
Research on the Response Mechanism of Vegetation to Drought Stress in the West Liao River Basin, China


Abstract
1. Introduction
2. Materials and Methods
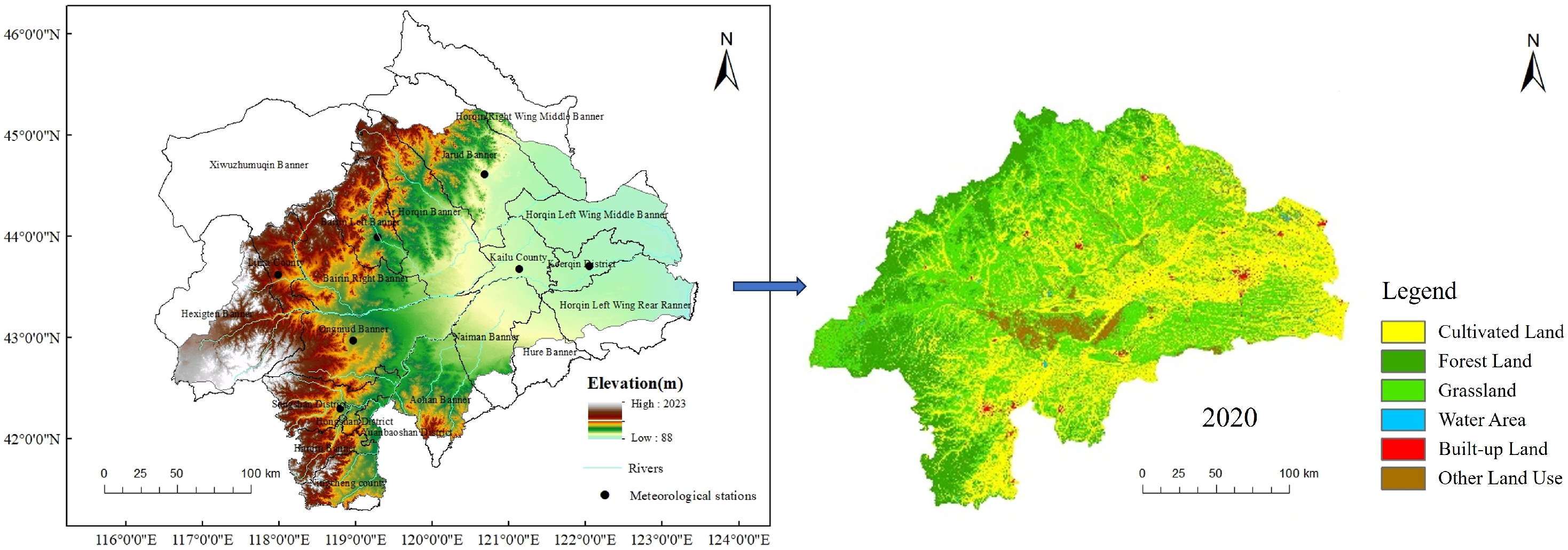
2.1. Study Area
2.2. Data Sources and Processing
2.3. Maximum Value Composite (MVC)
2.4. Sen’s Trend Analysis and Mann–Kendall Test
2.5. Pearson Correlation Analysis
2.6. Vegetation Resistance and Resilience
3. Results
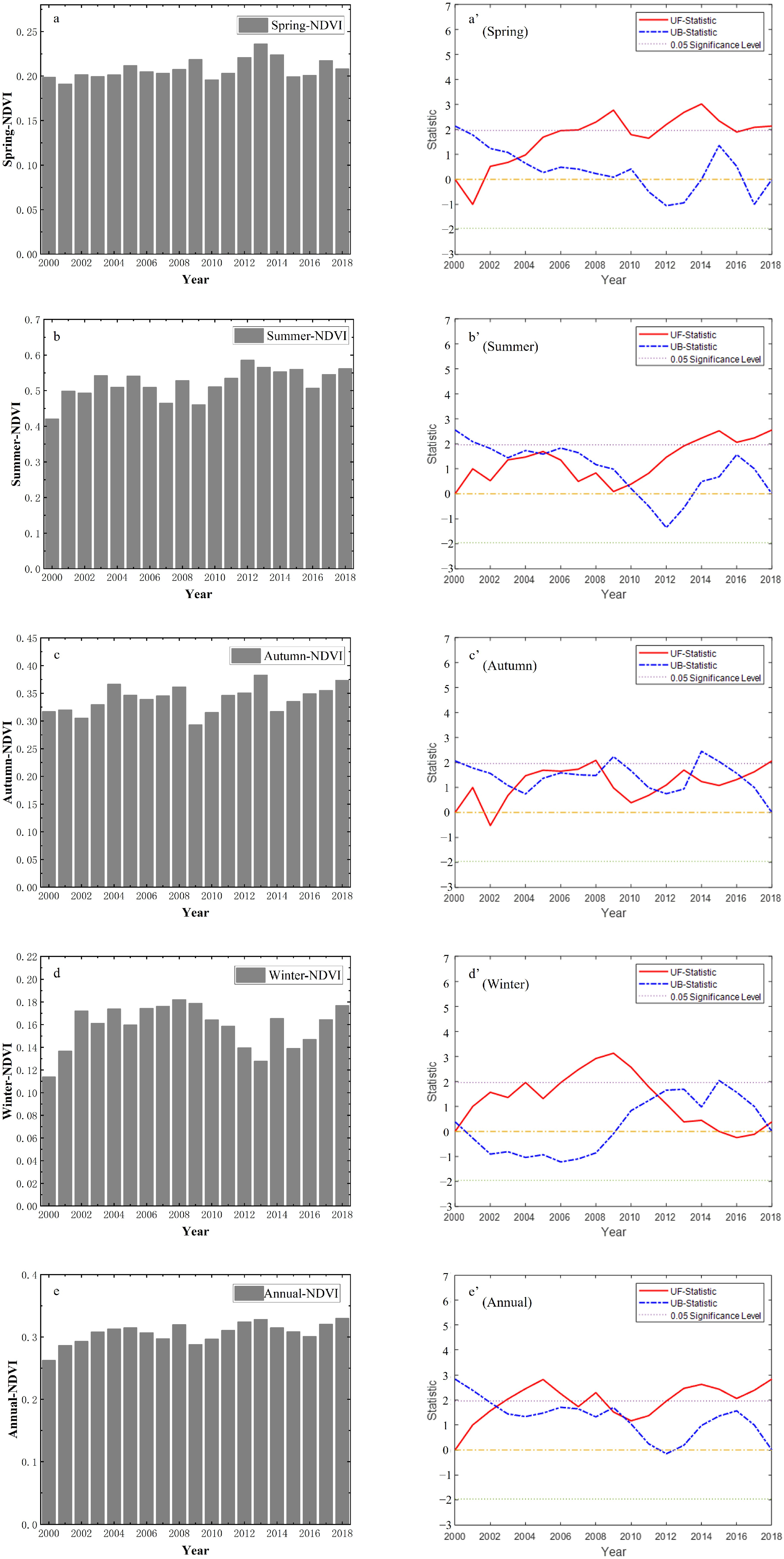
3.1. Temporal Variation Trends of NDVI
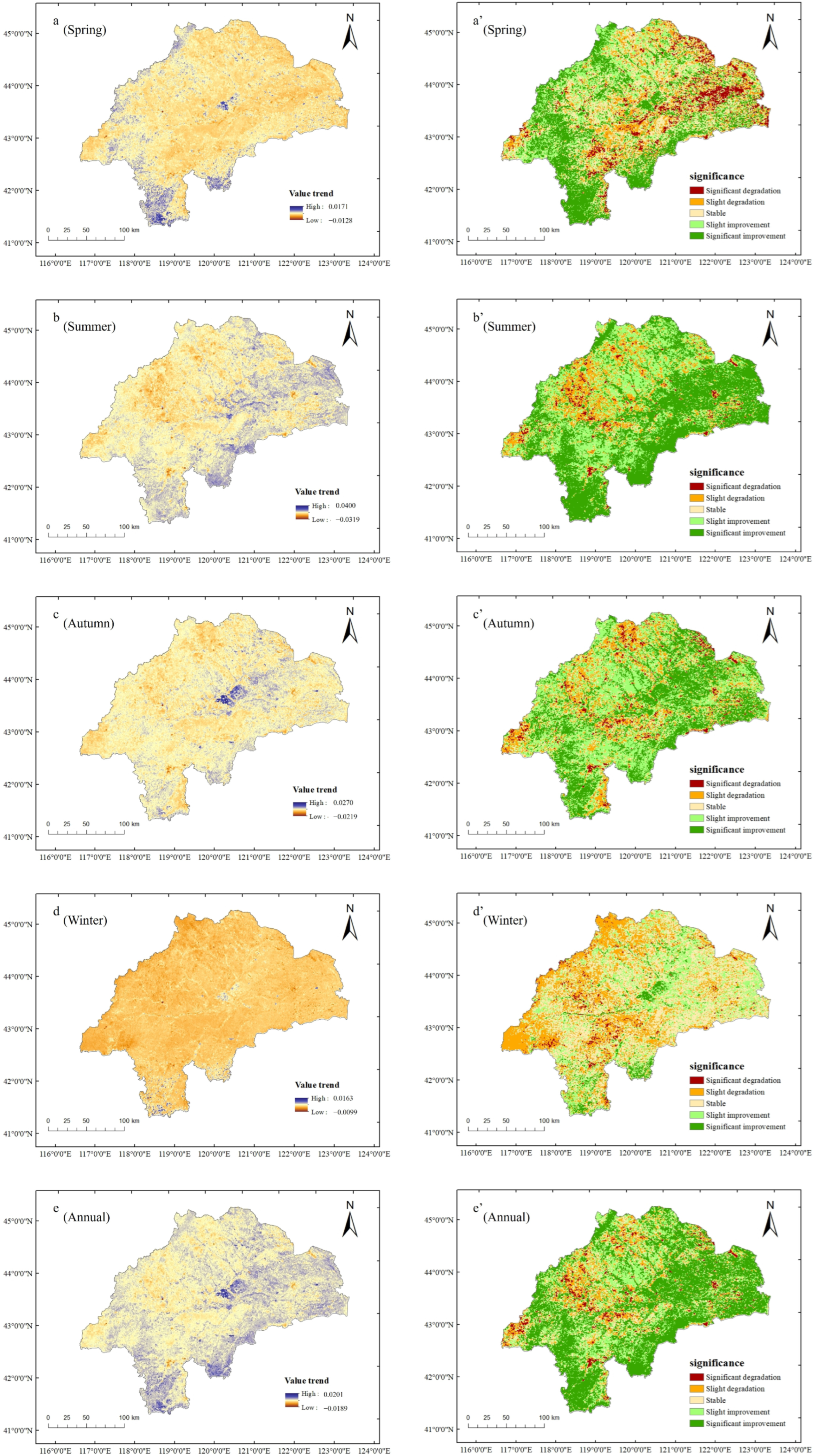
3.2. Spatial Variation Trends of NDVI
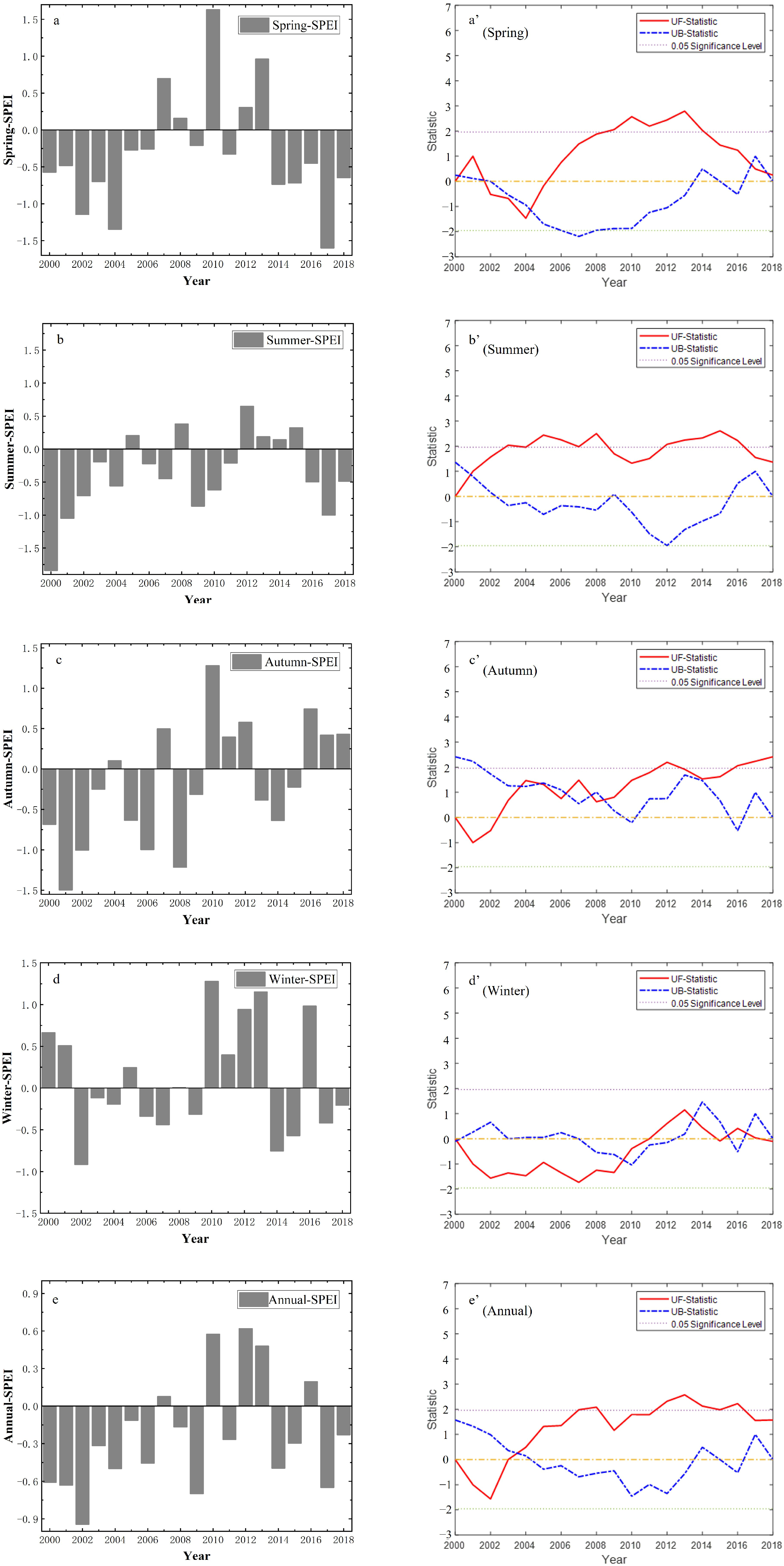
3.3. Temporal Variation Trends of SPEI
3.4. Spatial Variation Trends of SPEI
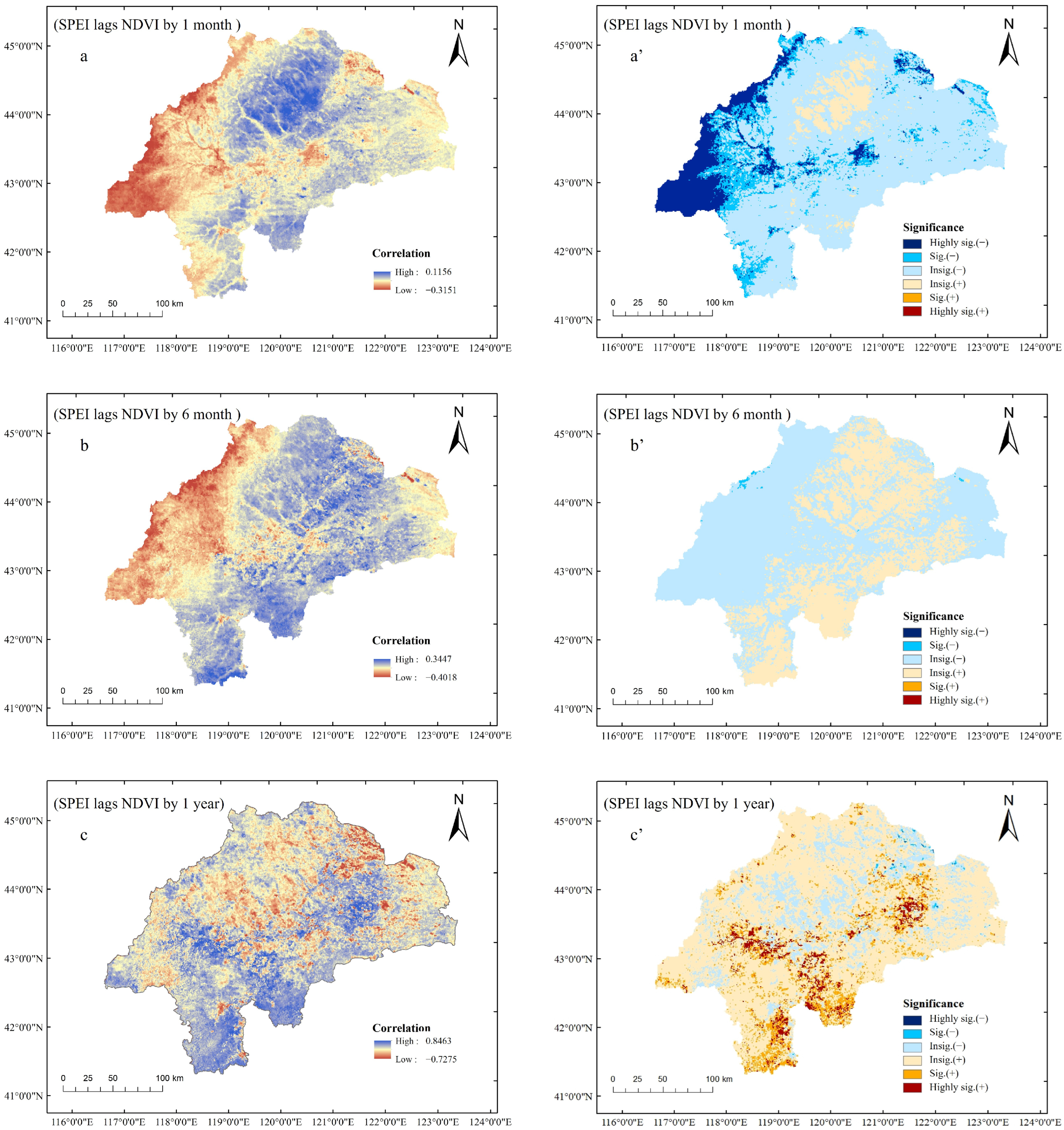
3.5. Sensitivity Analysis of Vegetation to Different Types of Drought Stress
3.6. Analysis of Vegetation Resistance and Resilience to Drought
4. Discussion
4.1. Responses of Different Vegetation Types to Short-, Medium-, and Long-Term Droughts
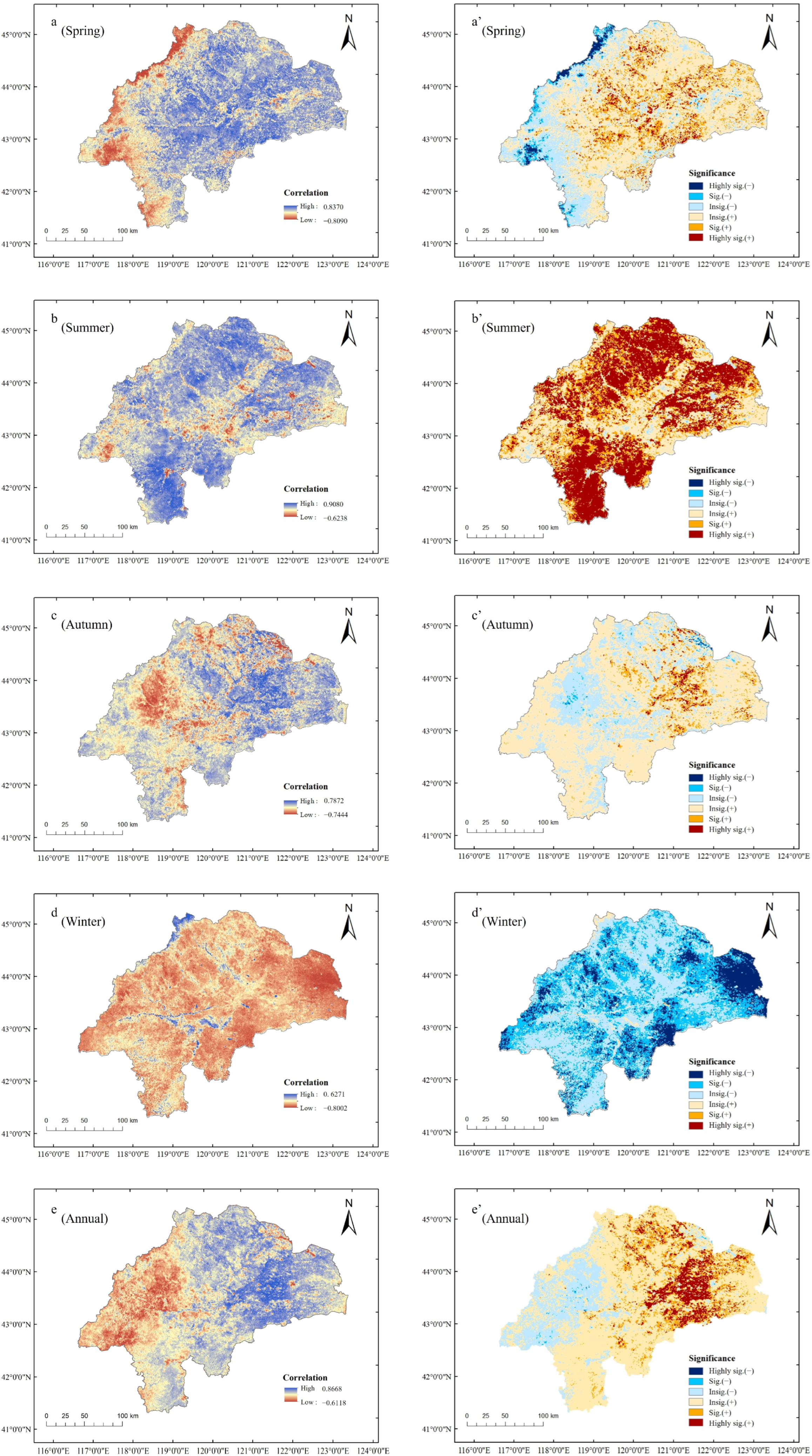
4.2. Seasonal Drought Trends and Heterogeneous Vegetation Responses
4.3. Spatial Heterogeneity of Vegetation Resistance and Resilience to Drought
4.4. Global Implications for Arid and Semi-Arid Regions
- (1)
- Regional adaptive management of resistance and resilience: The spatial heterogeneity of vegetation resistance and resilience offers insights for other regions. For example, in Africa’s Sahel, drought-tolerant species and ecological water projects could mitigate land degradation [57]. Similarly, in Australia’s Murray–Darling Basin, this study’s methods could inform vegetation monitoring and restoration strategies [58].
- (2)
- Global applicability of seasonal drought impacts: The seasonal variability of drought impacts provides a framework for managing droughts elsewhere. In the Mediterranean, optimizing irrigation and crop structures could alleviate summer drought effects [59]. In the U.S. Southwest, winter drought management strategies (e.g., soil moisture conservation) could support spring regrowth.
- (3)
- Adaptive ecosystem management under drought trends: The long-term and cumulative effects of drought highlight the need for adaptive management. For example, in Central Asia, restoring natural vegetation and optimizing water resources could enhance ecosystem resilience [60]. In South America’s Pampas, this study’s framework could guide ecosystem monitoring and management [61].
- (4)
- International collaboration and knowledge sharing: This study’s findings can support global initiatives like the UNCCD, fostering knowledge exchange and technical cooperation. Additionally, the methods and results can improve global ecosystem models (e.g., DGVMs), enhancing predictions of drought impacts on vegetation.
4.5. Limitations and Future Prospects
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yin, G.; Zhang, H. A new integrated index for drought stress monitoring based on decomposed vegetation response factors. J. Hydrol. 2023, 618, 129252. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Schwalm, C.R.; Biondi, F.; Camarero, J.J.; Koch, G.W.; Litvak, M.; Ogle, K.; Shaw, J.D.; Shevliakova, E.; Williams, A.P.; et al. Pervasive drought legacies in forest ecosystems and their implications for carbon cycle models. Science 2015, 349, 528–532. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Trugman, A.T.; Badgley, G.; Koings, A.G.; Shaw, J. Divergent forest sensitivity to repeated extreme droughts. Nat. Clim. Change 2020, 10, 1091–1095. [Google Scholar] [CrossRef]
- Gu, L.; Chen, J.; Yin, J.; Xu, C.-Y.; Chen, H. Drought hazard transferability from meteorological to hydrological propagation. J. Hydrol. 2020, 585, 124761. [Google Scholar] [CrossRef]
- Rahman, G.; Jung, K.M.; Kim, W.T.; Kwan, H.H. Drought impact, vulnerability, risk assessment, management and mitigation under climate change: A comprehensive review. KSCE J. Civ. Eng. 2025, 29, 100120. [Google Scholar] [CrossRef]
- Liu, X.; Sun, G.; Fu, Z.; Ciais, P.; Feng, X.; Li, J.; Fu, B. Compound droughts slow down the greening of the Earth. Glob. Change Biol. 2023, 29, 3072–3084. [Google Scholar] [CrossRef]
- Tangjialeke, W.; Zou, J.; Ding, J.; Yahefujiang, H.; Huang, S.; Li, J. Analysis of drought response thresholds and drought-causing factors of central Asian vegetation. Ecol. Indic. 2024, 169, 112926. [Google Scholar] [CrossRef]
- Wang, C.; Chen, J.; Lee, S.-C.; Xiong, L.; Su, T.; Lin, Q.; Xu, C.-Y. Response and recovery times of vegetation productivity under drought stress: Dominant factors and relationships. J. Hydrol. 2025, 655, 132945. [Google Scholar] [CrossRef]
- Zhao, Q.; Zhang, X.; Li, C.; Xu, Y.; Fei, J.; Hao, F.; Song, R. Diverse vegetation response to meteorological drought from propagation perspective using event matching method. J. Hydrol. 2025, 653, 132776. [Google Scholar] [CrossRef]
- Wu, B.; Quan, Q.; Huang, X.; Deng, J. Response of extreme precipitation to climatic warming in the Weihe river basin, China and its mechanism. Open Geosci. 2022, 14, 853–862. [Google Scholar] [CrossRef]
- De Keersmaecker, W.; Lhermitte, S.; Tits, L.; Honnay, O.; Somers, B.; Coppin, P. A model quantifying global vegetation resistance and resilience to short-term climate anomalies and their relationship with vegetation cover. Glob. Ecol. Biogeogr. 2021, 30, 714–728. [Google Scholar] [CrossRef]
- Choat, B.; Brodribb, T.J.; Brodersen, C.R.; Duursma, R.A.; López, R.; Medlyn, B.E. Triggers of tree mortality under drought. Nature 2022, 608, 528–533. [Google Scholar] [CrossRef] [PubMed]
- Schwalm, C.R.; Anderegg, W.R.L.; Michalak, A.M.; Fisher, J.B.; Biondi, F.; Koch, G.; Huntzinger, D.N. Global patterns of drought recovery. Nature 2020, 548, 202–205. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, B. The responses of natural vegetation dynamics to drought during the growing season across China. J. Hydrol. 2019, 574, 706–714. [Google Scholar] [CrossRef]
- Craine, J.M.; Ocheltree, T.W.; Nippert, J.B.; Towne, E.G.; Skibbe, A.M.; Kembel, S.W.; Fargione, J.E. Global diversity of drought tolerance and grassland climate-change resilience. Nat. Clim. Change 2013, 3, 63–67. [Google Scholar] [CrossRef]
- Gazol, A.; Camarero, J.J.; Anderegg, W.R.L.; Vicente-Serrano, S.M. Impacts of droughts on the growth resilience of Northern Hemisphere forests. Glob. Ecol. Biogeogr. 2017, 26, 166–176. [Google Scholar] [CrossRef]
- Stampfli, A.; Bloor, J.M.G.; Fischer, M.; Zeiter, M. High land-use intensity exacerbates shifts in grassland vegetation composition after severe experimental drought. Glob. Change Biol. 2018, 24, 2021–2034. [Google Scholar] [CrossRef]
- Kannenberg, S.A.; Maxwell, J.T.; Pederson, N.; D’Orangeville, L.; Ficklin, D.L.; Phillips, R.P. Drought legacies are dependent on water table depth, wood anatomy and drought timing across the eastern US. Ecol. Lett. 2019, 22, 119–127. [Google Scholar] [CrossRef]
- Jiang, W.; Fu, B.; Gao, G.; Lv, Y.; Wang, C.; Sun, S.; Wang, K.; Schüler, S.; Shu, Z. Exploring spatial-temporal driving factors for changes in multiple ecosystem services and their relationships in West Liao River Basin, China. Sci. Total Environ. 2023, 904, 166716. [Google Scholar] [CrossRef]
- Gong, J.; Yang, G.; Zhang, S.; Zhang, W.; Dong, X.; Zhang, S.; Wang, R.; Yan, C.; Wang, T. Human activities weaken the positive effects of soil abiotic factors and biodiversity on ecosystem multifunctionality more than drought: A case study in China’s West Liao River Basin. Sci. Total Environ. 2024, 957, 177564. [Google Scholar] [CrossRef]
- Cao, J.; Luo, Y.; Zhang, X.; Fan, L.; Tao, J.; Nam, W.-H.; Sur, C.; He, Y.; Gulakhmadov, A.; Niyogi, D. Assessing the responsiveness of multiple microwave remote sensing vegetation optical depth indices to drought on crops in Midwest US. Int. J. Appl. Earth Obs. Geoinf. 2024, 132, 104072. [Google Scholar] [CrossRef]
- Zhao, X.; Fan, P.; Wang, Y.; Zhou, Q. Study on the spatiotemporal variation characteristics and influencing factors of vegetation coverage in the West Liao River Basin from 2000 to 2018. Chin. J. Agric. Resour. Reg. Plan. 2021, 42, 75–88. [Google Scholar]
- Wang, Z.; Liu, D.; Wang, M. Mapping Main Grain Crops and Change Analysis in the West Liaohe River Basin with Limited Samples Based on Google Earth Engine. Remote Sens. 2023, 15, 5515. [Google Scholar] [CrossRef]
- Tucker, C.J.; Pinzon, J.E.; Brown, M.E.; Slayback, D.A.; Pak, E.W.; Mahoney, R.; Vermote, E.F.; El Saleous, N. An extended AVHRR 8-km NDVI dataset compatible with MODIS and SPOT vegetation NDVI data. Int. J. Remote Sens. 2005, 26, 4485–4498. [Google Scholar] [CrossRef]
- ESRI. ArcGIS Pro: How Resampling Works; Environmental Systems Research Institute: Redlands, CA, USA, 2023. [Google Scholar]
- Kendall, M.G. Rank Correlation Methods, 4th ed.; Griffin: London, UK, 1975. [Google Scholar]
- Kassahun, M.Z.; Ture, K.B.; Nedaw, D.H. Spatio-temporal variability and trends of hydroclimatic variables at Zarima Sub-Basin North Western Ethiopia. Environ. Syst. Res. 2022, 11, 27. [Google Scholar]
- Shi, J.; Liu, M.; Li, Y.; Guan, C. Response of total primary productivity of vegetation to meteorological drought in arid and semi-arid regions of China. J. Arid Environ. 2025, 228, 105346. [Google Scholar] [CrossRef]
- Huang, C.; Yang, Q.; Guo, Y.; Zhang, Y.; Guo, L. The pattern, change and driven factors of vegetation cover in the Qin Mountains region. Sci. Rep. 2020, 10, 20591. [Google Scholar] [CrossRef]
- Sun, B.; Li, Z.; Gao, Z.; Guo, Z.; Wang, B.; Hu, X.; Bai, L. Grassland degradation and restoration monitoring and driving forces analysis based on long time-series remote sensing data in Xilin Gol League. Ecol. Front. 2017, 37, 219–228. [Google Scholar] [CrossRef]
- Li, C.H.; Zhou, M.; Wang, Y.T.; Zhu, T.B.; Sun, H.; Yin, H.H.; Gao, H.J.; Han, H.Y. Interannual variation of vegetation net primary productivity and its spatiotemporal and climatic contributions in arid Northwest China: A case study of Hexi Corridor. Chin. J. Ecol. 2020, 39, 3265–3275. [Google Scholar]
- Huang, K.; Xia, J. High ecosystem stability of evergreen broadleaf forests under severe droughts. Glob. Change Biol. 2019, 25, 3494–3503. [Google Scholar] [CrossRef]
- Jha, S.; Das, J.; Sharma, A.; Hazra, B.; Goyal, M.K. Probabilistic evaluation of vegetation drought likelihood and its implications to resilience across India. Glob. Planet. Change 2019, 176, 23–35. [Google Scholar] [CrossRef]
- Yang, L.; Yang, Y.Z. Spatiotemporal patterns and influencing factors of soil moisture in the West Liao River Basin based on TVDI. J. Arid Land Resour. Environ. 2016, 30, 76–81. [Google Scholar]
- Chen, Z.Y. Response of Vegetation Ecology to Groundwater Depth Changes in the West Liaohe Plain Based on RS/GIS. Ph.D. Thesis, Jilin University, Jilin, China, 2012. [Google Scholar]
- Zou, D.; Zhou, Y.K.; Dong, X.J.; Lin, J.; Wang, H.; Liang, J. Spatiotemporal linkages and attribution analysis of vegetation and meteorological drought in the Yangtze River Basin. Remote Sens. Technol. Appl. 2024, 39, 1183–1195. [Google Scholar]
- Wang, X.; Guo, L.; Hu, C.; Zhou, L.; Sun, X.; Zhao, J. Relationship between groundwater and vegetation development in the West Liao River Basin based on RS and GIS. Groundwater 2024, 46, 20–22. [Google Scholar]
- Han, L.; Cao, X.X.; Zhu, H.L.; Chen, R.; Zhao, Y.H.; Liu, Z.; Zhao, Z.L.; Chang, Y.Q.; Yang, M.L.; Gao, Y.F. Vegetation coverage changes and driving factors in the Loess Plateau based on characteristic zoning. Acta Ecol. Sin. 2023, 43, 8564–8577. [Google Scholar]
- Wang, Z.K.; Chen, W.; Piao, J.L.; Cai, Q.; Chen, S.; Xue, X.; Ma, T. Synergistic effects of high atmospheric and soil dryness on vegetation productivity decline in Southwest China. NPJ Clim. Atmos. Sci. 2025, 8, 6. [Google Scholar] [CrossRef]
- Yin, M.; Yin, Y.; Zong, X.; Deng, H. Global vegetation vulnerability to drought is underestimated due to the lagged effect. Agric. For. Meteorol. 2025, 364, 110451. [Google Scholar] [CrossRef]
- Dang, C.; Shao, Z.; Huang, X.; Qian, J.; Cheng, G.; Ding, Q.; Fan, Y. Assessment of the importance of increasing temperature and decreasing soil moisture on global ecosystem productivity using solar-induced chlorophyll fluorescence. Glob. Change Biol. 2022, 28, 2066–2080. [Google Scholar] [CrossRef]
- Deng, K.-Q.; Azorin-Molina, C.; Yang, S.; Hu, C.-D.; Zhang, G.-F.; Minola, L.; Vicente-Serrano, S.; Chen, D. Shifting of summertime weather extremes in Western Europe during 2012–2020. Adv. Clim. Change Res. 2022, 13, 218–227. [Google Scholar] [CrossRef]
- Haughey, E.; McElwain, J.C.; Finn, J.A. Annual yields of multispecies grassland mesocosms outperformed monocultures across a drought gradient due to complementarity effects and rapid recovery. Grassland Res. 2023, 2, 309–321. [Google Scholar] [CrossRef]
- Ma, Z.; Sun, P.; Yao, R. Spatiotemporal characteristics of drought and its impact on vegetation in Inner Mongolia. J. Soil Water Conserv. 2022, 36, 231–240. [Google Scholar]
- Yang, M. Temporal and Spatial Distribution Characteristics of Gross Primary Productivity in Guizhou Province and Its Response to Drought Variability. Master’s Thesis, Guizhou Normal University, Guizhoi, China, 2022. [Google Scholar]
- Yuan, B.; Guo, S.; Zhang, X.; Mu, H.; Cao, S.; Xia, Z.; Pan, X.; Du, P. Quantifying the drought sensitivity of vegetation types in northern China from 1982 to 2022. Agric. For. Meteorol. 2024, 359, 110293. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, Y.; Piao, S.; Wang, T.; Huang, K.; Wang, S.; Janssens, I.A. Drought memory weakens vegetation recovery capacity after recurrent droughts. Nat. Clim. Change 2023, 13, 754–760. [Google Scholar]
- Deng, H.; Yin, Y.; Han, X. Vulnerability of vegetation activities to drought in Central Asia. Environ. Res. Lett. 2020, 15, 084005. [Google Scholar] [CrossRef]
- Xiao, J.; Terrer, C.; Gentine, P.; Tateno, R.; Fan, L.; Ma, M.; Yue, Y.; Yuan, W.; Peñuelas, J.; Shi, W. Temporal and phenological modulation of the impact of increasing drought conditions on vegetation growth in a humid big river basin: Insights from global comparisons. Earth’s Future 2025, 13, e2024EF005720. [Google Scholar] [CrossRef]
- Cao, W.; Li, X.; Lyu, X.; Dang, D.; Wang, K.; Li, M.; Liu, S. To explore the effectiveness of various ecological security pattern construction methods in many growth situations in the future: A case study of the West Liaohe River Basin in Inner Mongolia. Sci. Total Environ. 2024, 948, 174607. [Google Scholar] [CrossRef]
- Zheng, W.; Askari, K.; Song, C.; Shi, P.; Ge, W.; Shi, S.; Chu, J.; Chen, H.; Wang, F. Increasing vulnerability of vegetation to meteorological and groundwater drought: A case study in Argentina. J. Hydrol. Reg. Stud. 2024, 55, 101931. [Google Scholar] [CrossRef]
- Shi, Y.; Zhang, B.; Liang, L.; Wang, S.; Zhang, H.; Sun, H.; Han, X. Unfolding the effectiveness of ecological restoration programs in enhancing vegetation carbon sinks across different climate zones in China. Resour. Conserv. Recycl. 2025, 212, 107974. [Google Scholar] [CrossRef]
- Li, X.; Piao, S.; Wang, K.; Wang, X.; Wang, T.; Ciais, P.; Chen, A.; Lian, X.; Peng, S.; Peñuelas, J. Temporal trade-off between gymnosperm resistance and resilience increases forest sensitivity to extreme drought. Nat. Ecol. Evol. 2022, 4, 1075–1083. [Google Scholar] [CrossRef]
- Prasad, J.; Akila, N.; Bharathi, C.S.; Alagudurai, S.; Rao, C.R.; Raju, B.; Rao, K.; Rao, A.S.; Rejani, R.; Ramana, D.; et al. Assessment of adaptation practices for risk minimization to drought in semi-arid environments. Clim. Risk Manag. 2023, 42, 100563. [Google Scholar] [CrossRef]
- Hoover, D.L.; Knapp, A.K.; Smith, M.D. Drought resistance and resilience in grassland ecosystems: The role of plant functional traits. Agronomy 2022, 12, 701. [Google Scholar]
- Lobell, D.B.; Deines, J.M.; Di Tommaso, S. Divergent crop responses to drought in a warming world. Proc. Natl. Acad. Sci. USA 2023, 120, e2208976120. [Google Scholar]
- Elagib, A.N.; Ali, A.M.M.; Musa, A.A. Intensifying drought render more Sahel drylands unsuitable for cultivation. Sci. Total Environ. 2024, 954, 176390. [Google Scholar] [CrossRef]
- Taschetto, S.A.; Stojanovic, M.; Holgate, M.C.; Drumond, A.; Evans, J.P.; Gimeno, L.; Nieto, R. Changes in moisture sources contributed to the onset and development of the 2017-2019 southeast Australian drought. Weather Clim. Extrem. 2024, 44, 100672. [Google Scholar] [CrossRef]
- Essa, Y.H.; Hirschi, M.; Thiery, W.; El-Kenawy, A.M.; Yang, C. Drought characteristics in Mediterranean under future climate change. npj Clim. Atmos. Sci. 2023, 6, 133. [Google Scholar] [CrossRef]
- Yuan, Y.; Bao, A.; Jiang, P.; Hamdi, R.; Termonia, P.; De Maeyer, P.; Guo, H.; Zheng, G.; Yu, T.; Prishchepov, A.V. Probabilistic assessment of vegetation vulnerability to drought stress in Central Asia. J. Environ. Manag. 2022, 310, 114504. [Google Scholar] [CrossRef]
- Porcelli, A.C.; Rubio, G.; Boem, G.H.F.; Lavado, R.S. The Effect of Water and Salt Stress on Paspalum dilatatum, a Constituent of Pampas Natural Grasslands. Phyton 2024, 93, 2009–2018. [Google Scholar] [CrossRef]
- Pettorelli, N.; Bühne, H.S.T.; Tulloch, A.; Dubois, G.; Macinnis-Ng, C.; Queirós, A.M.; Keith, D.A.; Wegmann, M.; Schrodt, F.; Stellmes, M.; et al. Satellite remote sensing of ecosystem functions: Opportunities, challenges, and way forward. Remote Sens. Ecol. Conserv. 2018, 4, 71–93. [Google Scholar] [CrossRef]
- Allen, C.D.; Breshears, D.D.; McDowell, N.G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 2020, 6, 1–55. [Google Scholar] [CrossRef]
- AghaKouchak, A.; Chiang, F.; Huning, L.S.; Love, C.A.; Mallakpour, I.; Mazdiyasni, O.; Moftakhari, H.; Papalexiou, S.M.; Ragno, E.; Sadegh, M. Climate extremes and compound hazards in a warming world. Annu. Rev. Earth Planet. Sci. 2020, 48, 519–548. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drought Severity | SPEI Values |
|---|---|
| Near normal | (−0.5, + |
| Light drought | (−1.0, −0.5 |
| Moderate drought | (−1.5, −1.0 |
| Severe drought | (−2.0, −1.5 |
| Extreme drought | (, −2.0 |
| Annual | Spring | Summer | Autumn | Winter | |
|---|---|---|---|---|---|
| Sen slope (/a) | 0.00179 | 0.00998 * | 0.00420 * | 0.00208 * | 0.00022 |
| MK test | 1.69969 | 2.09913 | 2.51896 | 2.02916 | 0.34985 |
| Annual | Spring | Summer | Autumn | Winter | |
|---|---|---|---|---|---|
| Sen slope (/a) | 0.02742 | 0.01344 | 0.04575 | 0.07995 * | −0.00591 |
| MK test | 1.53936 | 0.20991 | 1.32945 | 2.37902 | −0.06997 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, Y.; Zheng, H.; Yan, M.; Wu, L. Research on the Response Mechanism of Vegetation to Drought Stress in the West Liao River Basin, China. Remote Sens. 2025, 17, 1780. https://doi.org/10.3390/rs17101780
Tian Y, Zheng H, Yan M, Wu L. Research on the Response Mechanism of Vegetation to Drought Stress in the West Liao River Basin, China. Remote Sensing. 2025; 17(10):1780. https://doi.org/10.3390/rs17101780
Chicago/Turabian StyleTian, Yuhong, Huichao Zheng, Mengxuan Yan, and Lizhu Wu. 2025. "Research on the Response Mechanism of Vegetation to Drought Stress in the West Liao River Basin, China" Remote Sensing 17, no. 10: 1780. https://doi.org/10.3390/rs17101780
APA StyleTian, Y., Zheng, H., Yan, M., & Wu, L. (2025). Research on the Response Mechanism of Vegetation to Drought Stress in the West Liao River Basin, China. Remote Sensing, 17(10), 1780. https://doi.org/10.3390/rs17101780