
Evaluation of Original and Water Stress-Incorporated Modified Weather Research and Forecasting Vegetation Photosynthesis and Respiration Model in Simulating CO2 Flux and Concentration Variability over the Tibetan Plateau

 , , ,
, , ,
Abstract
:1. Introduction
2. Methods
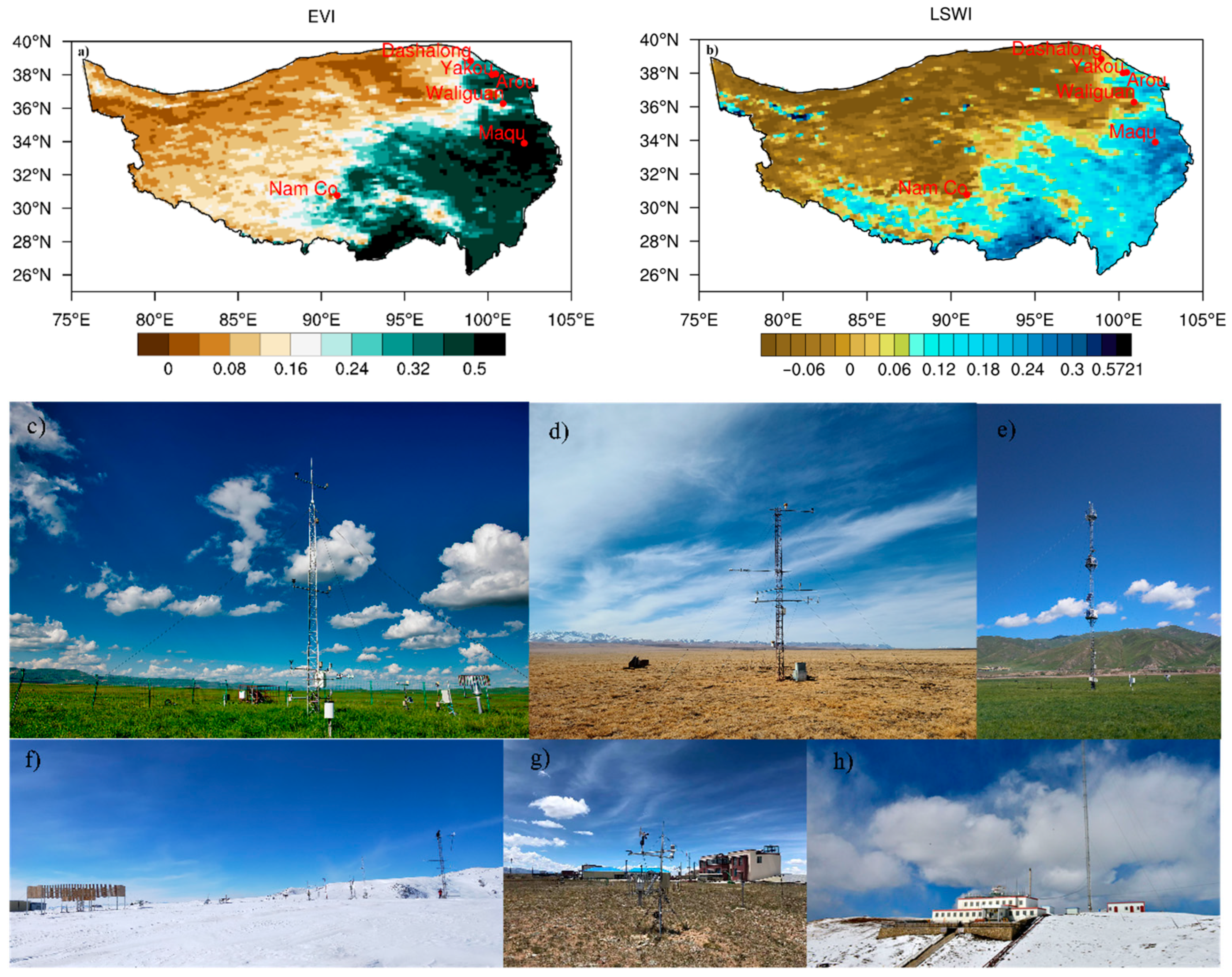
2.1. In Situ and Satellite Observations
2.2. Description of the WRF-VPRM
2.3. Analytical Method
3. Results
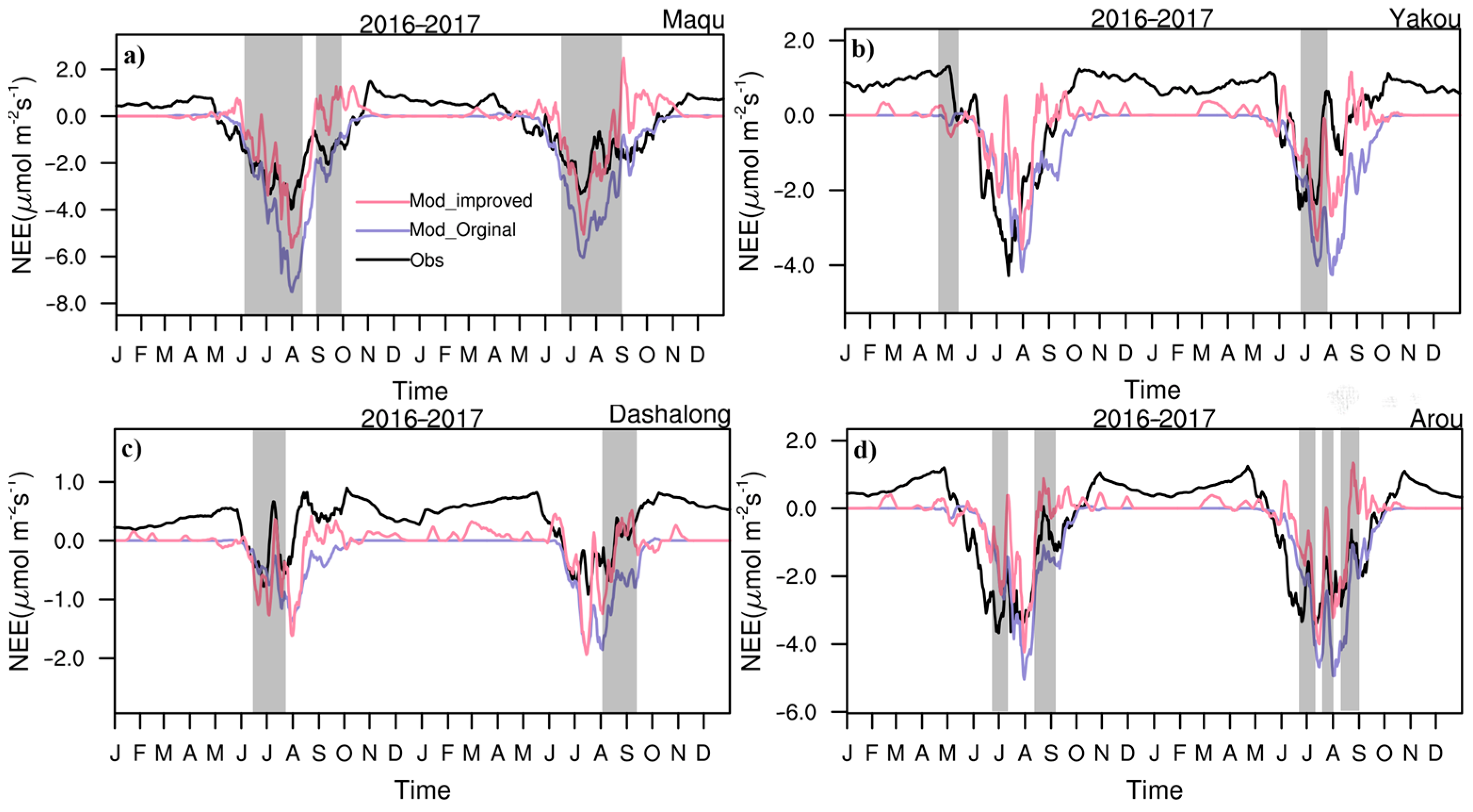
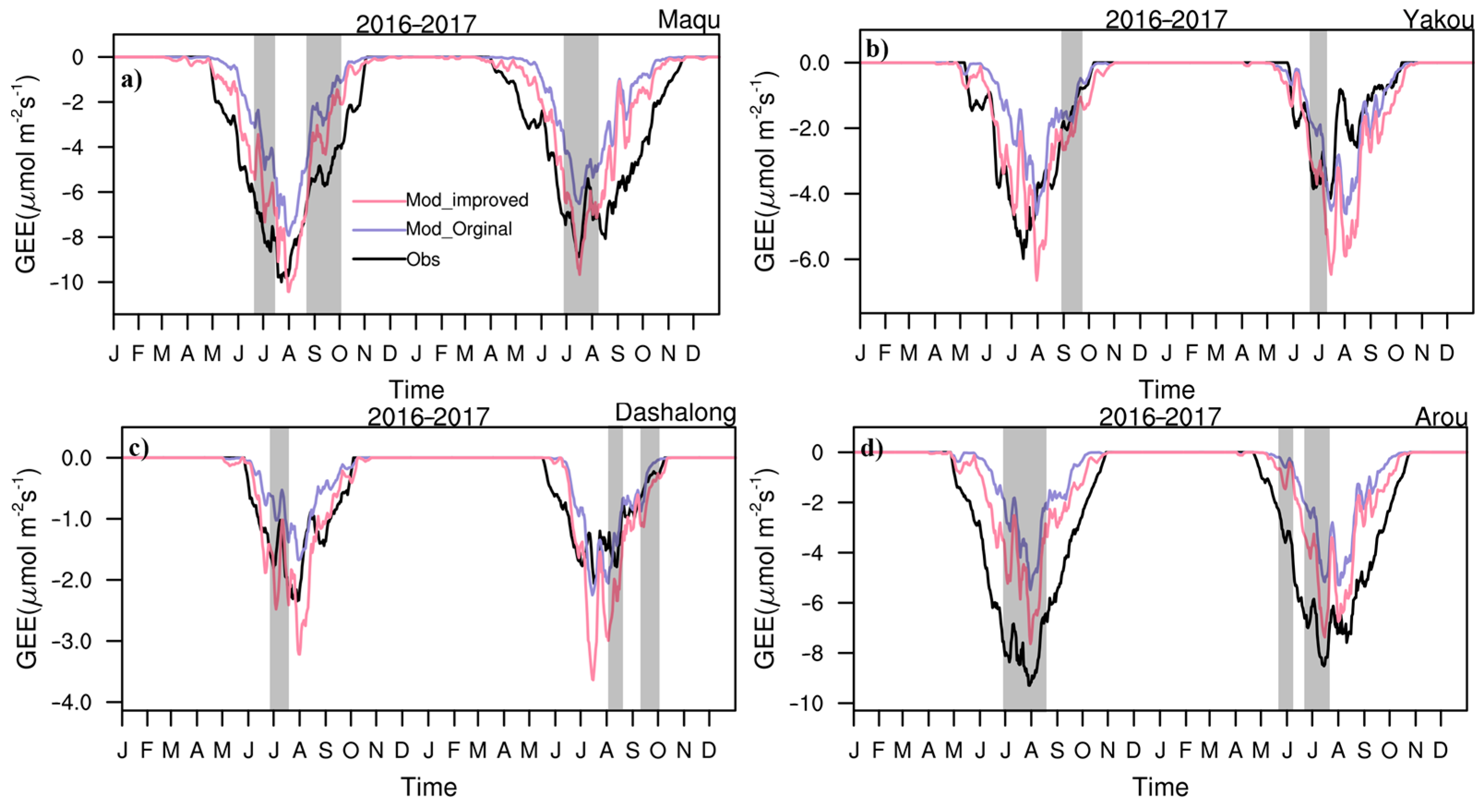
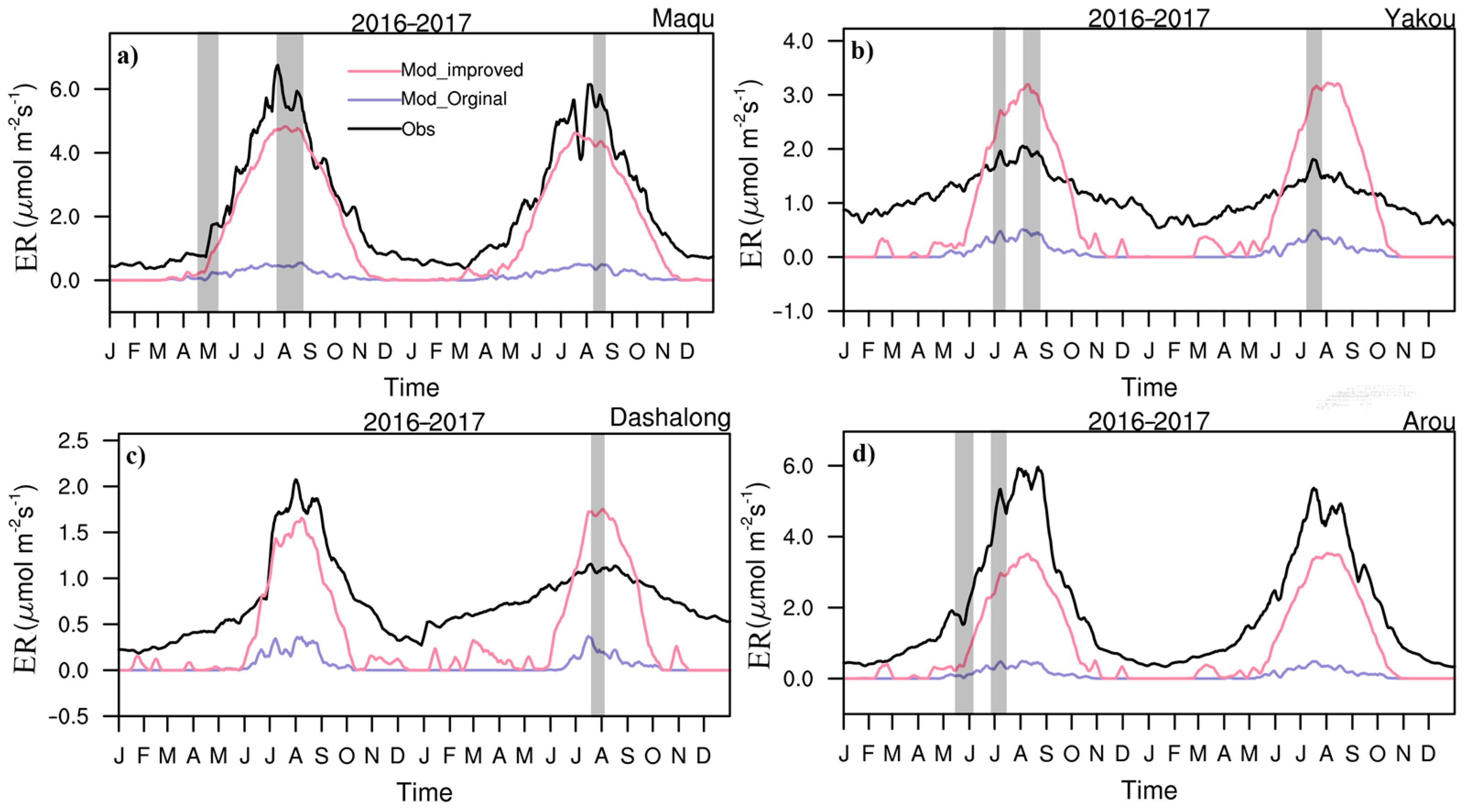
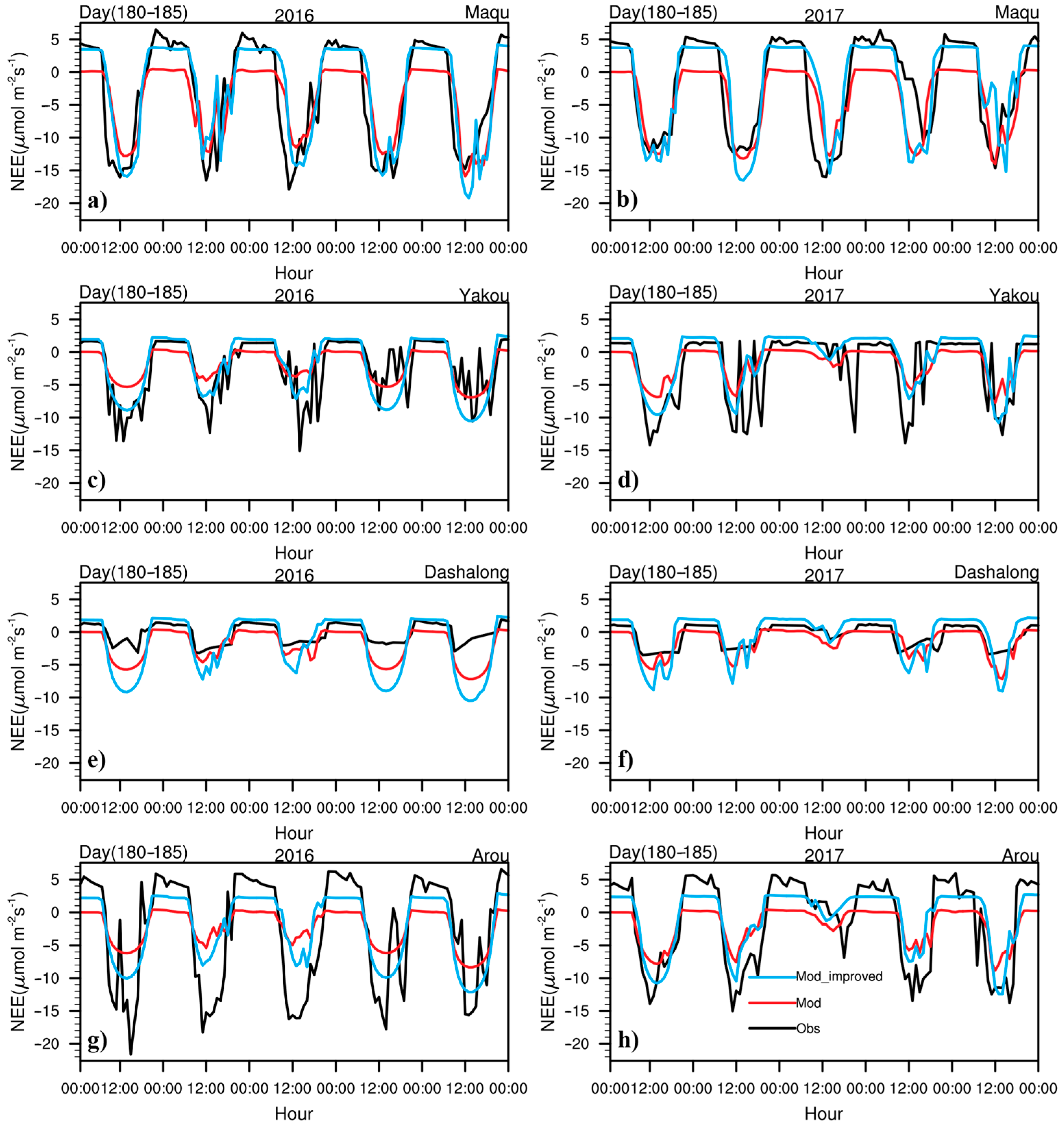
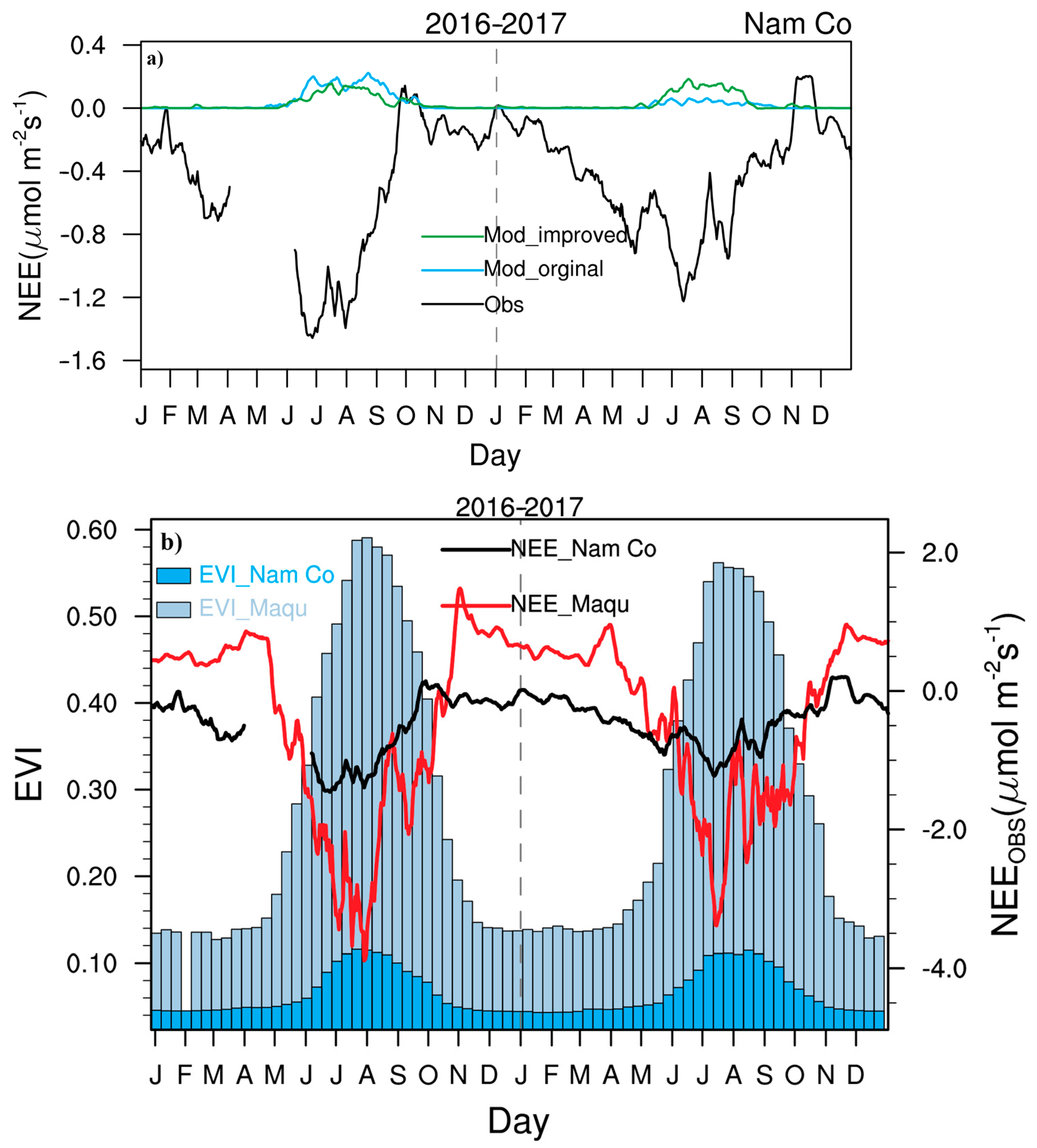
3.1. Seasonal Variations in CO2 Fluxes and Hourly Variations in NEE and CO2 Concentrations
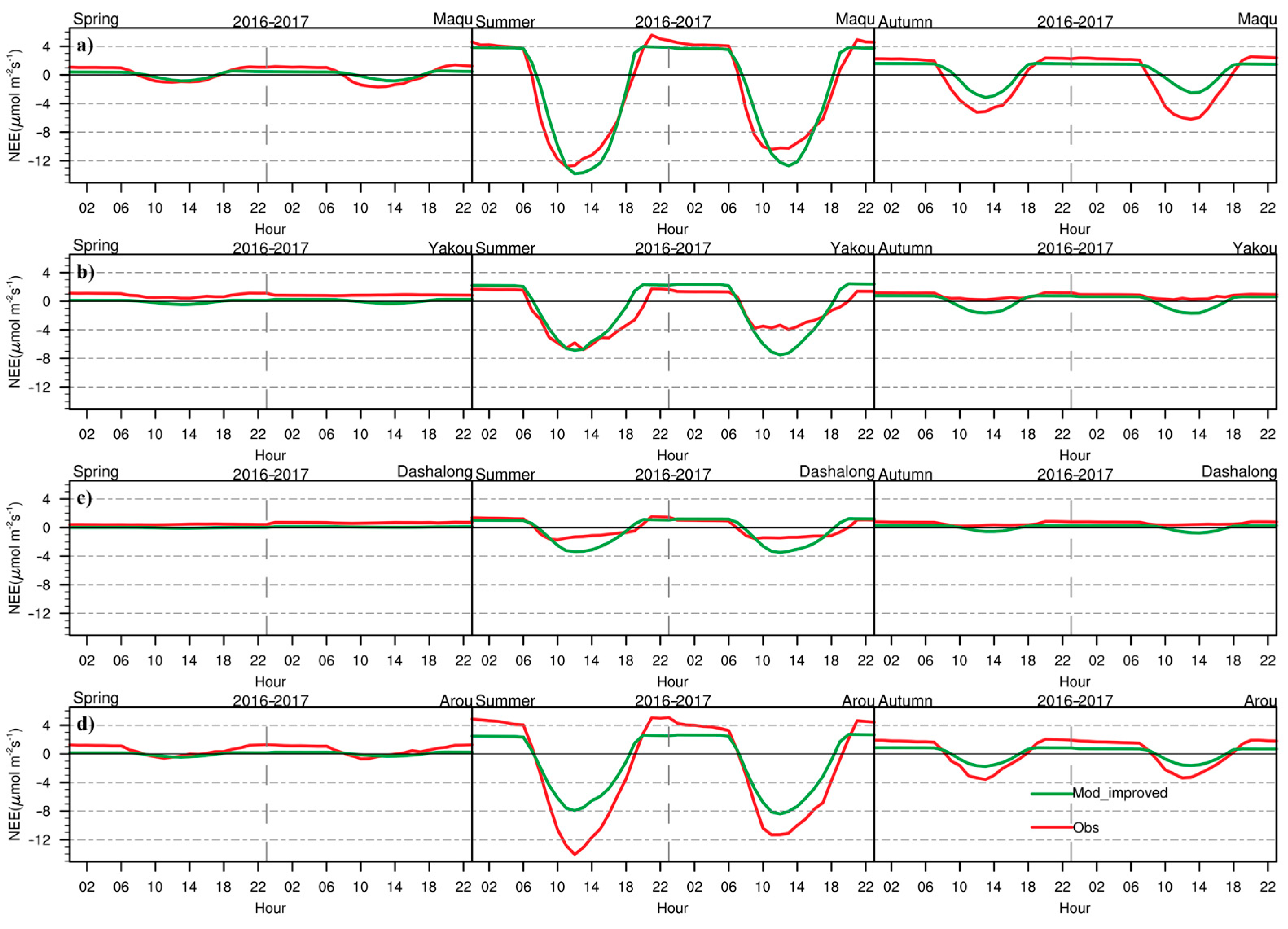
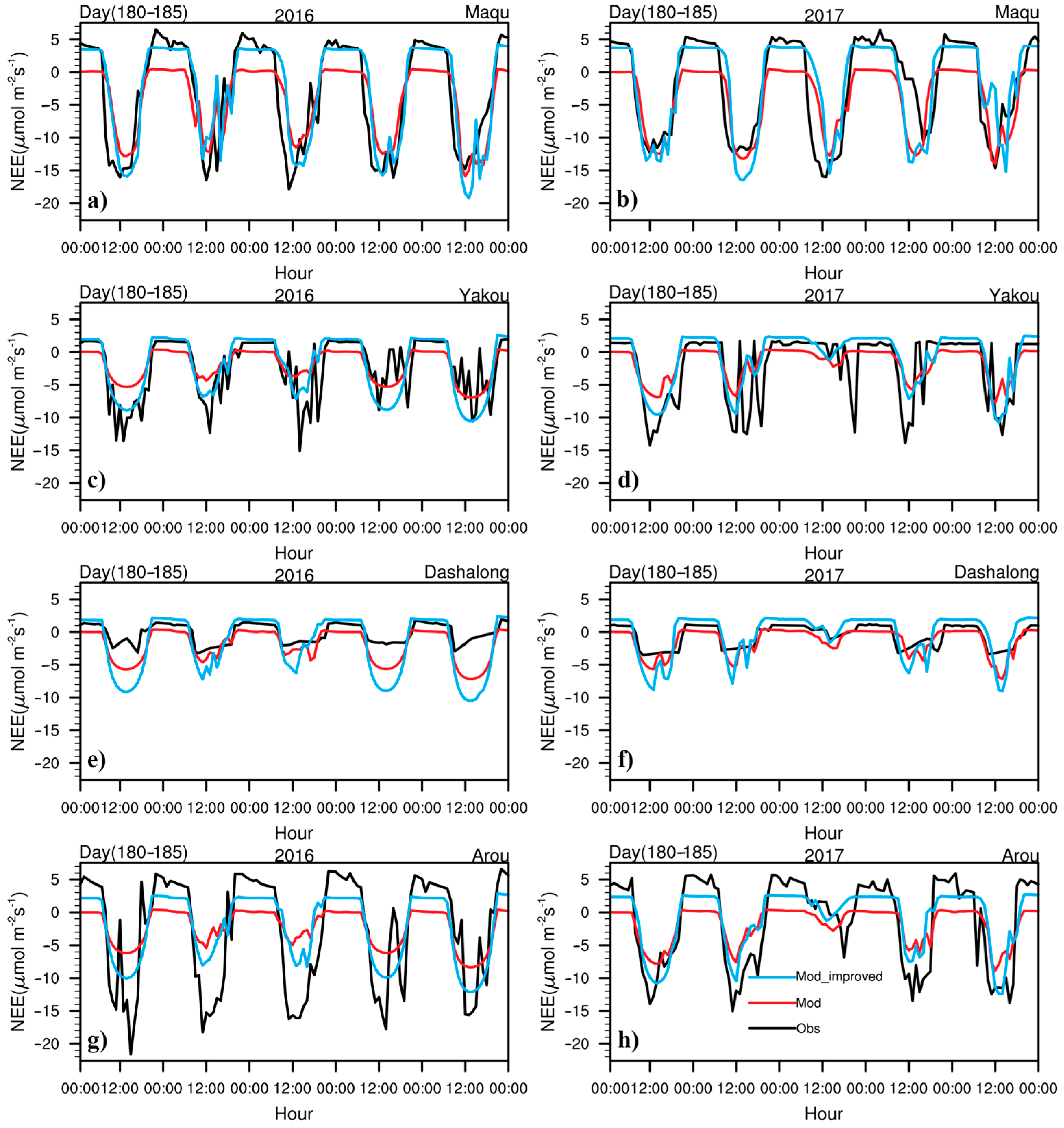
3.2. Daily Variations in CO2 Fluxes during the Growing Season
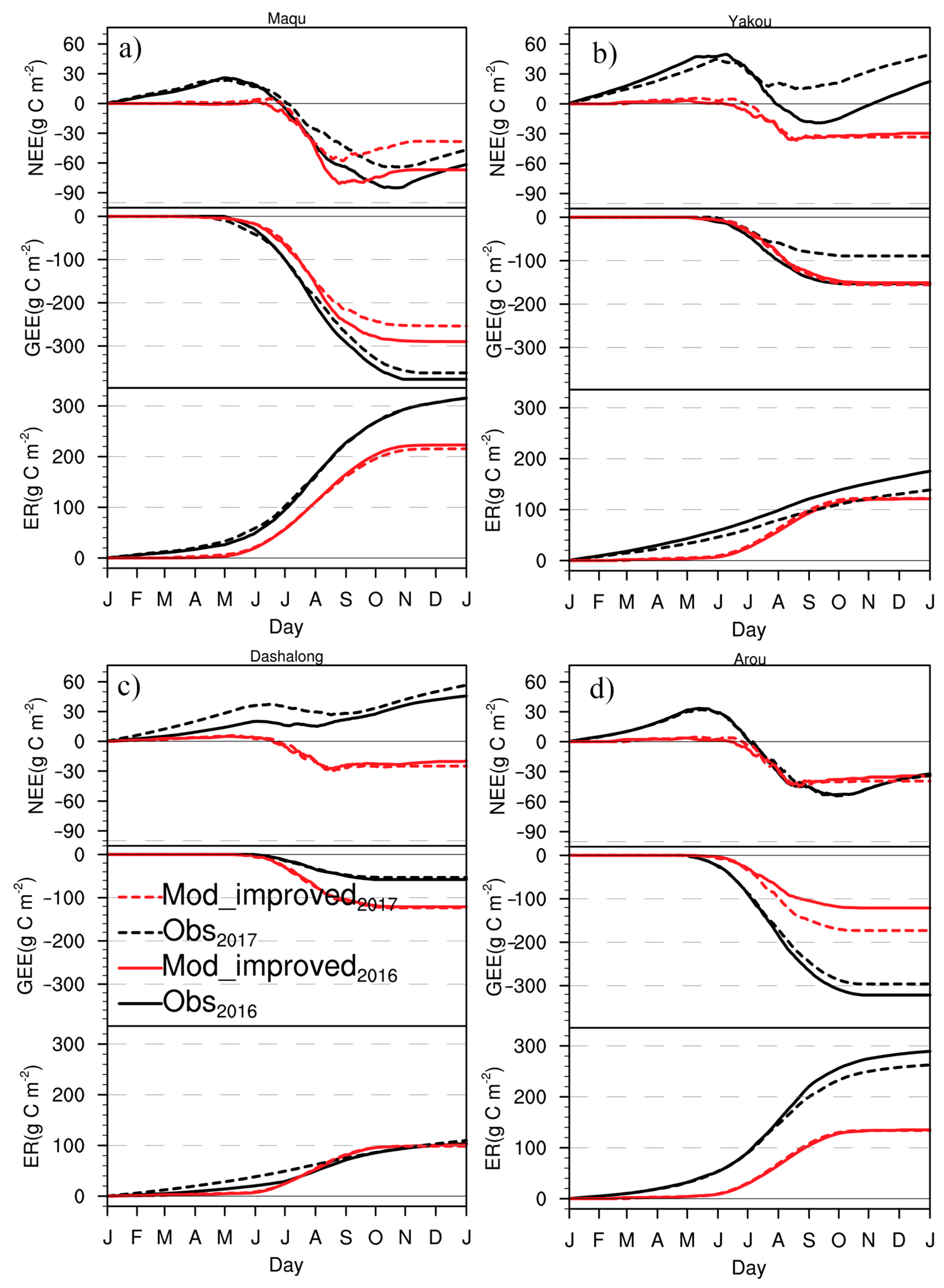
3.3. Cumulative CO2 Fluxes
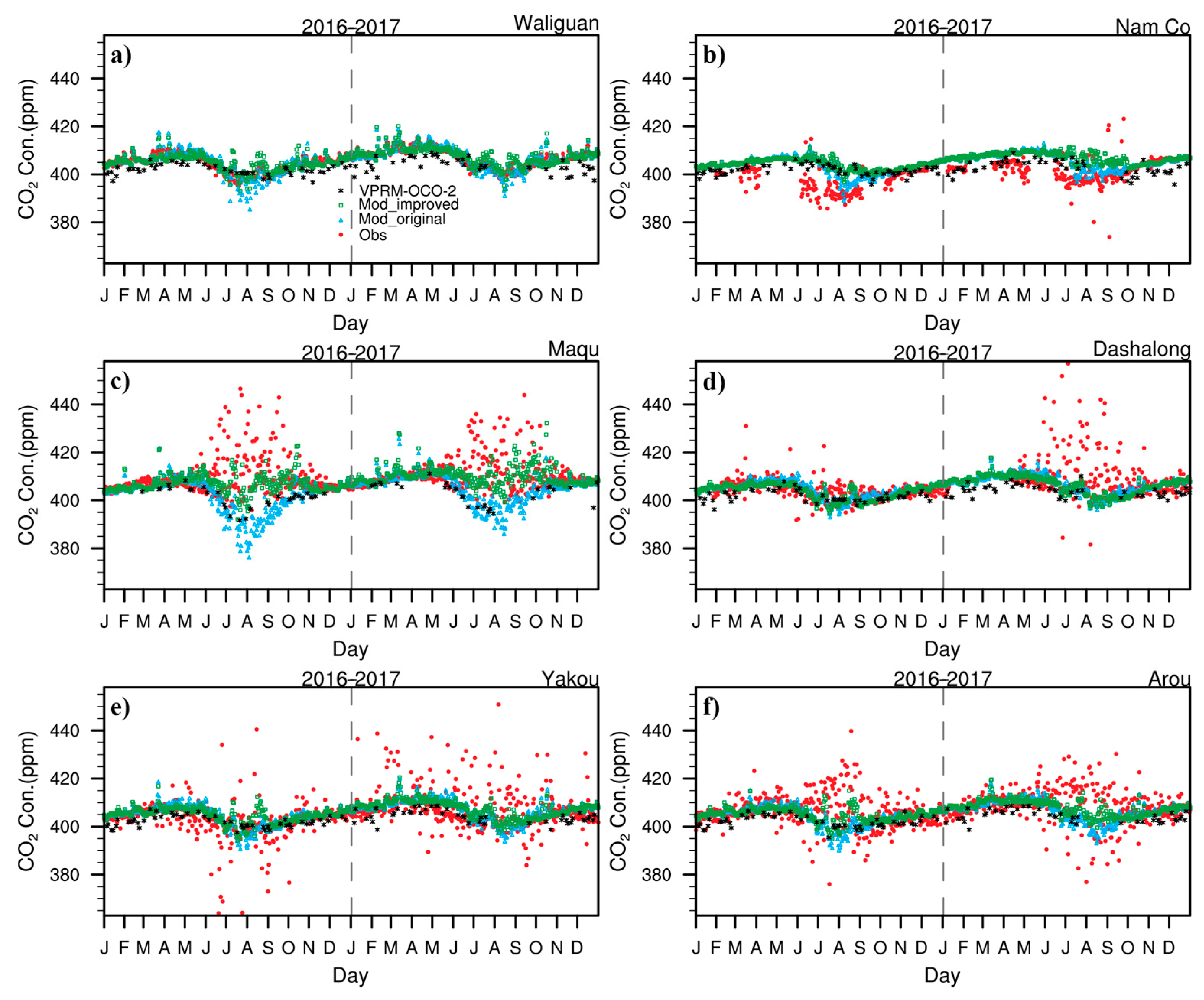
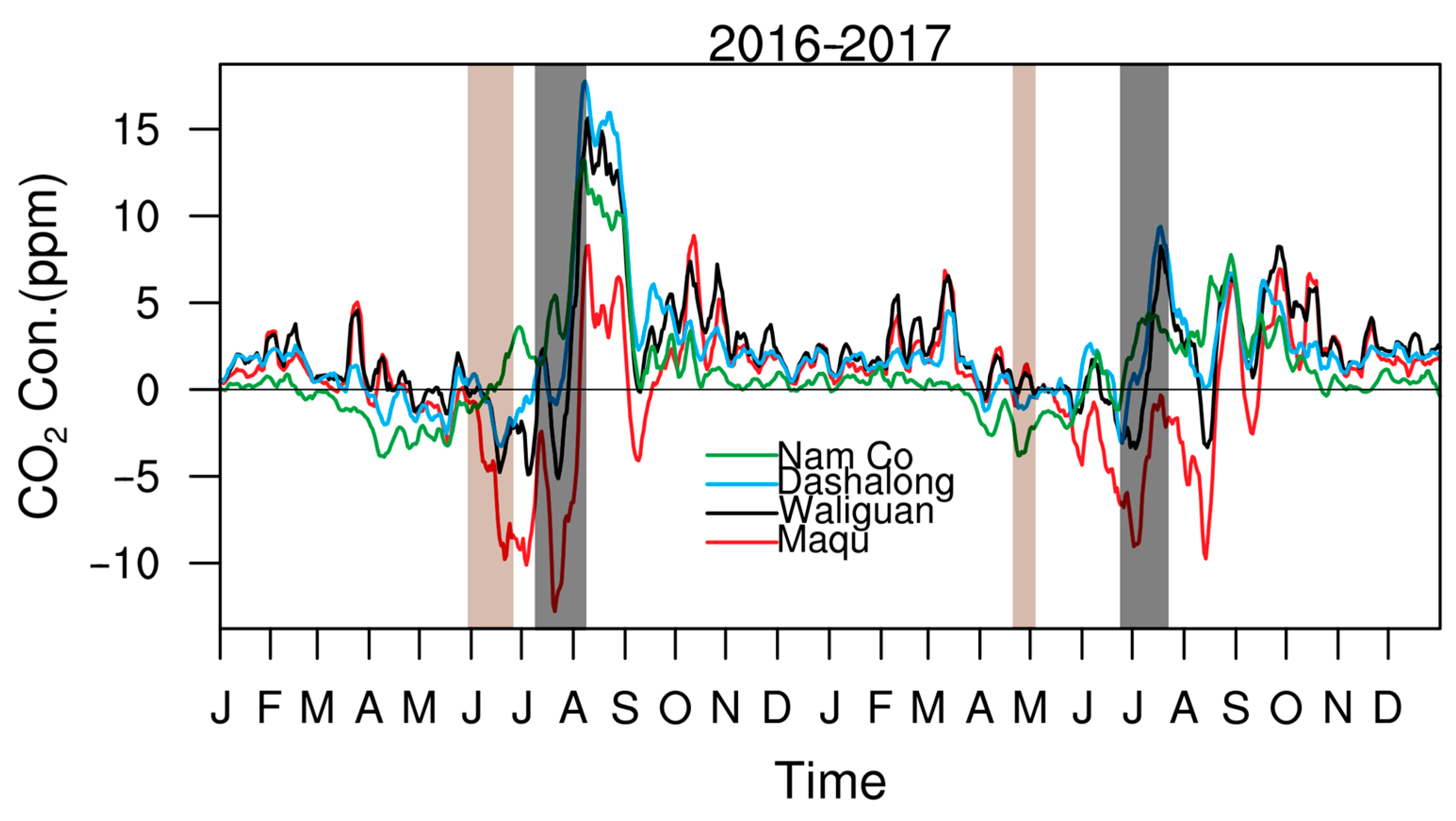
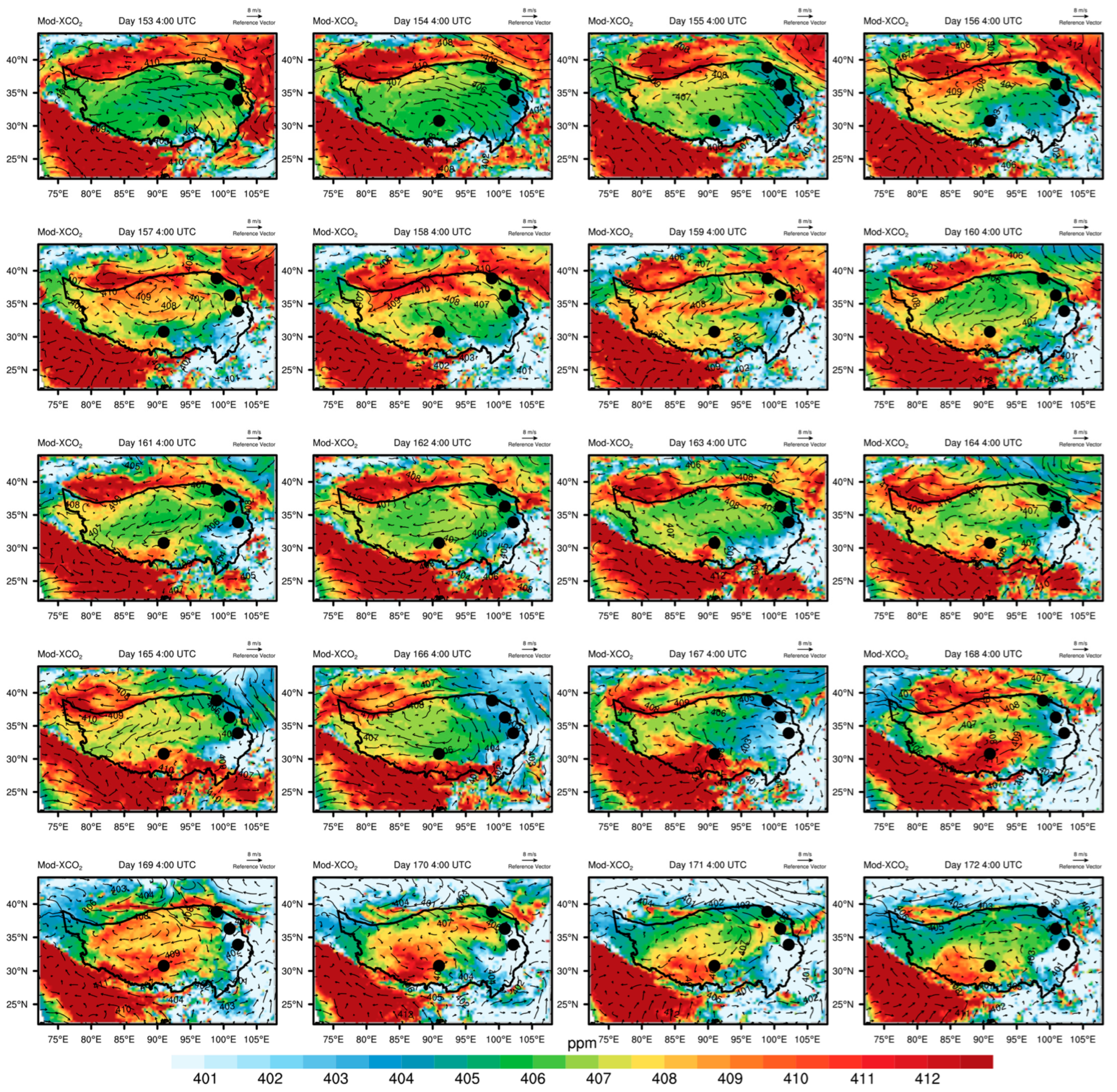
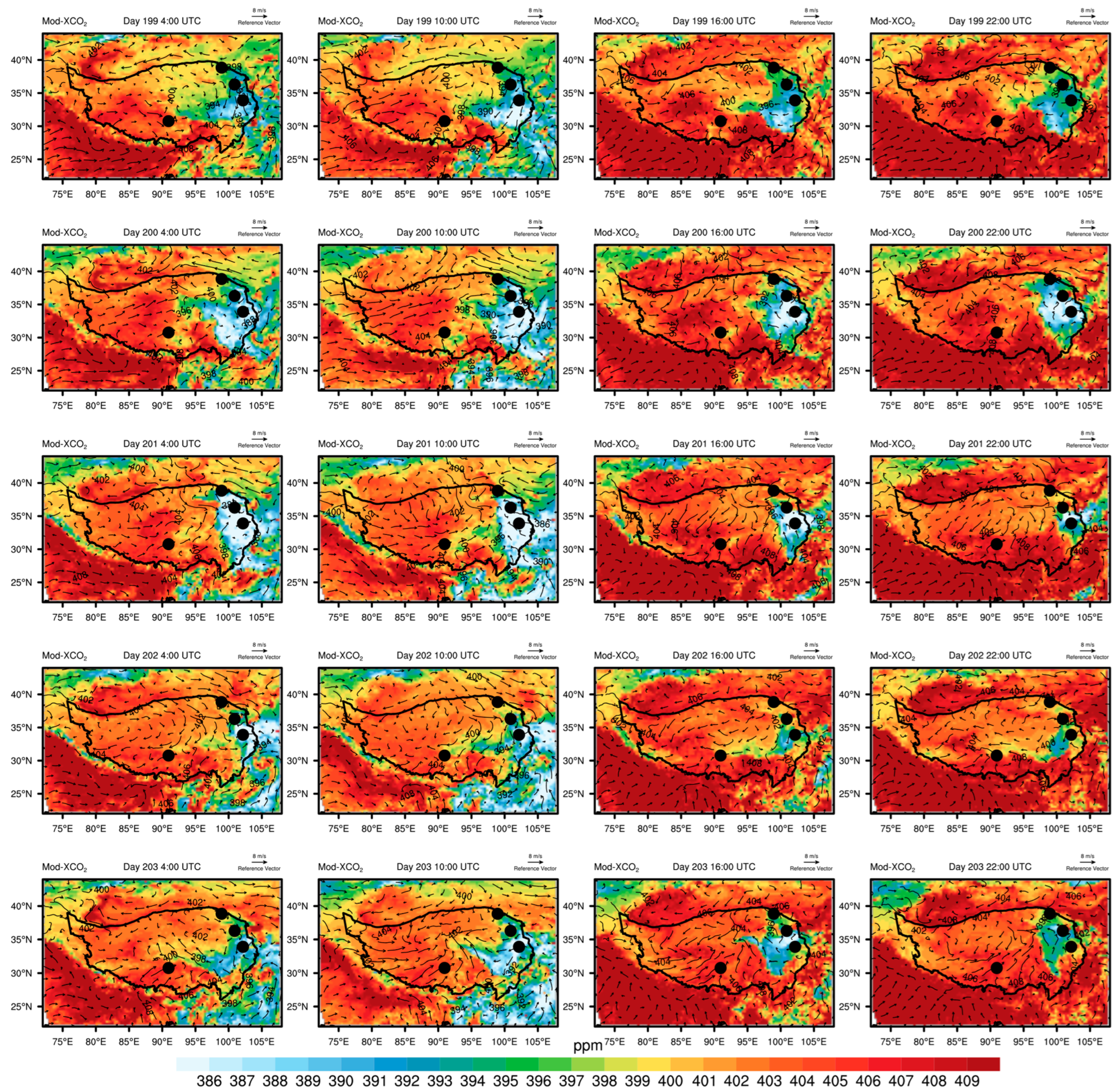
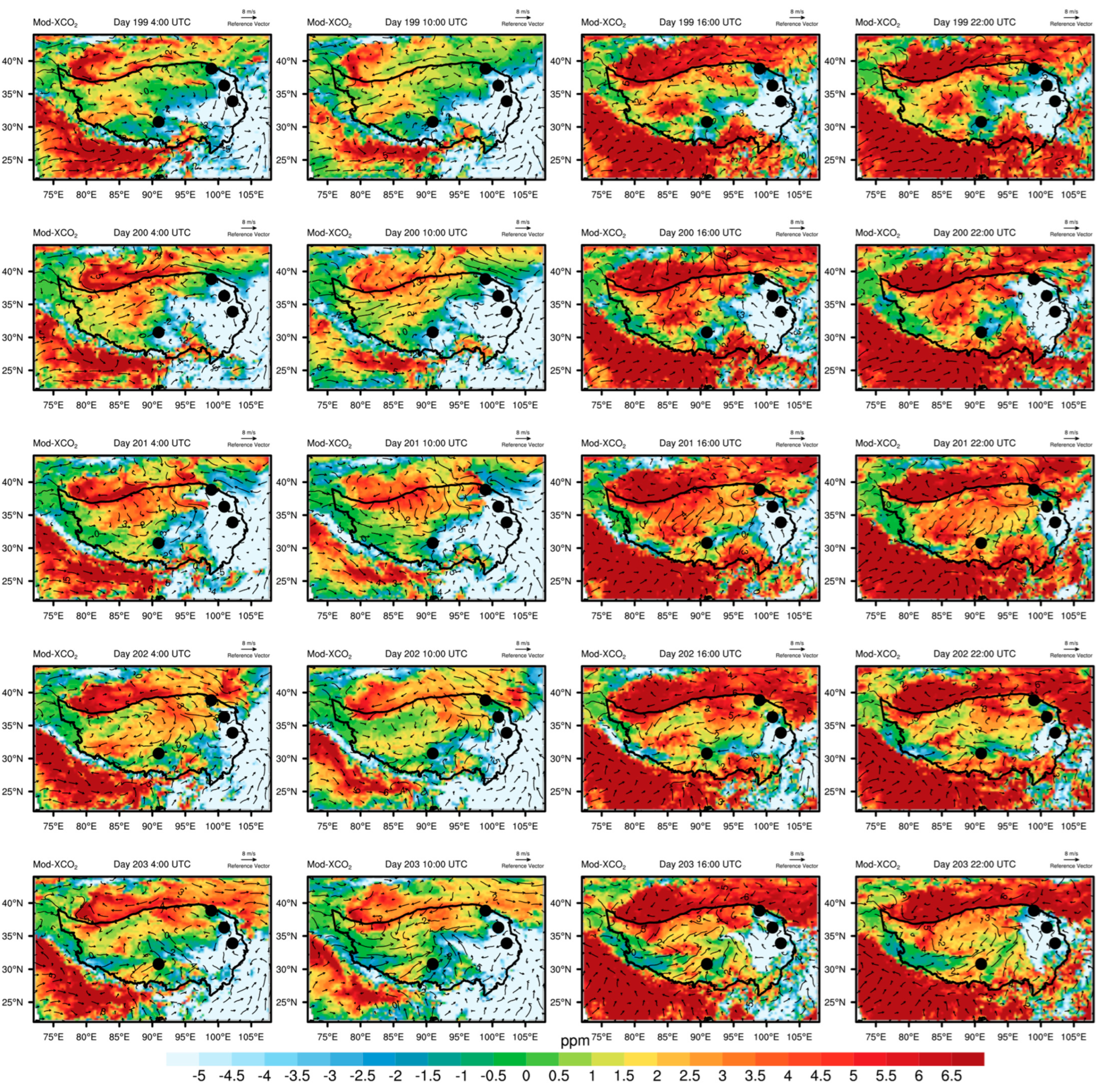
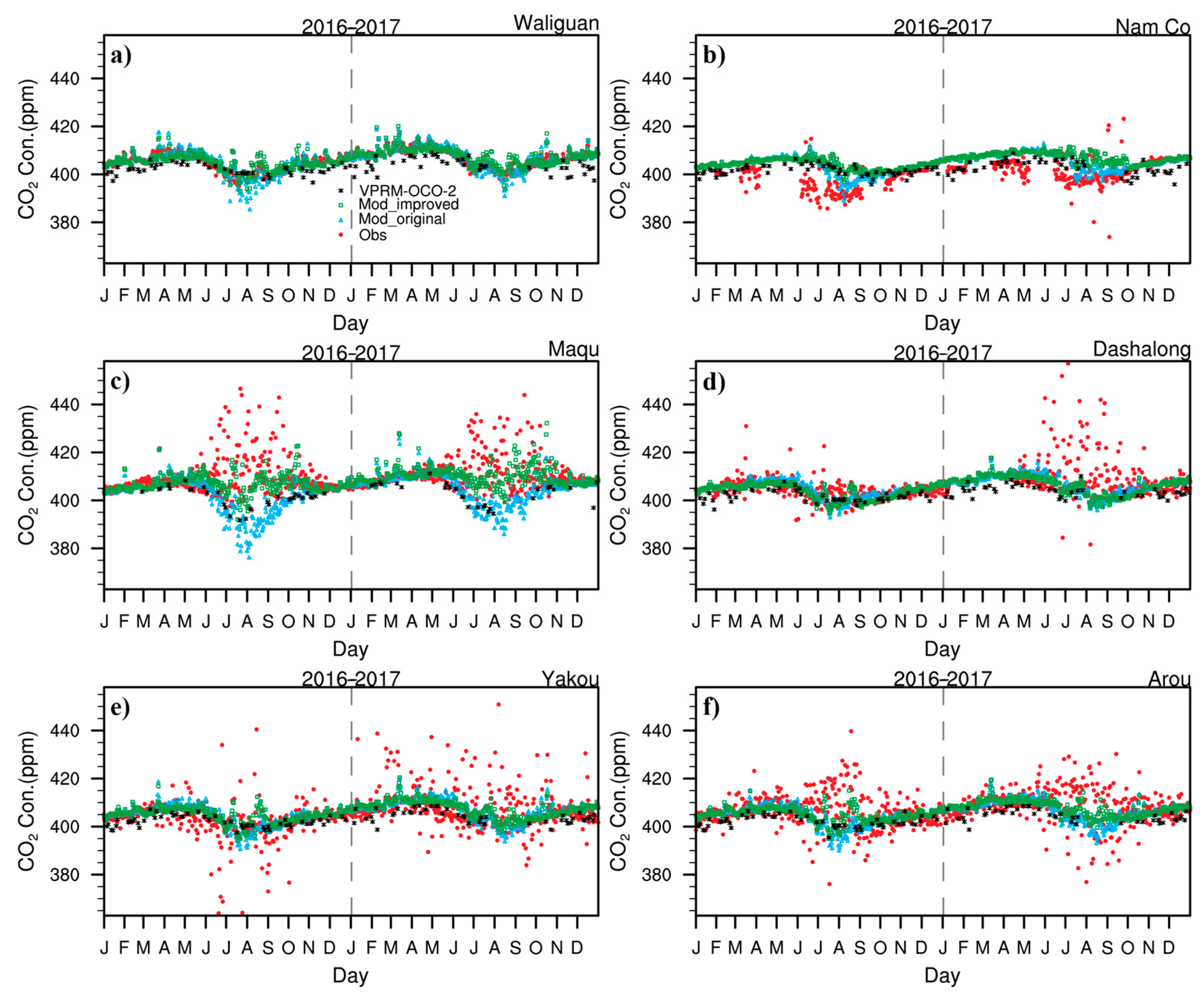
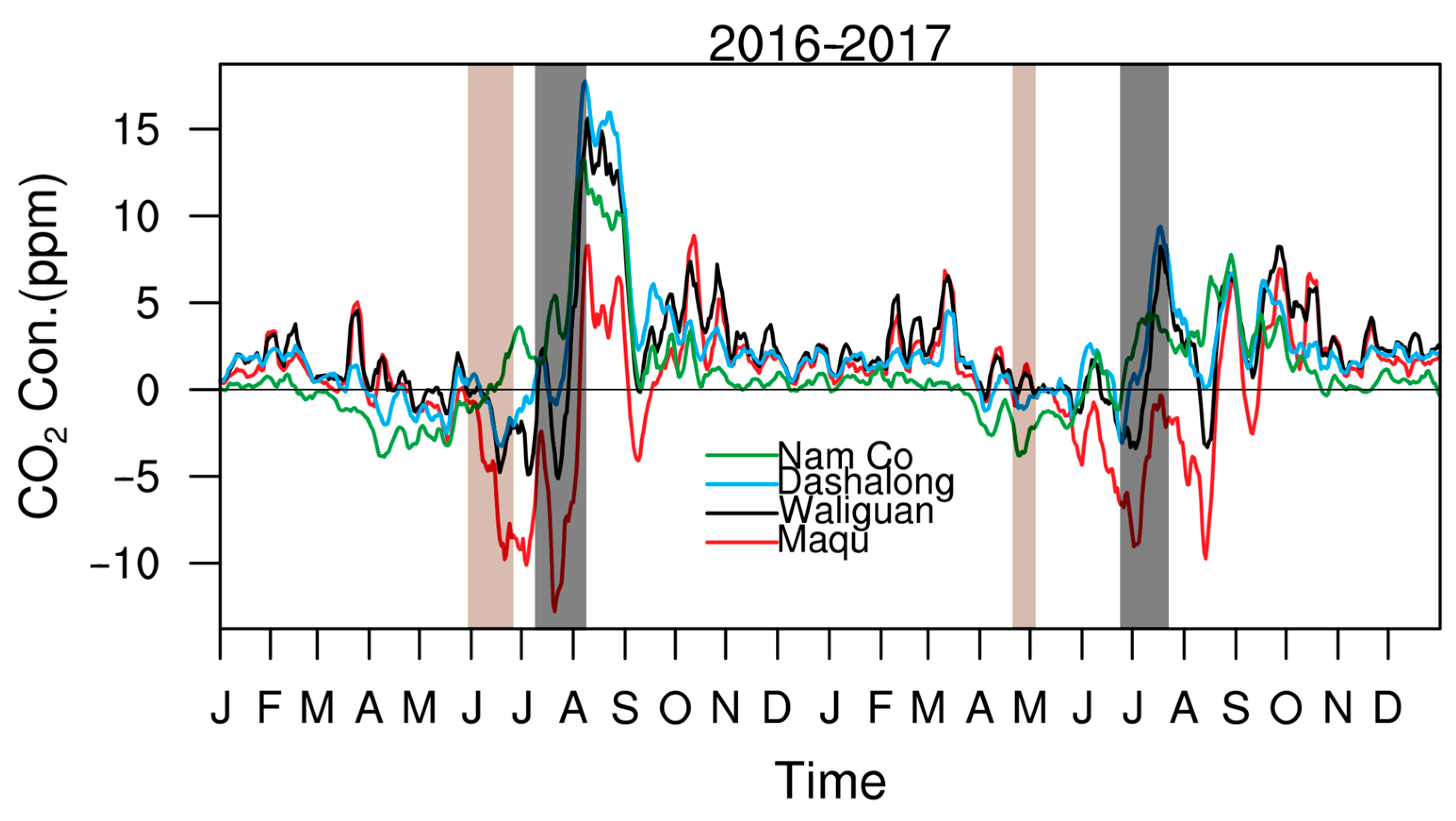
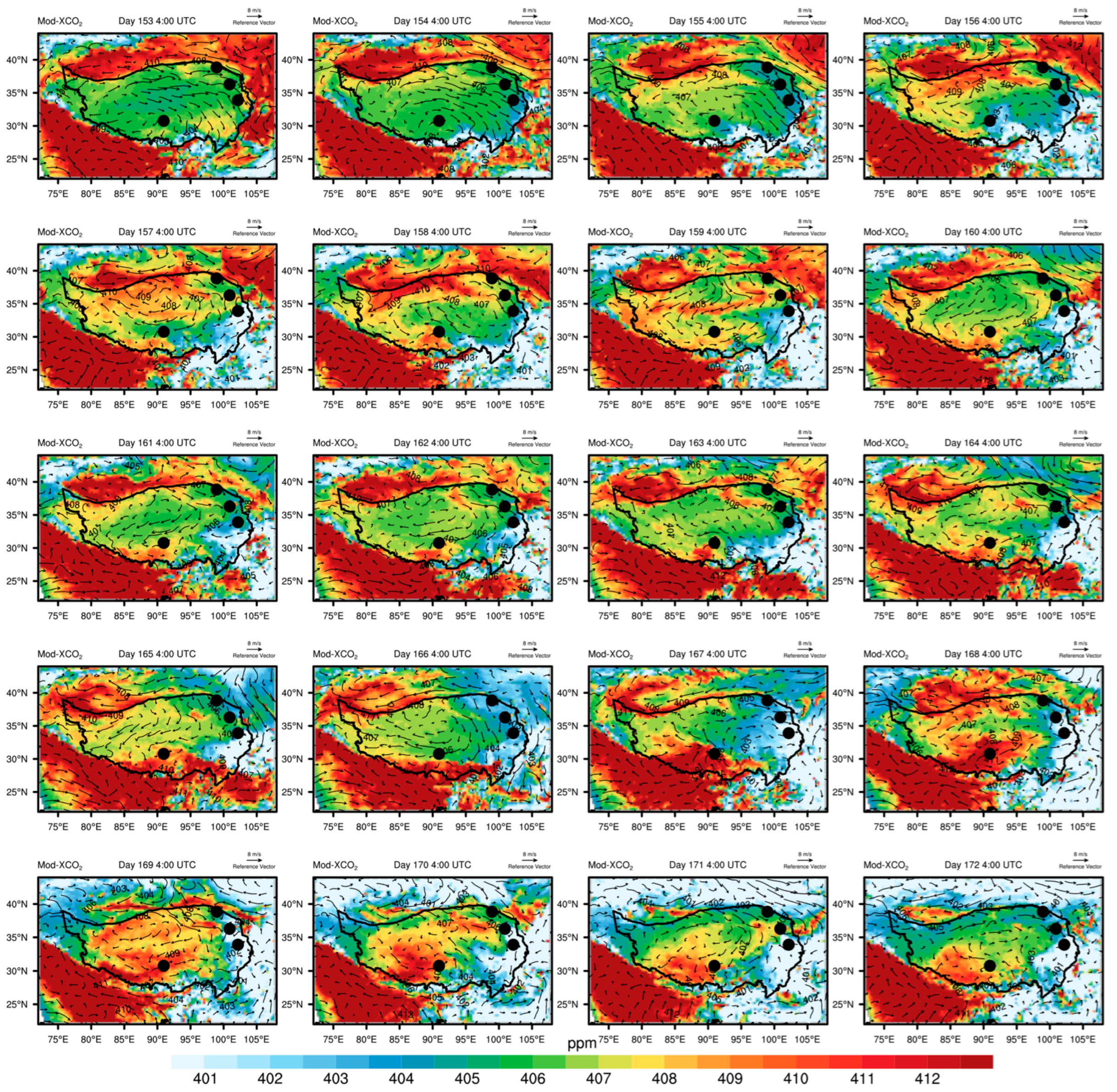
3.4. Irregular Variations in CO2 Concentrations
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Piao, S.; Fang, J.; Ciais, P.; Peylin, P.; Huang, Y.; Sitch, S.; Wang, T. The carbon balance of terrestrial ecosystems in China. Nature 2009, 458, 1009–1013. [Google Scholar] [CrossRef] [PubMed]
- Cox, P.M.; Betts, R.A.; Jones, C.D.; Spall, S.A.; Totterdell, I.J. Acceleration of global warming due to carbon-cycle feedbacks in a coupled climate model. Nature 2000, 408, 184–187. [Google Scholar] [CrossRef]
- Friedlingstein, P.; Cox, P.; Betts, R.; Bopp, L.; von Bloh, W.; Brovkin, V.; Cadule, P.; Doney, S.; Eby, M.; Fung, I.; et al. Climate–Carbon Cycle Feedback Analysis: Results from the C4MIP Model Intercomparison. J. Clim. 2006, 19, 3337–3353. [Google Scholar] [CrossRef]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G. A large and persistent carbon sink in the world’s forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [PubMed]
- Palmer, P.I.; Feng, L.; Baker, D.; Chevallier, F.; Bosch, H.; Somkuti, P. Net carbon emissions from African biosphere dominate pan-tropical atmospheric CO2 signal. Nat. Commun. 2019, 10, 3344. [Google Scholar] [CrossRef]
- Baldocchi, D.; Falge, E.; Gu, L.H.; Olson, R.; Hollinger, D.; Running, S.; Anthoni, P.; Bernhofer, C.; Davis, K.; Evans, R.; et al. FLUXNET: A new tool to study the temporal and spatial variability of ecosystem-scale carbon dioxide, water vapor, and energy flux densities. Bull. Am. Meteorol. Soc. 2001, 82, 2415–2434. [Google Scholar] [CrossRef]
- Su, Y.-g.; Wu, L.; Zhang, Y.-m. Characteristics of carbon flux in two biologically crusted soils in the Gurbantunggut Desert, Northwestern China. Catena 2012, 96, 41–48. [Google Scholar] [CrossRef]
- Wang, H.; Li, X.; Xiao, J.; Ma, M.; Tan, J.; Wang, X.; Geng, L. Carbon fluxes across alpine, oasis, and desert ecosystems in northwestern China: The importance of water availability. Sci. Total Environ. 2019, 697, 133978. [Google Scholar] [CrossRef]
- Running, S.W.; Nemani, R.R.; Heinsch, F.A.; Zhao, M.S.; Reeves, M.; Hashimoto, H. A continuous satellite-derived measure of global terrestrial primary production. Bioscience 2004, 54, 547–560. [Google Scholar] [CrossRef]
- Keenan, T.F.; Gray, J.; Friedl, M.A.; Toomey, M.; Bohrer, G.; Hollinger, D.Y.; Munger, J.W.; O’Keefe, J.; Schmid, H.P.; Wing, I.S.; et al. Net carbon uptake has increased through warming-induced changes in temperate forest phenology. Nat. Clim. Chang. 2014, 4, 598–604. [Google Scholar] [CrossRef]
- van der Velde, I.R.; van der Werf, G.R.; Houweling, S.; Maasakkers, J.D.; Borsdorff, T.; Landgraf, J.; Tol, P.; van Kempen, T.A.; van Hees, R.; Hoogeveen, R.; et al. Vast CO2 release from Australian fires in 2019-2020 constrained by satellite. Nature 2021, 597, 366–369. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.X.; Lauvaux, T.; Kort, E.A.; Oda, T.; Feng, S.; Lin, J.C.; Yang, E.G.; Wu, D. Constraining Fossil Fuel CO2 Emissions From Urban Area Using OCO-2 Observations of Total Column CO2. J. Geophys. Res.-Atmos. 2020, 125, e2019JD030528. [Google Scholar] [CrossRef]
- Labzovskii, L.D.; Jeong, S.J.; Parazoo, N.C. Working towards confident spaceborne monitoring of carbon emissions from cities using Orbiting Carbon Observatory-2. Remote Sens. Environ. 2019, 233, 111359. [Google Scholar] [CrossRef]
- Fu, P.; Xie, Y.H.; Moore, C.E.; Myint, S.W.; Bernacchi, C.J. A Comparative Analysis of Anthropogenic CO2 Emissions at City Level Using OCO-2 Observations: A Global Perspective. Earths Future 2019, 7, 1058–1070. [Google Scholar] [CrossRef]
- Ciais, P.; Reichstein, M.; Viovy, N.; Granier, A.; Ogee, J.; Allard, V.; Aubinet, M.; Buchmann, N.; Bernhofer, C.; Carrara, A.; et al. Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature 2005, 437, 529–533. [Google Scholar] [CrossRef]
- Beer, C.; Reichstein, M.; Tomelleri, E.; Ciais, P.; Jung, M.; Carvalhais, N.; Rodenbeck, C.; Arain, M.A.; Baldocchi, D.; Bonan, G.B.; et al. Terrestrial Gross Carbon Dioxide Uptake: Global Distribution and Covariation with Climate. Science 2010, 329, 834–838. [Google Scholar] [CrossRef]
- Cho, S.; Kang, M.; Ichii, K.; Kim, J.; Lim, J.-H.; Chun, J.-H.; Park, C.-W.; Kim, H.S.; Choi, S.-W.; Lee, S.-H.; et al. Evaluation of forest carbon uptake in South Korea using the national flux tower network, remote sensing, and data-driven technology. Agric. For. Meteorol. 2021, 311, 108653. [Google Scholar] [CrossRef]
- Li, X.; Hu, X.M.; Cai, C.; Jia, Q.; Zhang, Y.; Liu, J.; Xue, M.; Xu, J.; Wen, R.; Crowell, S.M.R. Terrestrial CO2 Fluxes, Concentrations, Sources and Budget in Northeast China: Observational and Modeling Studies. J. Geophys. Res. Atmos. 2020, 125, e2019JD031686. [Google Scholar] [CrossRef]
- Dong, X.Y.; Yue, M.; Jiang, Y.J.; Hu, X.M.; Ma, Q.L.; Pu, J.J.; Zhou, G.Q. Analysis of CO2 spatio-temporal variations in China using a weather-biosphere online coupled model. Atmos. Chem. Phys. 2021, 21, 7217–7233. [Google Scholar] [CrossRef]
- Wei, D.; Qi, Y.; Ma, Y.; Wang, X.; Ma, W.; Gao, T.; Huang, L.; Zhao, H.; Zhang, J.; Wang, X. Plant uptake of CO2 outpaces losses from permafrost and plant respiration on the Tibetan Plateau. Proc. Natl. Acad. Sci. USA 2021, 118, e2015283118. [Google Scholar] [CrossRef]
- Liu, H.Y.; Mi, Z.R.; Lin, L.; Wang, Y.H.; Zhang, Z.H.; Zhang, F.W.; Wang, H.; Liu, L.L.; Zhu, B.A.; Cao, G.M.; et al. Shifting plant species composition in response to climate change stabilizes grassland primary production. Proc. Natl. Acad. Sci. USA 2018, 115, 4051–4056. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.Z.; Chen, L.Y.; Ji, C.J.; Hugelius, G.; Li, Y.N.; Liu, L.; Qin, S.Q.; Zhang, B.B.; Yang, G.B.; Li, F.; et al. Decadal soil carbon accumulation across Tibetan permafrost regions. Nat. Geosci. 2017, 10, 420–424. [Google Scholar] [CrossRef]
- Schuur, E.A.G.; Abbott, B.; Permafrost Carbon, N. High risk of permafrost thaw. Nature 2011, 480, 32–33. [Google Scholar] [CrossRef] [PubMed]
- Schneider von Deimling, T.; Meinshausen, M.; Levermann, A.; Huber, V.; Frieler, K.; Lawrence, D.M.; Brovkin, V. Estimating the near-surface permafrost-carbon feedback on global warming. Biogeosciences 2012, 9, 649–665. [Google Scholar] [CrossRef]
- Piao, S.L.; Wang, X.H.; Park, T.; Chen, C.; Lian, X.; He, Y.; Bjerke, J.W.; Chen, A.P.; Ciais, P.; Tommervik, H.; et al. Characteristics, drivers and feedbacks of global greening. Nat. Rev. Earth Environ. 2020, 1, 14–27. [Google Scholar] [CrossRef]
- Kroel-Dulay, G.; Mojzes, A.; Szitar, K.; Bahn, M.; Batary, P.; Beier, C.; Bilton, M.; De Boeck, H.J.; Dukes, J.S.; Estiarte, M.; et al. Field experiments underestimate aboveground biomass response to drought. Nat. Ecol. Evol. 2022, 6, 540–545. [Google Scholar] [CrossRef]
- De Cannière, S.; Herbst, M.; Vereecken, H.; Defourny, P.; Jonard, F. Constraining water limitation of photosynthesis in a crop growth model with sun-induced chlorophyll fluorescence. Remote Sens. Environ. 2021, 267, 112722. [Google Scholar] [CrossRef]
- Huete, A.R.; Liu, H.Q.; Batchily, K.; vanLeeuwen, W. A comparison of vegetation indices global set of TM images for EOS-MODIS. Remote Sens. Environ. 1997, 59, 440–451. [Google Scholar] [CrossRef]
- Xiao, X.; Hollinger, D.; Aber, J.; Goltz, M.; Davidson, E.A.; Zhang, Q.; Moore, B. Satellite-based modeling of gross primary production in an evergreen needleleaf forest. Remote Sens. Environ. 2004, 89, 519–534. [Google Scholar] [CrossRef]
- Churkina, G.; Schimel, D.; Braswell, B.H.; Xiao, X.M. Spatial analysis of growing season length control over net ecosystem exchange. Glob. Chang. Biol. 2005, 11, 1777–1787. [Google Scholar] [CrossRef]
- Jenkins, J.P.; Richardson, A.D.; Braswell, B.H.; Ollinger, S.V.; Hollinger, D.Y.; Smith, M.L. Refining light-use efficiency calculations for a deciduous forest canopy using simultaneous tower-based carbon flux and radiometric measurements. Agric. For. Meteorol. 2007, 143, 64–79. [Google Scholar] [CrossRef]
- Sims, D.; Rahman, A.; Cordova, V.; Elmasri, B.; Baldocchi, D.; Bolstad, P.; Flanagan, L.; Goldstein, A.; Hollinger, D.; Misson, L. A new model of gross primary productivity for North American ecosystems based solely on the enhanced vegetation index and land surface temperature from MODIS. Remote Sens. Environ. 2008, 112, 1633–1646. [Google Scholar] [CrossRef]
- Houborg, R.; Soegaard, H.; Boegh, E. Combining vegetation index and model inversion methods for the extraction of key vegetation biophysical parameters using Terra and Aqua MODIS reflectance data. Remote Sens. Environ. 2007, 106, 39–58. [Google Scholar] [CrossRef]
- Ahmadov, R.; Gerbig, C.; Kretschmer, R.; Koerner, S.; Neininger, B.; Dolman, A.J.; Sarrat, C. Mesoscale covariance of transport and CO2 fluxes: Evidence from observations and simulations using the WRF-VPRM coupled atmosphere-biosphere model. J. Geophys. Res.-Atmos. 2007, 112, D2210. [Google Scholar] [CrossRef]
- Mahadevan, P.; Wofsy, S.C.; Matross, D.M.; Xiao, X.; Dunn, A.L.; Lin, J.C.; Gerbig, C.; Munger, J.W.; Chow, V.Y.; Gottlieb, E.W. A satellite-based biosphere parameterization for net ecosystem CO2 exchange: Vegetation Photosynthesis and Respiration Model (VPRM). Glob. Biogeochem. Cycles 2008, 22, GB2005. [Google Scholar] [CrossRef]
- Vázquez-Lule, A.; Vargas, R. Proximal remote sensing and gross primary productivity in a temperate salt marsh. Agric. For. Meteorol. 2023, 341, 109639. [Google Scholar] [CrossRef]
- Gupta, S.; Burman, P.K.D.; Tiwari, Y.K.; Dumka, U.C.; Kumari, N.; Srivastava, A.; Raghubanshi, A.S. Understanding carbon sequestration trends using model and satellite data under different ecosystems in India. Sci. Total Environ. 2023, 897, 166381. [Google Scholar] [CrossRef] [PubMed]
- Fan, F.F.; Xiao, C.W.; Feng, Z.M.; Yang, Y.Z. Impact of human and climate factors on vegetation changes in mainland southeast asia and yunnan province of China. J. Clean. Prod. 2023, 415, 137690. [Google Scholar] [CrossRef]
- Hu, X.M.; Gourdji, S.M.; Davis, K.J.; Wang, Q.; Zhang, Y.; Xue, M.; Feng, S.; Moore, B.; Crowell, S.M.R. Implementation of Improved Parameterization of Terrestrial Flux in WRF-VPRM Improves the Simulation of Nighttime CO2 Peaks and a Daytime CO2 Band Ahead of a Cold Front. J. Geophys. Res. Atmos. 2021, 126, e2020JD034362. [Google Scholar] [CrossRef]
- Hu, X.M.; Crowell, S.; Wang, Q.; Zhang, Y.; Davis, K.J.; Xue, M.; Xiao, X.; Moore, B.; Wu, X.; Choi, Y.; et al. Dynamical Downscaling of CO2 in 2016 Over the Contiguous United States Using WRF-VPRM, a Weather-Biosphere-Online-Coupled Model. J. Adv. Model. Earth Syst. 2020, 12, e2019MS001875. [Google Scholar] [CrossRef]
- Raoult, N.; Charbit, S.; Dumas, C.; Maignan, F.; Ottlé, C.; Bastrikov, V. Improving modelled albedo over the Greenland ice sheet through parameter optimisation and MODIS snow albedo retrievals. Cryosphere 2023, 17, 2705–2724. [Google Scholar] [CrossRef]
- Bacour, C.; MacBean, N.; Chevallier, F.; Léonard, S.; Koffi, E.N.; Peylin, P. Assimilation of multiple datasets results in large differences in regional- to global-scale NEE and GPP budgets simulated by a terrestrial biospheremodel. Biogeosciences 2023, 20, 1089–1111. [Google Scholar] [CrossRef]
- Yang, S.Y.; Zhao, B.X.; Yang, D.W.; Wang, T.H.; Yang, Y.T.; Ma, T.; Santisirisomboon, J. Future changes in water resources, floods and droughts under the joint impact of climate and land-use changes in the Chao Phraya basin, Thailand. J. Hydrol. 2023, 620, 129454. [Google Scholar] [CrossRef]
- Song, X.; Zheng, B.F.; Hu, F.Q.; Xu, L.L.; Wu, H.Q.; Liu, Z.; Wan, W. Divergent Responses of NPP to Climate Factors among Forest Types at Interannual and Inter-Monthly Scales: An Empirical Study on Four Typical Forest Types in Subtropical China. Forests 2023, 14, 1474. [Google Scholar] [CrossRef]
- Kretschmer, R.; Gerbig, C.; Karstens, U.; Koch, F.T. Error characterization of CO2 vertical mixing in the atmospheric transport model WRF-VPRM. Atmos. Chem. Phys. 2012, 12, 2441–2458. [Google Scholar] [CrossRef]
- Pillai, D.; Gerbig, C.; Ahmadov, R.; Rodenbeck, C.; Kretschmer, R.; Koch, T.; Thompson, R.; Neininger, B.; Lavric, J.V. High-resolution simulations of atmospheric CO2 over complex terrain—Representing the Ochsenkopf mountain tall tower. Atmos. Chem. Phys. 2011, 11, 7445–7464. [Google Scholar] [CrossRef]
- Raich, J.W.; Rastetter, E.B.; Melillo, J.M.; Kicklighter, D.W.; Steudler, P.A.; Peterson, B.J.; Grace, A.L.; Moore, B.; Vorosmarty, C.J. Potential Net Primary Productivity In South-America—Application Of A Global-Model. Ecol. Appl. 1991, 1, 399–429. [Google Scholar] [CrossRef]
- Lee, Y.-H.; Park, S.-U. Evaluation of a modified soil–plant–atmosphere model for CO2 flux and latent heat flux in open canopies. Agric. For. Meteorol. 2007, 143, 230–241. [Google Scholar] [CrossRef]
- Joetzjer, E.; Delire, C.; Douville, H.; Ciais, P.; Decharme, B.; Carrer, D.; Verbeeck, H.; DeWeirdt, M.; Bonal, D. Improving the ISBA(CC) land surface model simulation of water and carbon fluxes and stocks over the Amazon forest. Geosci. Model Dev. 2015, 8, 1709–1727. [Google Scholar] [CrossRef]
- Zhang, T.; Zhang, Y.; Xu, M.; Zhu, J.; Chen, N.; Jiang, Y.; Huang, K.; Zu, J.; Liu, Y.; Yu, G. Water availability is more important than temperature in driving the carbon fluxes of an alpine meadow on the Tibetan Plateau. Agric. For. Meteorol. 2018, 256–257, 22–31. [Google Scholar] [CrossRef]
- Cao, G.; Tang, Y.; Mo, W.; Wang, Y.; Li, Y.; Zhao, X. Grazing intensity alters soil respiration in an alpine meadow on the Tibetan plateau. Soil Biol. Biochem. 2004, 36, 237–243. [Google Scholar] [CrossRef]
- Kato, T.; Tang, Y.; Gu, S.; Hirota, M.; Du, M.; Li, Y.; Zhao, X. Temperature and biomass influences on interannual changes in CO2. exchange in an alpine meadow on the Qinghai-Tibetan Plateau. Glob. Chang. Biol. 2006, 12, 1285–1298. [Google Scholar] [CrossRef]
- Zhao, L.; Li, Y.; Xu, S.; Zhou, H.; Gu, S.; Yu, G.; Zhao, X. Diurnal, seasonal and annual variation in net ecosystem CO2 exchange of an alpine shrubland on Qinghai-Tibetan plateau. Glob. Chang. Biol. 2006, 12, 1940–1953. [Google Scholar] [CrossRef]
- Zhuang, Q.; He, J.; Lu, Y.; Ji, L.; Xiao, J.; Luo, T. Carbon dynamics of terrestrial ecosystems on the Tibetan Plateau during the 20th century: An analysis with a process-based biogeochemical model. Glob. Ecol. Biogeogr. 2010, 119, 649–662. [Google Scholar] [CrossRef]
- Chen, J.; Luo, Y.; Xia, J.; Shi, Z.; Jiang, L.; Niu, S.; Zhou, X.; Cao, J. Differential responses of ecosystem respiration components to experimental warming in a meadow grassland on the Tibetan Plateau. Agric. For. Meteorol. 2016, 220, 21–29. [Google Scholar] [CrossRef]
- Li, H.; Zhang, F.; Li, Y.; Wang, J.; Zhang, L.; Zhao, L.; Cao, G.; Zhao, X.; Du, M. Seasonal and inter-annual variations in CO2 fluxes over 10 years in an alpine shrubland on the Qinghai-Tibetan Plateau, China. Agric. For. Meteorol. 2016, 228–229, 95–103. [Google Scholar] [CrossRef]
- Li, X.; Cheng, G.D.; Liu, S.M.; Xiao, Q.; Ma, M.G.; Jin, R.; Che, T.; Liu, Q.H.; Wang, W.Z.; Qi, Y.; et al. Heihe Watershed Allied Telemetry Experimental Research (HiWATER): Scientific Objectives and Experimental Design. Bull. Am. Meteorol. Soc. 2013, 94, 1145–1160. [Google Scholar] [CrossRef]
- Liu, S.M.; Xu, Z.W.; Song, L.S.; Zhao, Q.Y.; Ge, Y.; Xu, T.R.; Ma, Y.F.; Zhu, Z.L.; Jia, Z.Z.; Zhang, F. Upscaling evapotranspiration measurements from multi-site to the satellite pixel scale over heterogeneous land surfaces. Agric. For. Meteorol. 2016, 230, 97–113. [Google Scholar] [CrossRef]
- Liu, S.M.; Xu, Z.W.; Wang, W.Z.; Jia, Z.Z.; Zhu, M.J.; Bai, J.; Wang, J.M. A comparison of eddy-covariance and large aperture scintillometer measurements with respect to the energy balance closure problem. Hydrol. Earth Syst. Sci. 2011, 15, 1291–1306. [Google Scholar] [CrossRef]
- Shang, L.; Zhang, Y.; Lyu, S.; Wang, S. Seasonal and Inter-Annual Variations in Carbon Dioxide Exchange over an Alpine Grassland in the Eastern Qinghai-Tibetan Plateau. PLoS ONE 2016, 11, e0166837. [Google Scholar] [CrossRef]
- Nieberding, F.; Wille, C.; Fratini, G.; Asmussen, M.O.; Wang, Y.Y.; Ma, Y.M.; Sachs, T. A long-term (2005-2019) eddy covariance data set of CO2 and H2O fluxes from the Tibetan alpine steppe. Earth Syst. Sci. Data 2020, 12, 2505–2524. [Google Scholar] [CrossRef]
- Wu, F.Q.; Guo, S.M.; Huang, W.B.; Han, Y.C.; Wang, Z.B.; Wang, G.P.; Li, X.F.; Lei, Y.P.; Yang, B.F.; Xiong, S.W.; et al. Adaptation of cotton production to climate change by sowing date optimization and precision resource management. Ind. Crops Prod. 2023, 203, 117167. [Google Scholar] [CrossRef]
- Omazic, B.; Prtenjak, M.T.; Bubola, M.; Mestric, J.; Karoglan, M.; Prsa, I. Application of statistical models in the detection of grapevine phenology changes. Agric. For. Meteorol. 2023, 341, 109682. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Pillai, D.; Gerbig, C.; Marshall, J.; Ahmadov, R.; Kretschmer, R.; Koch, T.; Karstens, U. High resolution modeling of CO2 over Europe: Implications for representation errors of satellite retrievals. Atmos. Chem. Phys. 2010, 10, 83–94. [Google Scholar] [CrossRef]
- Kanamitsu, M.; Ebisuzaki, W.; Woollen, J.; Yang, S.K.; Hnilo, J.J.; Fiorino, M.; Potter, G.L. NCEP-DOE AMIP-II reanalysis (R-2). Bull. Am. Meteorol. Soc. 2002, 83, 1631–1643. [Google Scholar] [CrossRef]
- Meng Xianhong, L.I.Z. Meteorological Observation Data at Maqu Grassland Site from 2017 to 2020; National Tibetan Plateau Data Center: 2022. Available online: https://data.tpdc.ac.cn/en/data/3966b5db-9a4f-44e0-afac-4a1eedc2cfce/ (accessed on 9 October 2022).
- Weir, B.; Crisp, D.; O’Dell, C.W.; Basu, S.; Chatterjee, A.; Kolassa, J.; Oda, T.; Pawson, S.; Poulter, B.; Zhang, Z.; et al. Regional impacts of COVID-19 on carbon dioxide detected worldwide from space. Sci. Adv. 2021, 7, eabf9415. [Google Scholar] [CrossRef]
- Sheng, M.Y.; Lei, L.P.; Zeng, Z.C.; Rao, W.Q.; Zhang, S.Q. Detecting the Responses of CO2 Column Abundances to Anthropogenic Emissions from Satellite Observations of GOSAT and OCO-2. Remote Sens. 2021, 13, 3524. [Google Scholar] [CrossRef]
- Miglietta, F.; Raschi, A.; Resti, R.; Badiani, M. Growth and onto-morphogenesis of soybean (Glycine max Merril) in an open, naturally CO2-enriched environment. Plant Cell Environ. 1993, 16, 909–918. [Google Scholar] [CrossRef]
- Li, K.F. An Intraseasonal Variability in CO2 Over the Arctic Induced by the Madden-Julian Oscillation. Geophys. Res. Lett. 2018, 45, 1630–1638. [Google Scholar] [CrossRef]
- Hakkarainen, J.; Ialongo, I.; Maksyutov, S.; Crisp, D. Analysis of Four Years of Global XCO2 Anomalies as Seen by Orbiting Carbon Observatory-2. Remote Sens. 2019, 11, 850. [Google Scholar] [CrossRef]
- Xie, J.; Yin, G.; Ma, D.; Chen, R.; Zhao, W.; Xie, Q.; Wang, C.; Lin, S.; Yuan, W. Climatic limitations on grassland photosynthesis over the Tibetan Plateau shifted from temperature to water. Sci. Total Environ. 2023, 906, 167663. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Huang, K.; Yan, H.; Yan, H.; Zhou, L.; Wang, H.; Zhang, J.; Yan, J.; Zhao, L.; Wang, Y.; et al. Improving the light use efficiency model for simulating terrestrial vegetation gross primary production by the inclusion of diffuse radiation across ecosystems in China. Ecol. Complex. 2015, 23, 1–13. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Site | Altitude (m) | Substrate |
|---|---|---|---|
| Maqu | 33.8975°N, 102.1619°E | 3423 | Kobresia tibetica and K. humilis |
| Yakou | 38.0142°N, 100.2421°E | 4148 | Alpine grassland |
| Dashalong | 38.8399°N, 98.9406°E | 3739 | Swampy alpine meadows |
| Arou | 38.0473°N, 100.4643°E | 3033 | Alpine grassland |
| Nam CO | 30.7667°N, 90.95°E | 4730 | K. pygmaea and alpine steppe |
| Mt. Waliguan | 36.28°N, 100.9°E | 3810 | Arid and semi-arid grasslands, tundra, and deserts |
| NEE | Model | Maqu | Yakou | Dashalong | Arou | ||||
|---|---|---|---|---|---|---|---|---|---|
| 2016 | 2017 | 2016 | 2017 | 2016 | 2017 | 2016 | 2017 | ||
| bias (μmol·m−2·s−1) | Original | −1 | −0.991 | −0.305 | −1.492 | −1.272 | −1.263 | −0.123 | −0.418 |
| Improved | 0.398 | 0.439 | 0.012 | −0.498 | −0.522 | −0.59 | 0.335 | 0.364 | |
| RMSE (μmol·m−2·s−1) | Original | 1.989 | 1.813 | 1.484 | 2.106 | 1.58 | 1.474 | 1.461 | 1.501 |
| Improved | 1.686 | 1.532 | 1.546 | 1.358 | 1.048 | 1.015 | 1.338 | 1.363 | |
| r | Original | 0.721 | 0.62 | 0.533 | 0.429 | 0.278 | 0.711 | 0.584 | 0.594 |
| Improved | 0.646 | 0.572 | 0.472 | 0.494 | 0.394 | 0.602 | 0.673 | 0.65 | |
| RSD | Original | 0.85 | 0.662 | 0.26 | 0.107 | 0.74 | 0.817 | 0.085 | 0.16 |
| Improved | 0.406 | 0.398 | 0.26 | 0.099 | 1.293 | 0.89 | 0.146 | 0.057 | |
| GEE | Model | Maqu | Yakou | Dashalong | Arou | ||||
|---|---|---|---|---|---|---|---|---|---|
| 2016 | 2017 | 2016 | 2017 | 2016 | 2017 | 2016 | 2017 | ||
| bias (μmol·m−2·s−1) | Original | 2.732 | 2.707 | 1.026 | −0.365 | −0.134 | −0.475 | 4.539 | 2.893 |
| Improved | 0.944 | 1.169 | −0.013 | −1.177 | −1.046 | −1.271 | 3.064 | 1.849 | |
| RMSE (μmol·m−2·s−1) | Original | 2.991 | 2.972 | 1.775 | 1.463 | 0.779 | 0.918 | 4.838 | 3.172 |
| Improved | 1.772 | 1.927 | 1.444 | 2.004 | 1.608 | 1.791 | 3.486 | 2.28 | |
| r | Original | 0.88 | 0.827 | 0.633 | 0.558 | 0.663 | 0.757 | 0.776 | 0.822 |
| Improved | 0.893 | 0.848 | 0.715 | 0.613 | 0.708 | 0.748 | 0.82 | 0.842 | |
| RSD | Original | 0.009 | 0.032 | 0.276 | 0.102 | 0.606 | 0.783 | 0.562 | 0.205 |
| Improved | 0.004 | 0.053 | 0.011 | 0.434 | 1.233 | 1.562 | 0.401 | 0.037 | |
| RE | Model | Maqu | Yakou | Dashalong | Arou | ||||
|---|---|---|---|---|---|---|---|---|---|
| 2016 | 2017 | 2016 | 2017 | 2016 | 2017 | 2016 | 2017 | ||
| bias (μmol·m−2·s−1) | Original | −3.591 | −3.52 | −1.385 | −1.119 | −1.135 | −0.771 | −3.946 | −3.5 |
| Improved | −0.67 | −0.741 | −0.017 | 0.31 | 0.126 | 0.182 | −1.387 | −1.143 | |
| RMSE (μmol·m−2·s−1) | Original | 3.897 | 3.775 | 1.389 | 1.122 | 1.211 | 0.775 | 4.149 | 3.629 |
| Improved | 0.881 | 0.906 | 0.812 | 0.956 | 0.474 | 0.805 | 1.567 | 1.234 | |
| r | Original | 0.814 | 0.765 | 0.947 | 0.922 | 0.597 | 0.693 | 0.784 | 0.776 |
| Improved | 0.956 | 0.953 | 0.876 | 0.852 | 0.912 | 0.887 | 0.96 | 0.953 | |
| RSD | Original | 0.917 | 0.916 | 0.377 | 0.39 | 0.744 | 0.235 | 0.904 | 0.895 |
| Improved | 0.083 | 0.067 | 2.126 | 2.767 | 0.579 | 4.079 | 0.332 | 0.146 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niu, H.; Hu, X.-M.; Shang, L.; Meng, X.; Wang, S.; Li, Z.; Zhao, L.; Chen, H.; Deng, M.; Sheng, D. Evaluation of Original and Water Stress-Incorporated Modified Weather Research and Forecasting Vegetation Photosynthesis and Respiration Model in Simulating CO2 Flux and Concentration Variability over the Tibetan Plateau. Remote Sens. 2023, 15, 5474. https://doi.org/10.3390/rs15235474
Niu H, Hu X-M, Shang L, Meng X, Wang S, Li Z, Zhao L, Chen H, Deng M, Sheng D. Evaluation of Original and Water Stress-Incorporated Modified Weather Research and Forecasting Vegetation Photosynthesis and Respiration Model in Simulating CO2 Flux and Concentration Variability over the Tibetan Plateau. Remote Sensing. 2023; 15(23):5474. https://doi.org/10.3390/rs15235474
Chicago/Turabian StyleNiu, Hanlin, Xiao-Ming Hu, Lunyu Shang, Xianhong Meng, Shaoying Wang, Zhaoguo Li, Lin Zhao, Hao Chen, Mingshan Deng, and Danrui Sheng. 2023. "Evaluation of Original and Water Stress-Incorporated Modified Weather Research and Forecasting Vegetation Photosynthesis and Respiration Model in Simulating CO2 Flux and Concentration Variability over the Tibetan Plateau" Remote Sensing 15, no. 23: 5474. https://doi.org/10.3390/rs15235474
APA StyleNiu, H., Hu, X.-M., Shang, L., Meng, X., Wang, S., Li, Z., Zhao, L., Chen, H., Deng, M., & Sheng, D. (2023). Evaluation of Original and Water Stress-Incorporated Modified Weather Research and Forecasting Vegetation Photosynthesis and Respiration Model in Simulating CO2 Flux and Concentration Variability over the Tibetan Plateau. Remote Sensing, 15(23), 5474. https://doi.org/10.3390/rs15235474