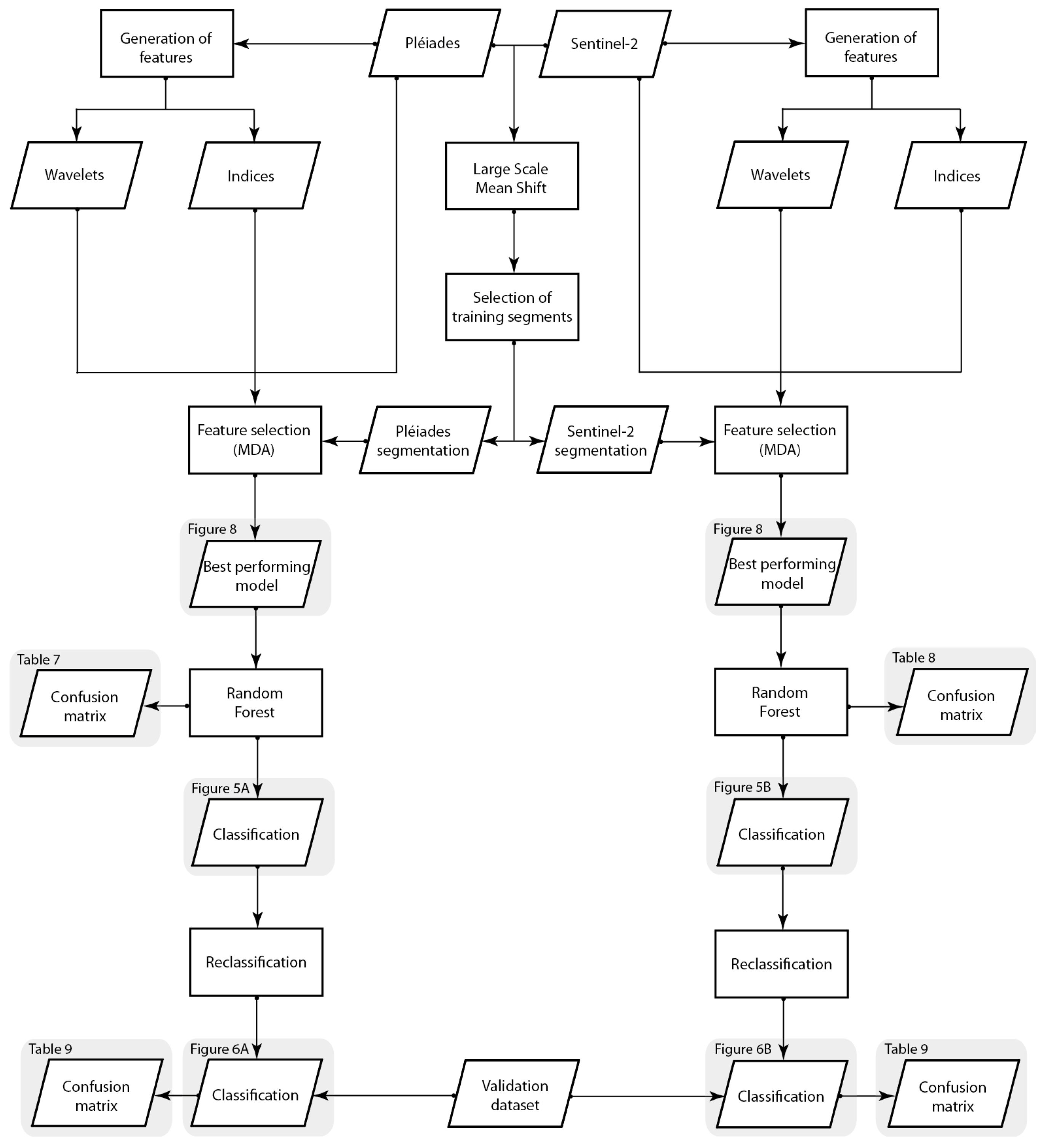
2.2.3. Segmentation
2.2.4. Spectral and Textural Features
2.2.5. Random Forest and Feature Selection
2.2.6. Accuracy Assessment
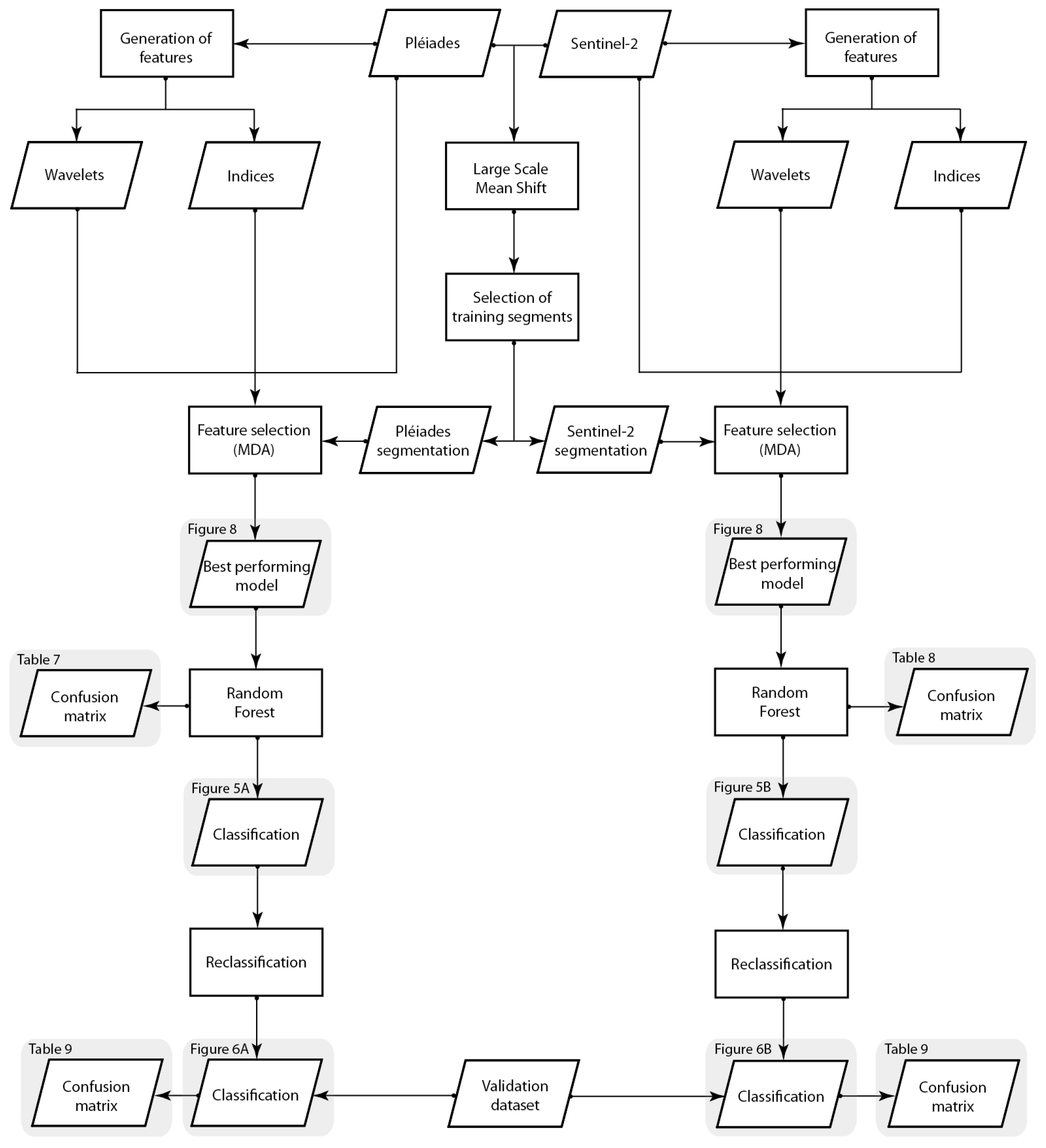
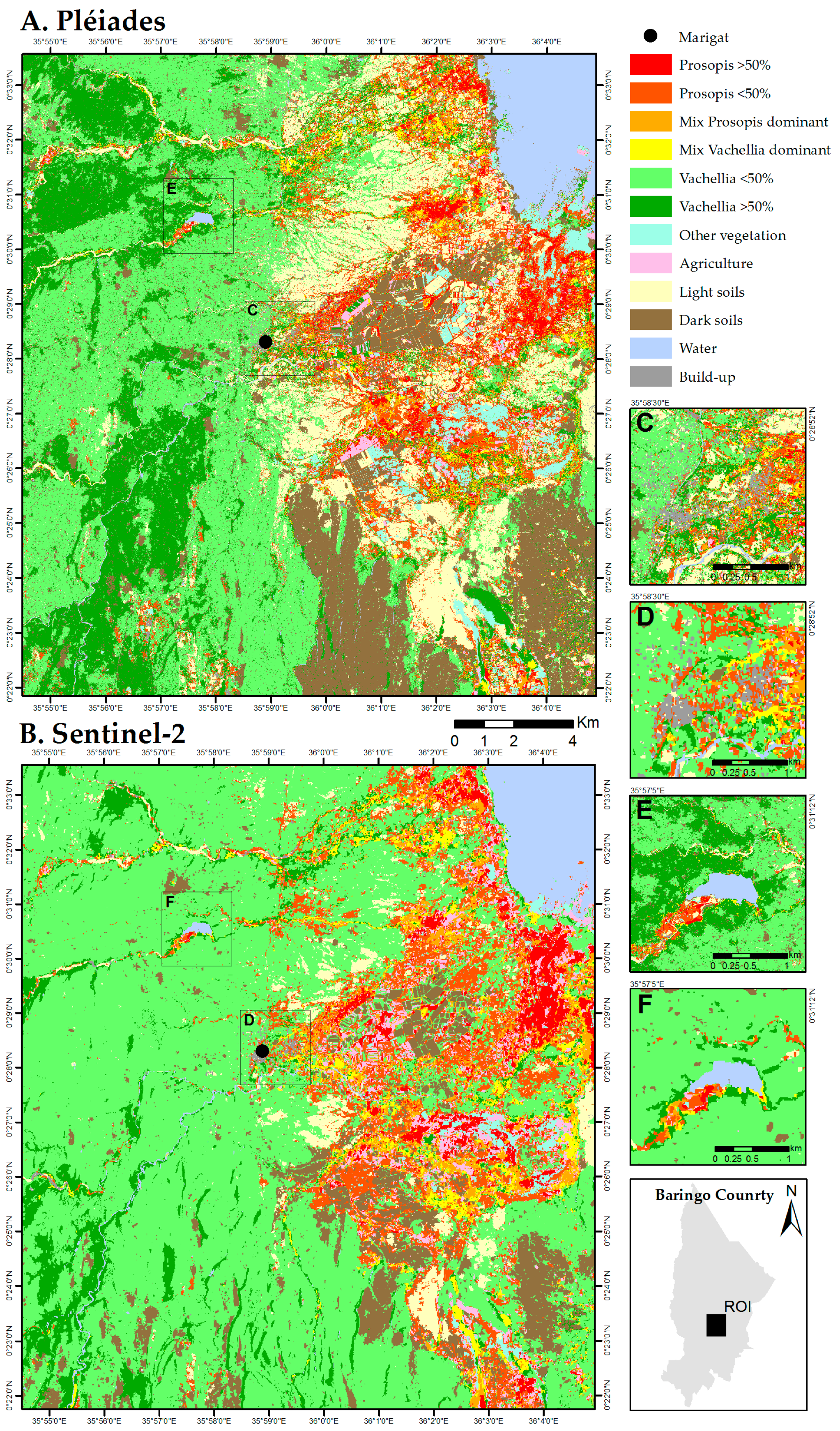
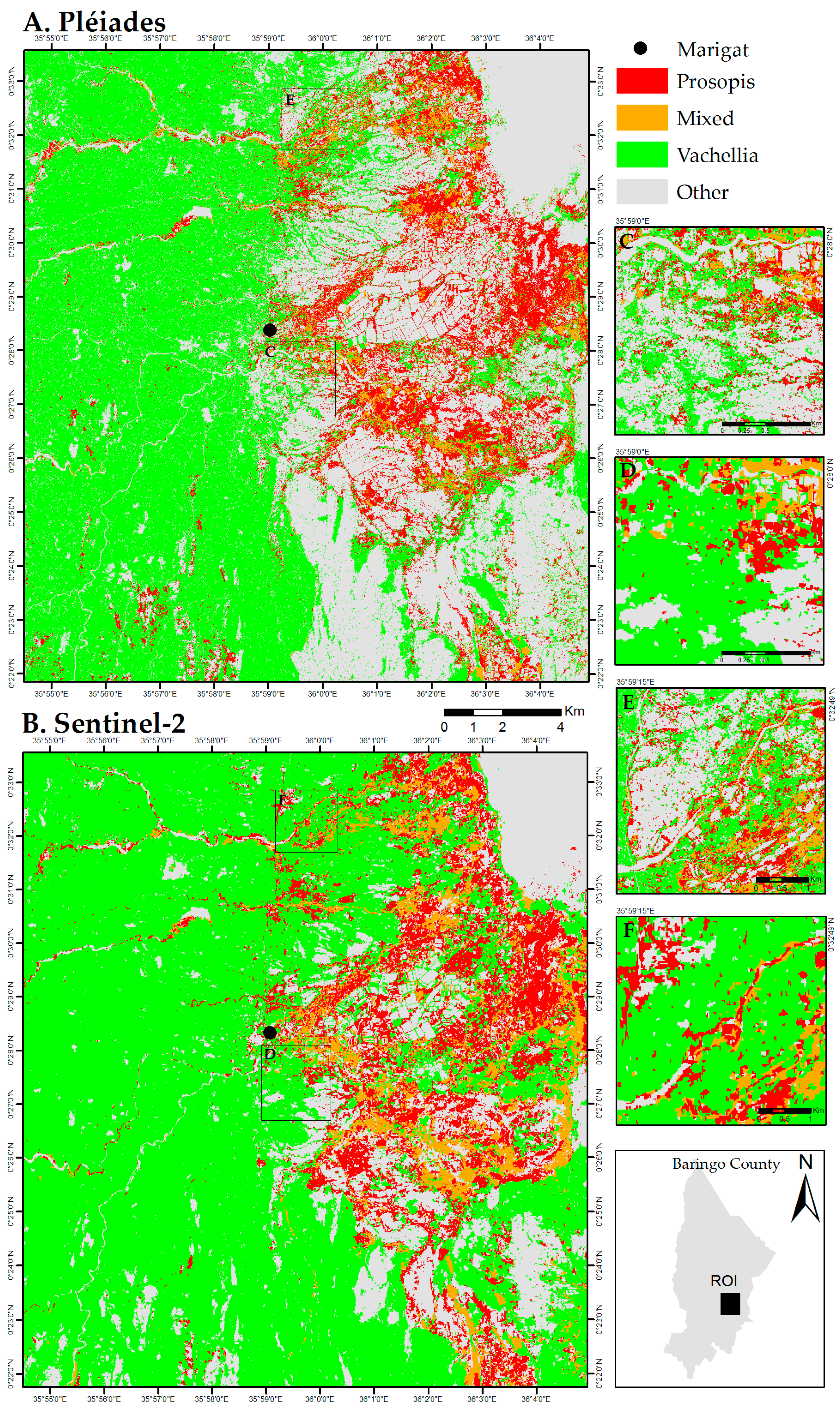
3. Results
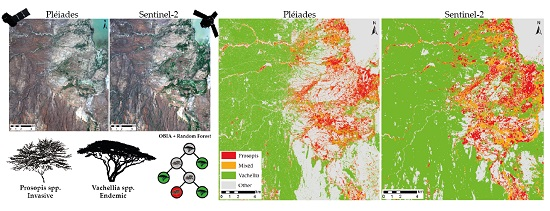
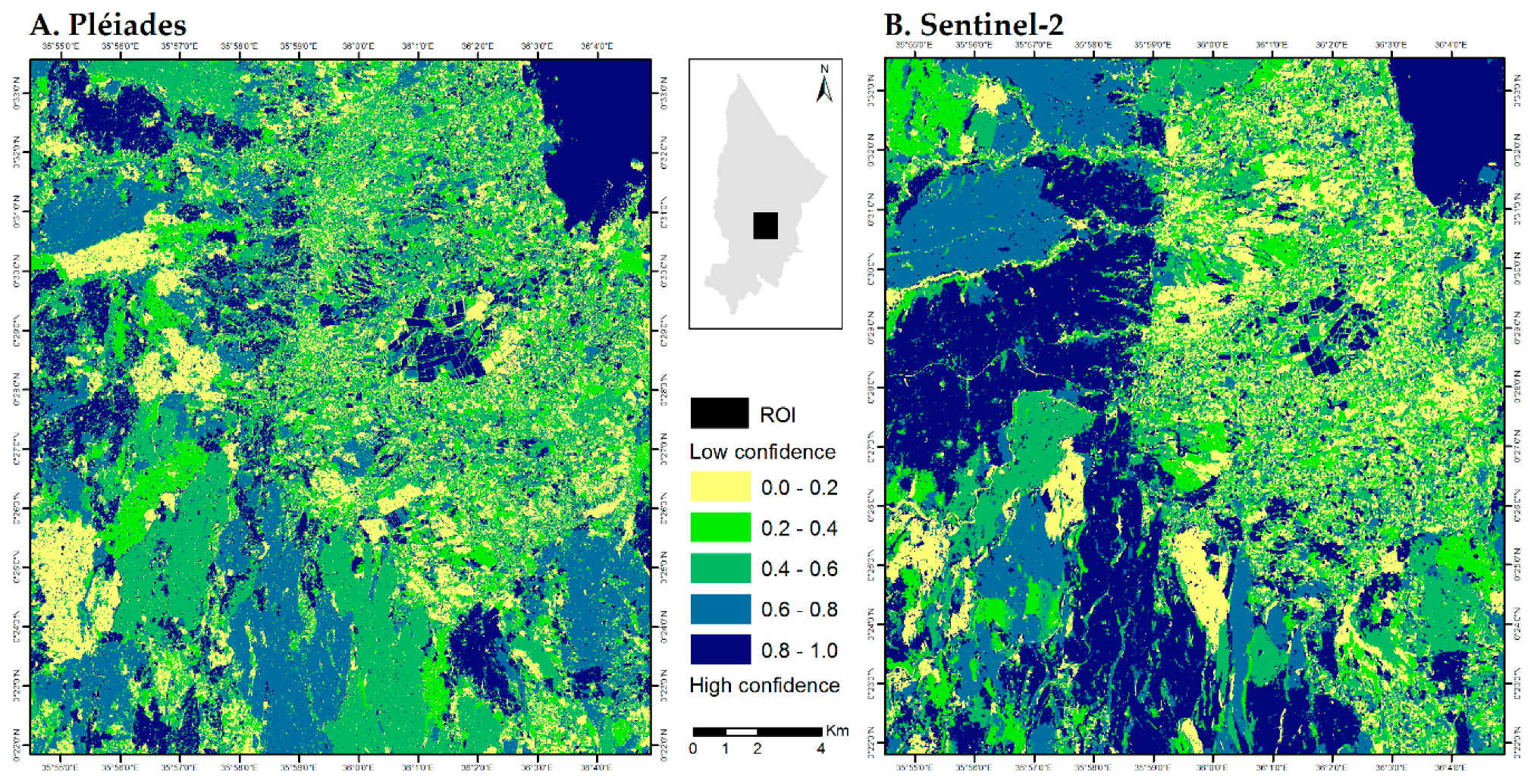
3.1. Prosopis and Vachellia Maps
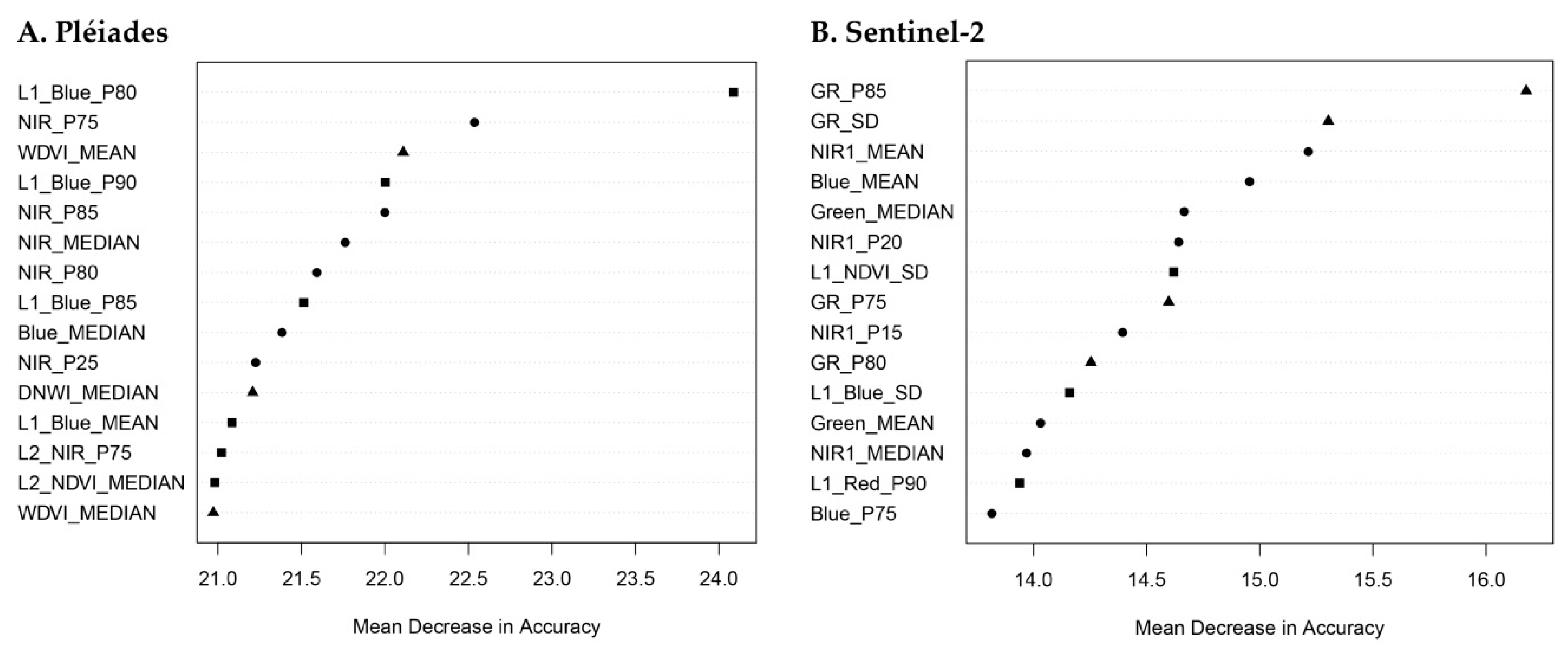
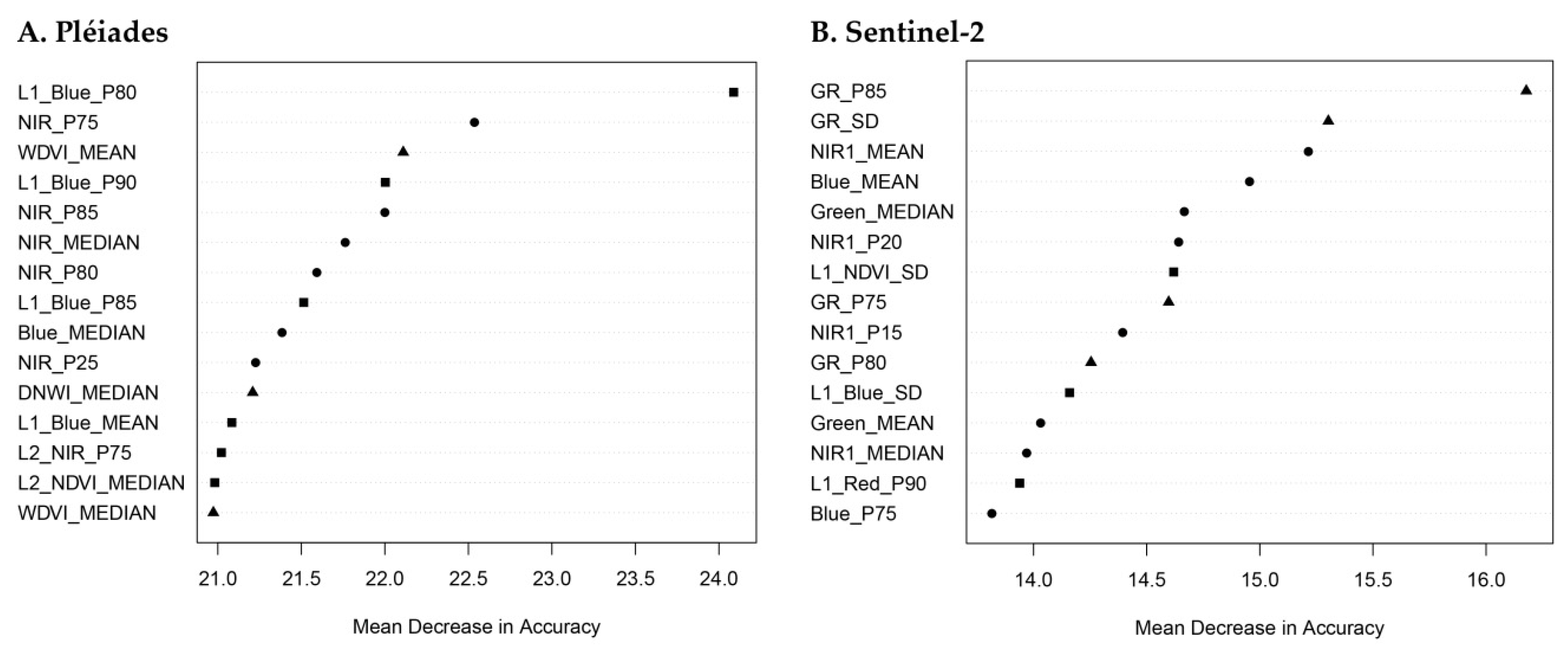
3.2. Feature Importance
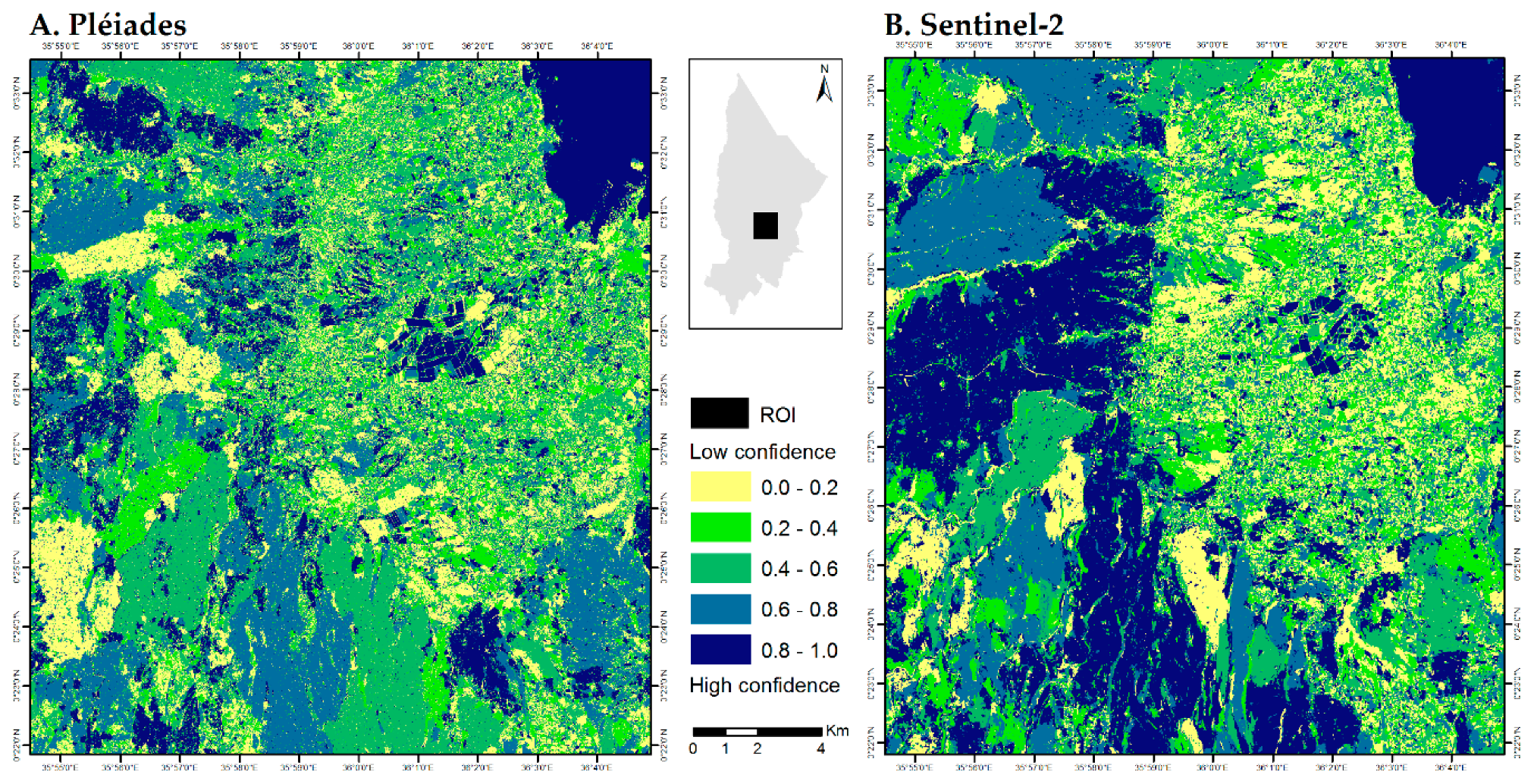
3.3. Accuracy Assessment
4. Discussion
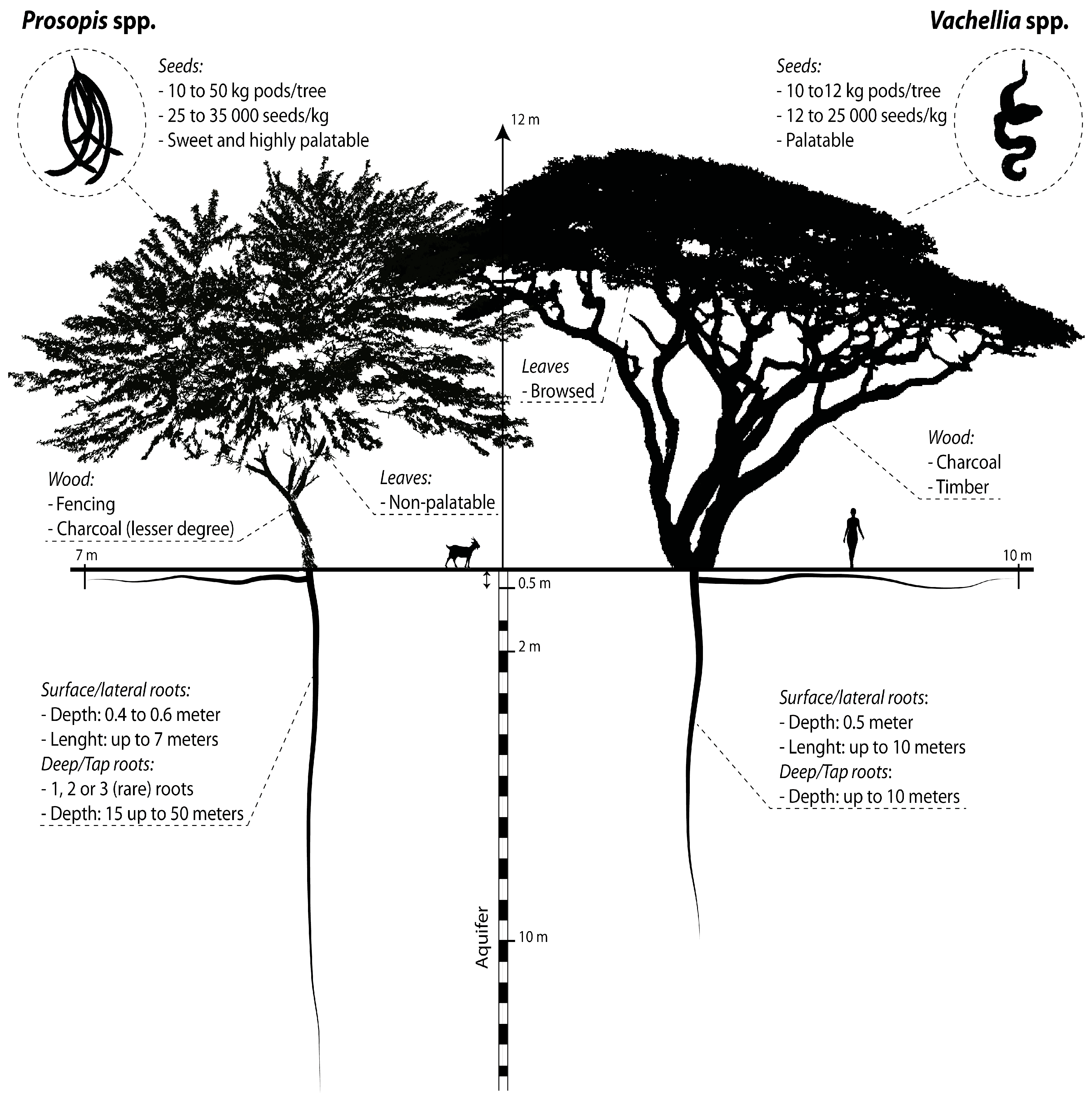
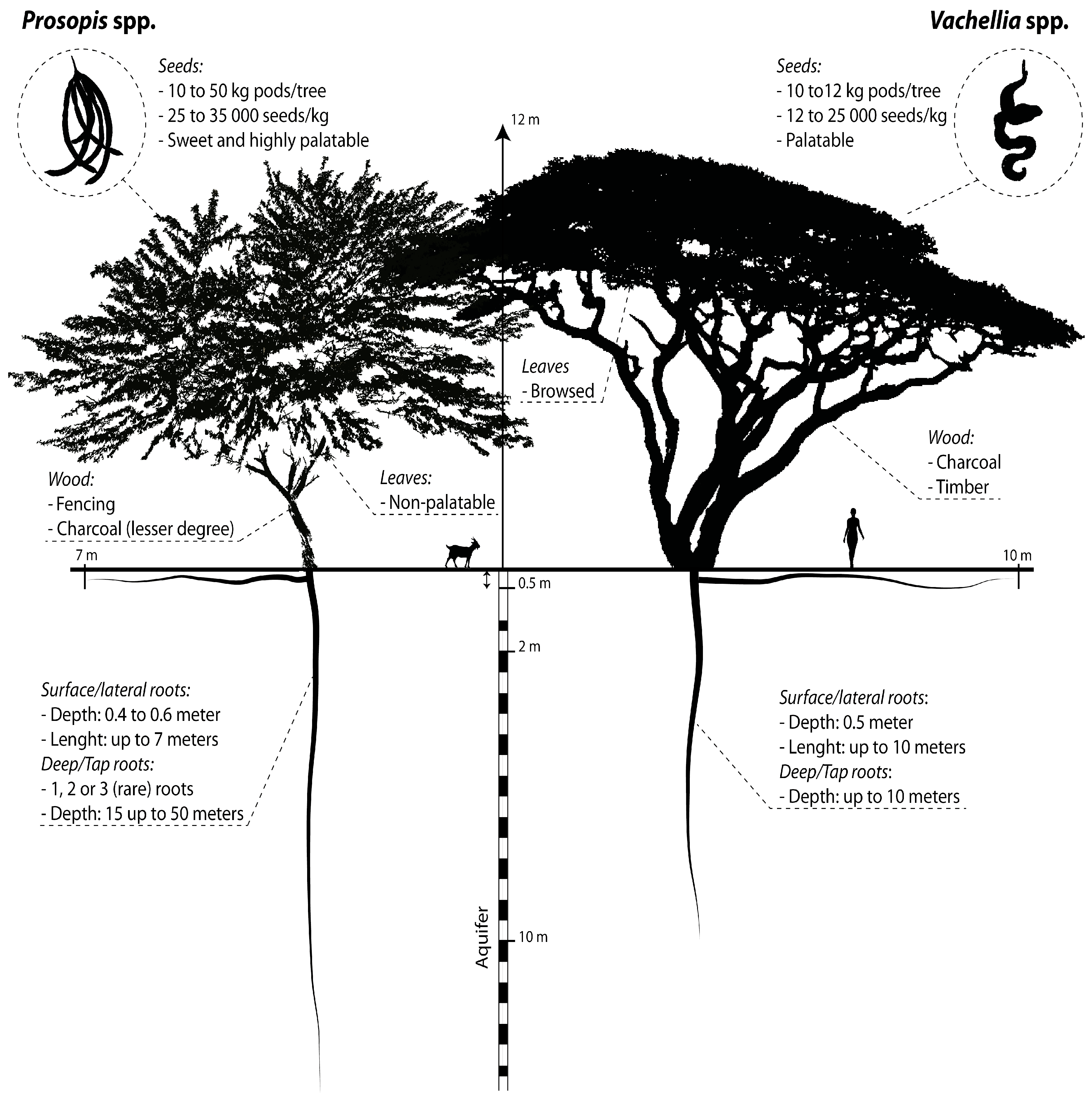
4.1. Map Product and Spatial Distribution
4.2. Map Comparison
4.2.1. Impact of Spatial Resolution
4.2.2. Impact of Spectral Resolution
4.2.3. Impact of Acquisition Dates
- Pléiades results show for example a higher proportion in the class Other, originating from the soil classes, as compared to Sentinel-2. There could be a relation between the larger exposure of bare soils and the utilization of both Prosopis and Vachellia for charcoal. In the years 2013–2015, the preliminary testing of the Cummins biogas power plant was initiated. The plant was established in Marigat to generate electricity from Prosopis, however, activities were discontinued the next year [103,104]. Hence, the increase in the sparsely vegetated classes in the Sentinel-2 data (2016) could be regrowth of the cleared area.
- Some of the Prosopis invaded areas visible in the 2015 (Pléiades) data were observed as cleared during the fieldwork. At the same time, some previously none invaded areas (mainly bare soils) had a Prosopis presence when visited in the field. Doing the fieldwork, we were able to determine from the height of Prosopis seedlings that they were half a year old, thus only detectable in one of the datasets (Sentinel-2).
- The sparsely vegetated Vachellia class and soil classes are often confused, because Vachellia exists as shrubs in some areas due to intense grazing. These misclassifications can be explained by the spectral similarity of Vachellia and the soil classes during the dry season in which the datasets were acquired. During the dry period, being a deciduous tree species, Vachellia is shedding its leaves, thus resembling soil.
- An additional factor potentially explaining the differences between the class Other and vegetated classes (or soils and sparse vegetation) in 2016, can be attributed to the heavy rainfall coinciding with the 2016 El Niño–Southern Oscillation (ENSO). ENSO is known facilitating fast proliferation of Prosopis and Vachellia [105,106].
- Observed differences in the Water class are mainly caused by the slowly retreating water level, a consequence of the sudden increase of the water level of Lake Baringo in 2010 [107].
4.3. Validation and Reference Data
4.4. Random Forest
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pr > 50% | Pr < 50% | Mix Pr Dominant | Mix Va Dominant | Va > 50% | Va < 50% | Agriculture | Other Vegetation | Light Soils | Dark Soils | Water | Build-up | Ui | |
| Pr > 50% | 22 | 1 | 3 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0.79 |
| Pr < 50% | 2 | 26 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0.84 |
| Mix Pr dominant | 5 | 0 | 22 | 5 | 0 | 0 | 2 | 1 | 0 | 0 | 0 | 0 | 0.63 |
| Mix Va dominant | 0 | 0 | 5 | 18 | 3 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0.67 |
| Va > 50% | 0 | 0 | 0 | 5 | 23 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.82 |
| Va < 50% | 0 | 0 | 0 | 0 | 4 | 29 | 0 | 0 | 0 | 2 | 0 | 0 | 0.83 |
| Agriculture | 1 | 1 | 0 | 1 | 0 | 0 | 23 | 3 | 0 | 0 | 0 | 0 | 0.79 |
| Other vegetation | 0 | 2 | 0 | 0 | 0 | 0 | 2 | 23 | 0 | 0 | 0 | 0 | 0.85 |
| Light soils | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 29 | 0 | 1 | 1 | 0.91 |
| Dark soils | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 27 | 0 | 0 | 0.93 |
| Water | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 29 | 0 | 0.97 |
| Build-up | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 29 | 1.00 |
| Pi | 0.73 | 0.87 | 0.73 | 0.60 | 0.77 | 0.97 | 0.77 | 0.77 | 0.97 | 0.90 | 0.97 | 0.97 | 0.83 |
| Pr > 50% | Pr < 50% | Mix Pr Dominant | Mix Va Dominant | Va > 50% | Va < 50% | Agriculture | Other Vegetation | Light Soils | Dark Soils | Water | Build-up | Ui | |
| Pr > 50% | 22 | 0 | 0 | 0 | 0 | 0 | 7 | 1 | 0 | 0 | 0 | 0 | 0.73 |
| Pr < 50% | 1 | 21 | 2 | 0 | 0 | 2 | 4 | 1 | 0 | 1 | 1 | 0 | 0.64 |
| Mix Pr dominant | 2 | 0 | 22 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.79 |
| Mix Va dominant | 3 | 0 | 2 | 20 | 5 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0.65 |
| Va > 50% | 0 | 0 | 1 | 6 | 24 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.77 |
| Va < 50% | 0 | 5 | 1 | 1 | 1 | 27 | 1 | 0 | 0 | 2 | 0 | 0 | 0.71 |
| Agriculture | 2 | 1 | 2 | 0 | 0 | 0 | 17 | 0 | 0 | 1 | 0 | 0 | 0.74 |
| Other vegetation | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 26 | 0 | 0 | 0 | 0 | 0.90 |
| Light soils | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 24 | 0 | 0 | 3 | 0.86 |
| Dark soils | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 25 | 0 | 0 | 0.96 |
| Water | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 29 | 0 | 0.97 |
| Build-up | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 6 | 0 | 0 | 27 | 0.79 |
| Pi | 0.73 | 0.70 | 0.73 | 0.65 | 0.80 | 0.90 | 0.57 | 0.87 | 0.80 | 0.83 | 0.97 | 0.90 | 0.79 |
| Pléiades | Sentinel-2 | ||||||||||
| Prosopis | Mix | Vachellia | Other | Ui | Prosopis | Mix | Vachellia | Other | Ui | ||
| OOB accuracy | Prosopis | 51 | 3 | 1 | 3 | 0.879 | 44 | 0 | 2 | 15 | 0.721 |
| Mix | 5 | 50 | 3 | 4 | 0.806 | 5 | 48 | 5 | 1 | 0.814 | |
| Vachellia | 0 | 5 | 56 | 2 | 0.889 | 5 | 9 | 52 | 3 | 0.754 | |
| Other | 4 | 1 | 0 | 171 | 0.972 | 6 | 2 | 1 | 161 | 0.947 | |
| Pi | 0.850 | 0.847 | 0.933 | 0.950 | 0.914 | 0.733 | 0.814 | 0.867 | 0.894 | 0.850 | |
| Pléiades | Sentinel-2 | ||||||||||
| Prosopis | Mix | Vachellia | Other | Ui | Prosopis | Mix | Vachellia | Other | Ui | ||
| Sample count | Prosopis | 49 | 7 | 1 | 3 | 0.817 | 44 | 3 | 2 | 14 | 0.698 |
| Mix | 1 | 33 | 0 | 0 | 0.971 | 9 | 40 | 5 | 1 | 0.727 | |
| Vachellia | 0 | 5 | 63 | 2 | 0.900 | 2 | 1 | 57 | 9 | 0.826 | |
| Other | 7 | 0 | 2 | 68 | 0.883 | 2 | 1 | 2 | 49 | 0.907 | |
| Pi | 0.860 | 0.733 | 0.955 | 0.932 | 0.884 | 0.772 | 0.889 | 0.864 | 0.671 | 0.788 | |
| Pléiades | Sentinel-2 | ||||||||||
| Prosopis | Mix | Vachellia | Other | Ui | Prosopis | Mix | Vachellia | Other | Ui | ||
| Area sum | Prosopis | 105,576 | 35,280 | 368 | 22,136 | 0.646 | 201,400 | 3400 | 6100 | 37,100 | 0.812 |
| Mix | 384 | 125,584 | 0 | 0 | 0.997 | 53,800 | 595,300 | 16,300 | 83,600 | 0.795 | |
| Vachellia | 0 | 63,408 | 31,731,786 | 17,252 | 0.997 | 11,200 | 600 | 4,7547,100 | 127,900 | 0.997 | |
| Other | 84,568 | 0 | 388,160 | 18,254,284 | 0.975 | 14,900 | 600 | 5200 | 13,728,300 | 0.998 | |
| Pi | 0.554 | 0.560 | 0.988 | 0.998 | 0.988 | 0.716 | 0.992 | 0.999 | 0.982 | 0.994 | |
| Pléiades | Sentinel-2 | ||||||||||
| Prosopis | Mix | Vachellia | Other | Ui | Prosopis | Mix | Vachellia | Other | Ui | ||
| Area proportionat | Prosopis | 0.071 | 0.024 | 0.000 | 0.015 | 0.646 | 0.095 | 0.002 | 0.003 | 0.017 | 0.812 |
| Mix | 0.000 | 0.035 | 0.000 | 0.000 | 0.997 | 0.004 | 0.047 | 0.001 | 0.007 | 0.795 | |
| Vachellia | 0.000 | 0.001 | 0.472 | 0.000 | 0.997 | 0.000 | 0.000 | 0.627 | 0.002 | 0.997 | |
| Other | 0.002 | 0.000 | 0.008 | 0.373 | 0.975 | 0.000 | 0.000 | 0.000 | 0.196 | 0.998 | |
| Pi | 0.975 | 0.583 | 0.983 | 0.961 | 0.950 | 0.954 | 0.967 | 0.993 | 0.884 | 0.964 | |
References
- Kariuki, P. A Social Forestry Project in Baringo, Kenya: A Critical Analysis; University of Queensland: Brisbane, Autralia, 1993. [Google Scholar]
- Shackleton, R.T.; Le Maitre, D.C.; Pasiecznik, N.M.; Richardson, D.M. Prosopis: A global assessment of the biogeography, benefits, impacts and management of one of the world’s worst woody invasive plant taxa. AoB Plants 2014, 6, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Shiferaw, H.; Teketay, D.; Nemomissa, S.; Assefa, F. Some biological characteristics that foster the invasion of Prosopis juliflora (Sw.) DC. at Middle Awash Rift Valley Area, north-eastern Ethiopia. J. Arid Environ. 2004, 58, 135–154. [Google Scholar] [CrossRef]
- Felker, P. Mesquite: An all purpose leguminous arid land tree. In New Agricultural Crops, American Association for the Advancement of Science Symposium; Ritchie, G.A., Ed.; Westview Press: Boulder, UT, USA, 1979; Volume 38, pp. 89–132. [Google Scholar]
- Forsyth, G.G.; Le Maitre, D.C.; O’Farrell, P.J.; van Wilgen, B.W. The prioritisation of invasive alien plant control projects using a multi-criteria decision model informed by stakeholder input and spatial data. J. Environ. Manag. 2012, 103, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Nilsen, E.T.; Sharifi, M.R.; Rundel, P.W.; Jarrell, W.M.; Virginia, R.A. Diurnal and seasonal water relations of the desert phreatophyte Prosopis glandulosa (honey mesquite) in the Sonoran Desert of California. Ecology 1983, 64, 1381–1393. [Google Scholar] [CrossRef]
- Dzikiti, S.; Schachtschneider, K.; Naiken, V.; Gush, M.; Moses, G.; Le Maitre, D.C. Water relations and the effects of clearing invasive Prosopis trees on groundwater in an arid environment in the northern Cape, South Africa. J. Arid Environ. 2013, 90, 103–113. [Google Scholar] [CrossRef]
- Elfadl, M.A.; Luukkanen, O. Ecological strategies of Prosopis juliflora in the arid environment of the Sudan: 1. Leaf gas exchange approach. J. Arid Environ. 2006, 66, 1–15. [Google Scholar] [CrossRef]
- Lenachuru, C. Impacts of Prosopis species in Baringo District. In Proceedings of workshop on Integrated Management of Prosopis Species in Kenya, Nairobi, Kenya, 1–2 October 2003. [CrossRef]
- Andersson, S. Spread of the Introduced Tree Species Prosopis juliflora (Sw.) DC in the Lake Baringo Area, Kenya. Ph.D. Thesis, Faculty of Forest Sciences, Swedish University of Agricultural Sciences, Umeå, Sweden, 2005. [Google Scholar]
- Pasiecznik, N.; Felker, P.; Cadoret, K.; Harsh, L.N.; Cruz, G.; Tewari, J.C.; Maldonado, L.J. The Prosopis juliflora-Prosopis pallida complex: The Prosopis juliflora-Prosopis pallida complex. Managing 2001, 231, 162. [Google Scholar]
- Lowe, S.; Browne, M.; Boudjelas, S.; De Poorter, M. 100 of the World’s Worst Invasive Alien Species: A Selection from the Global Invasive Species Database; Invasive Species Specialist Group a Specialist Group Species Survival Commission of the World Conservation Union: Auckland, New Zealand, 2000. [Google Scholar]
- Maslin, B.R. Generic and subgeneric names in Acacia following retypification of the genus. Muelleria 2008, 26, 7–9. [Google Scholar]
- Groot, H.; Hall, D. How to pick crops that cope with extreme climates. In New Scientist; Reed Business Information Ltd.: London, UK, 1989; pp. 46–47. [Google Scholar]
- Burke, M.J.W.; Grime, J.P. An experimental study of plant community invasibility. Ecology 1996, 77, 776–790. [Google Scholar] [CrossRef]
- Yoda, K.; Elbasit, M.A.; Hoshino, B.; Nawata, H.; Yasuda, H. Root system development of Prosopis seedlings under different soil moisture conditions. J. Arid Land Stud. 2012, 16, 13–16. [Google Scholar]
- Aboud, A.A.; Kisoyan, P.K.; Coppock, D.L. Agro-pastoralists’ wrath for the Prosopis tree: The Case of the Il Chamus of Baringo district, Kenya. In Global Livestock Collaborative Research Support Program; ENVS Faculty Publications: Davis, CA, USA, 2005; pp. 1–3. [Google Scholar]
- Geesing, D.; Al-Khawlani, M.; Abba, M.L. Management of introduced Prosopis species: Can economic exploitation control an invasive species? Unasylva 2004, 55, 36–44. [Google Scholar]
- Sawal, R.K.; Ram, R.; Yadav, S.B.S. Mesquite (Prosopis juliflora) pods as a feed resource for livestock—A review. Asian-Aust. J. Anim. Sci. 2004, 17, 719–725. [Google Scholar] [CrossRef]
- Peter, F.F.; Thames, J.L. Collection, Handling, Storage and Pre-treatment of Prosopis Seeds in Latin America; FAO: Rome, Italy, 1983. [Google Scholar]
- Orwa, C.; Mutua, A.; Kindt, R.; Jamnadass, R.; Anthony, S. Agroforestree Database: A Tree Reference and Selection Guide Version 4.0. Available online: http://www.worldagroforestry.org/output/agroforestree-database (accessed on 6 January 2017).
- Guevara, A.; Giordano, C.V.; Aranibar, J.; Quiroga, M.; Villagra, P.E. Phenotypic plasticity of the coarse root system of Prosopis flexuosa, a phreatophyte tree, in the Monte Desert (Argentina). Plant Soil 2010, 330, 447–464. [Google Scholar] [CrossRef]
- Seghieri, J. The rooting patterns of woody and herbaceous plants in a savanna; Are they complementary or in competition? Afr. J. Ecol. 1995, 33, 358–365. [Google Scholar] [CrossRef]
- Aplin, P. Remote sensing: Ecology. Prog. Phys. Geogr. 2005, 29, 104–113. [Google Scholar] [CrossRef]
- Huang, C.Y.; Asner, G.P. Applications of remote sensing to alien invasive plant studies. Sensors 2009, 9, 4869–4889. [Google Scholar] [CrossRef] [PubMed]
- Rembold, F.; Leonardi, U.; Ng, W.; Gadain, H.; Meroni, M. Mapping areas invaded by Prosopis juliflora in Somaliland with Landsat 8 imagery. Proc. SPIE 2015, 9637, 1–12. [Google Scholar]
- Meroni, M.; Ng, W.-T.; Rembold, F.; Leonardi, U.; Atzberger, C.; Gadain, H.; Shaiye, M. Mapping Prosopis juliflora in west Somaliland with Landsat 8 satellite imagery and ground information. Land Degrad. Dev. 2016. [Google Scholar] [CrossRef]
- Ng, W.-T.; Meroni, M.; Immitzer, M.; Böck, S.; Leonardi, U.; Rembold, F.; Gadain, H.; Atzberger, C. Mapping Prosopis spp. with Landsat 8 data in arid environments: Evaluating effectiveness of different methods and temporal imagery selection for Hargeisa, Somaliland. Int. J. Appl. Earth Obs. Geoinf. 2016, 53, 76–89. [Google Scholar] [CrossRef]
- Wakie, T.T.; Evangelista, P.H.; Jarnevich, C.S.; Laituri, M. Mapping current and potential distribution of non-native Prosopis juliflora in the Afar Region of Ethiopia. PLoS ONE 2014, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ayanu, Y.; Jentsch, A.; Müller-Mahn, D.; Rettberg, S.; Romankiewicz, C.; Koellner, T. Ecosystem engineer unleashed: Prosopis juliflora threatening ecosystem services? Reg. Environ. Chang. 2014, 15, 155–167. [Google Scholar] [CrossRef]
- Van Den Berg, E.C.; Kotze, I.; Beukes, H. Detection, quantification and monitoring Prosopis spp. in the Northern Cape Province of South Africa using remote sensing and GIS. South Afr. J. Geomatic. 2013, 2, 68–81. [Google Scholar]
- Robinson, T.P.; Wardell-Johnson, G.W.; Pracilio, G.; Brown, C.; Corner, R.; van Klinken, R.D. Testing the discrimination and detection limits of WorldView-2 imagery on a challenging invasive plant target. Int. J. Appl. Earth Obs. Geoinf. 2016, 44, 23–30. [Google Scholar] [CrossRef]
- Muturi, G.M.; Poorter, L.; Mohren, G.M.J.; Kigomo, B.N. Ecological impact of Prosopis species invasion in Turkwel riverine forest, Kenya. J. Arid Environ. 2013, 92, 89–97. [Google Scholar] [CrossRef]
- Dubow, A.Z. Mapping and Managing the Spread of Prosopis Juliflora in Garissa County, Kenya. Master’s Thesis, School of Environmental Studies of Kenyatta University, Nairobi, Kenya, 2011; p. 80. [Google Scholar]
- Amboka, A.A.; Ngigi, T.G. Mapping and monitoring spatial-temporal cover change of Prosopis species colonization in Baringo Central, Kenya. Int. J. Eng. Sci. Invent. 2015, 4, 2319–6734. [Google Scholar]
- Swallow, B.; Mwangi, E. Invasion and rural livelihoods in the Lake Baringo area of Kenya. Conserv. Soc. 2008, 6, 130. [Google Scholar] [CrossRef]
- Thenkabail, P.S. Land Resources Monitoring, Modeling, and Mapping with Remote Sensing; CRC Press: Boca Raton, FL, USA, 2015. [Google Scholar]
- Alsharrah, S.A.; Bruce, D.A.; Bouabid, R.; Somenahalli, S.; Corcoran, P.A. High-spatial resolution multispectral and panchromatic satellite imagery for mapping perennial desert plants. Proc. SPIE 2015, 9644. [Google Scholar] [CrossRef]
- Regniers, O.; Bombrun, L.; Guyon, D.; Samalens, J.C.; Germain, C. Wavelet-based texture features for the classification of age classes in a maritime pine forest. IEEE Geosci. Remote Sens. Lett. 2015, 12, 621–625. [Google Scholar] [CrossRef]
- Immitzer, M.; Vuolo, F.; Atzberger, C. First experience with Sentinel-2 data for crop and tree species classifications in Central Europe. Remote Sens. 2016, 8, 166. [Google Scholar] [CrossRef]
- Radoux, J.; Chomé, G.; Jacques, D.C.; Waldner, F.; Bellemans, N.; Matton, N.; Lamarche, C.; d’Andrimont, R.; Defourny, P. Sentinel-2’s potential for sub-pixel landscape feature detection. Remote Sens. 2016, 8, 488. [Google Scholar] [CrossRef]
- Pratt, D.; Gwynne, M. Rangeland Management and Ecology in East Africa; Hodder & Stoughton: London, UK, 1977. [Google Scholar]
- Thom, D.; Martin, N. Ecology and production in Baringo-Kerio Valley, Kenya. Geogr. Rev. 1983, 73, 15–29. [Google Scholar] [CrossRef]
- Meroni, M.; Verstraete, M.M.; Rembold, F.; Urbano, F.; Kayitakire, F. A phenology-based method to derive biomass production anomalies for food security monitoring in the Horn of Africa. Int. J. Remote Sens. 2014, 35, 2472–2492. [Google Scholar] [CrossRef]
- Wiesmann, U.; Kiteme, B.; Mwangi, Z. Socio-Economic Atlas of Kenya: Depicting the National Population Census by County and Sub-Location, 2nd ed.; KNBS: Nairobi, Kenya; CETRAD: Nanyuki, Kenya; CDE: Bern, Switherland, 2016. [Google Scholar]
- Geosystems. Erdas imagine. Imagine 2005, 15, 1–16. [Google Scholar]
- Main-Knorn, M.; Pflug, B.; Debaecker, V.; Louis, J. Calibration and validation plan for the L2a processor and products of the Sentinel-2 mission. ISPRS Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. 2015, 40, 1249–1255. [Google Scholar] [CrossRef]
- Vuolo, F.; Żółtak, M.; Pipitone, C.; Zappa, L.; Wenng, H.; Immitzer, M.; Weiss, M.; Baret, F.; Atzberger, C. Data service platform for Sentinel-2 surface reflectance and value-added products: System use and examples. Remote Sens. 2016, 8, 938. [Google Scholar] [CrossRef]
- Inglada, J.; Christophe, E. The Orfeo Toolbox remote sensing image processing software. In Proceedings of 2009 IEEE International Geoscience and Remote Sensing Symposium, Cape Town, South Africa, 12–17 July 2009; pp. 733–736.
- Harrington, M.; Cross, M. Google Earth Forensics; Elsevier: Waltham, MA, USA, 2015. [Google Scholar]
- Woody Weeds Project. Available online: http://woodyweeds.org/ (accessed on 4 January 2017).
- Blaschke, T. Object based image analysis for remote sensing. ISPRS J. Photogramm. Remote Sens. 2010, 65, 2–16. [Google Scholar] [CrossRef]
- Blaschke, T.; Hay, G.J.; Kelly, M.; Lang, S.; Hofmann, P.; Addink, E.; Feitosa, R.Q.; van der Meer, F.; van der Werff, H.; van Coillie, F.; et al. Geographic object-based image analysis—Towards a new paradigm. ISPRS J. Photogramm. Remote Sens. 2014, 87, 180–191. [Google Scholar] [CrossRef] [PubMed]
- Fukunaga, K.; Hostetler, L. The estimation of the gradient of a density function, with applications in pattern recognition. IEEE Trans. Inf. Theory 1975, 21, 32–40. [Google Scholar] [CrossRef]
- Comaniciu, D.; Meer, P. Mean shift: A robust approach toward feature space analysis. IEEE Trans. Pattern Anal. Mach. 2002, 24, 603–619. [Google Scholar] [CrossRef]
- Michel, J.; Youssefi, D.; Grizonnet, M. Stable mean-shift algorithm and its application to the segmentation of arbitrarily large remote sensing images. IEEE Trans. Geosci. Remote Sens. 2015, 53, 952–964. [Google Scholar] [CrossRef]
- Huang, X.H.X.; Zhang, L.Z.L. An adaptive mean-shift analysis approach for object extraction and classification from urban hyperspectral imagery. IEEE Trans. Geosci. Remote Sens. 2008, 46, 4173–4185. [Google Scholar] [CrossRef]
- Pouliot, D.A.; King, D.J.; Bell, F.W.; Pitt, D.G. Automated tree crown detection and delineation in high-resolution digital camera imagery of coniferous forest regeneration. Remote Sens. Environ. 2002, 82, 322–334. [Google Scholar] [CrossRef]
- Waser, L.T.; Küchler, M.; Jütte, K.; Stampfer, T. Evaluating the potential of WorldView-2 data to classify tree species and different levels of ash mortality. Remote Sens. 2014, 6, 4515–4545. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Kaufman, Y.J.; Merzlyak, M.N. Use of a green channel in remote sensing of global vegetation from EOS-MODIS. Remote Sens. Environ. 1996, 58, 289–298. [Google Scholar] [CrossRef]
- Immitzer, M.; Atzberger, C. Early detection of Bark Beetle Infestation in Norway Spruce (Picea abies, L.) using WorldView-2 Data. Photogramm. Fernerkund. Geoinf. 2014, 5, 351–367. [Google Scholar] [CrossRef]
- Qi, J.; Chehbouni, A.; Huete, A.R.; Kerr, Y.H.; Sorooshian, S. A modified soil adjusted vegetation index. Remote Sens. Environ. 1994, 48, 119–126. [Google Scholar] [CrossRef]
- Gitelson, A.; Merzlyak, M.N. Quantitative estimation of chlorophyll-a using reflectance spectra: Experiments with autumn chestnut and maple leaves. J. Photochem. Photobiol. B Biol. 1994, 22, 247–252. [Google Scholar] [CrossRef]
- Barnes, E.; Clarke, T.; Richards, S.; Colaizzi, P.D.; Haberland, J.; Kostrzewski, M.; Waller, P.; Choi, C.; Riley, E.; Thompson, T.; et al. Coincident detection of crop water stress, nitrogen status and canopy density using ground-based multispectral data. In Proceedings of the 5th International Conference on Precision Agriculture, Bloomington, MN, USA, 16–19 July 2000.
- Rouse, J.W.; Haas, R.H.; Schell, J.A.; Deering, D.W. Monitoring vegetation systems in the Great Plains with ERTS. In Proceedings of Third Earth Resources Technology Satellite-1 Symposium, Washington, DC, USA, 10–14 December 1973; pp. 309–317.
- Gao, B.C. NDWI—A normalized difference water index for remote sensing of vegetation liquid water from space. Remote Sens. Environ. 1996, 58, 257–266. [Google Scholar] [CrossRef]
- Sripada, R.P.; Heiniger, R.W.; White, J.G.; Meijer, A.D. Aerial color infrared photography for determining early in-season nitrogen requirements in corn. Agron. J. 2006, 98, 968–977. [Google Scholar] [CrossRef]
- Merzlyak, M.N.; Gitelson, A.A.; Chivkunova, O.B.; Rakitin, V.Y.U. Non-destructive optical detection of pigment changes during leaf senescence and fruit ripening. Physiol. Plant. 1999, 106, 135–141. [Google Scholar] [CrossRef]
- Jordan, C.F. Derivation of leaf-area index from quality of light on the forest floor. Ecology 1969, 50, 663–666. [Google Scholar] [CrossRef]
- Pearson, R.L.; Miller, L.D. Remote mapping of standing crop biomass for estimation of the productivity of the short-grass Prairie, Pawnee National Grassland, Colorado. In Proceedings of Eighth International Symposium on Remote Sensing of Environment, Ann Arbor, MI, USA, 2–6 October 1972; pp. 1357–1381.
- Walz, U.; Hou, W. Charakterisierung der Landschaftsvielfalt mit RapidEye-Daten—Erste Ergebnisse und Erfahrungen. In Proceedings of the 3 RESA Workshop, Neustrelitz, Germany, 23 March 2011; pp. 75–85.
- Chávez, R.; Clevers, J.G.P.W. Phase 2. In Object-Based Analysis of 8-Bands WorldView-2 Imagery for Assessing Health Condition of Desert Trees, 8-Band Research Challenge; Digital Globe of Longmont: Westminster, CO, USA, 2011; pp. 1–8. [Google Scholar]
- Richardson, A.J.; Wiegand, C.L. Distinguishing vegetation from soil background information. Photogramm. Eng. Remote Sens. 1977, 43, 1541–1552. [Google Scholar]
- Clevers, J.G.P.W. The derivation of a simplified reflectance model for the estimation of leaf area index. Remote Sens. Environ. 1988, 25, 53–69. [Google Scholar] [CrossRef]
- Wolf, A.F. Using WorldView-2 Vis-NIR multispectral imagery to support land mapping and feature extraction using normalized difference index ratios. Proc. SPIE 2012, 8390. [Google Scholar] [CrossRef]
- Toscani, P.; Immitzer, M.; Atzberger, C. Wavelet-based texture measures for object-based classification of aerial images. Photogramm. Fernerkund. Geoinf. 2013, 2013, 105–121. [Google Scholar] [CrossRef]
- Huang, X.; Zhang, L. An SVM ensemble approach combining spectral, structural, and semantic features for the classification of high-resolution remotely sensed imagery. IEEE Trans. Geosci. Remote Sens. 2013, 51, 257–272. [Google Scholar] [CrossRef]
- Immitzer, M.; Atzberger, C.; Koukal, T. Tree species classification with random forest using very high spatial resolution 8-Band WorldView-2 satellite data. Remote Sens. 2012, 4, 2661–2693. [Google Scholar] [CrossRef]
- Huang, X.; Zhang, L.; Wang, L. Evaluation of morphological texture features for mangrove forest mapping and species discrimination using multispectral IKONOS imagery. IEEE Geosci. Remote Sens. Lett. 2009, 6, 393–397. [Google Scholar] [CrossRef]
- Daubechies, I. Ten Lectures on Wavelets; SIAM: Philadelphia, PA, USA, 1992; Volume 61. [Google Scholar]
- MATLAB R2012a, version 7.10.0; The MathWorks Inc.: Natick, MA, USA, 2010.
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Hastie, T.; Tibshirani, R.; Friedman, J. The Elements of Statistical Learning: Data Mining, Inference, and Prediction; Springer: Berlin, Germany, 2009. [Google Scholar]
- Gislason, P.O.; Benediktsson, J.A.; Sveinsson, J.R. Random forests for land cover classification. Pattern Recognit. Lett. 2006, 27, 294–300. [Google Scholar] [CrossRef]
- Li, X.; Cheng, X.; Chen, W.; Chen, G.; Liu, S. Identification of forested landslides using LiDar data, object-based image analysis, and machine learning algorithms. Remote Sens. 2015, 7, 9705–9726. [Google Scholar] [CrossRef]
- Li, X.; Chen, W.; Cheng, X.; Wang, L. A comparison of machine learning algorithms for mapping of complex surface-mined and agricultural landscapes using ZiYuan-3 stereo satellite imagery. Remote Sens. 2016, 8, 514. [Google Scholar] [CrossRef]
- Liaw, A.; Wiener, M. Classification and regression by randomForest. R News 2002, 2, 18–22. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Genuer, R.; Poggi, J.-M.; Tuleau-Malot, C. Variable selection using random forests. Pattern Recognit. Lett. 2010, 31, 2225–2236. [Google Scholar] [CrossRef]
- Congalton, R.G.; Green, K. Assessing the Accuracy of Remotely Sensed Data: Principles and Practices; CRC Press: Boca Raton, FL, USA, 2009; Volume 2. [Google Scholar]
- Olofsson, P.; Foody, G.M.; Herold, M.; Stehman, S.V.; Woodcock, C.E.; Wulder, M.A. Good practices for estimating area and assessing accuracy of land change. Remote Sens. Environ. 2014, 148, 42–57. [Google Scholar] [CrossRef]
- Stehman, S.V. Estimating area from an accuracy assessment error matrix. Remote Sens. Environ. 2013, 132, 202–211. [Google Scholar] [CrossRef]
- Radoux, J.; Bogaert, P. Accounting for the area of polygon sampling units for the prediction of primary accuracy assessment indices. Remote Sens. Environ. 2014, 142, 9–19. [Google Scholar] [CrossRef]
- Rembold, F.; Atzberger, C.; Savin, I.; Rojas, O. Using low resolution satellite imagery for yield prediction and yield anomaly detection. Remote Sens. 2013, 5, 1704–1733. [Google Scholar] [CrossRef]
- Vuolo, F.; Atzberger, C. Improving land cover maps in areas of disagreement of existing products using NDVI time series of MODIS—Example for Europe. Photogramm. Fernerkund. Geoinf. 2014, 201, 393–407. [Google Scholar] [CrossRef]
- Schultz, B.; Immitzer, M.; Formaggio, A.R.; Sanches, I.D.A.; Luiz, A.J.B.; Atzberger, C. Self-guided segmentation and classification of multi-temporal Landsat 8 images for crop type mapping in Southeastern Brazil. Remote Sens. 2015, 7, 14482–14508. [Google Scholar] [CrossRef]
- Immitzer, M.; Vuolo, F.; Einzmann, K.; Ng, W.-T.; Böck, S.; Atzberger, C. Verwendung von multispektralen Sentinel-2 Daten für die Baumartenklassifikation und Vergleich mit anderen Satellitensensoren. In Proceedings of Conferemce Dreiländertagung der SGPF, DGPF und OVG, Bern, Switzerland, 8–9 June 2016; Volume 25, pp. 417–427.
- Mirik, M.; Ansley, R.J. Utility of satellite and aerial images for quantification of canopy cover and infilling rates of the invasive woody species honey mesquite (Prosopis Glandulosa) on Rangeland. Remote Sens. 2012, 4, 1947–1962. [Google Scholar] [CrossRef]
- Stehman, S.V.; Wickham, J.D. Pixels, blocks of pixels, and polygons: Choosing a spatial unit for thematic accuracy assessment. Remote Sens. Environ. 2011, 115, 3044–3055. [Google Scholar] [CrossRef]
- Radoux, J.; Bogaert, P.; Fasbender, D.; Defourny, P. Thematic accuracy assessment of geographic object-based image classification. Int. J. Geogr. Inf. Sci. 2011, 25, 895–911. [Google Scholar] [CrossRef]
- Duro, D.C.; Franklin, S.E.; Dubé, M.G. A comparison of pixel-based and object-based image analysis with selected machine learning algorithms for the classification of agricultural landscapes using SPOT-5 HRG imagery. Remote Sens. Environ. 2012, 118, 259–272. [Google Scholar] [CrossRef]
- Fassnacht, F.E.; Latifi, H.; Stereńczak, K.; Modzelewska, A.; Lefsky, M.; Waser, L.T.; Staub, C.; Ghosh, A. Review of studies on tree species classification from remotely sensed data. Remote Sens. Environ. 2016, 186, 64–87. [Google Scholar] [CrossRef]
- US Embassy Cummins Cogeneration Kenya Limited (CCKL) 12MW Biomass-Fueled, On-Grid Electricity Generation Project in Baringo County, Kenya. Available online: http://nairobi.usembassy.gov/cumminsbaringo.html (accessed on 5 January 2017).
- Cummins Power Generation. Cummins Power Generation Project in Kenya Paves the Way for African Biogas Expansion. Available online: http://www.cumminspowerblog.com/en/2014/08/27/cummins-power-generation-project-kenya-paves-way-african-biogas-expansion/ (accessed on 5 January 2017).
- Red Cross Kenya. Situation Report of Effects of the El Nino Rains; Red Cross Kenya: Nairobi, Kenya, 2015. [Google Scholar]
- Rui, H.; McNally, A. Famine Early Warning Systems Network (FEWS NET) Land Data Assimilation System (FLDAS) Product; USGS Center for Earth Resources Observation and Science (EROS): Sioux Falls, SD, USA, 2015.
- Obando, J.; Onywere, S.; Shisanya, C.; Nbudi, A.; Masiga, D.; Irura, Z.; Mariita, N.; Maragia, H. Impact of short-term flooding on livelihoods in the Kenya Rift Valley Lakes. In Advances in Geographical and Environmental Sciences; Springer: Tokyo, Japan, 2016; pp. 193–215. [Google Scholar]
- Schmidt, T.; Schuster, C.; Kleinschmit, B.; Förster, M. Evaluating an intra-annual time series for grassland classification—How many acquisitions and what seasonal origin are optimal? IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2014, 7, 3428–3439. [Google Scholar] [CrossRef]
- Vuolo, F.; Richter, K.; Atzberger, C. Evaluation of time-series and phenological indicators for land cover classification based on MODIS data. Proc. SPIE 2011. [Google Scholar] [CrossRef]
- Elfadl, M.A. Growth performance and physiological characteristics of seedlings of six tropical dry. J. Nat. Resour. Environ. Stud. 2013, 1, 25–33. [Google Scholar]
- Muthana, K.D.; Arora, G.D. Prosopis juliflora (Swartz) DC, a fast growing tree to bloom the desert. In Central Arid Zone Research Institute Monograph; Central Arid Zone Research Institute: Jodhpur, India, 1983; Volume 22, pp. 1–19. [Google Scholar]
- Koger, C.H.; Bruce, L.M.; Shaw, D.R.; Reddy, K.N. Wavelet analysis of hyperspectral reflectance data for detecting pitted morningglory (Ipomoea lacunosa) in soybean (Glycine max). Remote Sens. Environ. 2003, 86, 108–119. [Google Scholar] [CrossRef]
- Peerbhay, K.Y.; Mutanga, O.; Ismail, R. Investigating the capability of few strategically placed WorldView-2 multispectral bands to discriminate forest species in KwaZulu-Natal, South Africa. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2014, 7, 307–316. [Google Scholar] [CrossRef]
- Pu, R.; Liu, D. Segmented canonical discriminant analysis of in situ hyperspectral data for identifying 13 urban tree species. Int. J. Remote Sens. 2011, 32, 2207–2226. [Google Scholar] [CrossRef]


| Initial Location | No. of Sites | Extent (Ha) | Prosopis Species |
|---|---|---|---|
| Salabani | 1 | 25.7 | P. juliflora, P. pallida |
| Loruk | 2 | 60.2 | P. juliflora, P. pallida |
| Kapthurin River | 2 | 17 | P. juliflora, P. pallida |
| Nga’mbo | 3 | 14.6 | P. juliflora,P. pallida |
| Chemeron Dam | 1 | NA | P. juliflora,P. pallida |
| Marigat | 2 | 29.1 | P. juliflora, P. pallida |
| Eldume | 1 | 75.9 | P. juliflora, P. pallida |
| Logumgum | 1 | 9.9 | P. juliflora, P. pallida |
| Sandai | 1 | 4.3 | P. juliflora, P. pallida |
| Eldepe | 1 | 4.9 | P. chilensis |
| Sintaan | 1 | 1.6 | P. chilensis |
| Kapkuikui | 1 | NA | P. chilensis |
| Loboi | 1 | 13.3 | P. chilensis |
| Others (Small scale) | 4 | NA | P. chilensis, P. juliflora, P. pallida |
| Sensor | Spatial Resolution | Acquisition Date | Cloud Cover (%) | Bands |
|---|---|---|---|---|
| Pléiades | 2 m | 11 February 2015 | 0 | 4 MS + 1 PAN |
| Sentinel-2 | 10/20 m | 17 February 2016 | 0 | 10 MS |
| ID | Re-Class | Class Name | Class Description |
|---|---|---|---|
| 1 | 1 | Pr > 50% | Dense Prosopis covering > 50% |
| 2 | 1 | Pr < 50% | Sparse Prosopis covering < 50% |
| 3 | 2 | Mix Pr dominant | Dominant Prosopis (>50%) mixed with Vachellia |
| 4 | 2 | Mix Va dominant | Dominant Vachellia (>50%) mixed with Prosopis |
| 5 | 3 | Va > 50% | Dense Vachellia covering > 50% |
| 6 | 3 | Va < 50% | Sparse Vachellia covering < 50% |
| 7 | 4 | Agriculture | Cropped agricultural areas |
| 8 | 4 | Other vegetation | Aquatic, alkaline and other vegetation |
| 9 | 4 | Light soils | Light soils (sand) |
| 10 | 4 | Dark soils | Dark soils (clay or loam) |
| 11 | 4 | Water | Waterbodies, stream and lakes |
| 12 | 4 | Build-up | Urban areas and settlements |
| Sensor | Spatial Radius | Range Radius | Minimum Size | Segments (n) |
|---|---|---|---|---|
| Pléiades | 35 | 0.34 | 10 | 1,350,000 |
| Sentinel-2 | 30 | 0.2 | 4 | 157,312 |
| Index | Equation | Reference | Data |
|---|---|---|---|
| Blue Ratio (BR) | Waser et al. 2014 [59] | S2A | |
| Green Normalized Difference Vegetation Index (GNDVI) | Gitelson and Merzlyak 1996 [60] | S2A | |
| Green Ratio (GR) | Waser et al. 2014 [59], Immitzer and Atzberger [61] | S2A | |
| Modified Soil Adjusted Vegetation Index (MSAVI) | Qi et al. 1994 [62] | S2A, PLE | |
| Normalized Difference Red Edge Index (NDREI) | Gitelson and Merzlyak 1994 [63], Barnes et al. 2000 [64] | S2A | |
| Normalized Difference Vegetation Index (NDVI) | Rouse et al. 1974 [65] | S2A, PLE | |
| Normalized Difference Vegetation Index 2 (NDVI2) | Rouse et al. 1974 [65] | S2A | |
| Normalized Difference Water Index (NDWI) | Gao 1996 [66] | S2A, PLE | |
| Normalized Near Infrared (NNIR) | Sripada et al. 2006 [67] | S2A | |
| Plant Senescence Reflectance Index (PSRI) | Merzlyak et al. 1999 [68] | S2A | |
| Red Ratio (RR) | Waser et al. 2014 [59] | S2A | |
| Ratio Vegetation Index (RVI) | Jordan 1969 [69], Pearson and Miller 1972 [70] | S2A, PLE | |
| Sentinel Improved Vegetation Index (SVI) | evolved for this study | S2A | |
| Vegetation Index based on RedEdge (VIRE) | Walz and Hou 2011 [71] | S2A | |
| Vegetation Index Ratio based on RedEdge (VIRRE) | Chávez and Clevers 2011 [72] | S2A | |
| Weighted Difference Vegetation Index (WDVI) | Richardson and Wiegand 1977 [73], Clevers 1988 [74] | S2A, PLE | |
| WorldView Improved Vegetative Index (WVVI) | Wolf 2012 [75] | S2A |
| 12 Classes | Pléiades | Sentinel-2 |
|---|---|---|
| Pr > 50% | 11.40 | 12.23 |
| Pr < 50% | 35.55 | 37.26 |
| Mix Pr dominant | 5.85 | 15.03 |
| Mix Va dominant | 8.94 | 9.97 |
| Va > 50% | 74.07 | 26.91 |
| Va < 50% | 127.31 | 240.42 |
| Agriculture | 5.39 | 16.18 |
| Other vegetation | 11.65 | 3.83 |
| Light soils | 60.33 | 14.93 |
| Dark soils | 61.18 | 29.93 |
| Water | 19.42 | 17.12 |
| Build-up | 4.90 | 1.43 |
| 4 Classes | Pléiades | Sentinel-2 |
| Prosopis | 46.95 | 49.49 |
| Mix | 14.79 | 25.00 |
| Vachellia | 201.38 | 267.33 |
| Other | 162.87 | 83.42 |
| User’s Accuracy (Ui) | Producer’s Accuracy (Pi) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Class | Reference (n) | Map Prop (Wi) | Mean Size (ha) | OOB | Sample Count | Area Prop | OOB | Sample Count | Area Prop |
| Pleiàdes | |||||||||
| Prosopis | 57 | 0.110 | 0.272 | 0.879 | 0.817 | 0.646 | 0.850 | 0.860 | 0.975 |
| Mix | 45 | 0.035 | 0.370 | 0.806 | 0.971 | 0.997 | 0.847 | 0.733 | 0.583 |
| Vachellia | 66 | 0.473 | 45.446 | 0.889 | 0.900 | 0.997 | 0.933 | 0.955 | 0.983 |
| Other | 73 | 0.382 | 24.321 | 0.972 | 0.883 | 0.975 | 0.950 | 0.932 | 0.961 |
| Sentinel-2 | |||||||||
| Prosopis | 57 | 0.116 | 0.394 | 0.721 | 0.698 | 0.812 | 0.733 | 0.772 | 0.954 |
| Mix | 45 | 0.059 | 1.362 | 0.814 | 0.727 | 0.795 | 0.814 | 0.889 | 0.967 |
| Vachellia | 66 | 0.629 | 69.111 | 0.754 | 0.826 | 0.997 | 0.867 | 0.864 | 0.993 |
| Other | 73 | 0.196 | 25.461 | 0.947 | 0.907 | 0.998 | 0.894 | 0.671 | 0.884 |
© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ng, W.-T.; Rima, P.; Einzmann, K.; Immitzer, M.; Atzberger, C.; Eckert, S. Assessing the Potential of Sentinel-2 and Pléiades Data for the Detection of Prosopis and Vachellia spp. in Kenya. Remote Sens. 2017, 9, 74. https://doi.org/10.3390/rs9010074
Ng W-T, Rima P, Einzmann K, Immitzer M, Atzberger C, Eckert S. Assessing the Potential of Sentinel-2 and Pléiades Data for the Detection of Prosopis and Vachellia spp. in Kenya. Remote Sensing. 2017; 9(1):74. https://doi.org/10.3390/rs9010074
Chicago/Turabian StyleNg, Wai-Tim, Purity Rima, Kathrin Einzmann, Markus Immitzer, Clement Atzberger, and Sandra Eckert. 2017. "Assessing the Potential of Sentinel-2 and Pléiades Data for the Detection of Prosopis and Vachellia spp. in Kenya" Remote Sensing 9, no. 1: 74. https://doi.org/10.3390/rs9010074
APA StyleNg, W.-T., Rima, P., Einzmann, K., Immitzer, M., Atzberger, C., & Eckert, S. (2017). Assessing the Potential of Sentinel-2 and Pléiades Data for the Detection of Prosopis and Vachellia spp. in Kenya. Remote Sensing, 9(1), 74. https://doi.org/10.3390/rs9010074