
Remote Sensing of Forest Gap Dynamics in the Białowieża Forest: Comparison of Multitemporal Airborne Laser Scanning and High-Resolution Aerial Imagery Point Clouds

 , , ,
, , ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
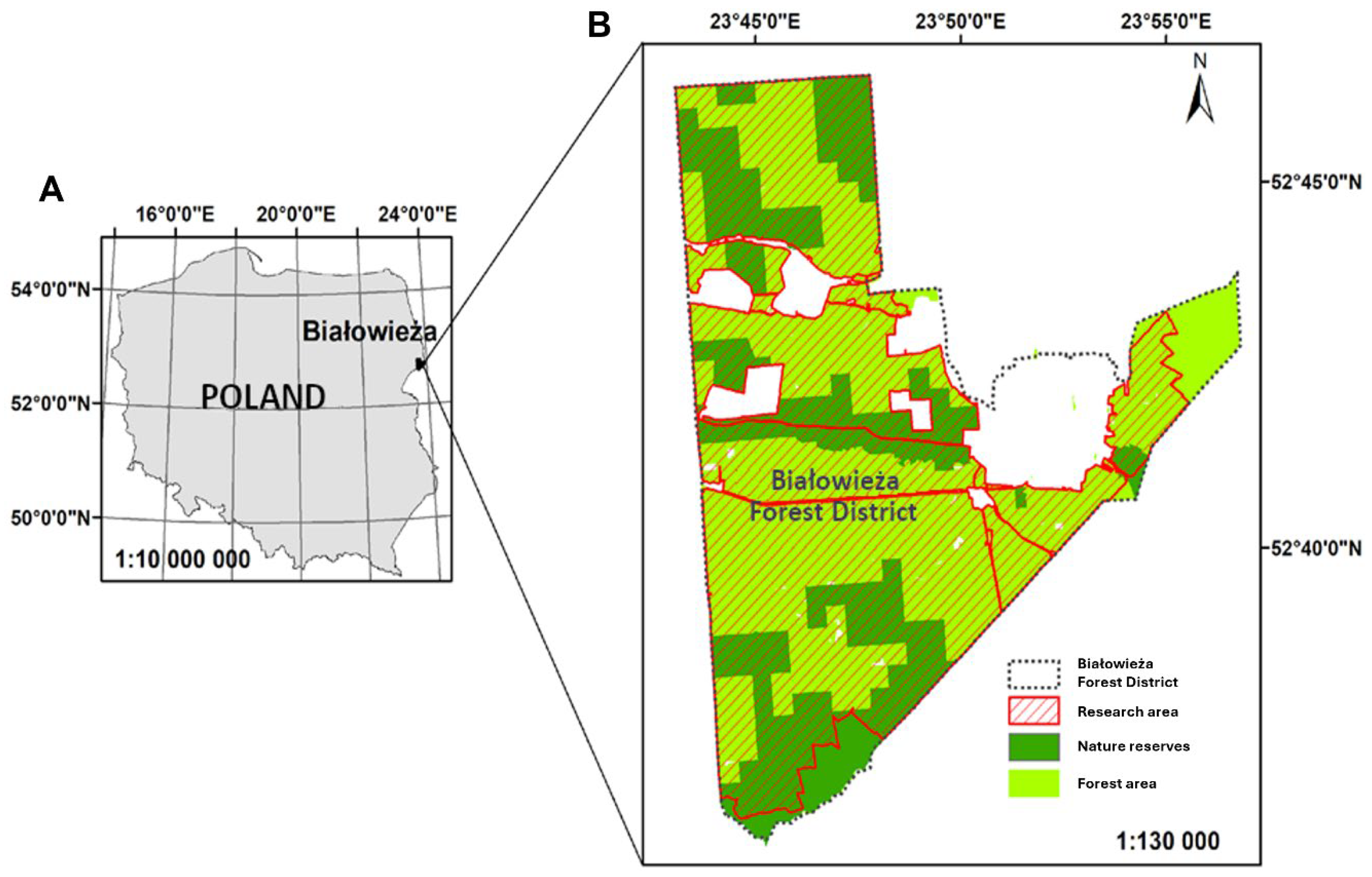
2.1. Study Area
2.2. Data
2.2.1. Aerial Imagery
2.2.2. Airborne Laser Scanning Data (ALS)
2.3. Processing of High-Resolution Aerial Imagery
- (i)
- The initial alignment of images was achieved through automatic aerotriangulation using a field photogrammetric warp measured in the field with a GNSS receiver. Despite automatic aerotriangulation, additional manual alignment of images was performed by application of extra photopoints and Ground Control Points (GCPs) to enhance the accuracy of the image block. Additional binding points, measured from ALS data and orthophotos, were also used in the image block alignment process.
- (ii)
- Dense point clouds were generated for each aerial imagery acquisition date using depth maps calculated through stereo-matching. The process of dense point cloud generation considered overlapping pairs of images, and the depth maps were combined to form a final dense point cloud, with excess information in overlapping regions that was used to filter out erroneous depth measurements.
- (iii)
- Digital Surface Models (DSMs) with a resolution of 0.5 m were generated from the dense point clouds for each data collection date. These models were then normalised and transformed into CHMs with a spatial resolution of 0.5 m. The normalisation process employed a Digital Terrain Model (DTM) with a resolution of 0.5 m, created from ALS data.
2.4. Processing of ALS Data
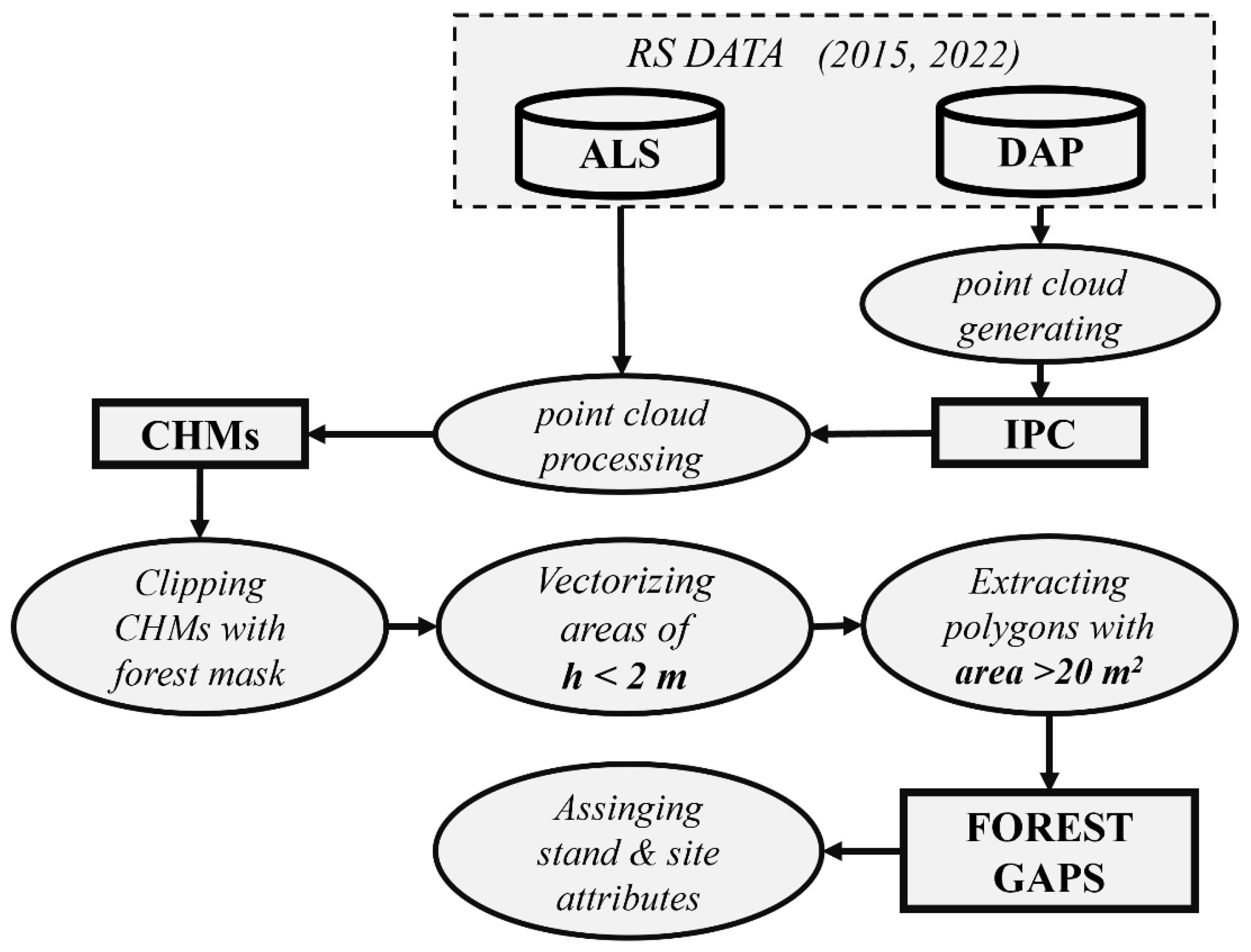
2.5. Gap Detection Based on Canopy Height Models
2.6. Statistical Analyses
3. Results
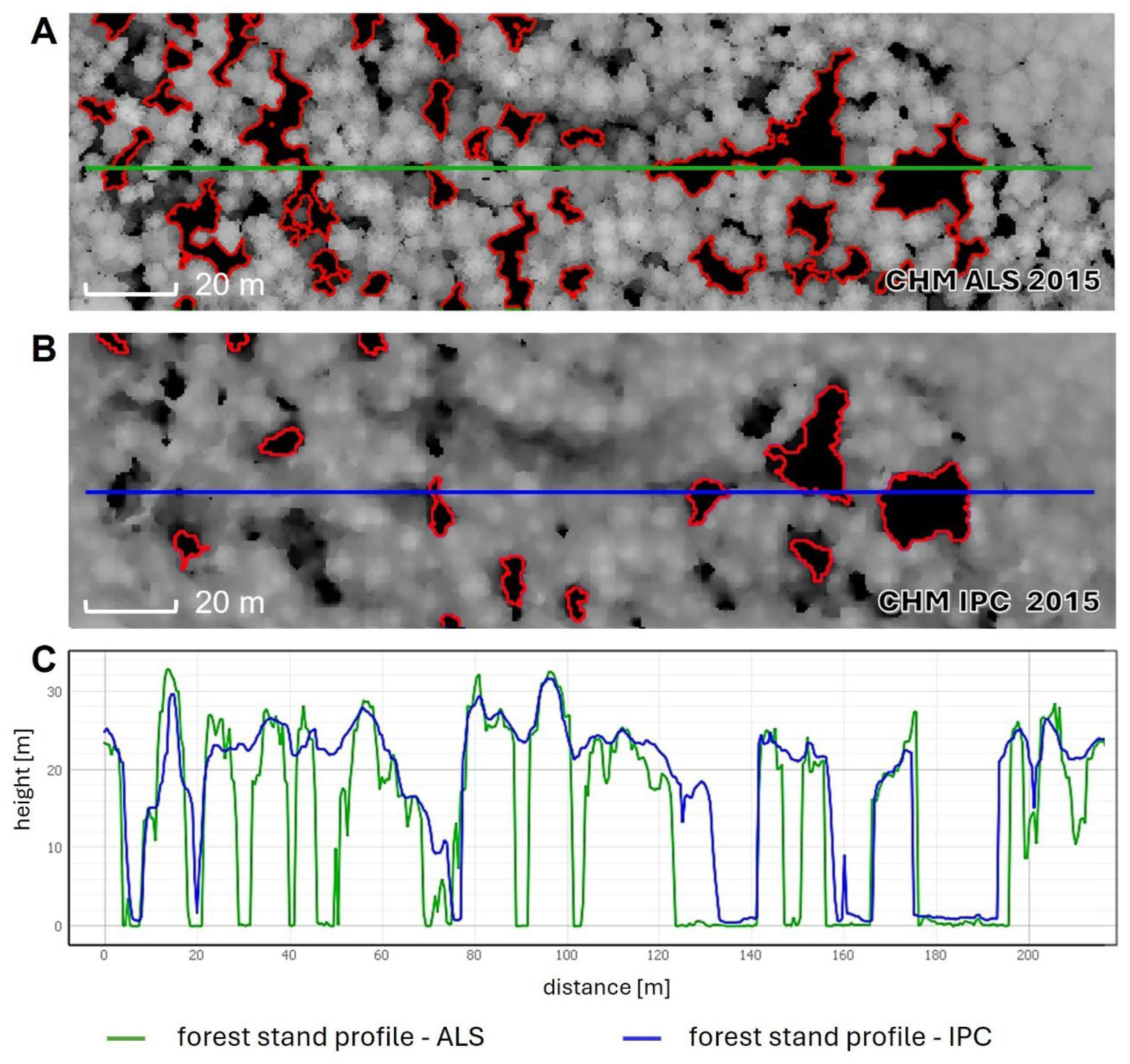
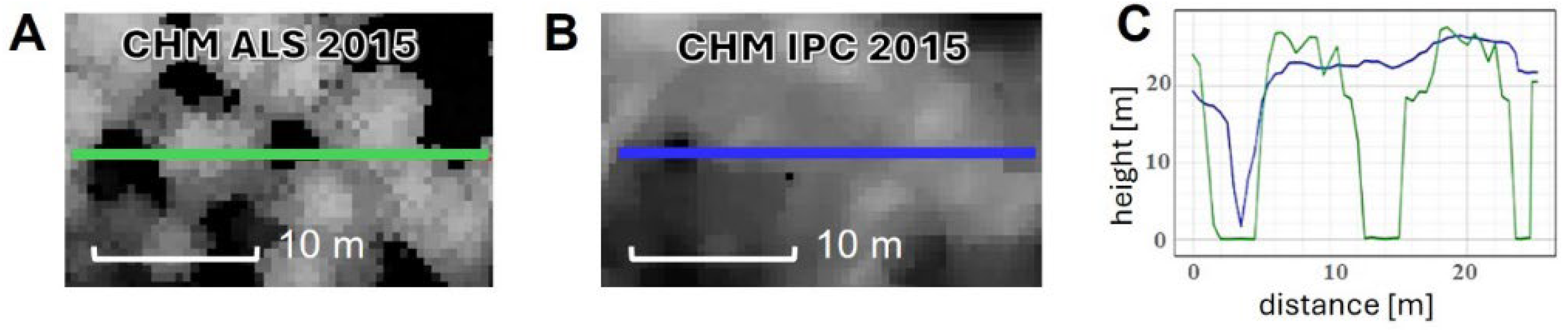
3.1. Comparison of Canopy Height Models—ALS vs. IPC
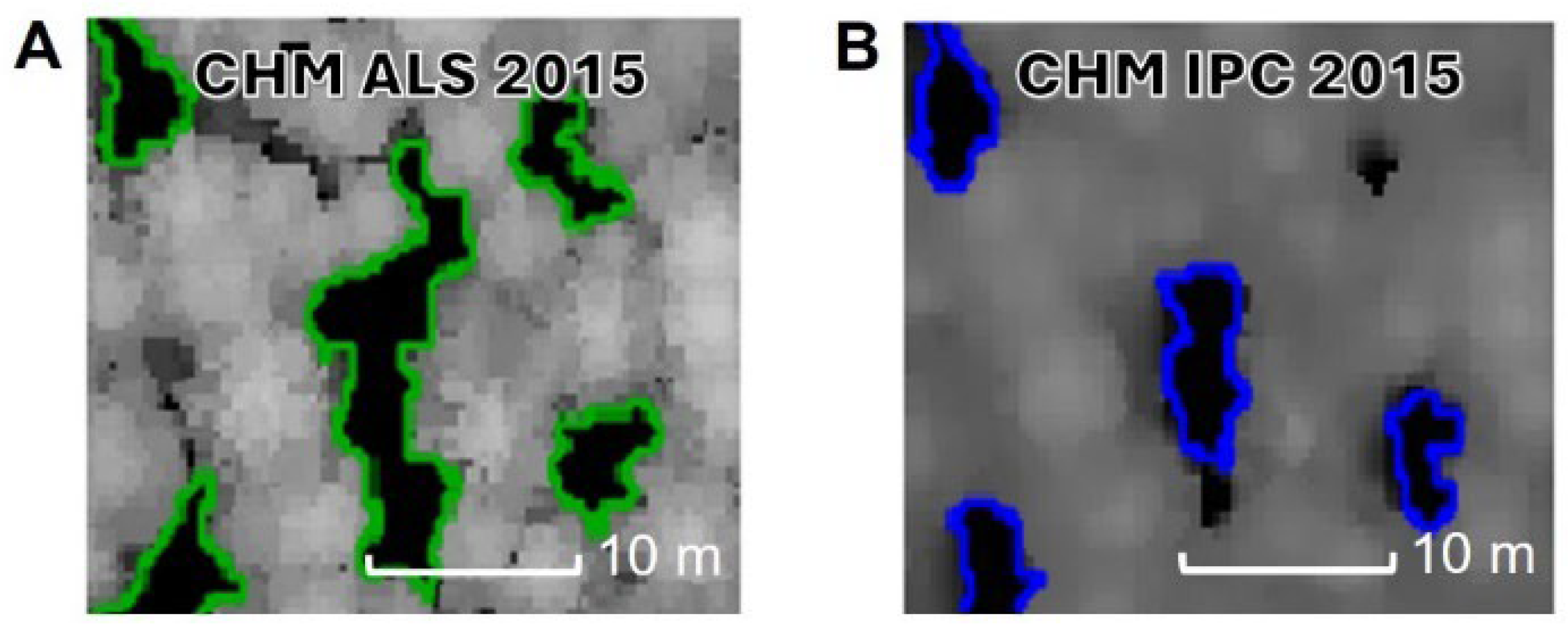
3.2. Canopy Gaps Features—ALS vs. IPC
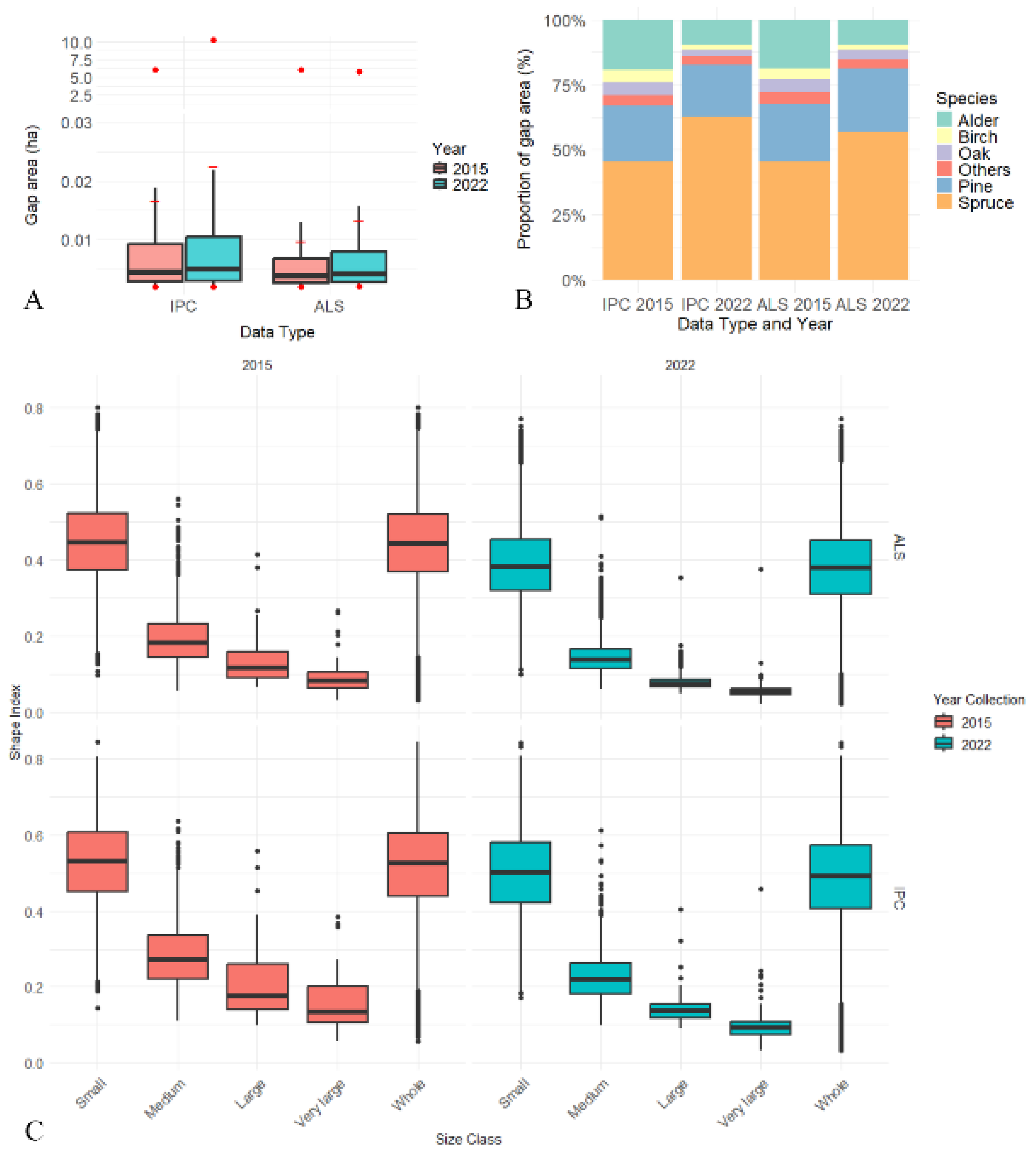
3.2.1. Gap Area
3.2.2. Dominant Tree Species vs. Spatial Distribution of Gaps
3.2.3. Gap Shape
3.2.4. Distribution of Gap Size
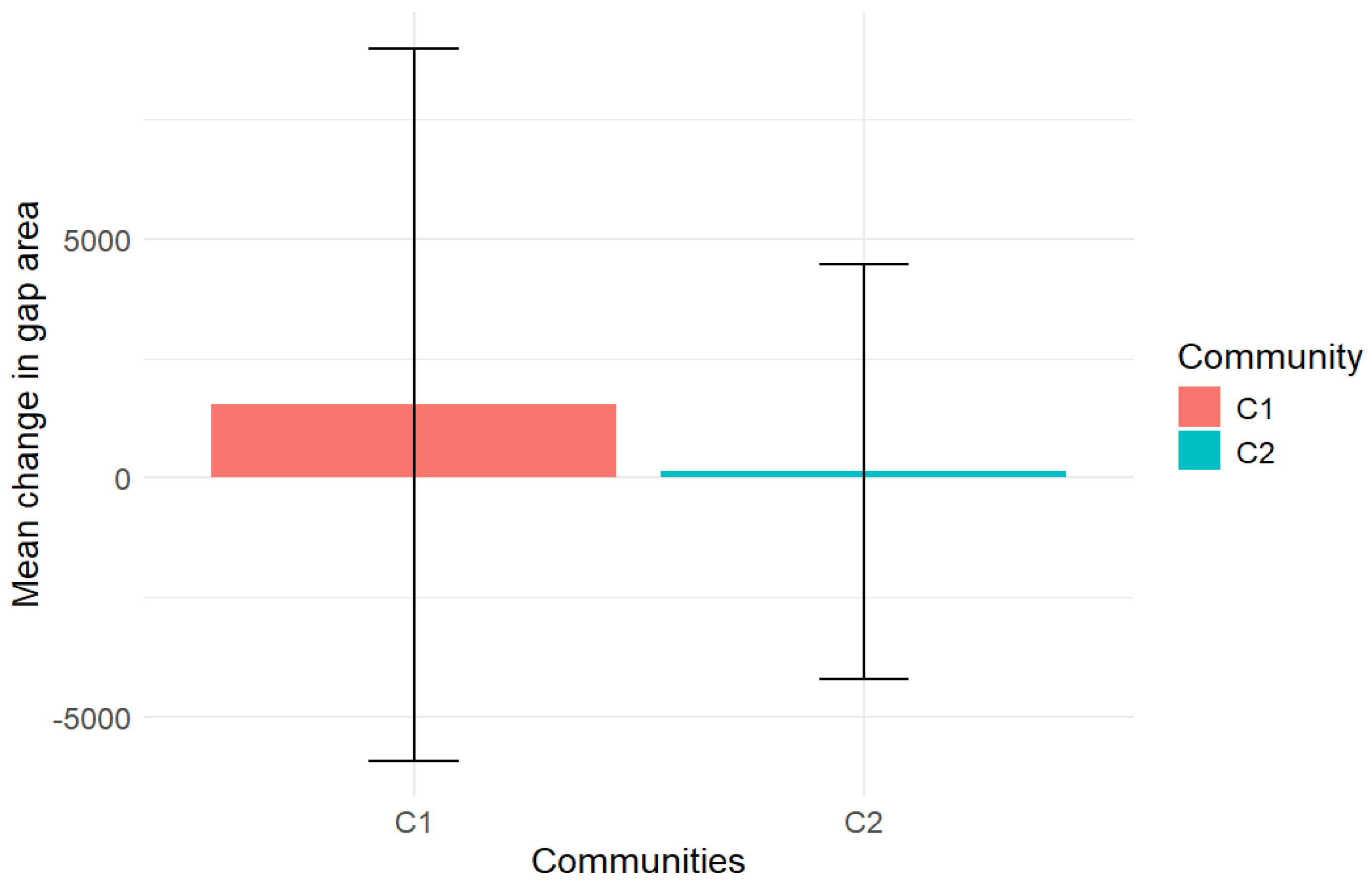
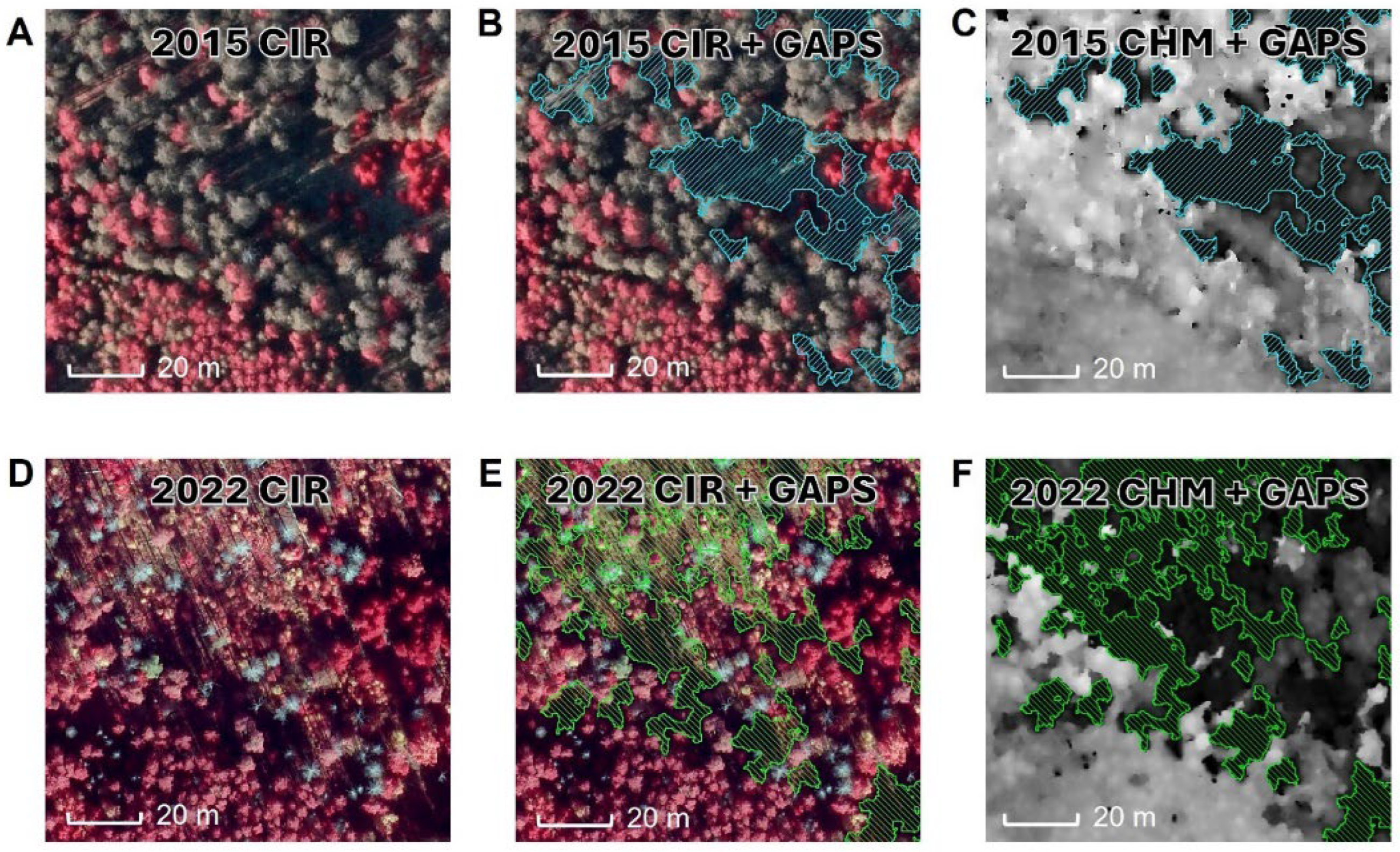
3.3. Gap Dynamics from 2015 to 2022
4. Discussion
4.1. Comparison of Height Information Obtained from Canopy Height Models—ALS vs. IPC
4.2. Size, Shape, and Number of Gaps
4.3. Site and Stand Factors Influencing the Spatial Distribution of Canopy Gaps
4.4. Gap Dynamics from 2015 to 2022
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kuuluvainen, T. Gap disturbance, ground microtopography, and the regeneration dynamics of boreal coniferous forests in Finland: A review. Ann. Zool. Fenn. 1994, 31, 35–51. [Google Scholar]
- Yamamoto, S.-I. Forest gap dynamics and tree regeneration. J. For. Res. 2000, 5, 223–229. [Google Scholar] [CrossRef]
- Dobrowolska, D. Rola luk w odnawianiu drzewostanów mieszanych w rezerwacie Jata. Sylwan 2007, 151, 14–25. [Google Scholar]
- Bolibok, L. Regulacja Warunków Wzrostu Odnowień na Gniazdach—Wpływ Parametrów Gniazd na Oddziaływanie Czynników Biotycznych. Sylwan 2009, 153, 733–744. [Google Scholar]
- Schliemann, S.A.; Bockheim, J.G. Methods for studying treefall gaps: A review. For. Ecol. Manag. 2011, 261, 1143–1151. [Google Scholar] [CrossRef]
- Brokaw, N.V.L. The Definition of Treefall Gap and Its Effect on Measures of Forest Dynamics. Biotropica 1982, 14, 158–160. [Google Scholar]
- Dobrowolska, D. Dynamika luk w drzewostanach mieszanych rezerwatu Jata. Sylwan 2006, 4, 61–75. [Google Scholar]
- Feldmann, E.; Drößler, L.; Hauck, M.; Kucbel, S.; Pichler, V.; Leuschner, C. Canopy Gap Dynamics and Tree Understory Release in a Virgin Beech Forest, Slovakian Carpathians. For. Ecol. Manag. 2018, 415–416, 38–46. [Google Scholar] [CrossRef]
- Muscolo, A.; Bagnato, S.; Sidari, M.; Mercurio, R. A review of the roles of forest canopy gaps. J. For. Res. 2014, 25, 725–736. [Google Scholar] [CrossRef]
- Bonnet, S.; Gaulton, R.; Lehaire, F.; Lejeune, P. Canopy Gap Mapping from Airborne Laser Scanning: An Assessment of the Positional and Geometrical Accuracy. Remote Sens. 2015, 7, 11267–11294. [Google Scholar] [CrossRef]
- Vilhar, U.; Roženbergar, D.; Simončič, P.; Diaci, J. Variation in irradiance, soil features and regeneration patterns in experimental forest canopy gaps. Ann. For. Sci. 2015, 72, 253–266. [Google Scholar] [CrossRef]
- de Carvalho, L.M.T.; Fontes, M.A.L.; Oliveira-Filho, A.T. Tree Species Distribution in Canopy Gaps and Mature Forest in an Area of Cloud Forest of the Ibitipoca Range, South-Eastern Brazil. Plant Ecol. 2000, 149, 9–22. [Google Scholar] [CrossRef]
- Wagner, S.; Fischer, H.; Huth, F. Canopy effects on vegetation caused by harvesting and regeneration treatments. Eur. J. For. Res. 2011, 130, 17–40. [Google Scholar] [CrossRef]
- Kuijper, D.P.J.; Cromsigt, J.P.G.M.; Churski, M.; Adam, B.; Jędrzejewska, B.; Jędrzejewski, W. Do ungulates preferentially feed in forest gaps in European temperate forest? For. Ecol. Manag. 2009, 258, 1528–1535. [Google Scholar] [CrossRef]
- Schnitzer, S.A.; Mascaro, J.; Carson, W.P. Treefall gaps and the maintenance of plant species diversity in tropical forests. In Tropical Forest Community Ecology; Carson, W.P., Schnitzer, S.A., Eds.; Blackwell Publishing: West Sussex, UK, 2008; pp. 196–209. [Google Scholar]
- Gray, A.N.; Spies, T.A.; Pabst, R.J. Canopy Gaps Affect Long-Term Patterns of Tree Growth and Mortality in Mature and Old-Growth Forests in the Pacific Northwest. For. Ecol. Manag. 2012, 281, 111–120. [Google Scholar] [CrossRef]
- Hammond, M.E.; Pokorný, R. Impact of Canopy Gap Ecology on the Diversity and Dynamics of Natural Regeneration in a Tropical Moist Semi-Deciduous Forest, Ghana. Biol. Life Sci. Forum 2021, 2, 10. [Google Scholar] [CrossRef]
- Kucbel, S.; Jaloviar, P.; Saniga, M.; Vencurik, J.; Klimaš, V. Canopy gaps in an old-growth fir-beech forest remnant of Western Carpathians. Eur. J. For. Res. 2010, 129, 249–259. [Google Scholar] [CrossRef]
- Zhu, C.; Zhu, J.; Wang, G.G.; Zheng, X.; Lu, D.; Gao, T. Dynamics of gaps and large openings in a secondary forest of Northeast China over 50 years. Ann. For. Sci. 2019, 76, 72. [Google Scholar] [CrossRef]
- Yamamoto, S.-I.; Nishimura, N.; Torimaru, T.; Manabe, T.; Itaya, A.; Becek, K. A comparison of different survey methods for assessing gap parameters in old-growth forests. For. Ecol. Manag. 2011, 262, 886–893. [Google Scholar] [CrossRef]
- Silva, C.A.; Valbuena, R.; Pinagé, E.R.; Mohan, M.; de Almeida, D.R.A.; Broadbent, E.N.; Jaafar, W.S.W.M.; de Almeida Papa, D.; Cardil, A.; Klauberg, C. ForestGapR: An r Package for forest gap analysis from canopy height models. Methods Ecol. Evol. 2019, 10, 1347–1356. [Google Scholar] [CrossRef]
- Zheng, G.; Shen, G.; Moskal, L.M. Characterizing spatiotemporal variations of forest canopy gaps using aerial laser scanning data. Int. J. Appl. Earth Obs. Geoinf. 2021, 104, 102588. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Franklin, J.F. Conserving Forest Biodiversity: A Comprehensive Multiscaled Approach; Island Press: Washington, DC, USA, 2002. [Google Scholar] [CrossRef]
- Diaci, J.; Adamič, T.; Rozman, A.; Fidej, G.; Roženbergar, D. Conversion of Pinus nigra Plantations with Natural Regeneration in the Slovenian Karst: The Importance of Intermediate, Gradually Formed Canopy Gaps. Forests 2019, 10, 1136. [Google Scholar] [CrossRef]
- Runkle, J.R. Gap regeneration in some old-growth forests of the eastern United States. Ecology 1981, 62, 1041–1051. [Google Scholar] [CrossRef]
- Yamamoto, S.-I. Gap dynamics in climax Fagus crenata forests. Bot. Mag. Shokubutsu-Gaku-Zasshi 1989, 102, 93–114. [Google Scholar] [CrossRef]
- Dobrowolska, D.; Veblen, T.T. Treefall-Gap Structure and Regeneration in Mixed Abies alba Stands in Central Poland. For. Ecol. Manag. 2008, 255, 3469–3476. [Google Scholar] [CrossRef]
- Itaya, A.; Miura, M.; Yamamoto, S.-I. Canopy height changes of an old-growth evergreen broad-leaved forest analyzed with digital elevation models. For. Ecol. Manag. 2004, 194, 403–411. [Google Scholar] [CrossRef]
- St-Onge, B.; Vepakomma, U.; Sénécal, J.F.; Kneeshaw, D.; Doyon, F. Canopy gap detection and analysis with airborne laser scanning. In Forestry Applications of Airborne Laser Scanning: Concepts and Case Studies; Maltamo, M., Næsset, E., Vauhkonen, J., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 419–437. [Google Scholar]
- White, J.C.; Tompalski, P.; Coops, N.C.; Wulder, M.A. Comparison of airborne laser scanning and digital stereo imagery for characterizing forest canopy gaps in coastal temperate rainforests. Remote Sens. 2018, 10, 1065. [Google Scholar] [CrossRef]
- Winstanley, P.; Dalagnol, R.; Mendiratta, S.; Braga, D.; Galvão, L.S.; Bispo, P.d.C. Post-Logging Canopy Gap Dynamics and Forest Regeneration Assessed Using Airborne LiDAR Time Series in the Brazilian Amazon with Attribution to Gap Types and Origins. Remote Sens. 2024, 16, 2319. [Google Scholar] [CrossRef]
- Chung, C.-H.; Wang, J.; Deng, S.-L.; Huang, C. Analysis of Canopy Gaps of Coastal Broadleaf Forest Plantations in Northeast Taiwan Using UAV LiDAR and the Weibull Distribution. Remote Sens. 2022, 14, 667. [Google Scholar] [CrossRef]
- Chen, J.; Wang, L.; Jucker, T.; Da, H.; Zhang, Z.; Hu, J.; Zhang, J. Detecting Forest Canopy Gaps Using Unoccupied Aerial Vehicle RGB Imagery in a Species-Rich Subtropical Forest. Remote Sens. Ecol. Conserv. 2023, 9, 671–686. [Google Scholar]
- Baltsavias, E. A Comparison between Photogrammetry and Laser Scanning. ISPRS J. Photogramm. Remote Sens. 1999, 54, 83–94. [Google Scholar] [CrossRef]
- Hyyppä, J.; Holopainen, M.; Olsson, H. Laser Scanning in Forests. Remote Sens. 2012, 4, 2919–2922. [Google Scholar] [CrossRef]
- Wallace, L.; Lucieer, A.; Malenovský, Z.; Turner, D.; Vopěnka, P. Assessment of Forest Structure Using Two UAV Techniques: A Comparison of Airborne Laser Scanning and Structure from Motion (SfM) Point Clouds. Forests 2016, 7, 62. [Google Scholar] [CrossRef]
- White, J.C.; Wulder, M.A.; Vastaranta, M.; Coops, N.C.; Pitt, D.; Woods, M. The utility of image-based point clouds for forest inventory: A comparison with airborne laser scanning. Forests 2013, 4, 518–536. [Google Scholar] [CrossRef]
- Erfanifard, Y.; Stereńczak, K.; Kraszewski, B.; Kamińska, A. Development of a Robust Canopy Height Model Derived from ALS Point Clouds for Predicting Individual Crown Attributes at the Species Level. Int. J. Remote Sens. 2018, 39, 9206–9227. [Google Scholar] [CrossRef]
- Mielcarek, M.; Stereńczak, K.; Khosravipour, A. Testing and evaluating different LiDAR-derived canopy height model generation methods for tree height estimation. Int. J. Appl. Earth Obs. Geoinf. 2018, 71, 132–143. [Google Scholar] [CrossRef]
- Sterenczak, K.; Zasada, M.; Brach, M. The accuracy assessment of DTM generated from LIDAR data for forest area–a case study for scots pine stands in Poland. Balt. For. 2013, 19, 252–262. [Google Scholar]
- Rahlf, J.; Breidenbach, J.; Solberg, S.; Næsset, E.; Astrup, R. Comparison of four types of 3D data for timber volume estimation. Remote Sens. Environ. 2014, 155, 325–333. [Google Scholar] [CrossRef]
- White, J.; Stepper, C.; Tompalski, P.; Coops, N.; Wulder, M. Comparing ALS and image-based point cloud metrics and modelled forest inventory attributes in a complex coastal forest environment. Forests 2015, 6, 3704–3732. [Google Scholar] [CrossRef]
- Goodbody, T.R.; Coops, N.C.; White, J.C. Digital Aerial Photogrammetry for Updating Area-Based Forest Inventories: A Review of Opportunities, Challenges, and Future Directions. Curr. For. Rep. 2019, 5, 55–75. [Google Scholar] [CrossRef]
- Htun, N.M.; Owari, T.; Tsuyuki, S.; Hiroshima, T. Detecting Canopy Gaps in Uneven-Aged Mixed Forests through the Combined Use of Unmanned Aerial Vehicle Imagery and Deep Learning. Drones 2024, 8, 484. [Google Scholar] [CrossRef]
- Zielewska-Büttner, K.; Adler, P.; Ehmann, M.; Braunisch, V. Automated detection of forest gaps in spruce dominated stands using canopy height models derived from stereo aerial imagery. Remote Sens. 2016, 8, 175. [Google Scholar] [CrossRef]
- Dietmaier, A.; McDermid, G.J.; Rahman, M.M.; Linke, J.; Ludwig, R. Comparison of LiDAR and Digital Aerial Photogrammetry for Characterizing Canopy Openings in the Boreal Forest of Northern Alberta. Remote Sens. 2019, 11, 1919. [Google Scholar] [CrossRef]
- Seidl, R.; Thom, D.; Kautz, M.; Martin-Benito, D.; Peltoniemi, M.; Vacchiano, G.; Wild, J.; Ascoli, D.; Petr, M.; Honkaniemi, J.; et al. Forest disturbances under climate change. Nat. Clim. Chang. 2017, 7, 395–402. [Google Scholar] [CrossRef]
- Jaworski, T.; Hilszczański, J. The effect of temperature and humidity changes on insect development and their impact on forest ecosystems in the context of expected climate change. For. Res. Pap. 2013, 74, 345–355. [Google Scholar]
- Wermelinger, B.; Seifert, M. Temperature-dependent reproduction of the spruce bark beetle Ips typographus, and analysis of the potential population growth. Ecol. Entomol. 1999, 24, 103–110. [Google Scholar]
- Meddens, A.J.H.; Hicke, J.A.; Ferguson, C.A. Spatiotemporal patterns of observed bark beetle-caused tree mortality in British Columbia and the western United States. Ecol. Appl. 2012, 22, 1876–1891. [Google Scholar] [CrossRef]
- Lausch, A.; Heurich, M.; Fahse, L. Spatio-temporal infestation patterns of Ips typographus (L.) in the Bavarian Forest National Park, Germany. Ecol. Indic. 2013, 31, 73–81. [Google Scholar] [CrossRef]
- Hicke, J.A.; Meddens, A.J.H.; Kolden, C.A. Recent Tree Mortality in the Western United States from Bark Beetles and Forest Fires. For. Sci. 2016, 62, 141–153. [Google Scholar] [CrossRef]
- Stereńczak, K.; Mielcarek, M.; Kamińska, A.; Kraszewski, B.; Piasecka, Ż.; Miścicki, S.; Heurich, M. Influence of selected habitat and stand factors on bark beetle Ips typographus (L.) outbreak in the Białowieża Forest. For. Ecol. Manag. 2020, 459, 117826. [Google Scholar] [CrossRef]
- Marini, L.; Økland, B.; Jönsson, A.M.; Bentz, B.; Carroll, A.; Forster, B.; Grégoire, J.-C.; Hurling, R.; Nageleisen, L.M.; Netherer, S.; et al. Climate drivers of bark beetle outbreak dynamics in Norway spruce forests. Ecography 2017, 40, 1426–1435. [Google Scholar] [CrossRef]
- Hlásny, T.; König, L.; Krokene, P.; Lindner, M.; Montagné-Huck, C.; Müller, J.; Qin, H.; Raffa, K.F.; Schelhaas, M.-J.; Svoboda, M.; et al. Bark Beetle Outbreaks in Europe: State of Knowledge and Ways Forward for Management. Curr. For. Rep. 2021, 7, 138–165. [Google Scholar] [CrossRef]
- Hlásny, T.; Zimová, S.; Merganičová, K.; Štěpánek, P.; Modlinger, R.; Turčáni, M. Devastating Outbreak of Bark Beetles in the Czech Republic: Drivers, Impacts, and Management Implications. For. Ecol. Manag. 2021, 490, 119075. [Google Scholar] [CrossRef]
- Seidl, R.; Rammer, W. Climate change amplifies the interactions between wind and bark beetle disturbances in forest landscapes. Landsc. Ecol. 2017, 32, 1485–1498. [Google Scholar] [CrossRef]
- Abdullah, H.; Skidmore, A.K.; Darvishzadeh, R.; Heurich, M. Sentinel-2 Accurately Maps Green-Attack Stage of European Spruce Bark Beetle (Ips typographus, L.) Compared with Landsat-8. Remote Sens. Ecol. Conserv. 2019, 5, 87–106. [Google Scholar] [CrossRef]
- Jaroszewicz, B.; Cholewińska, O.; Gutowski, J.M.; Samojlik, T.; Zimny, M.; Latałowa, M. Białowieża Forest—A Relic of the High Naturalness of European Forests. Forests 2019, 10, 849. [Google Scholar] [CrossRef]
- Ginszt, T.; Laskowska-Ginszt, A. Ten Years (2012–2021) of Spruce Bark Beetle Ips typographus (L.) Activity in the Bialowieza Forest District of the Bialowieza Primeval Forest. Sylwan 2022, 166, 183–193. [Google Scholar] [CrossRef]
- AgiSoft PhotoScan Professional Edition (Version 2.0) (Software). 2021. Available online: http://www.agisoft.com/downloads/installer (accessed on 11 February 2025).
- Roussel, J.-R.; Auty, D.; Coops, N.C.; Tompalski, P.; Tristan, R.H.; Goodbody, A.; Meador, S.; Bourdon, J.-F.; Boissieu, F.; Achim, A. lidR: An R package for analysis of Airborne Laser Scanning (ALS) data. Remote Sens. Environ. 2020, 251, 112061. [Google Scholar]
- Khosravipour, A.; Skidmore, A.K.; Isenburg, M.; Wang, T.; Hussin, Y.A. Generating pit-free canopy height models from airborne lidar. Photogramm. Eng. Remote Sens. 2014, 80, 863–872. [Google Scholar]
- Dobrowolska, D.; Piasecka, Ż.; Kuberski, Ł.; Stereńczak, K. Canopy Gap Characteristics and Regeneration Patterns in the Białowieża Forest Based on Remote Sensing Data and Field Measurements. For. Ecol. Manag. 2022, 511, 120123. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: https://www.R-project.org/ (accessed on 20 July 2024).
- Baltsavias, E.; Gruen, A.; Eisenbeiss, H.; Zhang, L.; Waser, L.T. High-Quality Image Matching and Automated Generation of 3D Tree Models. Int. J. Remote Sens. 2008, 29, 1243–1259. [Google Scholar] [CrossRef]
- St-Onge, B.; Vega, C.; Fournier, R.A.; Hu, Y. Mapping canopy height using a combination of digital stereo-photogrammetry and lidar. Int. J. Remote Sens. 2008, 29, 3343–3364. [Google Scholar] [CrossRef]
- Straub, C.; Stepper, C.; Seitz, R.; Waser, L.T. Potential of UltraCamX stereo images for estimating timber volume and basal area at the plot level in mixed European forests. Can. J. For. Res. 2013, 43, 731–741. [Google Scholar] [CrossRef]
- Vepakomma, U.; St-Onge, B.; Kneeshaw, D. Spatially explicit characterization of boreal forest gap dynamics using multi-temporal lidar data. Remote Sens. Environ. 2007, 112, 2326–2340. [Google Scholar] [CrossRef]
- Vepakomma, U.; St-Onge, B.; Kneeshaw, D. Response of a boreal forest to canopy opening: Assessing vertical and lateral tree growth with multi-temporal lidar data. Ecol. Appl. 2011, 21, 99–121. [Google Scholar] [CrossRef]
- Holopainen, M.; Vastaranta, M.; Karjalainen, M.; Karila, K.; Kaasalainen, S.; Honkavaara, E.; Hyyppa, J. Forest Inventory Attribute Estimation Using Airborne Laser Scanning, Aerial Stereo Imagery, Radargrammetry and Interferometry—Finnish Experiences of the 3D Techniques. ISPRS Ann. Photogramm. Remote Sens. Spat. Inf. Sci. 2015, II–3/W4, 63–69. [Google Scholar] [CrossRef]
- Heurich, M.; Schadeck, S.; Weinacker, H.; Krzystek, P. Forest Parameter Derivation from DTM/DSM Generated from LiDAR and Digital Modular Camera (DMC). Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. 2004, XXXV, 84–89. [Google Scholar]
- Mielcarek, M.; Kamińska, A.; Stereńczak, K. Digital aerial photogrammetry (DAP) and airborne laser scanning (ALS) as sources of information about tree height: Comparisons of the accuracy of remote sensing methods for tree height estimation. Remote Sens. 2020, 12, 1808. [Google Scholar] [CrossRef]
- Petzold, B.; Reiss, P.; Stössel, W. Laser scanning—Surveying and mapping agencies are using a new technique for the derivation of digital terrain models. ISPRS J. Photogramm. Remote Sens. 1999, 54, 95–104. [Google Scholar] [CrossRef]
- Rahlf, J. Forest Resource Mapping Using 3D Remote Sensing: Combining National Forest Inventory Data and Digital Aerial Photogrammetry; Norwegian University of Life Sciences: Ås, Norway, 2017. [Google Scholar]
- Fassnacht, F.E.; White, J.C.; Wulder, M.A.; Næsset, E. Remote Sensing in Forestry: Current Challenges, Considerations, and Directions. Forestry 2024, 97, 11–37. [Google Scholar] [CrossRef]
- Vastaranta, M.; Wulder, M.A.; White, J.C.; Pekkarinen, A.; Tuominen, S.; Ginzler, C.; Kankare, V.; Holopainen, M.; Hyyppä, J.; Hyyppä, H. Airborne laser scanning and digital stereo imagery measures of forest structure: Comparative results and implications to forest mapping and inventory update. Can. J. Remote Sens. 2013, 39, 382–395. [Google Scholar] [CrossRef]
- St-Onge, B.; Audet, F.-A.; Bégin, J. Characterizing the height structure and composition of a boreal forest using an individual tree crown approach applied to photogrammetric point clouds. Forests 2015, 6, 3899–3922. [Google Scholar] [CrossRef]
- Lertzman, K.P.; Sutherland, G.D.; Inselberg, A.; Saunders, S.C. Canopy gaps and the landscape mosaic in a coastal temperate rain forest. Ecology 1996, 77, 1254–1270. [Google Scholar] [CrossRef]
- Seidel, D.; Ammer, C.; Puettmann, K. Describing forest canopy gaps efficiently, accurately, and objectively: New prospects through the use of terrestrial laser scanning. Agric. For. Meteorol. 2015, 213, 23–32. [Google Scholar] [CrossRef]
- Hu, L.; Zhu, J. Determination of the Tridimensional Shape of Canopy Gaps Using Two Hemispherical Photographs. Agric. For. Meteorol. 2009, 149, 862–872. [Google Scholar] [CrossRef]
- Koukoulas, S.; Blackburn, G.A. Spatial relationships between tree species and gap characteristics in broad-leaved deciduous woodland. J. Veg. Sci. 2005, 16, 587–596. [Google Scholar] [CrossRef]
- Rodes-Blanco, M.; Ruiz-Benito, P.; Silva, C.A.; García, M. Canopy gap patterns in Mediterranean forests: A spatio-temporal characterization using airborne LiDAR data. Landsc. Ecol. 2023, 38, 3427–3442. [Google Scholar] [CrossRef]
- Brzeziecki, B.; Hilszczański, J.; Kowalski, T.; Łakomy, P.; Małek, S.; Miścicki, S.; Modrzyński, J.; Sowa, J.; Starzyk, J. Problem Masowego Zamierania Drzewostanów Świerkowych w Leśnym Kompleksie Promocyjnym “Puszcza Białowieska”. Sylwan 2018, 162, 373–386. [Google Scholar] [CrossRef]
- Erfanifard, Y.; Lisiewicz, M.; Stereńczak, K. High-Resolution Remote Sensing for Biodiversity Assessment and Monitoring: A Case Study of Dominant Tree Species in an Old-Growth Forest. For. Ecol. Manag. 2024, 566, 122094. [Google Scholar] [CrossRef]
- Storaunet, K.O.; Rolstad, J. Time since death and fall of Norway spruce logs in old-growth and selectively cut boreal forest. Can. J. For. Res. 2002, 32, 1801–1812. [Google Scholar] [CrossRef]
- Grodzki, W. Gradacyjne Występowanie Kornika Drukarza Ips typographus (L.) (Col.: Curculionidae, Scolytinae) w Aspekcie Kontrowersji Wokół Puszczy Białowieskiej. Leśne Pr. Badaw. 2016, 77, 324–331. [Google Scholar] [CrossRef]
- Krzywicka, A. Ochrona przyrody i gospodarka leśna w Puszczy Białowieskiej: Brak kompleksowej strategii działania. Kontrola Państwowa 2020, 65, 33–44. [Google Scholar]
- Vepakomma, U.; Kneeshaw, D.; Fortin, M.-J. Spatial contiguity and continuity of canopy gaps in mixed wood boreal forests: Persistence, expansion, shrinkage and displacement. J. Ecol. 2012, 100, 1257–1268. [Google Scholar] [CrossRef]
- Ali-Sisto, D.; Packalen, P. Forest Change Detection by Using Point Clouds from Dense Image Matching Together with a LiDAR-Derived Terrain Model. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2016, 10, 1197–1206. [Google Scholar] [CrossRef]
- Gutowski, J.M.; Jaroszewicz, B. Zmiany Udziału Świerka Pospolitego w Drzewostanach Puszczy Białowieskiej w Kontekście Dynamiki Liczebności Kornika Drukarza Ips typographus (L.). In Stan Ekosystemów Leśnych Puszczy Białowieskiej; Wikło, A., Ed.; CILP: Warszawa, Poland, 2016; pp. 87–108. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aerial Imagery | Airborne Laser Scanning | |||
|---|---|---|---|---|
| Parameter | 2015 | 2022 | 2015 | 2022 |
| Device | UltraCam Eagle | UltraCam Eagle | Riegl LMS-Q680i | Riegl VQ-780i |
| Sensor type | large format RGB-NIR camera | large format RGB-NIR camera | full waveform laser scanner | full waveform laser scanner |
| Coverage | 90/40% | 80/70% | 50% | 20% |
| Flight altitude | 3200 m | 3960 m | 500 m | 900 m |
| Resolution/ density | 0.20 m | 0.20 m | 11 pts/m2 | 38 pts/m2 |
| Flight date | July 2015 | October 2022 | July 2015 | October 2022 |
| Attribute | Description |
|---|---|
| Gap area | Geometric surface area of the gap [m2] |
| Gap size class | Small: area ≤ 500 m2 Medium: area > 500–3000 m2 Large: area > 3000–5000 m2 Very Large: area > 5000 m2 |
| Area of gap located in a nature reserve | Area of the gap located within nature reserve/-s [m2] |
| Tree species | Dominant tree species in the gap (in terms of the species’ spatial coverage). The area of the polygon covered by a given species (dominant tree species) was calculated based on information obtained from the Forest Digital Map for 2015 |
| Gap shape index | An index determining the level of complexity of the gap shape based on the surface and perimeter of the gap, calculated according to the formula shp_index = 1/(P/(2*sqrt(A*pi)), where P = perimeter, A = gap area. The shape index takes values from 0 to 1 (the closer the index value is to 1, the less complex the gap shape—more similar to a circle) |
| Method | Year | Gap Size Class | Total | ||||
|---|---|---|---|---|---|---|---|
| Small | Medium | Large | Very Large | ||||
| Total area of gaps by gap size class (ha) | ALS | 2015 | 291.92 | 100.54 | 27.41 | 66.64 | 486.50 |
| 2022 | 331.59 | 165.47 | 51.41 | 131.72 | 680.20 | ||
| IPC | 2015 | 131.20 | 75.85 | 27.28 | 77.90 | 312.22 | |
| 2022 | 232.49 | 150.00 | 52.24 | 268.33 | 703.06 | ||
| Total number of gaps by gap size class | ALS | 2015 | 48,755 | 952 | 71 | 60 | 49,838 |
| 2022 | 49,743 | 1517 | 134 | 123 | 51,517 | ||
| IPC | 2015 | 17,961 | 679 | 72 | 60 | 18,772 | |
| 2022 | 29,645 | 1351 | 135 | 201 | 31,332 | ||
| Newly Created | Overgrown | Overlapping | |||
|---|---|---|---|---|---|
| <25% | 25–75% | >75% | |||
| ALS 2015–2022 | 27,668 | 22,958 | 6453 | 10,097 | 7299 |
| IPC 2015–2022 | 23,478 | 9944 | 2653 | 3017 | 2184 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mielcarek, M.; Kurpiewska, S.; Guderski, K.; Dobrowolska, D.; Zin, E.; Kuberski, Ł.; Erfanifard, Y.; Stereńczak, K. Remote Sensing of Forest Gap Dynamics in the Białowieża Forest: Comparison of Multitemporal Airborne Laser Scanning and High-Resolution Aerial Imagery Point Clouds. Remote Sens. 2025, 17, 1149. https://doi.org/10.3390/rs17071149
Mielcarek M, Kurpiewska S, Guderski K, Dobrowolska D, Zin E, Kuberski Ł, Erfanifard Y, Stereńczak K. Remote Sensing of Forest Gap Dynamics in the Białowieża Forest: Comparison of Multitemporal Airborne Laser Scanning and High-Resolution Aerial Imagery Point Clouds. Remote Sensing. 2025; 17(7):1149. https://doi.org/10.3390/rs17071149
Chicago/Turabian StyleMielcarek, Miłosz, Sylwia Kurpiewska, Kacper Guderski, Dorota Dobrowolska, Ewa Zin, Łukasz Kuberski, Yousef Erfanifard, and Krzysztof Stereńczak. 2025. "Remote Sensing of Forest Gap Dynamics in the Białowieża Forest: Comparison of Multitemporal Airborne Laser Scanning and High-Resolution Aerial Imagery Point Clouds" Remote Sensing 17, no. 7: 1149. https://doi.org/10.3390/rs17071149
APA StyleMielcarek, M., Kurpiewska, S., Guderski, K., Dobrowolska, D., Zin, E., Kuberski, Ł., Erfanifard, Y., & Stereńczak, K. (2025). Remote Sensing of Forest Gap Dynamics in the Białowieża Forest: Comparison of Multitemporal Airborne Laser Scanning and High-Resolution Aerial Imagery Point Clouds. Remote Sensing, 17(7), 1149. https://doi.org/10.3390/rs17071149