

Combining Multitemporal Optical and Radar Satellite Data for Mapping the Tatra Mountains Non-Forest Plant Communities




Abstract
1. Introduction
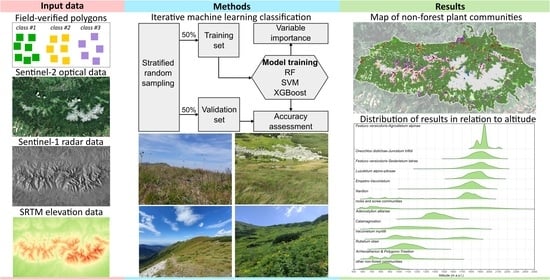
2. Materials and Methods
2.1. Study Area
2.2. Satellite Data
2.3. Reference Dataset
2.4. Classification, Accuracy Assessment, and Variable Importance
3. Results
3.1. Overall Results of Individual Classifiers and Acquisition Dates
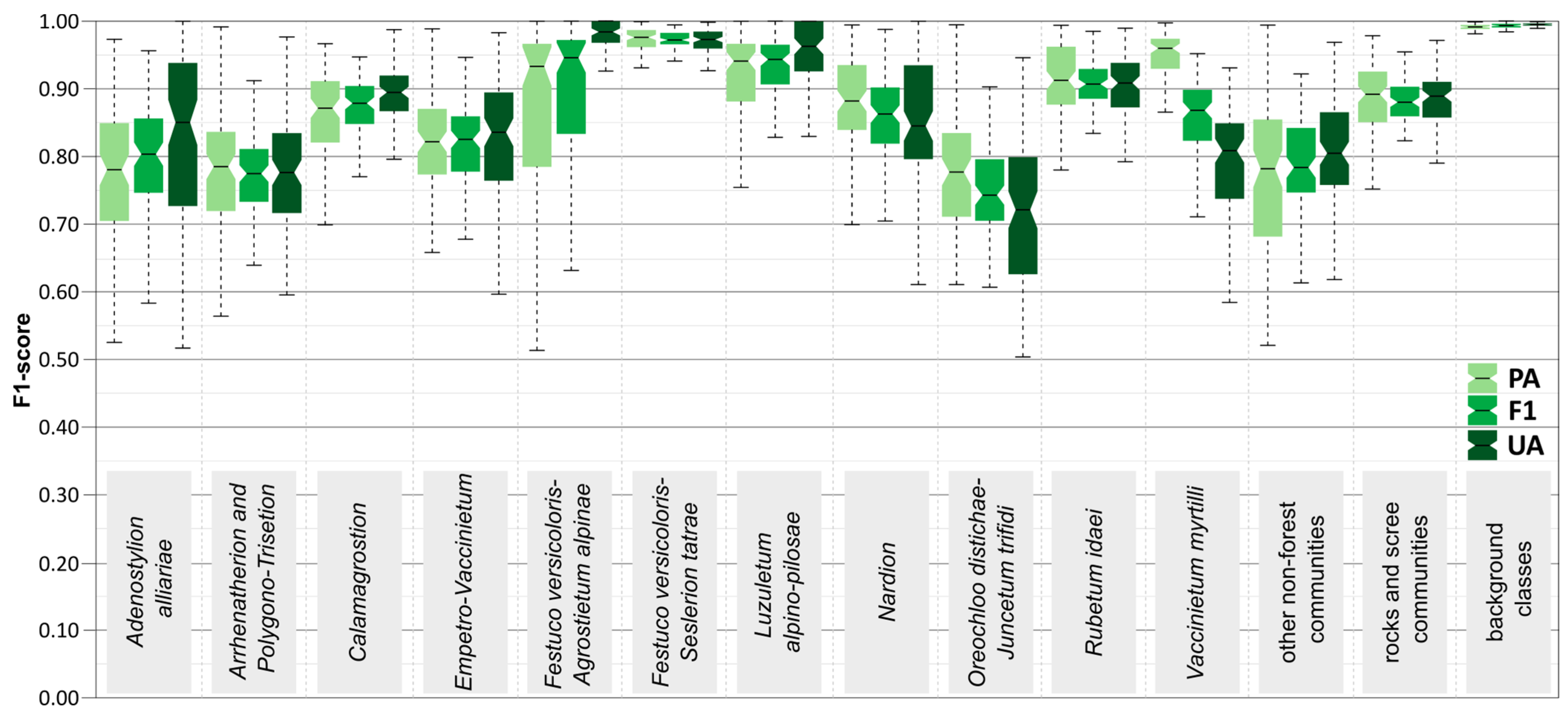
3.2. Results for Individual Vegetation Classes
3.3. Relevance of Input Data
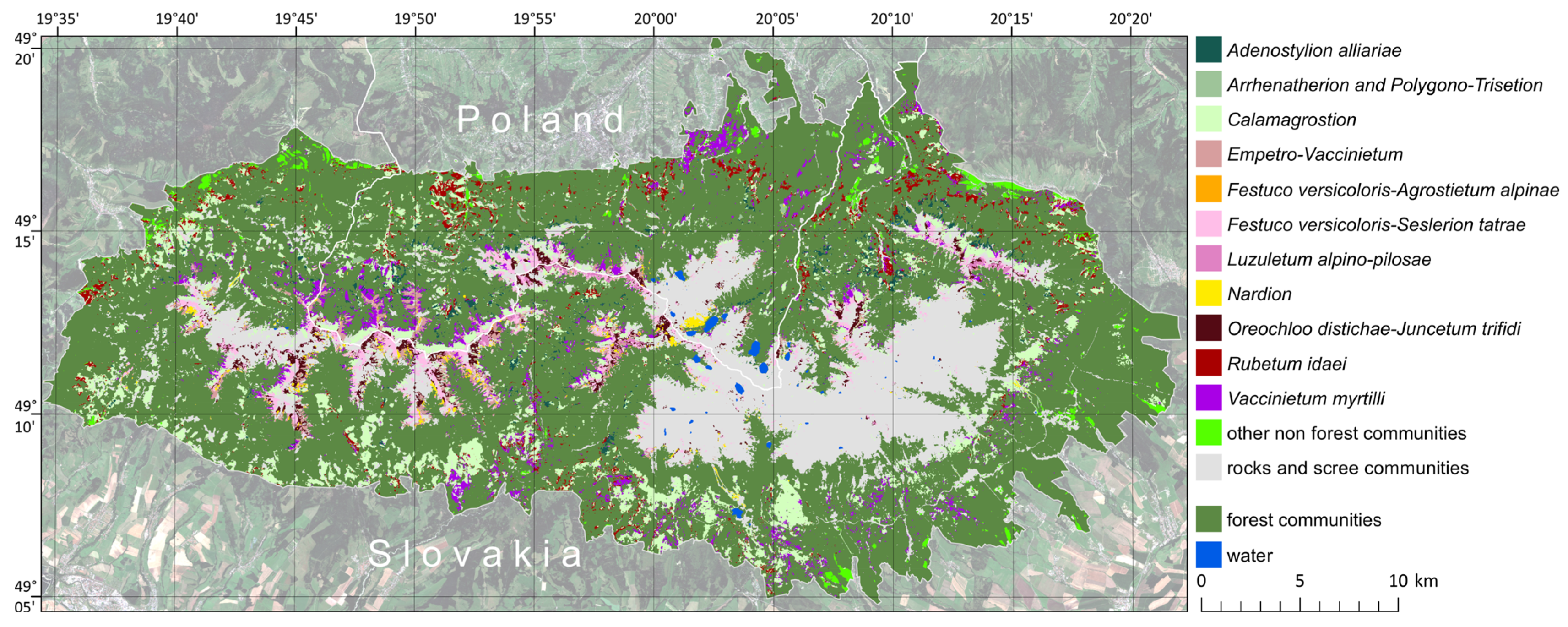
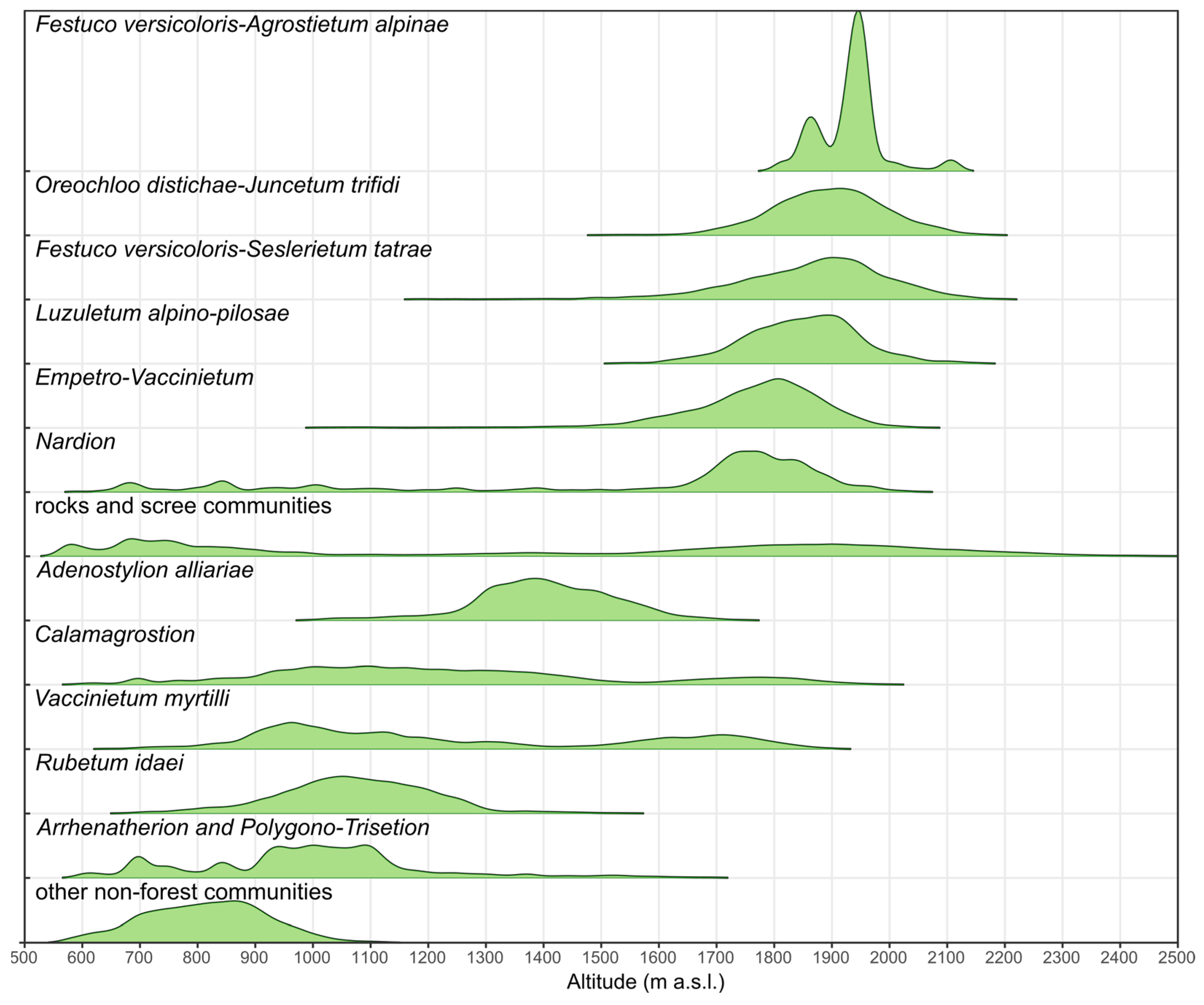
3.4. Maps of Non-Forest Vegetation Occurrence
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
References
- Schuchardt, M.A.; Berauer, B.J.; Duc, A.L.; Ingrisch, J.; Niu, Y.; Bahn, M.; Jentsch, A. Increases in functional diversity of mountain plant communities is mainly driven by species turnover under climate change. Oikos 2023, 2023, e09922. [Google Scholar] [CrossRef]
- Inouye, D.W. Effects of climate change on alpine plants and their pollinators. Ann. N. Y. Acad. Sci. 2020, 1469, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Choler, P.; Bayle, A.; Carlson, B.Z.; Randin, C.; Filippa, G.; Cremonese, E. The tempo of greening in the European Alps: Spatial variations on a common theme. Glob. Change Biol. 2021, 27, 5614–5628. [Google Scholar] [CrossRef] [PubMed]
- Steinbauer, M.; Grytnes, J.-A.; Jurasinski, G.; Kulonen, A.; Lenoir, J.; Pauli, H.; Rixen, C.; Winkler, M.; Bardy-Durchhalter, M.; Barni, E.; et al. Accelerated increase in plant species richness on mountain summits is linked to warming. Nature 2018, 556, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Peringer, A.; Frank, V.; Snell, R.S. Climate change simulations in Alpine summer pastures suggest a disruption of current vegetation zonation. Glob. Ecol. Conserv. 2022, 37, e02140. [Google Scholar] [CrossRef]
- Kabir, R.; Pomeroy, J.; Whitfield, P. Are the effects of vegetation and soil changes as important as climate change impacts on hydrological processes? Hydrol. Earth Syst. Sci. 2019, 23, 4933–4954. [Google Scholar] [CrossRef]
- Zheng, L.; Li, D.; Xu, J.; Xia, Z.; Hao, H.; Chen, Z. A twenty-years remote sensing study reveals changes to alpine pastures under asymmetric climate warming. ISPRS J. Photogramm. Remote Sens. 2022, 190, 69–78. [Google Scholar] [CrossRef]
- Laporte-Fauret, Q.; Castelle, B.; Michalet, R.; Marieu, V.; Bujan, S.; Rosebery, D. Morphological and ecological responses of a managed coastal sand dune to experimental notches. Sci. Total Environ. 2021, 782, 146813. [Google Scholar] [CrossRef] [PubMed]
- Fassnacht, F.E.; Müllerová, J.; Conti, L.; Malavasi, M.; Schmidtlein, S. About the link between biodiversity and spectral variation. Appl. Veg. Sci. 2022, 25, e12643. [Google Scholar] [CrossRef]
- Hubert-Moy, L.; Fabre, E.; Rapinel, S. Contribution of SPOT-7 multi-temporal imagery for mapping wetland vegetation. Eur. J. Remote Sens. 2020, 53, 201–210. [Google Scholar] [CrossRef]
- Greaves, H.E.; Eitel, J.U.H.; Vierling, L.A.; Boelman, N.T.; Griffin, K.L.; Magney, T.S.; Prager, C.M. 20 cm resolution mapping of tundra vegetation communities provides an ecological baseline for important research areas in a changing Arctic environment. Environ. Res. Commun. 2019, 1, 105004. [Google Scholar] [CrossRef]
- Meng, B.; Yang, Z.; Yu, H.; Qin, Y.; Sun, Y.; Zhang, J.; Chen, J.; Wang, Z.; Zhang, W.; Li, M.; et al. Mapping of Kobresia pygmaea Community Based on Umanned Aerial Vehicle Technology and Gaofen Remote Sensing Data in Alpine Meadow Grassland: A Case Study in Eastern of Qinghai–Tibetan Plateau. Remote Sens. 2021, 13, 2483. [Google Scholar] [CrossRef]
- Sun, C.; Li, J.; Liu, Y.; Liu, Y.; Liu, R. Plant species classification in salt marshes using phenological parameters derived from Sentinel-2 pixel-differential time-series. Remote Sens. Environ. 2021, 256, 112320. [Google Scholar] [CrossRef]
- Rapinel, S.; Rozo, C.; Delbosc, P.; Bioret, F.; Bouzillé, J.-B.; Hubert-Moy, L. Contribution of free satellite time-series images to mapping plant communities in the Mediterranean Natura 2000 site: The example of Biguglia Pond in Corse (France). Mediterr. Bot. 2020, 41, 181–191. [Google Scholar] [CrossRef]
- Le Dez, M.; Robin, M.; Launeau, P. Contribution of Sentinel-2 satellite images for habitat mapping of the Natura 2000 site ‘Estuaire de la Loire’ (France). Remote Sens. Appl. Soc. Environ. 2021, 24, 100637. [Google Scholar] [CrossRef]
- Hubert-Moy, L.; Rozo, C.; Perrin, G.; Bioret, F.; Rapinel, S. Large-scale and fine-grained mapping of heathland habitats using open-source remote sensing data. Remote Sens. Ecol. Conserv. 2022, 8, 448–463. [Google Scholar] [CrossRef]
- Zeferino, L.B.; Souza, L.F.T.; Amaral, C.H.; Filho, E.I.F.; Oliveira, T.S. Does environmental data increase the accuracy of land use and land cover classification? Int. J. Appl. Earth Obs. Geoinf. 2020, 91, 102128. [Google Scholar] [CrossRef]
- Waśniewski, A.; Hościło, A.; Aune-Lundberg, L. The impact of selection of reference samples and DEM on the accuracy of land cover classification based on Sentinel-2 data. Remote Sens. Appl. Soc. Environ. 2023, 32, 101035. [Google Scholar] [CrossRef]
- Xiao, H.; Su, F.; Fu, D.; Lyne, V.; Liu, G.; Pan, T.; Teng, J. Optimal and robust vegetation mapping in complex environments using multiple satellite imagery: Application to mangroves in Southeast Asia. Int. J. Appl. Earth Obs. Geoinf. 2021, 99, 102320. [Google Scholar] [CrossRef]
- Subedi, M.R.; Portillo-Quintero, C.; Kahl, S.S.; McIntyre, N.E.; Cox Robert, R.D.; Perry, G. Leveraging NAIP Imagery for Accurate Large-Area Land Use/land Cover Mapping: A Case Study in Central Texas. Photogramm. Eng. Remote Sens. 2023, 89, 547–560. [Google Scholar] [CrossRef]
- Wieland, R.; Kuhls, K.; Lentz, H.K.H.; Conraths, F.; Kampen, H.; Werner, D. Combined climate and regional mosquito habitat model based on machine learning. Ecol. Modell. 2021, 452, 109594. [Google Scholar] [CrossRef]
- Kattenborn, T.; Eichel, J.; Fassnacht, F.E. Convolutional Neural Networks enable efficient, accurate and fine-grained segmentation of plant species and communities from high-resolution UAV imagery. Sci. Rep. 2019, 9, 17656. [Google Scholar] [CrossRef] [PubMed]
- Łupikasza, E.; Szypuła, B. Vertical climatic belts in the Tatra Mountains in the light of current climate change. Theor. Appl. Climatol. 2019, 136, 249–264. [Google Scholar] [CrossRef]
- Adach, S.; Wojtkowska, M.; Religa, P. Consequences of the accessibility of the mountain national parks in Poland. Environ. Sci. Pollut. Res. 2023, 30, 27483–27500. [Google Scholar] [CrossRef]
- Kycko, M.; Zagajewski, B.; Zwijacz-Kozica, M.; Cierniewski, J.; Romanowska, E.; Orłowska, K.; Ochtyra, A.; Jarocińska, A. Assessment of Hyperspectral Remote Sensing for Analyzing the Impact of Human Trampling on Alpine Swards. Mt. Res. Dev. 2017, 37, 66–74. [Google Scholar] [CrossRef]
- Kiełtyk, P.; Delimat, A. Impact of the alien plant Impatiens glandulifera on species diversity of invaded vegetation in the northern foothills of the Tatra Mountains, Central Europe. Plant. Ecol. 2019, 220, 1–12. [Google Scholar] [CrossRef]
- Palaj, A.; Kollár, J.; Michalová, M. Changes in the Nardus grasslands in the (Sub)Alpine Zone of Western Carpathians over the last decades. Biologia 2023, 79, 1081–1090. [Google Scholar] [CrossRef]
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-scale geospatial analysis for everyone. Remote Sens. Environ. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- Potin, P.; Colin, O.; Pinheiro, M.; Rosich, B.; O’Connell, A.; Ormston, T.; Gratadour, J.-B.; Torres, R. Status and Evolution of the Sentinel-1 Mission. In Proceedings of the IEEE International Geoscience and Remote Sensing Symposium, Kuala Lumpur, Malaysia, 17–22 July 2022; Volume 2022, pp. 4707–4710. [Google Scholar] [CrossRef]
- Luo, C.; Liu, H.; Lu, L.; Liu, Z.; Kong, F.; Zhang, X. Monthly composites from Sentinel-1 and Sentinel-2 images for regional major crop mapping with Google Earth Engine. J. Integr. Agric. 2021, 20, 1944–1957. [Google Scholar] [CrossRef]
- Farr, T.G.; Rosen, P.A.; Caro, E.; Crippen, R.; Duren, R.; Hensley, S.; Kobrick, M.; Paller, M.; Rodriguez, E.; Roth, L.; et al. The Shuttle Radar Topography Mission. Rev. Geophys. 2007, 45, RG2004. [Google Scholar] [CrossRef]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Vapnik, V.N. The Nature of Statistical Learning Theory; Springer: New York, NY, USA, 1995; Volume 314. [Google Scholar] [CrossRef]
- Chen, T.; Guestrin, C. XGBoost: A Scalable Tree Boosting System. In Proceedings of the 22nd ACM SIGKDD International Conference on Knowledge Discovery and Data Mining, San Francisco, CA, USA, 13–17 August 2016; Association for Computing Machinery: New York, NY, USA, 2016; pp. 785–794. [Google Scholar] [CrossRef]
- Stehman, S.V.; Foody, G.M. Key issues in rigorous accuracy assessment of land cover products. Remote Sens. Environ. 2019, 231, 111199. [Google Scholar] [CrossRef]
- Tharwat, A. Parameter investigation of support vector machine classifier with kernel functions. Knowl. Inf. Syst. 2019, 61, 1269–1302. [Google Scholar] [CrossRef]
- Macintyre, P.; Niekerk, A.; Mucina, L. Efficacy of multi-season Sentinel-2 imagery for compositional vegetation classification. Int. J. Appl. Earth Obs. Geoinf. 2020, 85, 101980. [Google Scholar] [CrossRef]
- Dobrinić, D.; Gašparović, M.; Medak, D. Sentinel-1 and 2 Time-Series for Vegetation Mapping Using Random Forest Classification: A Case Study of Northern Croatia. Remote Sens. 2021, 13, 2321. [Google Scholar] [CrossRef]
- Heydari, S.; Mountrakis, G. Effect of classifier selection, reference sample size, reference class distribution and scene heterogeneity in per-pixel classification accuracy using 26 Landsat sites. Remote Sens. Environ. 2018, 204, 648–658. [Google Scholar] [CrossRef]
- Raab, C.; Stroh, H.G.; Tonn, B.; Meißner, M.; Rohwer, N.; Balkenhol, N.; Isselstein, J. Mapping semi-natural grassland communities using multi-temporal RapidEye remote sensing data. Int. J. Remote Sens. 2018, 39, 5638–5659. [Google Scholar] [CrossRef]
- Jarocińska, A.; Kopeć, D.; Niedzielko, J.; Wylazłowska, J.; Halladin-Dąbrowska, A.; Charyton, J.; Piernik, A.; Kamiński, D. The utility of airborne hyperspectral and satellite multispectral images in identifying Natura 2000 non-forest habitats for conservation purposes. Sci. Rep. 2023, 13, 4549. [Google Scholar] [CrossRef] [PubMed]
- Bhatnagar, S.; Gill, L.; Regan, S.; Naughton, O.; Johnston, P.; Waldren, S.; Ghosh, B. Mapping Vegetation Communities Inside Wetlands Using Sentinel-2 Imagery in Ireland. Int. J. Appl. Earth Obs. Geoinf. 2020, 88, 102083. [Google Scholar] [CrossRef]
- Peng, K.; Jiang, W.; Hou, P.; Wu, Z.; Ling, Z.; Wang, X.; Niu, Z.; Mao, D. Continental-scale wetland mapping: A novel algorithm for detailed wetland types classification based on time series Sentinel-1/2 images. Ecol. Indic. 2023, 148, 110113. [Google Scholar] [CrossRef]
- Kupková, L.; Cervená, L.; Suchá, R.; Jakešová, L.; Zagajewski, B.; Brezina, S.; Albrechtová, J. Classification of tundra vegetation in the Krkonoše Mts. National park using APEX, AISA dual and Sentinel-2A data. Eur. J. Remote Sens. 2017, 50, 29–46. [Google Scholar] [CrossRef]
- Marcinkowska-Ochtyra, A.; Zagajewski, B.; Ochtyra, A.; Jarocińska, A.; Wojtuń, B.; Rogass, C.; Mielke, C.; Lavender, S. Subalpine and Alpine Vegetation Classification based on Hyperspectral APEX and Simulated EnMAP images. Int. J. Remote Sens. 2017, 38, 1839–1864. [Google Scholar] [CrossRef]
- Pandey, A.; Singh, G.; Palni, S.; Chandra, N.; Rawa, J.; Singh, A.P. Application of remote sensing in alpine grasslands cover mapping of western Himalaya, Uttarakhand, India. Environ. Monit. Assess. 2021, 193, 166. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.P.; Rai, I.D.; Pangtey, D.; Padalia, H. Vegetation Characterization at Community Level Using Sentinel-2 SatelliteData and Random Forest Classifier in Western Himalayan Foothills, Uttarakhand. J. Indian Soc. Remote Sens. 2021, 49, 759–771. [Google Scholar] [CrossRef]
- Kluczek, M.; Zagajewski, B.; Kycko, M. Airborne HySpex Hyperspectral Versus Multitemporal Sentinel-2 Images for Mountain Plant Communities Mapping. Remote Sens. 2022, 14, 1209. [Google Scholar] [CrossRef]
- Kluczek, M.; Zagajewski, B.; Zwijacz-Kozica, T. Mountain Tree Species Mapping Using Sentinel-2, PlanetScope, and Airborne HySpex Hyperspectral Imagery. Remote Sens. 2023, 15, 844. [Google Scholar] [CrossRef]
- Zagajewski, B.; Kluczek, M.; Raczko, E.; Njegovec, A.; Dabija, A.; Kycko, M. Comparison of Random Forest, Support Vector Machines, and Neural Networks for Post-Disaster Forest Species Mapping of the Krkonoše/Karkonosze Transboundary Biosphere Reserve. Remote Sens. 2021, 13, 2581. [Google Scholar] [CrossRef]
- Zagajewski, B.; Kluczek, M.; Zdunek, K.B.; Holland, D. Sentinel-2 versus PlanetScope Images for Goldenrod Invasive Plant Species Mapping. Remote Sens. 2024, 16, 636. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S1A | S1B | |||||
|---|---|---|---|---|---|---|
| ASC | DSC | ASC | DSC | Total | Mean Scenes per Month | |
| 2019 | 35 | 34 | 54 | 34 | 157 | 22 |
| 2020 | 36 | 35 | 52 | 35 | 158 | 23 |
| 2021 | 35 | 34 | 54 | 35 | 158 | 23 |
| 2022 | 35 | 33 | 0 | 0 | 68 | 10 |
| Class | Description | No. of Sentinel-2 Pixels | No. of Polygons |
|---|---|---|---|
| Adenostylion alliariae | Low shrubs, mountain herbs | 405 | 22 |
| Arrhenatherion and Polygono-Trisetion | Mountain hay meadows | 1898 | 63 |
| Calamagrostion | Alpine grasslands | 1965 | 39 |
| Empetro-Vaccinietum | Low shrubs, heath communities | 527 | 27 |
| Festuco versicoloris-Agrostietum alpinae | Alpine calcareous grasslands | 163 | 12 |
| Festuco versicoloris-Seslerietum tatrae | Alpine calcareous grasslands | 730 | 29 |
| Luzuletum alpino-pilosae | Alpine grasslands | 102 | 18 |
| Nardion | Alpine grasslands | 355 | 28 |
| Oreochloo distichae-Juncetum trifidi | Siliceous alpine grasslands | 412 | 42 |
| Rubetum idaei | Shrubs with domination of raspberry | 920 | 56 |
| Vaccinietum myrtilli | Low shrubs with domination of Vaccinium myrtillus | 1394 | 50 |
| Other non-forest | Initial and degraded plant communities | 3215 | 63 |
| Rocks and scree communities | Vegetation and lichens growing on a loose bedrock or bare rock | 4116 | 75 |
| Forest (background) | Coniferous and deciduous forests plant communities | 21,674 | 430 |
| Water (background) | Stream and mountain lake waters | 9519 | 14 |
| Total | 47,395 | 968 |
| One Year | 2019–2022 | |||||||
|---|---|---|---|---|---|---|---|---|
| Dataset | No. of Variables | RF mtry | SVM Cost | SVM Gamma | No. of Variables | RF mtry | SVM Cost | SVM Gamma |
| S1 | 28 | 5 | 1 | 0.10 | 112 | 15 | 10 | 0.01 |
| S1 + TF | 31 | 5 | 1 | 0.10 | 115 | 15 | 10 | 0.01 |
| S2 | 48–72 | 7 | 1 | 0.10 | 240 | 25 | 100 | 0.01 |
| S2 + TF | 51–75 | 7 | 1 | 0.10 | 243 | 25 | 100 | 0.01 |
| S1 + S2 | 76–100 | 10 | 100 | 0.01 | 352 | 40 | 100 | 0.01 |
| S1 + S2 + TF | 79–103 | 10 | 100 | 0.01 | 355 | 40 | 100 | 0.01 |
| Class | S1 + S2 + TF | S1+ S2 | S2 + TF | S2 | S1 + TF | S1 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RF | SVM | RF | SVM | RF | SVM | RF | SVM | RF | SVM | RF | SVM | |
| Adenostylion alliariae | 0.78 | 0.70 | 0.59 | 0.68 | 0.76 | 0.82 | 0.51 | 0.77 | 0.41 | 0.41 | 0.25 | 0.36 |
| Arrhenatherion and Polygono–Trisetion | 0.77 | 0.75 | 0.70 | 0.74 | 0.79 | 0.77 | 0.71 | 0.76 | 0.71 | 0.76 | 0.63 | 0.71 |
| Calamagrostion | 0.87 | 0.82 | 0.87 | 0.82 | 0.87 | 0.88 | 0.86 | 0.88 | 0.51 | 0.67 | 0.49 | 0.65 |
| Empetro–Vaccinietum | 0.81 | 0.70 | 0.80 | 0.70 | 0.80 | 0.80 | 0.79 | 0.79 | 0.52 | 0.52 | 0.47 | 0.50 |
| Festuco versicoloris–Agrostietum alpinae | 0.89 | 0.95 | 0.89 | 0.95 | 0.88 | 0.96 | 0.88 | 0.96 | 0.88 | 0.86 | 0.74 | 0.82 |
| Festuco versicoloris–Seslerietum tatrae | 0.78 | 0.66 | 0.77 | 0.65 | 0.78 | 0.75 | 0.76 | 0.75 | 0.58 | 0.60 | 0.46 | 0.56 |
| Luzuletum alpino-pilosae | 0.92 | 0.77 | 0.92 | 0.76 | 0.88 | 0.80 | 0.88 | 0.80 | 0.58 | 0.60 | 0.19 | 0.51 |
| Nardion | 0.86 | 0.86 | 0.85 | 0.86 | 0.84 | 0.89 | 0.84 | 0.89 | 0.70 | 0.72 | 0.64 | 0.69 |
| Oreochloo distichae–Juncetum trifidi | 0.73 | 0.67 | 0.72 | 0.68 | 0.74 | 0.69 | 0.73 | 0.69 | 0.64 | 0.54 | 0.53 | 0.54 |
| Rubetum idaei | 0.86 | 0.86 | 0.79 | 0.85 | 0.86 | 0.91 | 0.77 | 0.90 | 0.55 | 0.70 | 0.50 | 0.66 |
| Vaccinietum myrtilli | 0.88 | 0.84 | 0.87 | 0.84 | 0.87 | 0.86 | 0.85 | 0.86 | 0.61 | 0.72 | 0.57 | 0.66 |
| Other non-forest | 0.91 | 0.89 | 0.88 | 0.88 | 0.91 | 0.89 | 0.87 | 0.88 | 0.84 | 0.88 | 0.77 | 0.83 |
| Rocks and scree communities | 0.97 | 0.82 | 0.97 | 0.82 | 0.97 | 0.92 | 0.96 | 0.92 | 0.83 | 0.83 | 0.74 | 0.79 |
| Forest (background) | 0.99 | 0.99 | 0.99 | 0.99 | 0.99 | 0.99 | 0.99 | 0.99 | 0.94 | 0.96 | 0.93 | 0.96 |
| Water (background) | 1.00 | 0.95 | 1.00 | 0.95 | 0.99 | 0.99 | 0.99 | 0.99 | 0.97 | 0.96 | 0.95 | 0.95 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kluczek, M.; Zagajewski, B.; Kycko, M. Combining Multitemporal Optical and Radar Satellite Data for Mapping the Tatra Mountains Non-Forest Plant Communities. Remote Sens. 2024, 16, 1451. https://doi.org/10.3390/rs16081451
Kluczek M, Zagajewski B, Kycko M. Combining Multitemporal Optical and Radar Satellite Data for Mapping the Tatra Mountains Non-Forest Plant Communities. Remote Sensing. 2024; 16(8):1451. https://doi.org/10.3390/rs16081451
Chicago/Turabian StyleKluczek, Marcin, Bogdan Zagajewski, and Marlena Kycko. 2024. "Combining Multitemporal Optical and Radar Satellite Data for Mapping the Tatra Mountains Non-Forest Plant Communities" Remote Sensing 16, no. 8: 1451. https://doi.org/10.3390/rs16081451
APA StyleKluczek, M., Zagajewski, B., & Kycko, M. (2024). Combining Multitemporal Optical and Radar Satellite Data for Mapping the Tatra Mountains Non-Forest Plant Communities. Remote Sensing, 16(8), 1451. https://doi.org/10.3390/rs16081451