



Cumulative and Lagged Effects: Seasonal Characteristics of Drought Effects on East Asian Grasslands
 ,
,  ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
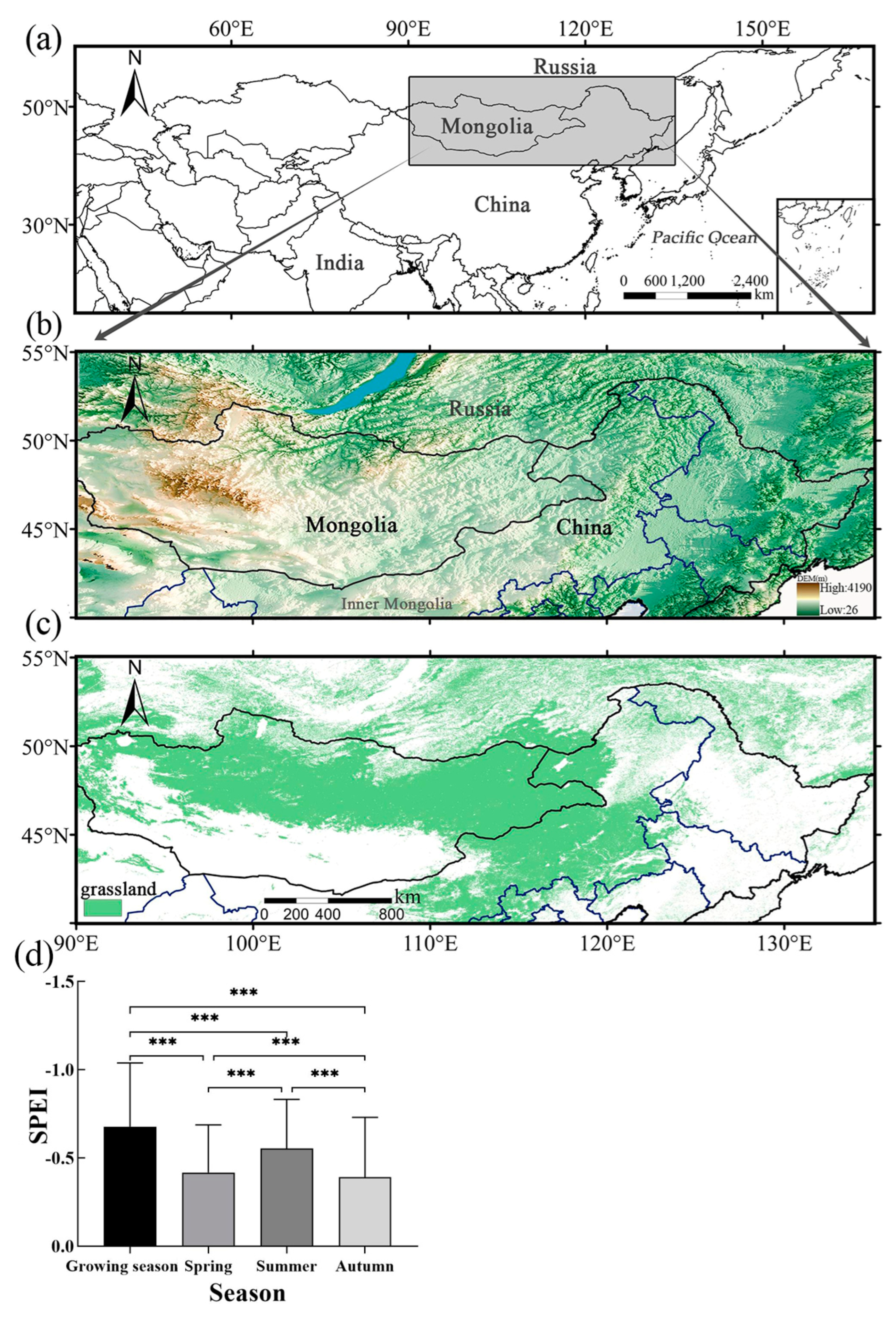
2.1. Study Area
2.2. Vegetation NDVI Data Source and Processing
2.3. Vegetation Type Data and Processing
2.4. SPEI Data Source and Processing
2.5. Research Methods
2.5.1. Cumulative Effect of Drought on NDVI
2.5.2. Lagged Effect of Drought on the NDVI
3. Results
3.1. Cumulative Effect of Drought on the Grassland NDVI
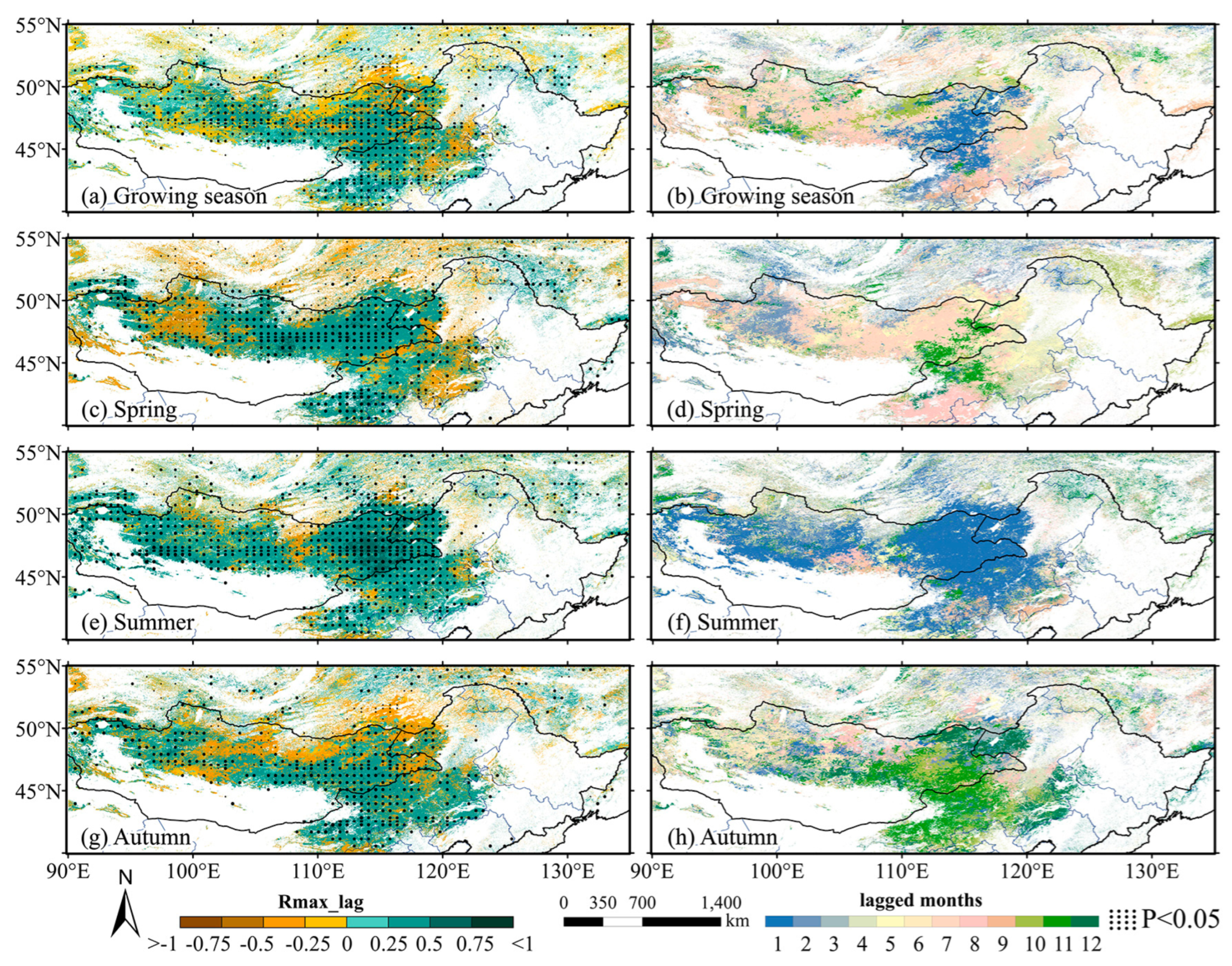
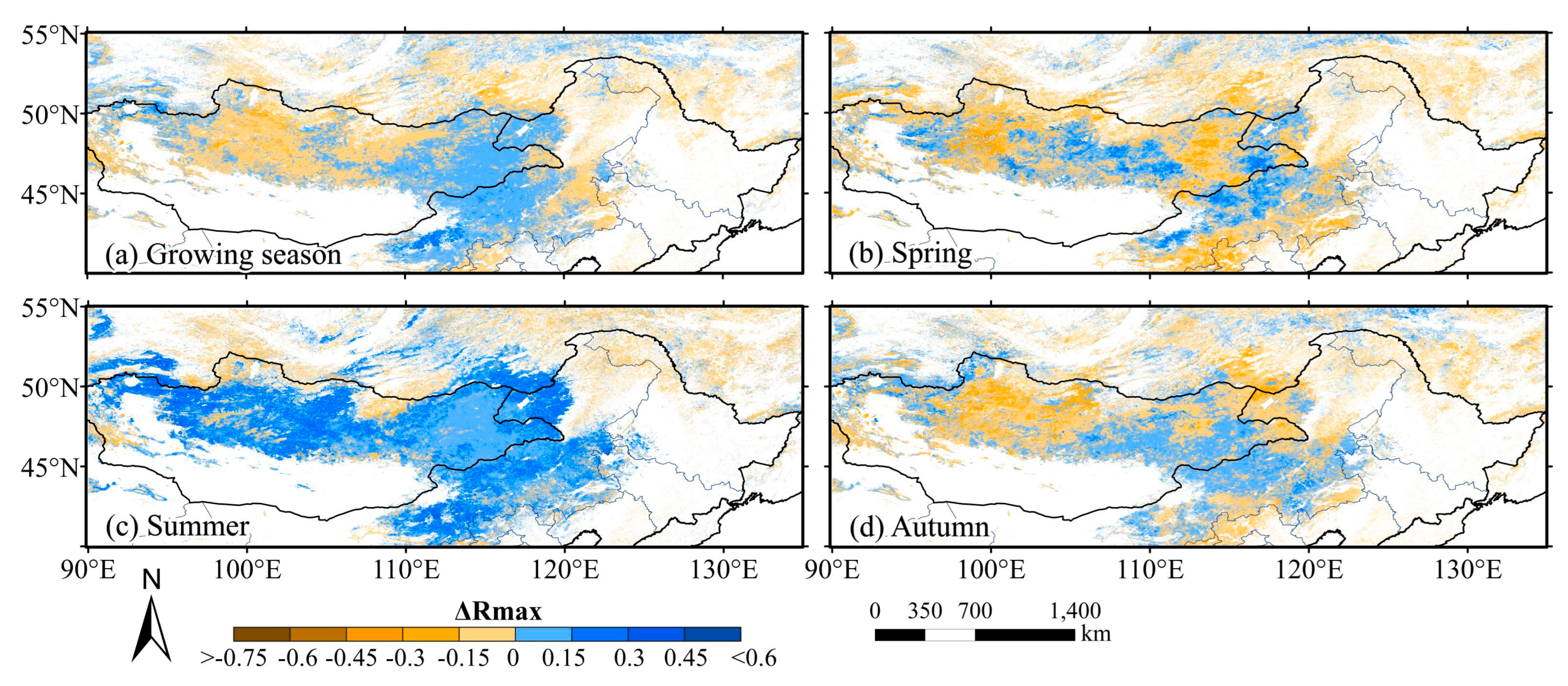
3.1.1. Spatial Distribution Characteristics of the Cumulative Effect
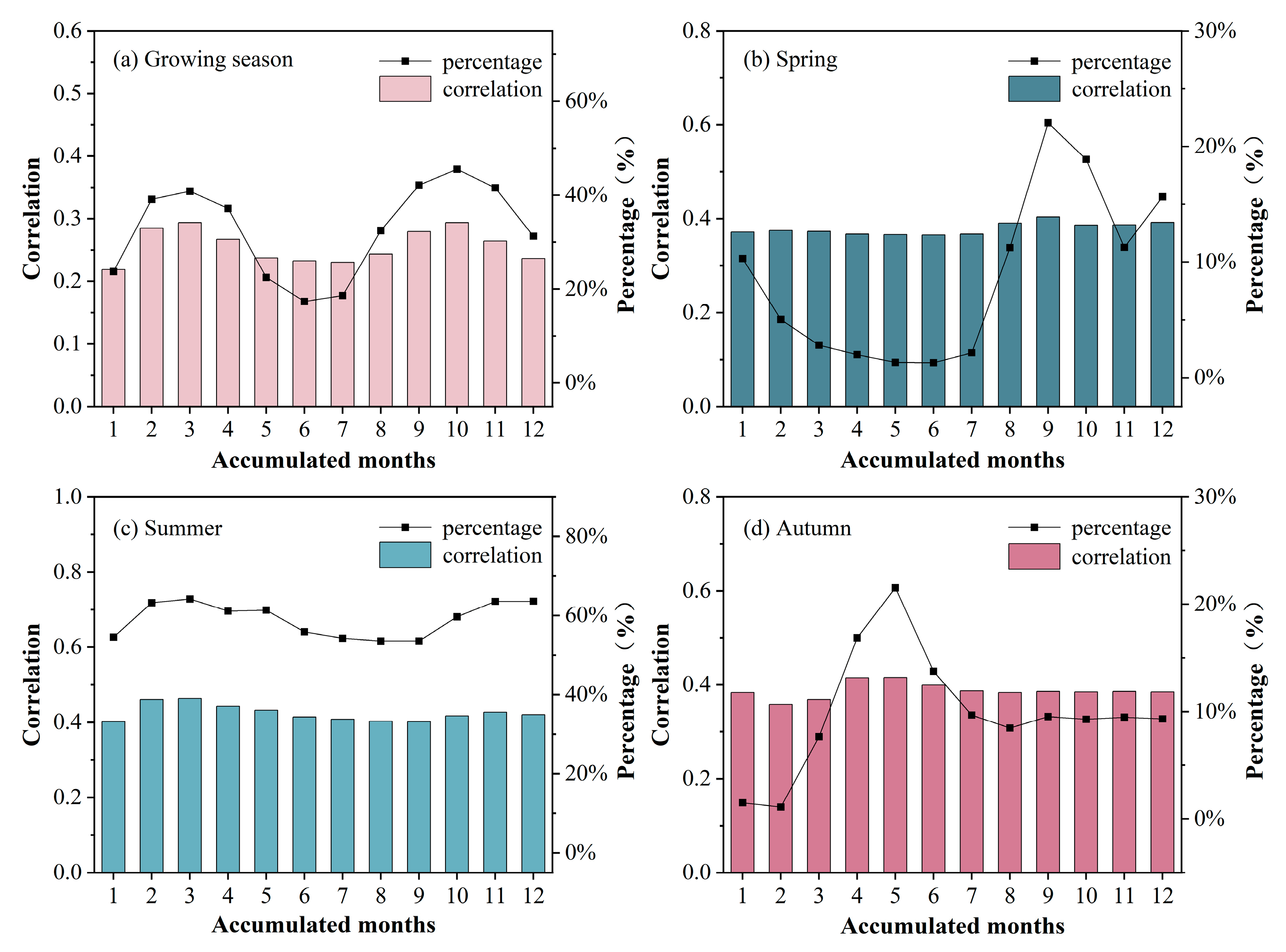
3.1.2. Temporal Characteristics of the Cumulative Effect
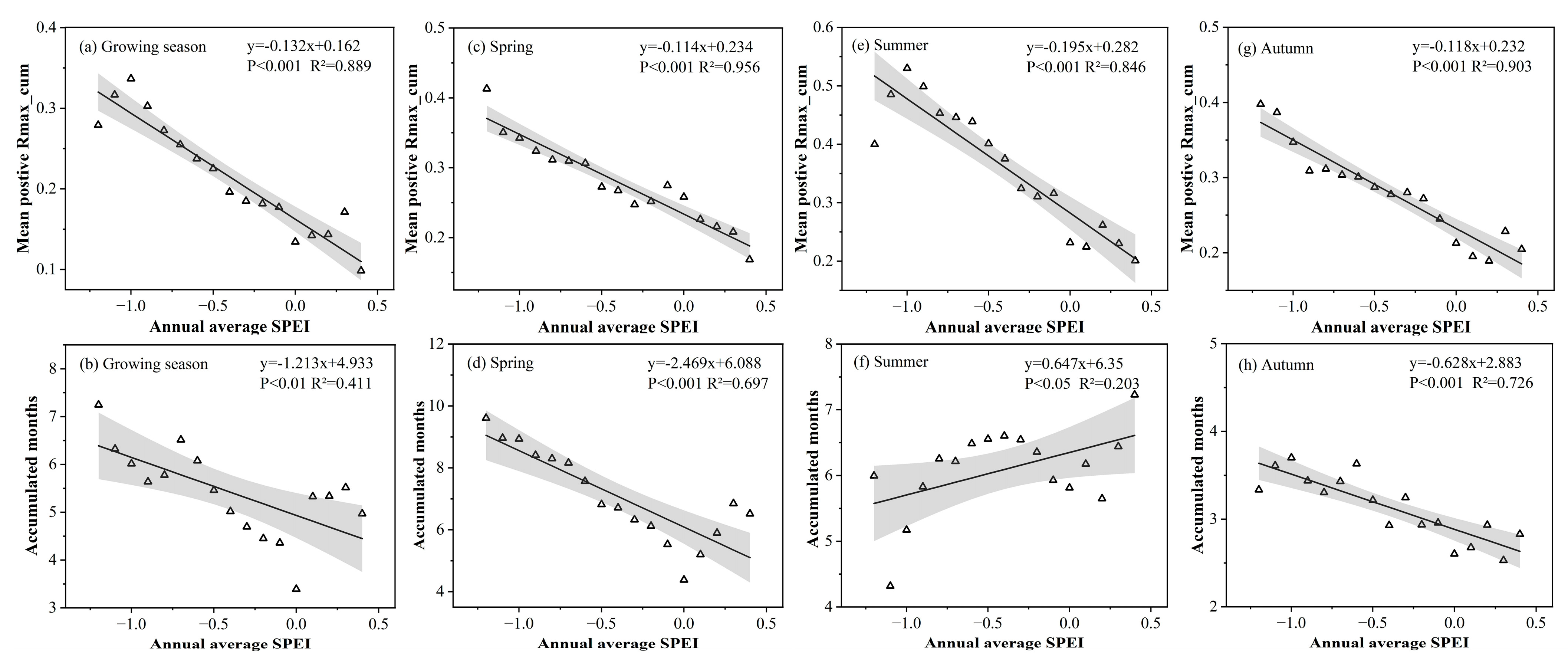
3.1.3. Cumulative Effect of Drought under Different Moisture Conditions
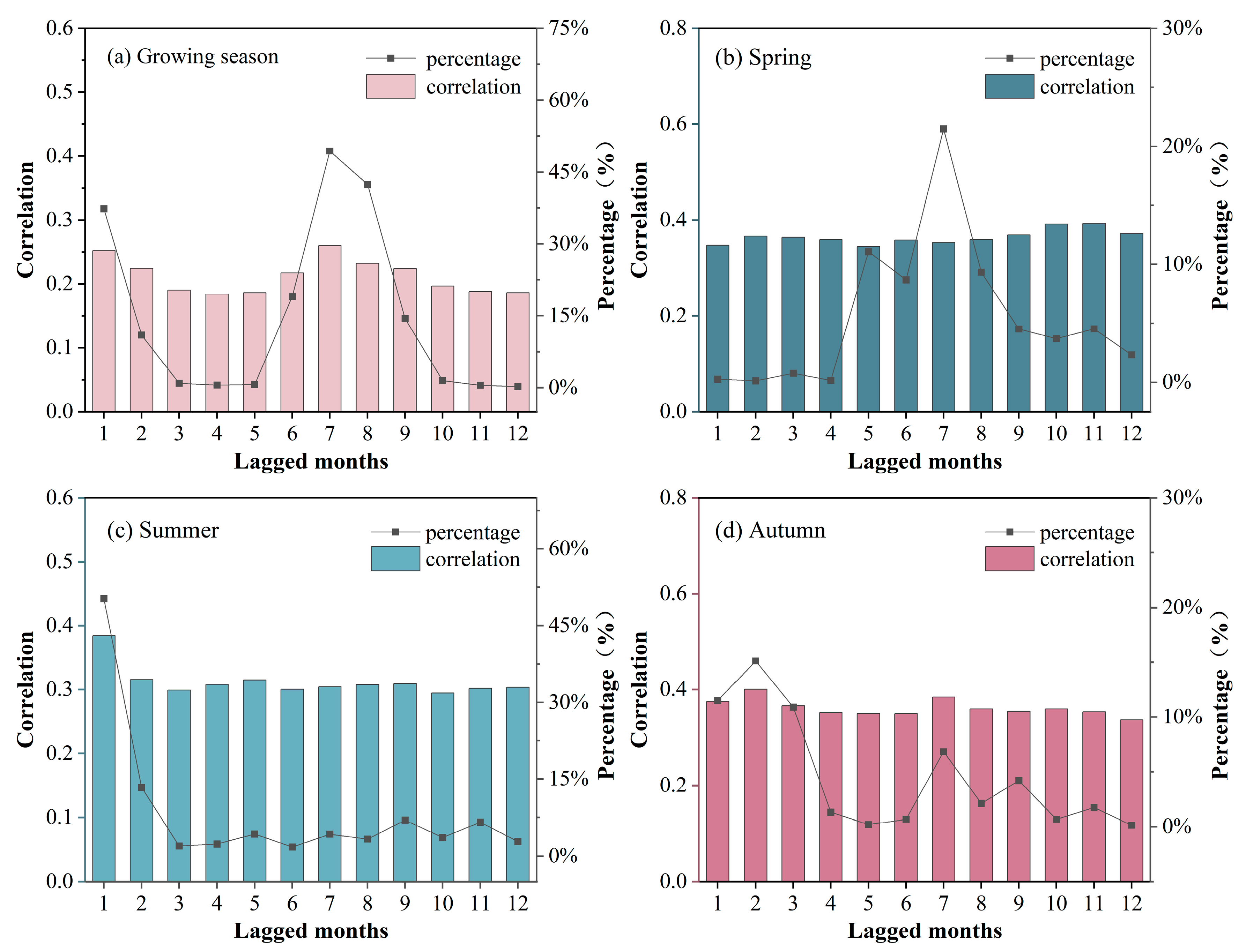
3.2. Lagged Effect of Drought on Grassland NDVI
3.2.1. Spatial Distribution Characteristics of the Lagged Effect
3.2.2. Temporal Characteristics of the Lagged Effect
3.2.3. Lagged Effect of Drought under Different Moisture Conditions
3.3. Comparison of the Lag and Cumulative Effects of Drought on Grassland NDVI
4. Discussion
5. Limitations and Prospects
6. Conclusions
- (1)
- There were differences in the response of grassland to drought CALEs in different seasons. The influence of CEs in summer was stronger than that in spring and autumn, and there was no obvious difference in LEs in the three seasons. Spatially, the area significantly affected by CALEs is greater in summer than spring and autumn. The cumulative and lagging time scale of spring is larger than that of summer and autumn;
- (2)
- Grasslands are most sensitive to drought in summer. As moisture decreases: the lag time scale gradually shortens in summer and autumn but shows an inverted U-shape in spring; the cumulative time scale gradually increases in spring and autumn while decreasing in summer. The spatial variability of the spring cumulative time scale is the largest, while that of the summer lag time scale is the largest;
- (3)
- For the growing season as a whole, the LE of drought on vegetation was greater than the CE in 54.89% of the study area. Different seasons had different dominant effects: in spring, the LE was dominant in 65.87% of the area; in summer, the CE was dominant in 71.81% of the area; and in autumn, the LE was dominant in 59.82% of the area.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kannenberg, S.A.; Novick, K.A.; Alexander, M.R.; Maxwell, J.T.; Moore, D.J.P.; Phillips, R.P.; Anderegg, W.R.L. Linking drought legacy effects across scales: From leaves to tree rings to ecosystems. Glob. Change Biol. 2019, 25, 2978–2992. [Google Scholar] [CrossRef]
- Liu, B.; Zhou, W.; Henderson, M.; Sun, Y.; Shen, X. Climatology of the Soil Surface Diurnal Temperature Range in a Warming World: Annual Cycles, Regional Patterns, and Trends in China. Earth’s Future 2022, 10, e2021EF002220. [Google Scholar] [CrossRef]
- Piao, S.; Mohammat, A.; Fang, J.; Cai, Q.; Feng, J. NDVI-based increase in growth of temperate grasslands and its responses to climate changes in China. Glob. Environ. Change 2006, 16, 340–348. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Schwalm, C.; Biondi, F.; Camarero, J.J.; Koch, G.; Litvak, M.; Ogle, K.; Shaw, J.D.; Shevliakova, E.; Williams, A.P.; et al. Pervasive drought legacies in forest ecosystems and their implications for carbon cycle models. Science 2015, 349, 528–532. [Google Scholar] [CrossRef]
- Wang, Y.; Lv, W.; Xue, K.; Wang, S.; Zhang, L.; Hu, R.; Zeng, H.; Xu, X.; Li, Y.; Jiang, L.; et al. Grassland changes and adaptive management on the Qinghai–Tibetan Plateau. Nat. Rev. Earth Environ. 2022, 3, 668–683. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, T.; Wang, Q.; Peng, L. Time-lagged and cumulative effects of drought and anthropogenic activities on China’s vegetation greening from 1990 to 2018. Int. J. Digit. Earth 2023, 16, 2233–2258. [Google Scholar] [CrossRef]
- Song, L.; Luo, W.; Griffin-Nolan, R.J.; Ma, W.; Cai, J.; Zuo, X.; Yu, Q.; Hartmann, H.; Li, M.H.; Smith, M.D.; et al. Differential responses of grassland community nonstructural carbohydrate to experimental drought along a natural aridity gradient. Sci. Total Environ. 2022, 822, 153589. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, S.; Sun, Y.; Li, M.; An, Y.; Shi, F. Spatial differentiation of the NPP and NDVI and its influencing factors vary with grassland type on the Qinghai-Tibet Plateau. Environ. Monit. Assess. 2021, 193, 48. [Google Scholar] [CrossRef]
- Lu, J.; Peng, J.; Li, G.; Guan, J.; Han, W.; Liu, L.; Wang, R.; Zheng, J. Assessment of time-lag and cumulative effects of drought on gross primary productivity of grassland in Central Asia from 1982 to 2018. Acta Ecol. Sin. 2023, 43, 9745–9757. [Google Scholar] [CrossRef]
- Peng, J.; Wu, C.; Zhang, X.; Wang, X.; Gonsamo, A. Satellite detection of cumulative and lagged effects of drought on autumn leaf senescence over the Northern Hemisphere. Glob. Change Biol. 2019, 25, 2174–2188. [Google Scholar] [CrossRef] [PubMed]
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I. A Multiscalar Drought Index Sensitive to Global Warming: The Standardized Precipitation Evapotranspiration Index. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef]
- Ivits, E.; Horion, S.; Erhard, M.; Fensholt, R. Assessing European ecosystem stability to drought in the vegetation growing season. Glob. Ecol. Biogeogr. 2016, 25, 1131–1143. [Google Scholar] [CrossRef]
- Ding, Y.; Li, Z.; Peng, S. Global analysis of time-lag and -accumulation effects of climate on vegetation growth. Int. J. Appl. Earth Obs. Geoinf. 2020, 92, 102179. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Gouveia, C.; Camarero, J.J.; Begueria, S.; Trigo, R.; Lopez-Moreno, J.I.; Azorin-Molina, C.; Pasho, E.; Lorenzo-Lacruz, J.; Revuelto, J.; et al. Response of vegetation to drought time-scales across global land biomes. Proc. Natl. Acad. Sci. USA 2013, 110, 52–57. [Google Scholar] [CrossRef]
- Gu, X.; Guo, E.; Yin, S.; Wang, Y.; Na, R.; Wan, Z. Assessment of the Cumulative and Lagging Effects of Drought on Vegetation Growth in Inner Mongolia. Acta Agrestia Sin. 2021, 29, 1301–1310. [Google Scholar] [CrossRef]
- Wei, W.; Liu, T.; Zhou, L.; Wang, J.; Yan, P.; Xie, B.; Zhou, J. Drought-Related Spatiotemporal Cumulative and Time-Lag Effects on Terrestrial Vegetation across China. Remote Sens. 2023, 15, 4362. [Google Scholar] [CrossRef]
- Zhao, A.; Yu, Q.; Feng, L.; Zhang, A.; Pei, T. Evaluating the cumulative and time-lag effects of drought on grassland vegetation: A case study in the Chinese Loess Plateau. J. Environ. Manag. 2020, 261, 110214. [Google Scholar] [CrossRef]
- Liu, L.; Guan, J.; Zheng, J.; Wang, Y.; Han, W.; Liu, Y. Cumulative effects of drought have an impact on net primary productivity stability in Central Asian grasslands. J. Environ. Manag. 2023, 344, 118734. [Google Scholar] [CrossRef]
- Wei, X.; He, W.; Zhou, Y.; Ju, W.; Xiao, J.; Li, X.; Liu, Y.; Xu, S.; Bi, W.; Zhang, X.; et al. Global assessment of lagged and cumulative effects of drought on grassland gross primary production. Ecol. Indic. 2022, 136, 108646. [Google Scholar] [CrossRef]
- Kong, D.; Zhang, Q.; Singh, V.P.; Shi, P. Seasonal vegetation response to climate change in the Northern Hemisphere (1982–2013). Glob. Planet. Change 2017, 148, 1–8. [Google Scholar] [CrossRef]
- Li, L.; Qian, R.; Liu, W.; Wang, W.; Biederman, J.A.; Zhang, B.; Kang, X.; Wen, F.; Ran, Q.; Zheng, Z.; et al. Drought timing influences the sensitivity of a semiarid grassland to drought. Geoderma 2022, 412, 115714. [Google Scholar] [CrossRef]
- Hahn, C.; Lüscher, A.; Ernst-Hasler, S.; Suter, M.; Kahmen, A. Timing of drought in the growing season and strong legacy effects determine the annual productivity of temperate grasses in a changing climate. Biogeosciences 2021, 18, 585–604. [Google Scholar] [CrossRef]
- Dietrich, J.D.; Smith, M.D. The effect of timing of growing season drought on flowering of a dominant C4 grass. Oecologia 2016, 181, 391–399. [Google Scholar] [CrossRef]
- Gang, C.; Zhang, Y.; Wang, Z.; Chen, Y.; Yang, Y.; Li, J.; Cheng, J.; Qi, J.; Odeh, I. Modeling the dynamics of distribution, extent, and NPP of global terrestrial ecosystems in response to future climate change. Glob. Planet. Change 2017, 148, 153–165. [Google Scholar] [CrossRef]
- Yu, Z.; Wang, J.; Liu, S.; Rentch, J.S.; Sun, P.; Lu, C. Global gross primary productivity and water use efficiency changes under drought stress. Environ. Res. Lett. 2017, 12, 014016. [Google Scholar] [CrossRef]
- Zhang, P.; Jeong, J.-H.; Yoon, J.-H.; Kim, H.; Wang, S.-Y.S.; Linderholm, H.W.; Fang, K.; Wu, X.; Chen, D. Abrupt shift to hotter and drier climate over inner East Asia beyond the tipping point. Science 2020, 370, 1095–1099. [Google Scholar] [CrossRef]
- De Boeck, H.J.; Dreesen, F.E.; Janssens, I.A.; Nijs, I. Whole-system responses of experimental plant communities to climate extremes imposed in different seasons. New Phytol. 2011, 189, 806–817. [Google Scholar] [CrossRef]
- Denton, E.M.; Dietrich, J.D.; Smith, M.D.; Knapp, A.K. Drought timing differentially affects above- and belowground productivity in a mesic grassland. Plant Ecol. 2017, 218, 317–328. [Google Scholar] [CrossRef]
- Xu, X.; Liu, H.; Wang, W.; Hu, G.; Wu, X.; Song, Z. Effects of manipulated precipitation on aboveground net primary productivity of grassland fields: Controlled rainfall experiments in Inner Mongolia, China. Land Degrad. Dev. 2021, 32, 1981–1992. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Huang, S.; Tang, L.; Hupy, J.P.; Wang, Y.; Shao, G. A commentary review on the use of normalized difference vegetation index (NDVI) in the era of popular remote sensing. J. For. Res. 2021, 32, 1–6. [Google Scholar] [CrossRef]
- Meng, X.; Gao, X.; Li, S.; Lei, J. Spatial and Temporal Characteristics of Vegetation NDVI Changes and the Driving Forces in Mongolia during 1982–2015. Remote Sens. 2020, 12, 603. [Google Scholar] [CrossRef]
- Luo, N.; Mao, D.; Wen, B.; Liu, X. Climate Change Affected Vegetation Dynamics in the Northern Xinjiang of China: Evaluation by SPEI and NDVI. Land 2020, 9, 90. [Google Scholar] [CrossRef]
- Chu, H.; Venevsky, S.; Wu, C.; Wang, M. NDVI-based vegetation dynamics and its response to climate changes at Amur-Heilongjiang River Basin from 1982 to 2015. Sci. Total Environ. 2019, 650, 2051–2062. [Google Scholar] [CrossRef]
- Zhang, F.; Su, Y.; Liu, B. Response characteristics of NDVI to asymmetric diurnal temperature increase and precipitation changes during the forest growing season in Northeast China. J. Beijing For. Univ. 2023, 45, 34–48. [Google Scholar] [CrossRef]
- Luo, X.; Li, Y.; Yin, H.; Sui, M. Response of NDVI to SPEI at different temporal scales in Northeast China. Chin. J. Ecol. 2020, 39, 412–421. [Google Scholar] [CrossRef]
- Ma, R.; Zhang, J.; Shen, X.; Liu, B.; Lu, X.; Jiang, M. Impacts of climate change on fractional vegetation coverage of temperate grasslands in China from 1982 to 2015. J. Environ. Manag. 2024, 350, 119694. [Google Scholar] [CrossRef]
- Cao, S.; Zhang, L.; He, Y.; Zhang, Y.; Chen, Y.; Yao, S.; Yang, W.; Sun, Q. Effects and contributions of meteorological drought on agricultural drought under different climatic zones and vegetation types in Northwest China. Sci. Total Environ. 2022, 821, 153270. [Google Scholar] [CrossRef]
- Shen, X.; Liu, B.; Henderson, M.; Wang, L.; Jiang, M.; Lu, X. Vegetation Greening, Extended Growing Seasons, and Temperature Feedbacks in Warming Temperate Grasslands of China. J. Clim. 2022, 35, 5103–5117. [Google Scholar] [CrossRef]
- Sun, S.; Du, W.; Song, Z.; Zhang, D.; Wu, X.; Chen, B.; Wu, Y. Response of Gross Primary Productivity to Drought Time-Scales Across China. J. Geophys. Res. Biogeosciences 2021, 126, e2020JG005953. [Google Scholar] [CrossRef]
- Zhang, Z.; Ju, W.; Zhou, Y.; Li, X. Revisiting the cumulative effects of drought on global gross primary productivity based on new long-term series data (1982–2018). Glob. Chang Biol. 2022, 28, 3620–3635. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Guo, E.; Yin, S.; Wang, Y.; Mandula, N.; Wan, Z.; Yun, X.; Li, H.; Bao, Y. Differentiating cumulative and lagged effects of drought on vegetation growth over the Mongolian Plateau. Ecosphere 2022, 13, e4289. [Google Scholar] [CrossRef]
- Zhao, G.; Han, Y.; Liu, M.; Zhao, S.; Liu, W. Characteristics of temporal and spatial variations of extreme temperature events in Inner Mongolia during 1951–2014. J. Arid Land Resour. Environ. 2017, 31, 110–116. [Google Scholar] [CrossRef]
- Ge, W.; Deng, L.; Wang, F.; Han, J. Quantifying the contributions of human activities and climate change to vegetation net primary productivity dynamics in China from 2001 to 2016. Sci. Total Environ. 2021, 773, 145648. [Google Scholar] [CrossRef]
- Zwicke, M.; Picon-Cochard, C.; Morvan-Bertrand, A.; Prud’homme, M.P.; Volaire, F. What functional strategies drive drought survival and recovery of perennial species from upland grassland? Ann. Bot. 2015, 116, 1001–1015. [Google Scholar] [CrossRef]
- Martínez-Vilalta, J.; Sala, A.; Asensio, D.; Galiano, L.; Hoch, G.; Palacio, S.; Piper, F.I.; Lloret, F. Dynamics of non-structural carbohydrates in terrestrial plants: A global synthesis. Ecol. Monogr. 2016, 86, 495–516. [Google Scholar] [CrossRef]
- Chapin, F.S.; Schulze, E.; Mooney, H.A. The Ecology and Economics of Storage in Plants. Annu. Rev. Ecol. Evol. Syst. 1990, 21, 423–447. [Google Scholar] [CrossRef]
- Piispanen, R.; Saranp, P. Variation of non-structural carbohydrates in silver birch (Betula pendulaRoth) wood. Trees 2001, 15, 444–451. [Google Scholar] [CrossRef]
- Sagar, G.R.; Salter, P.J.; Goode, J.E. Crop Responses to Water at Different Stages of Growth. J. Ecol. 1968, 56, 626. [Google Scholar] [CrossRef]
- Sullivan, C.Y.; Eastin, J.D. Plant physiological responses to water stress. Agric. Meteorol. 1974, 14, 113–127. [Google Scholar] [CrossRef]
- Fang, W.; Huang, S.; Huang, Q.; Huang, G.; Wang, H.; Leng, G.; Wang, L.; Guo, Y. Probabilistic assessment of remote sensing-based terrestrial vegetation vulnerability to drought stress of the Loess Plateau in China. Remote Sens. Environ. 2019, 232, 111290. [Google Scholar] [CrossRef]
- You, C.; Wang, Y.; Tan, X.; Zhang, B.; Ren, T.; Chen, B.; Xu, M.; Chen, S. Seasonal and interannual variations of ecosystem photosynthetic characteristics in a semi-arid grassland of Northern China. J. Plant Ecol. 2022, 15, 961–976. [Google Scholar] [CrossRef]
- Peng, F.; Sun, G. Variation of Leaf Area Index in China from 1982 to 1999 and Its Relationship with Climate Change. Clim. Environ. Res. 2017, 22, 162–176. [Google Scholar]
- Knapp, A.K.; Smith, M.D. Variation Among Biomes in Temporal Dynamics of Aboveground Primary Production. Science 2001, 291, 481–484. [Google Scholar] [CrossRef] [PubMed]
- Sippel, S.; Reichstein, M.; Ma, X.; Mahecha, M.D.; Lange, H.; Flach, M.; Frank, D. Drought, Heat, and the Carbon Cycle: A Review. Curr. Clim. Change Rep. 2018, 4, 266–286. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, H.; Wang, J.; Shi, H.; Liu, H.; He, J. Estimation on seasonal dynamics of alpine grassland aboveground biomass using phenology camera-derived NDVI. Chin. J. Plant Ecol. 2021, 45, 487–495. [Google Scholar] [CrossRef]
- Zhou, R.; Liu, Y.; Cui, M.; Lu, J.; Shi, H.; Ren, H.; Zhang, W.; Wen, Z. Global Assessment of Cumulative and Time-Lag Effects of Drought on Land Surface Phenology. GIScience Remote Sens. 2022, 59, 1918–1937. [Google Scholar] [CrossRef]
- Peguero-Pina, J.J.; Vilagrosa, A.; Alonso-Forn, D.; Ferrio, J.P.; Sancho-Knapik, D.; Gil-Pelegrín, E. Living in Drylands: Functional Adaptations of Trees and Shrubs to Cope with High Temperatures and Water Scarcity. Forests 2020, 11, 1028. [Google Scholar] [CrossRef]
- Wei, D.; Zhao, H.; Zhang, J.; Qi, Y.; Wang, X. Human activities alter response of alpine grasslands on Tibetan Plateau to climate change. J. Environ. Manag. 2020, 262, 110335. [Google Scholar] [CrossRef]
- Xue, L.; Kappas, M.; Wyss, D.; Wang, C.; Putzenlechner, B.; Thi, N.P.; Chen, J. Assessment of Climate Change and Human Activities on Vegetation Development in Northeast China. Sensors 2022, 22, 2509. [Google Scholar] [CrossRef]
- Xu, H.-j.; Wang, X.-p.; Zhao, C.-y. Drought sensitivity of vegetation photosynthesis along the aridity gradient in northern China. Int. J. Appl. Earth Obs. Geoinf. 2021, 102, 102418. [Google Scholar] [CrossRef]
- Barnes, M.L.; Moran, M.S.; Scott, R.L.; Kolb, T.E.; Ponce-Campos, G.E.; Moore, D.J.P.; Ross, M.A.; Mitra, B.; Dore, S. Vegetation productivity responds to sub-annual climate conditions across semiarid biomes. Ecosphere 2016, 7, e01339. [Google Scholar] [CrossRef]
- Tong, S.; Bao, G.; Bao, Y.; Huang, X. Monitoring of long-term vegetation dynamics and responses to droughts of various timescales in Inner Mongolia. Ecosphere 2023, 14, e4415. [Google Scholar] [CrossRef]
- Busetto, L.; Colombo, R.; Migliavacca, M.; Cremonese, E.; Meroni, M.; Galvagno, M.; Rossini, M.; Siniscalco, C.; Di Cella, U.M.; Pari, E. Remote sensing of larch phenological cycle and analysis of relationships with climate in the Alpine region. Glob. Change Biol. 2010, 16, 2504–2517. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, S.; Regnier, P.; Yuan, W. New insights on plant phenological response to temperature revealed from long-term widespread observations in China. Glob. Change Biol. 2018, 24, 2066–2078. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Season | 0–0.15 | 0.15–0.3 | 0.3–0.45 | 0.45–0.6 | >0.6 | Mean |
|---|---|---|---|---|---|---|
| Growing season | 6.75% | 37.44% | 19.54% | 0.06% | 0.00% | 0.262 |
| Spring | 3.34% | 34.48% | 28.86% | 7.71% | 0.54% | 0.309 |
| Summer | 5.36% | 16.78% | 24.07% | 31.40% | 12.63% | 0.424 |
| Autumn | 4.26% | 25.68% | 21.83% | 8.16% | 0.83% | 0.314 |
| Season | 0–0.15 | 0.15–0.3 | 0.3–0.45 | 0.45–0.6 | >0.6 | Mean |
|---|---|---|---|---|---|---|
| Growing season | 4.71% | 32.91% | 25.91% | 0.27% | 0.01% | 0.256 |
| Spring | 0.06% | 17.81% | 38.73% | 6.78% | 0.09% | 0.349 |
| Summer | 0.43% | 28.90% | 36.77% | 11.97% | 0.51% | 0.341 |
| Autumn | 0.09% | 21.22% | 35.03% | 5.63% | 0.18% | 0.338 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, W.; Henderson, M.; Liu, B.; Su, Y.; Zhou, W.; Ma, R.; Chen, M.; Zhang, Z. Cumulative and Lagged Effects: Seasonal Characteristics of Drought Effects on East Asian Grasslands. Remote Sens. 2024, 16, 3478. https://doi.org/10.3390/rs16183478
Huang W, Henderson M, Liu B, Su Y, Zhou W, Ma R, Chen M, Zhang Z. Cumulative and Lagged Effects: Seasonal Characteristics of Drought Effects on East Asian Grasslands. Remote Sensing. 2024; 16(18):3478. https://doi.org/10.3390/rs16183478
Chicago/Turabian StyleHuang, Weiwei, Mark Henderson, Binhui Liu, Yuanhang Su, Wanying Zhou, Rong Ma, Mingyang Chen, and Zhi Zhang. 2024. "Cumulative and Lagged Effects: Seasonal Characteristics of Drought Effects on East Asian Grasslands" Remote Sensing 16, no. 18: 3478. https://doi.org/10.3390/rs16183478
APA StyleHuang, W., Henderson, M., Liu, B., Su, Y., Zhou, W., Ma, R., Chen, M., & Zhang, Z. (2024). Cumulative and Lagged Effects: Seasonal Characteristics of Drought Effects on East Asian Grasslands. Remote Sensing, 16(18), 3478. https://doi.org/10.3390/rs16183478