

Using Multi-Angular Spectral Reflection of Dorsiventral Leaves to Improve the Transferability of PLSR Models for Estimating Leaf Biochemical Traits



Abstract
1. Introduction
2. Materials and Methods
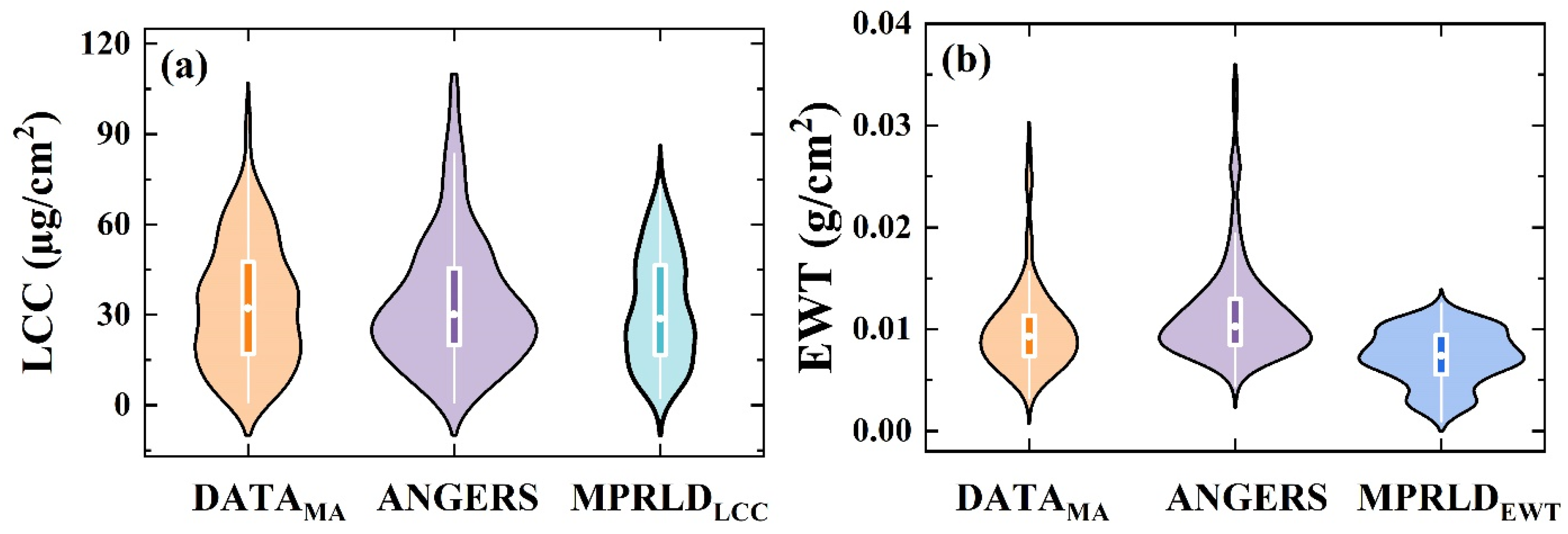
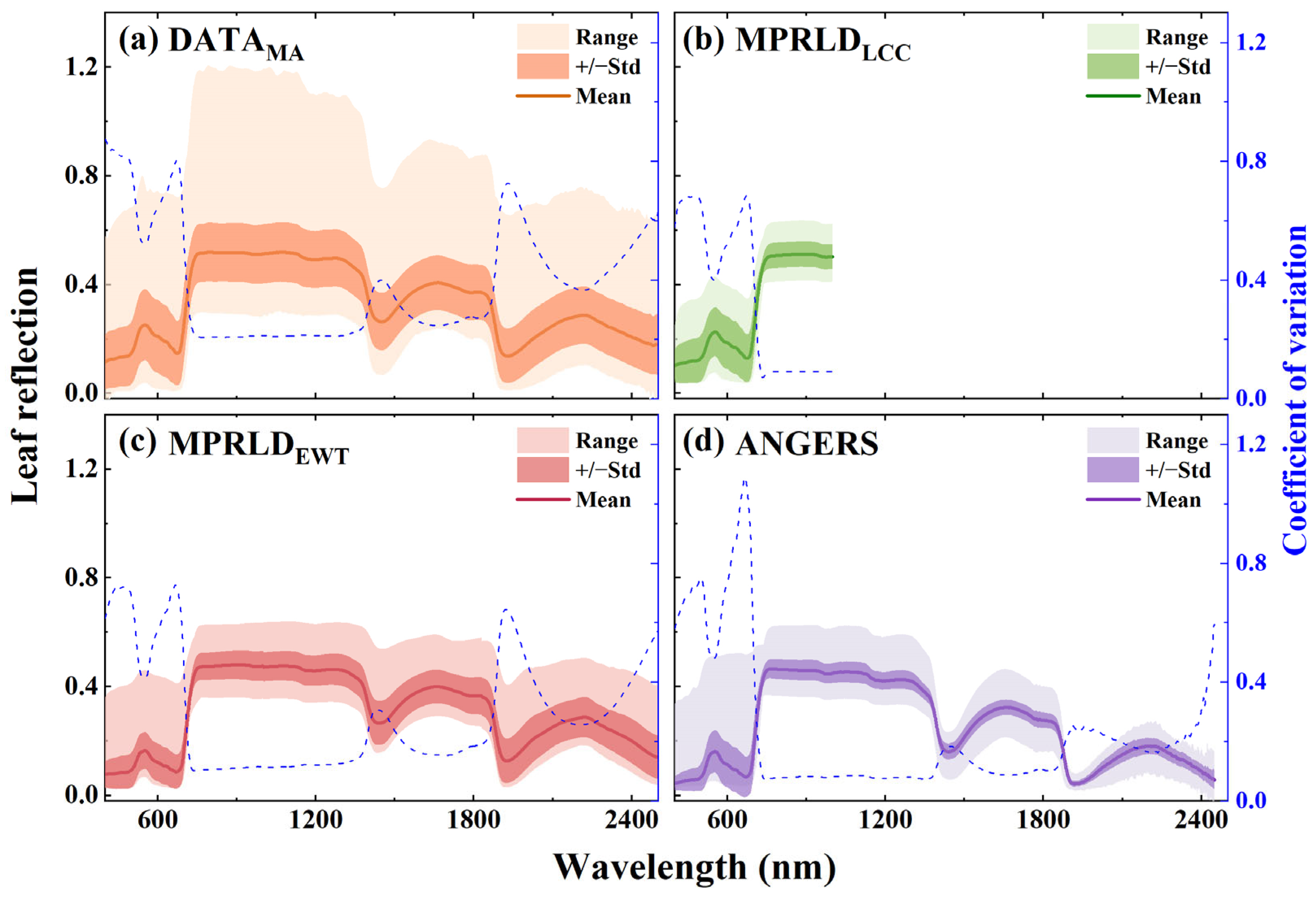
2.1. Description of Datasets
2.2. Estimation of Leaf Biochemical Traits Using PLSR
- (1)
- Strategy 1: training with adaxial reflection spectra measured at the nadir.
- (2)
- Strategy 2: training with abaxial reflection spectra measured at the nadir.
- (3)
- Strategy 3: training with both adaxial and abaxial reflection spectra measured at the nadir.
- (4)
- Strategy 4: training with multi-angular reflection spectra from both the adaxial and abaxial sides.
2.3. Statistical Analysis
3. Results
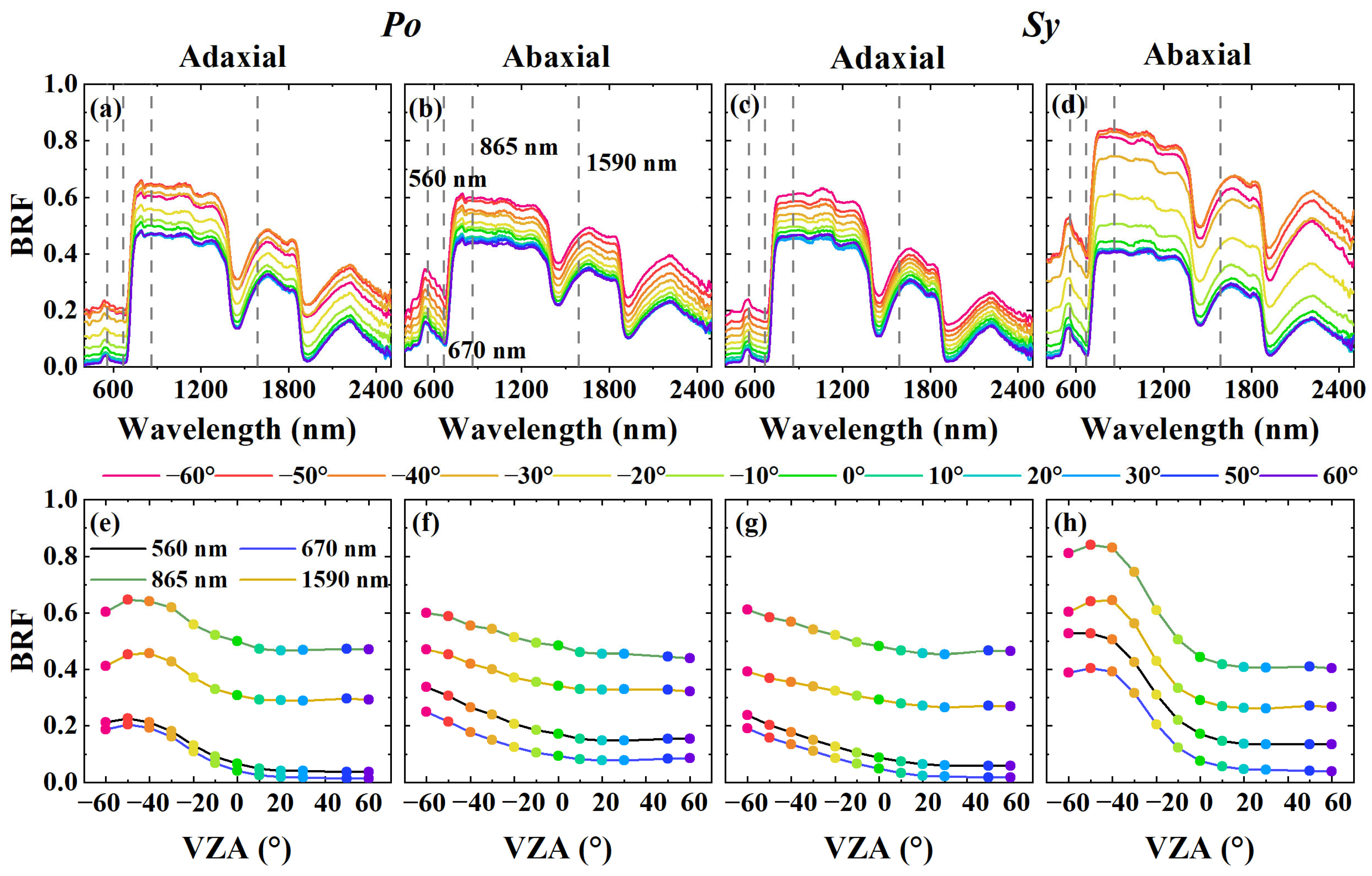
3.1. Spectral Variations Across Leaf Sides, VZAs, and Datasets
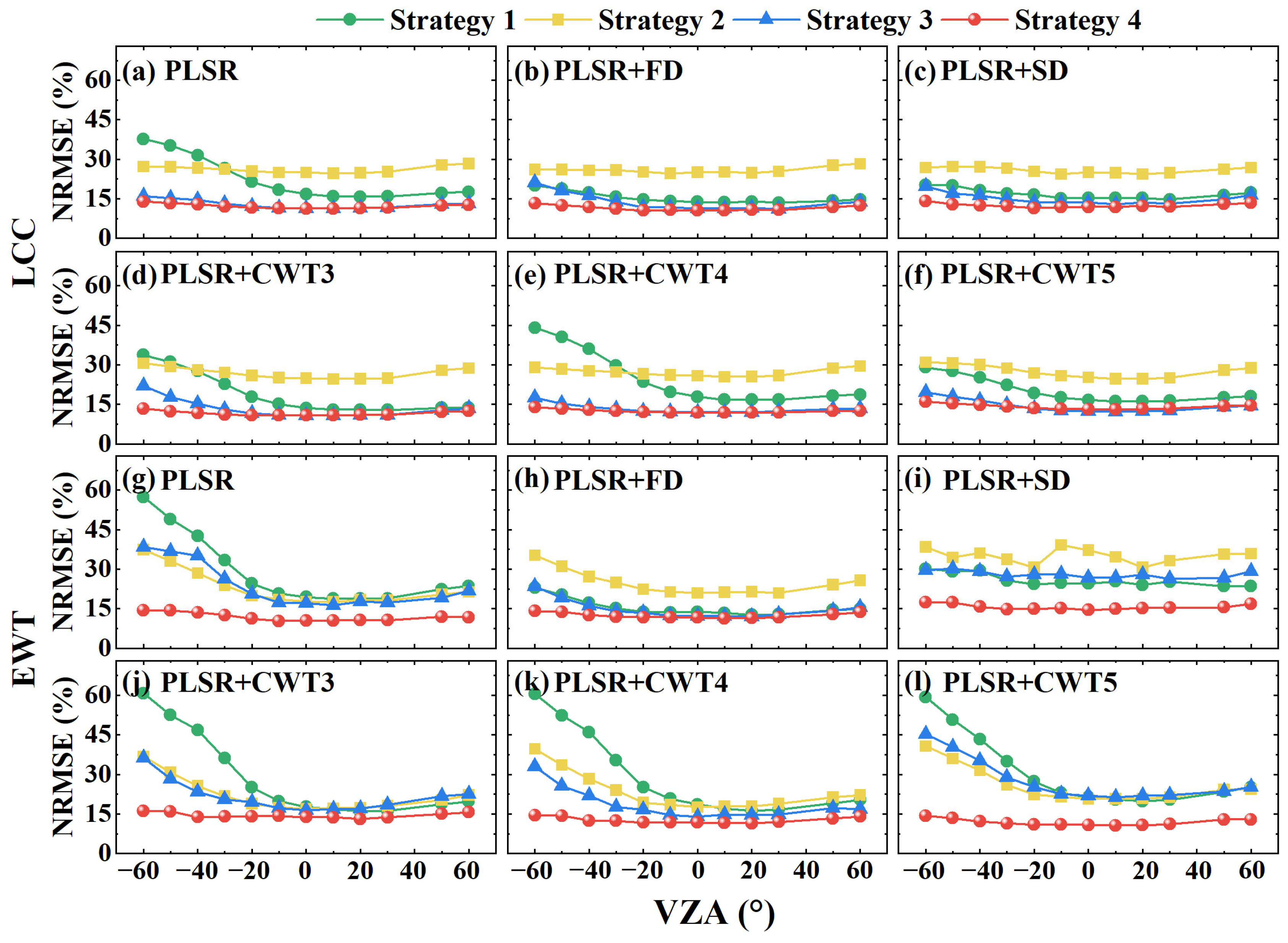
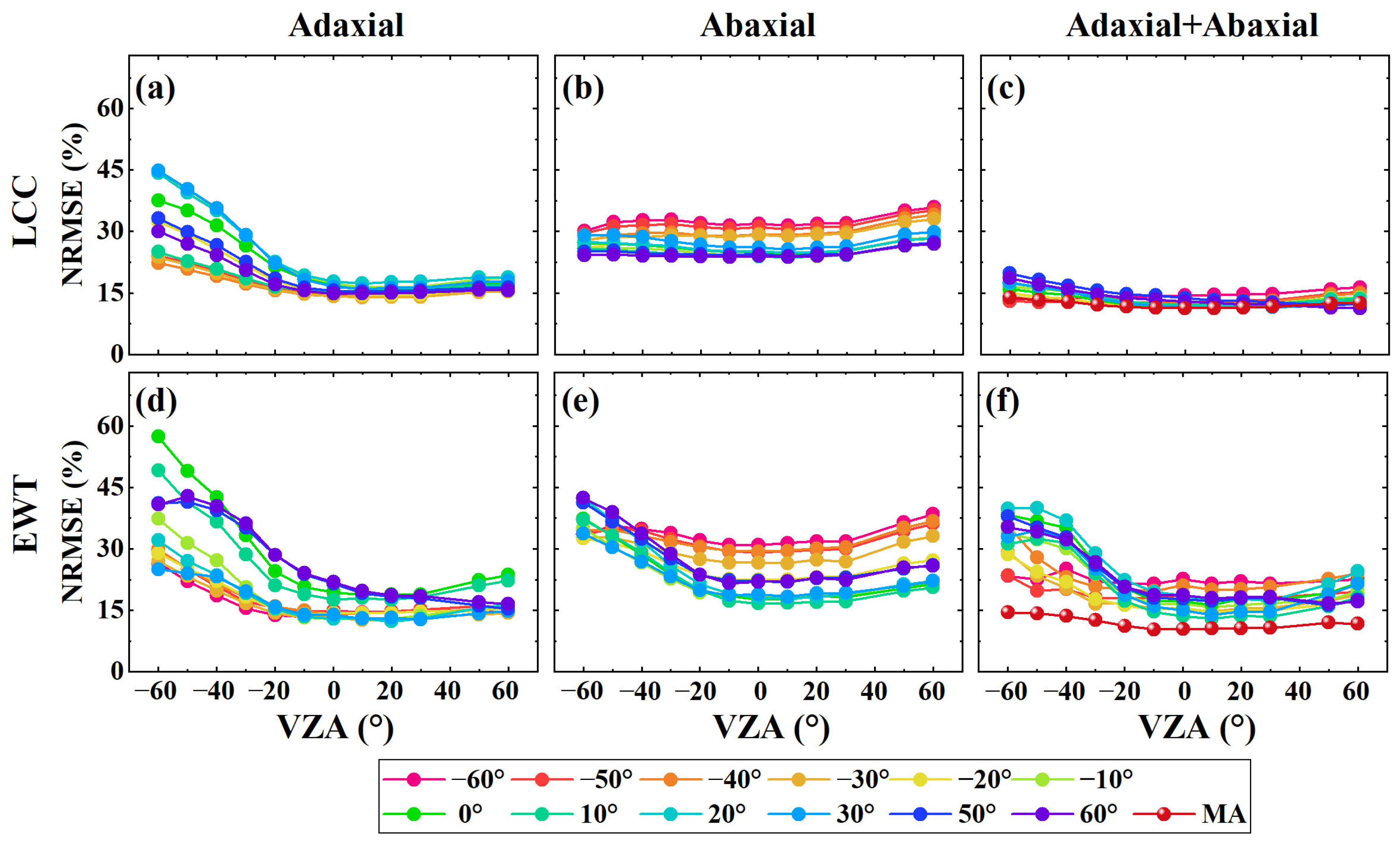
3.2. The Transferability of the PLSR Model to Different VZAs
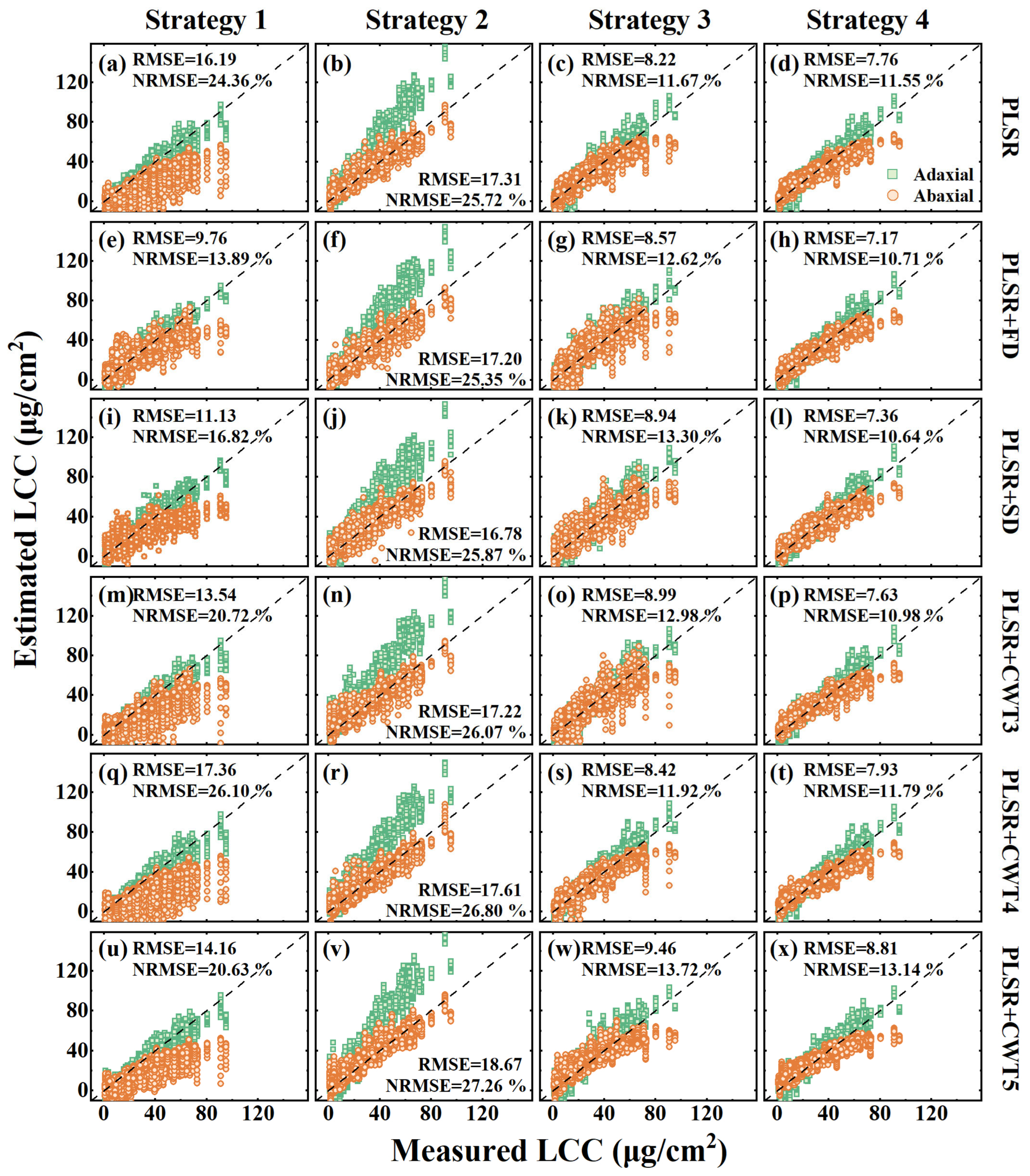
3.3. The Transferability of the PLSR Model to Different Leaf Sides
3.4. The Transferability of the PLSR Model to Different Datasets
4. Discussion
4.1. The Importance of Considering Leaf Reflection Spectral Variations Across Leaf Sides and VZAs for PLSR Modeling
4.2. Implications of Using Multi-Angular Reflection Spectra of Dorsiventral Leaves to Improve PLSR Model Transferability
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Féret, J.-B.; François, C.; Gitelson, A.; Asner, G.P.; Barry, K.M.; Panigada, C.; Richardson, A.D.; Jacquemoud, S. Optimizing spectral indices and chemometric analysis of leaf chemical properties using radiative transfer modeling. Remote Sens. Environ. 2011, 115, 2742–2750. [Google Scholar] [CrossRef]
- Kattge, J.; Diaz, S.; Lavorel, S.; Prentice, I.C.; Leadley, P.; Bönisch, G.; Garnier, E.; Westoby, M.; Reich, P.B.; Wright, I.J. TRY–A global database of plant traits. Glob. Change Biol. 2011, 17, 2905–2935. [Google Scholar] [CrossRef]
- Ollinger, S.V. Sources of variability in canopy reflectance and the convergent properties of plants. New Phytol. 2011, 189, 375–394. [Google Scholar] [CrossRef]
- Schimel, D.; Pavlick, R.; Fisher, J.B.; Asner, G.P.; Saatchi, S.; Townsend, P.; Miller, C.; Frankenberg, C.; Hibbard, K.; Cox, P. Observing terrestrial ecosystems and the carbon cycle from space. Glob. Change Biol. 2015, 21, 1762–1776. [Google Scholar] [CrossRef]
- Schweiger, A.K.; Cavender-Bares, J.; Townsend, P.A.; Hobbie, S.E.; Madritch, M.D.; Wang, R.; Tilman, D.; Gamon, J.A. Plant spectral diversity integrates functional and phylogenetic components of biodiversity and predicts ecosystem function. Nat. Ecol. Evol. 2018, 2, 976–982. [Google Scholar] [CrossRef] [PubMed]
- Croft, H.; Chen, J.M.; Luo, X.; Bartlett, P.; Chen, B.; Staebler, R.M. Leaf chlorophyll content as a proxy for leaf photosynthetic capacity. Glob. Change Biol. 2017, 23, 3513–3524. [Google Scholar] [CrossRef]
- Gamon, J.A.; Huemmrich, K.F.; Wong, C.Y.; Ensminger, I.; Garrity, S.; Hollinger, D.Y.; Noormets, A.; Penuelas, J. A remotely sensed pigment index reveals photosynthetic phenology in evergreen conifers. Proc. Natl. Acad. Sci. USA 2016, 113, 13087–13092. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Gritz, Y.; Merzlyak, M.N. Relationships between leaf chlorophyll content and spectral reflectance and algorithms for non-destructive chlorophyll assessment in higher plant leaves. J. Plant Physiol. 2003, 160, 271–282. [Google Scholar] [CrossRef]
- Houborg, R.; McCabe, M.F.; Cescatti, A.; Gitelson, A.A. Leaf chlorophyll constraint on model simulated gross primary productivity in agricultural systems. Int. J. Appl. Earth Obs. Geoinf. 2015, 43, 160–176. [Google Scholar] [CrossRef]
- Van Bodegom, P.M.; Douma, J.C.; Verheijen, L.M. A fully traits-based approach to modeling global vegetation distribution. Proc. Natl. Acad. Sci. USA 2014, 111, 13733–13738. [Google Scholar] [CrossRef]
- Galmés, J.; Medrano, H.; Flexas, J. Photosynthetic limitations in response to water stress and recovery in Mediterranean plants with different growth forms. New Phytol. 2007, 175, 81–93. [Google Scholar] [CrossRef]
- Sims, D.A.; Gamon, J.A. Estimation of vegetation water content and photosynthetic tissue area from spectral reflectance: A comparison of indices based on liquid water and chlorophyll absorption features. Remote Sens. Environ. 2003, 84, 526–537. [Google Scholar] [CrossRef]
- Humphrey, V.; Zscheischler, J.; Ciais, P.; Gudmundsson, L.; Sitch, S.; Seneviratne, S.I. Sensitivity of atmospheric CO2 growth rate to observed changes in terrestrial water storage. Nature 2018, 560, 628–631. [Google Scholar] [CrossRef]
- Maki, M.; Ishiahra, M.; Tamura, M. Estimation of leaf water status to monitor the risk of forest fires by using remotely sensed data. Remote Sens. Environ. 2004, 90, 441–450. [Google Scholar] [CrossRef]
- Houborg, R.; Fisher, J.B.; Skidmore, A.K. Advances in remote sensing of vegetation function and traits. Int. J. Appl. Earth Obs. Geoinf. 2015, 43, 1–6. [Google Scholar] [CrossRef]
- Luo, X.; Croft, H.; Chen, J.M.; He, L.; Keenan, T.F. Improved estimates of global terrestrial photosynthesis using information on leaf chlorophyll content. Glob. Change Biol. 2019, 25, 2499–2514. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Gamon, J.A. Remote sensing of terrestrial plant biodiversity. Remote Sens. Environ. 2019, 231, 111218. [Google Scholar] [CrossRef]
- Asner, G.P.; Martin, R.E. Spectral and chemical analysis of tropical forests: Scaling from leaf to canopy levels. Remote Sens. Environ. 2008, 112, 3958–3970. [Google Scholar] [CrossRef]
- Carter, G.A.; Knapp, A.K. Leaf optical properties in higher plants: Linking spectral characteristics to stress and chlorophyll concentration. Am. J. Bot. 2001, 88, 677–684. [Google Scholar] [CrossRef]
- Hill, J.; Buddenbaum, H.; Townsend, P.A. Imaging spectroscopy of forest ecosystems: Perspectives for the use of space-borne hyperspectral earth observation systems. Surv. Geophys. 2019, 40, 553–588. [Google Scholar] [CrossRef]
- Curran, P.J.; Dungan, J.L.; Peterson, D.L. Estimating the foliar biochemical concentration of leaves with reflectance spectrometry: Testing the Kokaly and Clark methodologies. Remote Sens. Environ. 2001, 76, 349–359. [Google Scholar] [CrossRef]
- Datt, B. A new reflectance index for remote sensing of chlorophyll content in higher plants: Tests using Eucalyptus leaves. J. Plant Physiol. 1999, 154, 30–36. [Google Scholar] [CrossRef]
- Gitelson, A.; Merzlyak, M.N. Quantitative estimation of chlorophyll-a using reflectance spectra: Experiments with autumn chestnut and maple leaves. J. Photochem. Photobiol. B Biol. 1994, 22, 247–252. [Google Scholar] [CrossRef]
- Sims, D.A.; Gamon, J.A. Relationships between leaf pigment content and spectral reflectance across a wide range of species, leaf structures and developmental stages. Remote Sens. Environ. 2002, 81, 337–354. [Google Scholar] [CrossRef]
- Ceccato, P.; Flasse, S.; Tarantola, S.; Jacquemoud, S.; Grégoire, J.-M. Detecting vegetation leaf water content using reflectance in the optical domain. Remote Sens. Environ. 2001, 77, 22–33. [Google Scholar] [CrossRef]
- Gao, B.C. NDWI—A normalized difference water index for remote sensing of vegetation liquid water from space. Remote Sens. Environ. 1996, 58, 257–266. [Google Scholar] [CrossRef]
- Peñuelas, J.; Pinol, J.; Ogaya, R.; Filella, I. Estimation of plant water concentration by the reflectance water index WI (R900/R970). Int. J. Remote Sens. 1997, 18, 2869–2875. [Google Scholar] [CrossRef]
- Asner, G.P.; Martin, R.E.; Anderson, C.B.; Knapp, D.E. Quantifying forest canopy traits: Imaging spectroscopy versus field survey. Remote Sens. Environ. 2015, 158, 15–27. [Google Scholar] [CrossRef]
- Féret, J.B.; le Maire, G.; Jay, S.; Berveiller, D.; Bendoula, R.; Hmimina, G.; Cheraiet, A.; Oliveira, J.C.; Ponzoni, F.J.; Solanki, T.; et al. Estimating leaf mass per area and equivalent water thickness based on leaf optical properties: Potential and limitations of physical modeling and machine learning. Remote Sens. Environ. 2019, 231, 110959. [Google Scholar] [CrossRef]
- Ji, F.; Li, F.; Hao, D.; Shiklomanov, A.N.; Yang, X.; Townsend, P.A.; Dashti, H.; Nakaji, T.; Kovach, K.R.; Liu, H.; et al. Unveiling the transferability of PLSR models for leaf trait estimation: Lessons from a comprehensive analysis with a novel global dataset. New Phytol. 2024, 243, 111–131. [Google Scholar] [CrossRef]
- Verrelst, J.; Malenovský, Z.; Van der Tol, C.; Camps-Valls, G.; Gastellu-Etchegorry, J.-P.; Lewis, P.; North, P.; Moreno, J. Quantifying vegetation biophysical variables from imaging spectroscopy data: A review on retrieval methods. Surv. Geophys. 2019, 40, 589–629. [Google Scholar] [CrossRef] [PubMed]
- Féret, J.-B.; Berger, K.; de Boissieu, F.; Malenovský, Z. PROSPECT-PRO for estimating content of nitrogen-containing leaf proteins and other carbon-based constituents. Remote Sens. Environ. 2021, 252, 112173. [Google Scholar] [CrossRef]
- Feret, J.-B.; François, C.; Asner, G.P.; Gitelson, A.A.; Martin, R.E.; Bidel, L.P.R.; Ustin, S.L.; le Maire, G.; Jacquemoud, S. PROSPECT-4 and 5: Advances in the leaf optical properties model separating photosynthetic pigments. Remote Sens. Environ. 2008, 112, 3030–3043. [Google Scholar] [CrossRef]
- Jacquemoud, S.; Baret, F. PROSPECT: A model of leaf optical properties spectra. Remote Sens. Environ. 1990, 34, 75–91. [Google Scholar] [CrossRef]
- Stuckens, J.; Verstraeten, W.W.; Delalieux, S.; Swennen, R.; Coppin, P. A dorsiventral leaf radiative transfer model: Development, validation and improved model inversion techniques. Remote Sens. Environ. 2009, 113, 2560–2573. [Google Scholar] [CrossRef]
- Jiang, J.; Comar, A.; Weiss, M.; Baret, F. FASPECT: A model of leaf optical properties accounting for the differences between upper and lower faces. Remote Sens. Environ. 2021, 253. [Google Scholar] [CrossRef]
- Combal, B.; Baret, F.; Weiss, M.; Trubuil, A.; Mace, D.; Pragnere, A.; Myneni, R.; Knyazikhin, Y.; Wang, L. Retrieval of canopy biophysical variables from bidirectional reflectance: Using prior information to solve the ill-posed inverse problem. Remote Sens. Environ. 2003, 84, 1–15. [Google Scholar] [CrossRef]
- Berger, K.; Atzberger, C.; Danner, M.; D’Urso, G.; Mauser, W.; Vuolo, F.; Hank, T. Evaluation of the PROSAIL model capabilities for future hyperspectral model environments: A review study. Remote Sens. 2018, 10, 85. [Google Scholar] [CrossRef]
- Jacquemoud, S.; Baret, F.; Andrieu, B.; Danson, F.M.; Jaggard, K. Extraction of vegetation biophysical parameters by inversion of the PROSPECT + SAIL models on sugar beet canopy reflectance data. Application to TM and AVIRIS sensors. Remote Sens. Environ. 1995, 52, 163–172. [Google Scholar] [CrossRef]
- Kimes, D.S.; Knyazikhin, Y.; Privette, J.L.; Abuelgasim, A.A.; Gao, F. Inversion methods for physically-based models. Remote Sens. Rev. 2000, 18, 381–439. [Google Scholar] [CrossRef]
- Weiss, M.; Baret, F. Evaluation of canopy biophysical variable retrieval performances from the accumulation of large swath satellite data. Remote Sens. Environ. 1999, 70, 293–306. [Google Scholar] [CrossRef]
- Verrelst, J.; Camps-Valls, G.; Muñoz-Marí, J.; Rivera, J.P.; Veroustraete, F.; Clevers, J.G.P.W.; Moreno, J. Optical remote sensing and the retrieval of terrestrial vegetation bio-geophysical properties—A review. ISPRS J. Photogramm. Remote Sens. 2015, 108, 273–290. [Google Scholar] [CrossRef]
- Cao, Z.X.; Wang, Q.; Zheng, C.L. Best hyperspectral indices for tracing leaf water status as determined from leaf dehydration experiments. Ecol. Indic. 2015, 54, 96–107. [Google Scholar] [CrossRef]
- Main, R.; Cho, M.A.; Mathieu, R.; O’Kennedy, M.M.; Ramoelo, A.; Koch, S. An investigation into robust spectral indices for leaf chlorophyll estimation. ISPRS J. Photogramm. Remote Sens. 2011, 66, 751–761. [Google Scholar] [CrossRef]
- Sun, J.; Yang, J.; Shi, S.; Chen, B.; Du, L.; Gong, W.; Song, S. Estimating rice leaf nitrogen concentration: Influence of regression algorithms based on passive and active leaf reflectance. Remote Sens. 2017, 9, 951. [Google Scholar] [CrossRef]
- Verrelst, J.; Rivera, J.P.; Gitelson, A.; Delegido, J.; Moreno, J.; Camps-Valls, G. Spectral band selection for vegetation properties retrieval using Gaussian processes regression. Int. J. Appl. Earth Obs. Geoinf. 2016, 52, 554–567. [Google Scholar] [CrossRef]
- Wan, L.; Zhou, W.; He, Y.; Wanger, T.C.; Cen, H. Combining transfer learning and hyperspectral reflectance analysis to assess leaf nitrogen concentration across different plant species datasets. Remote Sens. Environ. 2022, 269, 112826. [Google Scholar] [CrossRef]
- García-Soria, J.L.; Morata, M.; Berger, K.; Pascual-Venteo, A.B.; Rivera-Caicedo, J.P.; Verrelst, J. Evaluating epistemic uncertainty estimation strategies in vegetation trait retrieval using hybrid models and imaging spectroscopy data. Remote Sens. Environ. 2024, 310, 114228. [Google Scholar] [CrossRef]
- Cherif, E.; Feilhauer, H.; Berger, K.; Dao, P.D.; Ewald, M.; Hank, T.B.; He, Y.; Kovach, K.R.; Lu, B.; Townsend, P.A.; et al. From spectra to plant functional traits: Transferable multi-trait models from heterogeneous and sparse data. Remote Sens. Environ. 2023, 292, 113580. [Google Scholar] [CrossRef]
- Hwang, J.-S.; and Hu, T.-H. A stepwise regression algorithm for high-dimensional variable selection. J. Stat. Comput. Simul. 2015, 85, 1793–1806. [Google Scholar] [CrossRef]
- Atzberger, C.; Guérif, M.; Baret, F.; Werner, W. Comparative analysis of three chemometric techniques for the spectroradiometric assessment of canopy chlorophyll content in winter wheat. Comput. Electron. Agric. 2010, 73, 165–173. [Google Scholar] [CrossRef]
- Rudin, C. Stop explaining black box machine learning models for high stakes decisions and use interpretable models instead. Nat. Mach. Intell. 2019, 1, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Guzman, Q.J.A.; Sanchez-Azofeifa, G.A. Prediction of leaf traits of lianas and trees via the integration of wavelet spectra in the visible-near infrared and thermal infrared domains. Remote Sens. Environ. 2021, 259, 112406. [Google Scholar] [CrossRef]
- Kothari, S.; Beauchamp-Rioux, R.; Blanchard, F.; Crofts, A.L.; Girard, A.; Guilbeault-Mayers, X.; Hacker, P.W.; Pardo, J.; Schweiger, A.K.; Demers-Thibeault, S.; et al. Predicting leaf traits across functional groups using reflectance spectroscopy. New Phytol. 2023, 238, 549–566. [Google Scholar] [CrossRef]
- Nakaji, T.; Oguma, H.; Nakamura, M.; Kachina, P.; Asanok, L.; Marod, D.; Aiba, M.; Kurokawa, H.; Kosugi, Y.; Kassim, A.R.; et al. Estimation of six leaf traits of East Asian forest tree species by leaf spectroscopy and partial least square regression. Remote Sens. Environ. 2019, 233, 111381. [Google Scholar] [CrossRef]
- Neuwirthová, E.; Lhotáková, Z.; Červená, L.; Lukeš, P.; Campbell, P.; Albrechtová, J. Asymmetry of leaf internal structure affects PLSR modelling of anatomical traits using VIS-NIR leaf level spectra. Eur. J. Remote Sens. 2024, 57, 2292154. [Google Scholar] [CrossRef]
- Yan, Z.; Guo, Z.; Serbin, S.P.; Song, G.; Zhao, Y.; Chen, Y.; Wu, S.; Wang, J.; Wang, X.; Li, J.; et al. Spectroscopy outperforms leaf trait relationships for predicting photosynthetic capacity across different forest types. New Phytol. 2021, 232, 134–147. [Google Scholar] [CrossRef]
- Wang, Z.; Féret, J.-B.; Liu, N.; Sun, Z.; Yang, L.; Geng, S.; Zhang, H.; Chlus, A.; Kruger, E.L.; Townsend, P.A. Generality of leaf spectroscopic models for predicting key foliar functional traits across continents: A comparison between physically-and empirically-based approaches. Remote Sens. Environ. 2023, 293, 113614. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, Y.; Nunes, M.H.; Stoddart, J.; Khoury, S.; Chan, A.H.Y.; Coomes, D.A. Predicting leaf traits of temperate broadleaf deciduous trees from hyperspectral reflectance: Can a general model be applied across a growing season? Remote Sens. Environ. 2022, 269, 112767. [Google Scholar] [CrossRef]
- Wu, J.; Chavana-Bryant, C.; Prohaska, N.; Serbin, S.P.; Guan, K.; Albert, L.P.; Yang, X.; van Leeuwen, W.J.D.; Garnello, A.J.; Martins, G.; et al. Convergence in relationships between leaf traits, spectra and age across diverse canopy environments and two contrasting tropical forests. New Phytol. 2017, 214, 1033–1048. [Google Scholar] [CrossRef]
- Wang, S.; Guan, K.; Wang, Z.; Ainsworth, E.A.; Zheng, T.; Townsend, P.A.; Li, K.; Moller, C.; Wu, G.; Jiang, C. Unique contributions of chlorophyll and nitrogen to predict crop photosynthetic capacity from leaf spectroscopy. J. Exp. Bot. 2020, 72, 341–354. [Google Scholar] [CrossRef]
- Yang, X.; Tang, J.; Mustard, J.F.; Wu, J.; Zhao, K.; Serbin, S.; Lee, J.-E. Seasonal variability of multiple leaf traits captured by leaf spectroscopy at two temperate deciduous forests. Remote Sens. Environ. 2016, 179, 1–12. [Google Scholar] [CrossRef]
- Borsuk, A.M.; Roddy, A.B.; Théroux-Rancourt, G.; Brodersen, C.R. Structural organization of the spongy mesophyll. New Phytol. 2022, 234, 946–960. [Google Scholar] [CrossRef] [PubMed]
- Wall, S.; Vialet-Chabrand, S.; Davey, P.; Van Rie, J.; Galle, A.; Cockram, J.; Lawson, T. Stomata on the abaxial and adaxial leaf surfaces contribute differently to leaf gas exchange and photosynthesis in wheat. New Phytol. 2022, 235, 1743–1756. [Google Scholar] [CrossRef]
- Nakata, M.; Okada, K. The leaf adaxial-abaxial boundary and lamina growth. Plants 2013, 2, 174–202. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Nukazuka, A.; Tsukaya, H. Leaf adaxial–abaxial polarity specification and lamina outgrowth: Evolution and development. Plant Cell Physiol. 2012, 53, 1180–1194. [Google Scholar] [CrossRef]
- Drake, P.L.; de Boer, H.J.; Schymanski, S.J.; Veneklaas, E.J. Two sides to every leaf: Water and CO2 transport in hypostomatous and amphistomatous leaves. New Phytol. 2019, 222, 1179–1187. [Google Scholar] [CrossRef]
- Wieneke, S.; Pacheco-Labrador, J.; Mahecha, M.D.; Poblador, S.; Vicca, S.; Janssens, I.A. Comparing the quantum use efficiency of red and far-red sun-induced fluorescence at leaf and canopy under heat-drought stress. Remote Sens. Environ. 2024, 311, 114294. [Google Scholar] [CrossRef]
- Sagan, V.; Maimaitijiang, M.; Paheding, S.; Bhadra, S.; Gosselin, N.; Burnette, M.; Demieville, J.; Hartling, S.; LeBauer, D.; Newcomb, M.; et al. Data-driven artificial intelligence for calibration of hyperspectral big data. IEEE Trans. Geosci. Remote Sens. 2022, 60, 5510320. [Google Scholar] [CrossRef]
- Li, W.; Jiang, J.; Weiss, M.; Madec, S.; Tison, F.; Philippe, B.; Comar, A.; Baret, F. Impact of the reproductive organs on crop BRDF as observed from a UAV. Remote Sens. Environ. 2021, 259, 112433. [Google Scholar] [CrossRef]
- Schläpfer, D.; Richter, R.; Feingersh, T. Operational BRDF effects correction for wide-field-of-view optical scanners (BREFCOR). IEEE Trans. Geosci. Remote Sens. 2015, 53, 1855–1864. [Google Scholar] [CrossRef]
- Zhao, F.; Li, Y.; Dai, X.; Verhoef, W.; Guo, Y.; Shang, H.; Gu, X.; Huang, Y.; Yu, T.; Huang, J. Simulated impact of sensor field of view and distance on field measurements of bidirectional reflectance factors for row crops. Remote Sens. Environ. 2015, 156, 129–142. [Google Scholar] [CrossRef]
- Lukeš, P.; Neuwirthová, E.; Lhotáková, Z.; Janoutová, R.; Albrechtová, J. Upscaling seasonal phenological course of leaf dorsiventral reflectance in radiative transfer model. Remote Sens. Environ. 2020, 246, 111862. [Google Scholar] [CrossRef]
- Bousquet, L.; Lachérade, S.; Jacquemoud, S.; Moya, I. Leaf BRDF measurements and model for specular and diffuse components differentiation. Remote Sens. Environ. 2005, 98, 201–211. [Google Scholar] [CrossRef]
- Comar, A.; Baret, F.; Vienot, F.; Yan, L.; De Solan, B. Wheat leaf bidirectional reflectance measurements: Description and quantification of the volume, specular and hot-spot scattering features. Remote Sens. Environ. 2012, 121, 26–35. [Google Scholar] [CrossRef]
- Jay, S.; Bendoula, R.; Hadoux, X.; Feret, J.B.; Gorretta, N. A physically-based model for retrieving foliar biochemistry and leaf orientation using close-range imaging spectroscopy. Remote Sens. Environ. 2016, 177, 220–236. [Google Scholar] [CrossRef]
- Li, D.; Tian, L.; Wan, Z.; Jia, M.; Yao, X.; Tian, Y.; Zhu, Y.; Cao, W.; Cheng, T. Assessment of unified models for estimating leaf chlorophyll content across directional-hemispherical reflectance and bidirectional reflectance spectra. Remote Sens. Environ. 2019, 231, 111240. [Google Scholar] [CrossRef]
- Lu, S.; Lu, X.; Zhao, W.; Liu, Y.; Wang, Z.; Omasa, K. Comparing vegetation indices for remote chlorophyll measurement of white poplar and Chinese elm leaves with different adaxial and abaxial surfaces. J. Exp. Bot. 2015, 66, 5625–5637. [Google Scholar] [CrossRef]
- Ran, D.; Sun, Z.; Lu, S.; Omasa, K. Optimizing angular resistant spectral indices to estimate leaf biochemical parameters from multi-angular spectral reflection. Agric. For. Meteorol. 2024, 348, 109916. [Google Scholar] [CrossRef]
- Ran, D.; Sun, Z.; Lu, S.; Omasa, K. An advanced dorsiventral leaf radiative transfer model for simulating multi-angular and spectral reflection: Considering asymmetry of leaf internal and surface structure. Remote Sens. Environ. 2025, 318, 114531. [Google Scholar] [CrossRef]
- Cheng, T.; Rivard, B.; Sánchez-Azofeifa, A.G.; Féret, J.-B.; Jacquemoud, S.; Ustin, S.L. Predicting leaf gravimetric water content from foliar reflectance across a range of plant species using continuous wavelet analysis. J. Plant Physiol. 2012, 169, 1134–1142. [Google Scholar] [CrossRef]
- Li, D.; Cheng, T.; Jia, M.; Zhou, K.; Lu, N.; Yao, X.; Tian, Y.C.; Zhu, Y.; Cao, W.X. PROCWT: Coupling PROSPECT with continuous wavelet transform to improve the retrieval of foliar chemistry from leaf bidirectional reflectance spectra. Remote Sens. Environ. 2018, 206, 1–14. [Google Scholar] [CrossRef]
- Wan, L.; Zhang, J.; Xu, Y.; Huang, Y.; Zhou, W.; Jiang, L.; He, Y.; Cen, H. PROSDM: Applicability of PROSPECT model coupled with spectral derivatives and similarity metrics to retrieve leaf biochemical traits from bidirectional reflectance. Remote Sens. Environ. 2021, 267, 112761. [Google Scholar] [CrossRef]
- Yao, C.; Sun, Z.; Lu, S. Evaluation of PROSPECT inversion approaches using multiangular spectral reflectance factor of leaves. IEEE Trans. Geosci. Remote Sens. 2024, 62, 1–16. [Google Scholar] [CrossRef]
- You, W. Methods for Estimating Water Content of Bifacial Leaf Plants. Master’s Thesis, Northeast Normal University, Changchun, China, 2018. [Google Scholar]
- Sun, Z.; Wu, Z.F.; Zhao, Y.S. Semi-automatic laboratory goniospectrometer system for performing multi-angular reflectance and polarization measurements for natural surfaces. Rev. Sci. Instrum. 2014, 85, 014503. [Google Scholar] [CrossRef]
- Dechant, B.; Cuntz, M.; Vohland, M.; Schulz, E.; Doktor, D. Estimation of photosynthesis traits from leaf reflectance spectra: Correlation to nitrogen content as the dominant mechanism. Remote Sens. Environ. 2017, 196, 279–292. [Google Scholar] [CrossRef]
- Filzmoser, P.; Liebmann, B.; Varmuza, K. Repeated double cross validation. J. Chemom. A J. Chemom. Soc. 2009, 23, 160–171. [Google Scholar] [CrossRef]
- Wold, S.; Sjöström, M.; Eriksson, L. PLS-regression: A basic tool of chemometrics. Chemom. Intell. Lab. Syst. 2001, 58, 109–130. [Google Scholar] [CrossRef]
- Demetriades-Shah, T.H.; Steven, M.D.; Clark, J.A. High resolution derivative spectra in remote sensing. Remote Sens. Environ. 1990, 33, 55–64. [Google Scholar] [CrossRef]
- Tsai, F.; Philpot, W. Derivative analysis of hyperspectral data. Remote Sens. Environ. 1998, 66, 41–51. [Google Scholar] [CrossRef]
- Torrence, C.; Compo, G.P. A practical guide to wavelet analysis. Bull. Am. Meteorol. Soc. 1998, 79, 61–78. [Google Scholar] [CrossRef]
- Rivard, B.; Feng, J.; Gallie, A.; Sanchez-Azofeifa, A. Continuous wavelets for the improved use of spectral libraries and hyperspectral data. Remote Sens. Environ. 2008, 112, 2850–2862. [Google Scholar] [CrossRef]
- Feng, J.; Rogge, D.; Rivard, B. Comparison of lithological mapping results from airborne hyperspectral VNIR-SWIR, LWIR and combined data. Int. J. Appl. Earth Obs. Geoinf. 2018, 64, 340–353. [Google Scholar] [CrossRef]
- Cheng, T.; Rivard, B.; Sánchez-Azofeifa, A.G.; Féret, J.-B.; Jacquemoud, S.; Ustin, S.L. Deriving leaf mass per area (LMA) from foliar reflectance across a variety of plant species using continuous wavelet analysis. ISPRS J. Photogramm. Remote Sens. 2014, 87, 28–38. [Google Scholar] [CrossRef]
- Li, D.; Cheng, T.; Yao, X.; Zhang, Z.; Tian, Y.; Zhu, Y.; Cao, W. Wavelet-based PROSPECT inversion for retrieving leaf mass per area (LMA) and equivalent water thickness (EWT) from leaf reflectance. In Proceedings of the 2016 IEEE International Geoscience and Remote Sensing Symposium (IGARSS), Beijing, China, 10–15 July 2016; pp. 6910–6913. [Google Scholar]
- Sun, Z.; Wu, D.; Lv, Y.; Lu, S. Optical properties of reflected light from leaves: A case study from one species. IEEE Trans. Geosci. Remote Sens. 2019, 57, 4388–4406. [Google Scholar] [CrossRef]
- Diner, D.J.; Beckert, J.C.; Reilly, T.H.; Bruegge, C.J.; Conel, J.E.; Kahn, R.A.; Martonchik, J.V.; Ackerman, T.P.; Davies, R.; Gerstl, S.A.W.; et al. Multi-angle Imaging SpectroRadiometer (MISR) instrument description and experiment overview. IEEE Trans. Geosci. Remote Sens. 1998, 36, 1072–1087. [Google Scholar] [CrossRef]
- Deschamps, P.-Y.; Bréon, F.-M.; Leroy, M.; Podaire, A.; Bricaud, A.; Buriez, J.-C.; Seze, G. The POLDER mission: Instrument characteristics and scientific objectives. IEEE Trans. Geosci. Remote Sens. 1994, 32, 598–615. [Google Scholar] [CrossRef]
- Cairns, B.; Russell, E.E.; Travis, L.D. Research scanning polarimeter: Calibration and ground-based measurements. In Proceedings of the Polarization: Measurement, Analysis, and Remote Sensing II, Denver, CO, USA, 19–21 July 1999; pp. 186–196. [Google Scholar]
- Roth, B.D.; Saunders, M.G.; Bachmann, C.M.; van Aardt, J.A. On leaf BRDF estimates and their fit to microfacet models. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2020, 13, 1761–1771. [Google Scholar] [CrossRef]
- Woolley, J.T. Reflectance and transmittance of light by leaves. Plant Physiol. 1971, 47, 656–662. [Google Scholar] [CrossRef]
- Meyer, H.; Pebesma, E. Predicting into unknown space? Estimating the area of applicability of spatial prediction models. Methods Ecol. Evol. 2021, 12, 1620–1633. [Google Scholar] [CrossRef]
- Jay, S.; Gorretta, N.; Morel, J.; Maupas, F.; Bendoula, R.; Rabatel, G.; Dutartre, D.; Comar, A.; Baret, F. Estimating leaf chlorophyll content in sugar beet canopies using millimeter- to centimeter-scale reflectance imagery. Remote Sens. Environ. 2017, 198, 173–186. [Google Scholar] [CrossRef]
- Stuckens, J.; Somers, B.; Delalieux, S.; Verstraeten, W.W.; Coppin, P. The impact of common assumptions on canopy radiative transfer simulations: A case study in Citrus orchards. J. Quant. Spectrosc. Radiat. Transf. 2009, 110, 1–21. [Google Scholar] [CrossRef]
- Chávez, R.O.; Clevers, J.G.; Verbesselt, J.; Naulin, P.I.; Herold, M. Detecting leaf pulvinar movements on NDVI time series of desert trees: A new approach for water stress detection. PLoS ONE 2014, 9, e106613. [Google Scholar] [CrossRef] [PubMed]
- Pastenes, C.; Pimentel, P.; Lillo, J. Leaf movements and photoinhibition in relation to water stress in field-grown beans. J. Exp. Bot. 2005, 56, 425–433. [Google Scholar] [CrossRef]
- Zhang, Y.; Hui, J.; Qin, Q.; Sun, Y.; Zhang, T.; Sun, H.; Li, M. Transfer-learning-based approach for leaf chlorophyll content estimation of winter wheat from hyperspectral data. Remote Sens. Environ. 2021, 267, 112724. [Google Scholar] [CrossRef]
- Chen, A.; Xu, C.; Zhang, M.; Guo, J.; Xing, X.; Yang, D.; Xu, B.; Yang, X. Cross-scale mapping of above-ground biomass and shrub dominance by integrating UAV and satellite data in temperate grassland. Remote Sens. Environ. 2024, 304, 114024. [Google Scholar] [CrossRef]
- Schaepman-Strub, G.; Schaepman, M.E.; Painter, T.H.; Dangel, S.; Martonchik, J.V. Reflectance quantities in optical remote sensing-definitions and case studies. Remote Sens. Environ. 2006, 103, 27–42. [Google Scholar] [CrossRef]
- Moreno-Martínez, Á.; Camps-Valls, G.; Kattge, J.; Robinson, N.; Reichstein, M.; van Bodegom, P.; Kramer, K.; Cornelissen, J.H.C.; Reich, P.; Bahn, M. A methodology to derive global maps of leaf traits using remote sensing and climate data. Remote Sens. Environ. 2018, 218, 69–88. [Google Scholar] [CrossRef]
- Danner, M.; Berger, K.; Wocher, M.; Mauser, W.; Hank, T. Efficient RTM-based training of machine learning regression algorithms to quantify biophysical & biochemical traits of agricultural crops. ISPRS J. Photogramm. Remote Sens. 2021, 173, 278–296. [Google Scholar]
- Yuan, H.; Yang, G.; Li, C.; Wang, Y.; Liu, J.; Yu, H.; Feng, H.; Xu, B.; Zhao, X.; Yang, X. Retrieving soybean leaf area index from unmanned aerial vehicle hyperspectral remote sensing: Analysis of RF, ANN, and SVM regression models. Remote Sens. 2017, 9, 309. [Google Scholar] [CrossRef]
- Jiao, Q.; Sun, Q.; Zhang, B.; Huang, W.; Ye, H.; Zhang, Z.; Zhang, X.; Qian, B. A random forest algorithm for retrieving canopy chlorophyll content of wheat and soybean trained with PROSAIL simulations using adjusted average leaf angle. Remote Sens. 2021, 14, 98. [Google Scholar] [CrossRef]
- Rasmussen, C.E. Gaussian processes in machine learning. In Summer School on Machine Learning; Springer: Berlin/Heidelberg, Germany, 2003; pp. 63–71. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DATAMA | MPRLDLCC | MPRLDEWT | ANGERS | |
|---|---|---|---|---|
| Biochemical traits | LCC, EWT | LCC | EWT | LCC, EWT |
| Reflectance | [BRFad, BRFab] | [BRFad, BRFab] | [BRFad, BRFab] | [DHRad] |
| Spectral Range (nm) | 400–2500 | 400–1000 | 400–2500 | 400–2450 |
| Spectrometer | ASD FieldSpec4 | ASD FieldSpec3 | ASD FieldSpec3 | ASD FieldSpec |
| Measurement | Goniometer | Leaf clip | Leaf clip | Integrating sphere |
| Plant Species | 10 | 2 | 5 | 43 |
| Number of Leaves | 176 | 115 | 300 | 276 |
| Reference | [80] | [78] | [85] | [33] |
| Strategy 1 | Strategy 2 | Strategy 3 | Strategy 4 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AD | AB | AD + AB | AD | AB | AD + AB | AD | AB | AD + AB | AD | AB | AD + AB | ||
| PLSR | R2 | 0.85 | 0.57 | 0.61 | 0.93 | 0.88 | 0.75 | 0.92 | 0.83 | 0.84 | 0.92 | 0.87 | 0.84 |
| RMSE | 8.36 | 21.29 | 16.19 | 23.53 | 6.71 | 17.31 | 7.12 | 9.11 | 8.22 | 6.73 | 8.63 | 7.76 | |
| NRMSE | 12.55 | 32.06 | 24.36 | 34.97 | 10.00 | 25.72 | 10.18 | 12.87 | 11.67 | 10.16 | 12.73 | 11.55 | |
| PLSR+FD | R2 | 0.94 | 0.70 | 0.78 | 0.93 | 0.88 | 0.74 | 0.91 | 0.77 | 0.83 | 0.93 | 0.87 | 0.87 |
| RMSE | 5.14 | 12.74 | 9.76 | 23.26 | 7.08 | 17.20 | 7.28 | 9.60 | 8.57 | 5.87 | 8.18 | 7.17 | |
| NRMSE | 7.42 | 18.08 | 13.89 | 34.27 | 10.45 | 25.35 | 10.88 | 14.03 | 12.62 | 8.86 | 12.15 | 10.71 | |
| PLSR+SD | R2 | 0.92 | 0.68 | 0.71 | 0.90 | 0.85 | 0.72 | 0.89 | 0.78 | 0.81 | 0.92 | 0.86 | 0.86 |
| RMSE | 5.78 | 14.62 | 11.13 | 22.46 | 7.53 | 16.78 | 7.88 | 9.79 | 8.94 | 6.28 | 8.25 | 7.36 | |
| NRMSE | 8.88 | 22.02 | 16.82 | 34.71 | 11.42 | 25.87 | 11.97 | 14.35 | 13.30 | 9.20 | 11.82 | 10.64 | |
| PLSR+CWT3 | R2 | 0.85 | 0.59 | 0.67 | 0.91 | 0.84 | 0.71 | 0.90 | 0.75 | 0.81 | 0.92 | 0.86 | 0.86 |
| RMSE | 8.47 | 17.15 | 13.54 | 23.18 | 7.32 | 17.22 | 7.32 | 10.23 | 8.99 | 6.36 | 8.62 | 7.63 | |
| NRMSE | 13.10 | 26.18 | 20.72 | 35.04 | 11.24 | 26.07 | 10.65 | 14.74 | 12.98 | 9.25 | 12.34 | 10.98 | |
| PLSR+CWT4 | R2 | 0.82 | 0.48 | 0.56 | 0.92 | 0.87 | 0.73 | 0.92 | 0.82 | 0.83 | 0.92 | 0.87 | 0.84 |
| RMSE | 9.28 | 22.70 | 17.36 | 23.82 | 7.13 | 17.61 | 7.23 | 9.31 | 8.42 | 6.80 | 8.81 | 7.93 | |
| NRMSE | 13.95 | 34.13 | 26.10 | 36.27 | 10.82 | 26.80 | 10.41 | 13.04 | 11.92 | 10.27 | 13.00 | 11.79 | |
| PLSR+CWT5 | R2 | 0.91 | 0.64 | 0.66 | 0.91 | 0.89 | 0.74 | 0.89 | 0.80 | 0.78 | 0.90 | 0.86 | 0.81 |
| RMSE | 6.51 | 18.91 | 14.16 | 25.41 | 7.08 | 18.67 | 8.70 | 10.08 | 9.46 | 7.40 | 9.93 | 8.81 | |
| NRMSE | 9.45 | 27.57 | 20.63 | 37.12 | 10.28 | 27.26 | 12.74 | 14.49 | 13.72 | 11.29 | 14.58 | 13.14 | |
| Strategy 1 | Strategy 2 | Strategy 3 | Strategy 4 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AD | AB | AD+AB | AD | AB | AD+AB | AD | AB | AD+AB | AD | AB | AD+AB | ||
| PLSR | R2 | 0.76 | 0.37 | 0.48 | 0.72 | 0.58 | 0.62 | 0.79 | 0.69 | 0.73 | 0.88 | 0.84 | 0.84 |
| RMSE | 0.0022 | 0.0051 | 0.0039 | 0.0036 | 0.0030 | 0.0033 | 0.0019 | 0.0022 | 0.0021 | 0.0015 | 0.0017 | 0.0016 | |
| NRMSE | 15.56 | 37.03 | 28.48 | 26.53 | 21.58 | 24.24 | 14.73 | 16.01 | 15.48 | 10.27 | 11.32 | 10.91 | |
| PLSR+FD | R2 | 0.83 | 0.54 | 0.66 | 0.82 | 0.67 | 0.68 | 0.83 | 0.70 | 0.74 | 0.90 | 0.86 | 0.86 |
| RMSE | 0.0016 | 0.0029 | 0.0023 | 0.0039 | 0.0026 | 0.0033 | 0.0018 | 0.0022 | 0.0020 | 0.0014 | 0.0017 | 0.0015 | |
| NRMSE | 11.35 | 19.39 | 15.96 | 26.17 | 17.34 | 22.24 | 13.02 | 14.63 | 13.89 | 9.13 | 10.69 | 10.00 | |
| PLSR+SD | R2 | 0.42 | 0.16 | 0.26 | 0.22 | 0.19 | 0.20 | 0.47 | 0.25 | 0.35 | 0.81 | 0.73 | 0.74 |
| RMSE | 0.0031 | 0.0042 | 0.0037 | 0.0040 | 0.0041 | 0.0041 | 0.0029 | 0.0037 | 0.0033 | 0.0018 | 0.0022 | 0.0020 | |
| NRMSE | 20.37 | 26.85 | 23.87 | 26.12 | 25.63 | 25.91 | 24.09 | 29.55 | 27.00 | 12.92 | 14.39 | 13.77 | |
| PLSR+CWT3 | R2 | 0.59 | 0.23 | 0.36 | 0.82 | 0.60 | 0.68 | 0.80 | 0.56 | 0.67 | 0.88 | 0.81 | 0.82 |
| RMSE | 0.0035 | 0.0063 | 0.0051 | 0.0035 | 0.0030 | 0.0032 | 0.0020 | 0.0027 | 0.0024 | 0.0015 | 0.0018 | 0.0017 | |
| NRMSE | 24.48 | 44.79 | 36.12 | 23.84 | 20.54 | 22.30 | 15.94 | 20.60 | 18.47 | 9.03 | 11.17 | 10.18 | |
| PLSR+CWT4 | R2 | 0.57 | 0.25 | 0.35 | 0.83 | 0.64 | 0.70 | 0.81 | 0.71 | 0.74 | 0.87 | 0.83 | 0.82 |
| RMSE | 0.0033 | 0.0063 | 0.0050 | 0.0032 | 0.0027 | 0.0030 | 0.0017 | 0.0020 | 0.0019 | 0.0014 | 0.0017 | 0.0016 | |
| NRMSE | 25.25 | 47.96 | 38.45 | 22.44 | 19.34 | 21.02 | 13.95 | 14.86 | 14.47 | 10.17 | 11.49 | 10.88 | |
| PLSR+CWT5 | R2 | 0.76 | 0.33 | 0.45 | 0.78 | 0.63 | 0.67 | 0.86 | 0.78 | 0.80 | 0.85 | 0.82 | 0.81 |
| RMSE | 0.0022 | 0.0054 | 0.0041 | 0.0034 | 0.0028 | 0.0031 | 0.0017 | 0.0018 | 0.0018 | 0.0015 | 0.0017 | 0.0016 | |
| NRMSE | 15.56 | 37.97 | 29.04 | 27.40 | 21.23 | 24.56 | 12.22 | 12.91 | 12.60 | 10.86 | 11.71 | 11.39 | |
| Biochemical Trait | Training Dataset | Calibration | Validation | ||||
|---|---|---|---|---|---|---|---|
| R2 | RMSE | NRMSE (%) | R2 | RMSE | NRMSE (%) | ||
| LCC | DATAMA | 0.88 | 7.15 | 7.58 | 0.76 | 10.39 | 9.81 |
| ANGERS | 0.95 | 4.78 | 4.97 | 0.52 | 16.82 | 17.84 | |
| MPRLDLCC | 0.87 | 6.51 | 9.92 | 0.16 | 58.76 | 55.46 | |
| EWT | DATAMA | 0.88 | 0.0014 | 5.74 | 0.88 | 0.0018 | 5.55 |
| ANGERS | 0.93 | 0.0013 | 5.17 | 0.66 | 0.0036 | 13.27 | |
| MPRLDEWT | 0.92 | 0.0007 | 6.87 | 0.08 | 0.0071 | 23.07 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ran, D.; Sun, Z.; Lu, S. Using Multi-Angular Spectral Reflection of Dorsiventral Leaves to Improve the Transferability of PLSR Models for Estimating Leaf Biochemical Traits. Remote Sens. 2025, 17, 1758. https://doi.org/10.3390/rs17101758
Ran D, Sun Z, Lu S. Using Multi-Angular Spectral Reflection of Dorsiventral Leaves to Improve the Transferability of PLSR Models for Estimating Leaf Biochemical Traits. Remote Sensing. 2025; 17(10):1758. https://doi.org/10.3390/rs17101758
Chicago/Turabian StyleRan, Dongjie, Zhongqiu Sun, and Shan Lu. 2025. "Using Multi-Angular Spectral Reflection of Dorsiventral Leaves to Improve the Transferability of PLSR Models for Estimating Leaf Biochemical Traits" Remote Sensing 17, no. 10: 1758. https://doi.org/10.3390/rs17101758
APA StyleRan, D., Sun, Z., & Lu, S. (2025). Using Multi-Angular Spectral Reflection of Dorsiventral Leaves to Improve the Transferability of PLSR Models for Estimating Leaf Biochemical Traits. Remote Sensing, 17(10), 1758. https://doi.org/10.3390/rs17101758