



Large-Scale Controls on the Leaf Economic Spectrum of the Overstory Tree Species Metrosideros polymorpha


Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Collection
2.2. Analysis
3. Results
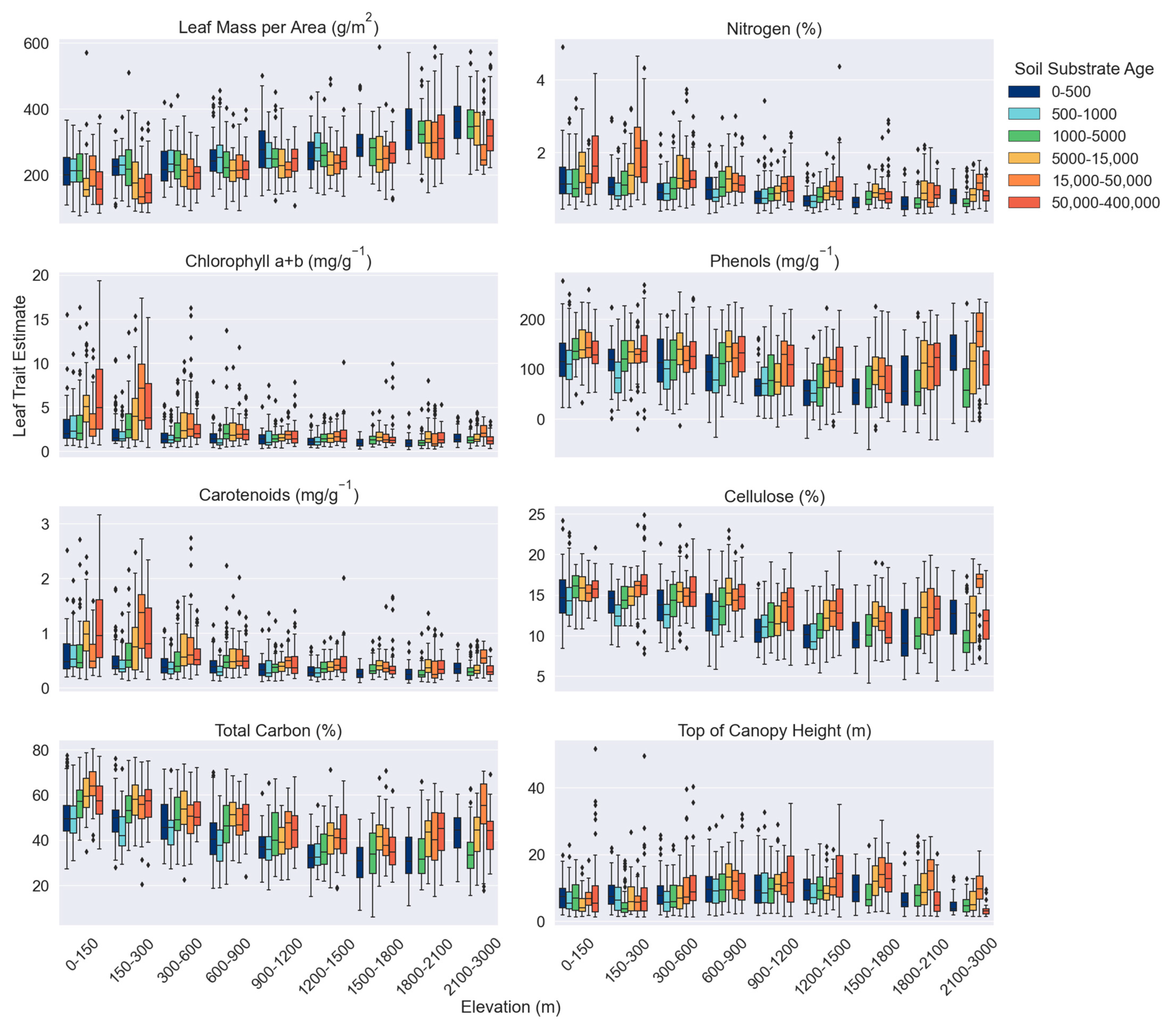
3.1. Canopy Trait Variation across Environmental Gradients
3.2. Canopy Trait Relationships as Mediated by Environmental Factors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schleuter, D.; Daufresne, M.; Massol, F.; Argillier, C. A user’s guide to functional diversity indices. Ecol. Monogr. 2010, 80, 469–484. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef]
- Wright, S.J.; Kitajima, K.; Kraft, N.J.B.; Reich, P.B.; Wright, I.J.; Bunker, D.E.; Condit, R.; Dalling, J.W.; Davies, S.J.; Díaz, S.; et al. Functional traits and the growth–mortality trade-off in tropical trees. Ecology 2010, 91, 3664–3674. [Google Scholar] [CrossRef]
- Fajardo, A.; Siefert, A. Intraspecific trait variation and the leaf economics spectrum across resource gradients and levels of organization. Ecology 2018, 99, 1024–1030. [Google Scholar] [CrossRef]
- Hecking, M.J.; Zukswert, J.M.; Drake, J.E.; Dovciak, M.; Burton, J.I. Montane Temperate-Boreal Forests Retain the Leaf Economic Spectrum Despite Intraspecific Variability. Front. For. Glob. Chang. 2022, 4, 754063. Available online: https://www.frontiersin.org/articles/10.3389/ffgc.2021.754063 (accessed on 10 June 2023). [CrossRef]
- Laughlin, D.C.; Lusk, C.H.; Bellingham, P.J.; Burslem, D.F.R.P.; Simpson, A.H.; Kramer-Walter, K.R. Intraspecific trait variation can weaken interspecific trait correlations when assessing the whole-plant economic spectrum. Ecol. Evol. 2017, 7, 8936–8949. [Google Scholar] [CrossRef]
- Wright, J.P.; Sutton-Grier, A. Does the leaf economic spectrum hold within local species pools across varying environmental conditions? Funct. Ecol. 2012, 26, 1390–1398. [Google Scholar] [CrossRef]
- Siefert, A.; Violle, C.; Chalmandrier, L.; Albert, C.H.; Taudiere, A.; Fajardo, A.; Aarssen, L.W.; Baraloto, C.; Carlucci, M.B.; Cianciaruso, M.V.; et al. A global meta-analysis of the relative extent of intraspecific trait variation in plant communities. Ecol. Lett. 2015, 18, 1406–1419. [Google Scholar] [CrossRef]
- Albert, C.H.; Thuiller, W.; Yoccoz, N.G.; Soudant, A.; Boucher, F.; Saccone, P.; Lavorel, S. Intraspecific functional variability: Extent, structure and sources of variation. J. Ecol. 2010, 98, 604–613. [Google Scholar] [CrossRef]
- De Bello, F.; Lavorel, S.; Albert, C.H.; Thuiller, W.; Grigulis, K.; Dolezal, J.; Janeček, Š.; Lepš, J. Quantifying the relevance of intraspecific trait variability for functional diversity. Methods Ecol. Evol. 2011, 2, 163–174. [Google Scholar] [CrossRef]
- Jung, V.; Violle, C.; Mondy, C.; Hoffmann, L.; Muller, S. Intraspecific variability and trait-based community assembly. J. Ecol. 2010, 98, 1134–1140. [Google Scholar] [CrossRef]
- Paine, C.E.T.; Baraloto, C.; Chave, J.; Hérault, B. Functional traits of individual trees reveal ecological constraints on community assembly in tropical rain forests. Oikos 2011, 120, 720–727. [Google Scholar] [CrossRef]
- Boardman, J.W.; Green, R.O. Exploring the Spectral Variability of the Earth as Measured by AVIRIS in 1999. 2000. Available online: https://trs.jpl.nasa.gov/handle/2014/16602 (accessed on 9 June 2023).
- Green, R.O.; Boardman, J.W. Exploration of the relationship between information content and signal-to-noise ratio and spatial resolution in AVIRIS spectral data. Spectrum 2000, 7, 1–12. [Google Scholar]
- Asner, G.P.; Martin, R.E.; Anderson, C.B.; Knapp, D.E. Quantifying forest canopy traits: Imaging spectroscopy versus field survey. Remote Sens. Environ. 2015, 158, 15–27. [Google Scholar] [CrossRef]
- Asner, G.P.; Martin, R.E. Airborne spectranomics: Mapping canopy chemical and taxonomic diversity in tropical forests. Front. Ecol. Environ. 2009, 7, 269–276. [Google Scholar] [CrossRef]
- Asner, G.P.; Martin, R.E. Spectranomics: Emerging science and conservation opportunities at the interface of biodiversity and remote sensing. Glob. Ecol. Conserv. 2016, 8, 212–219. [Google Scholar] [CrossRef]
- Féret, J.-B.; Asner, G.P. Mapping tropical forest canopy diversity using high-fidelity imaging spectroscopy. Ecol. Appl. 2014, 24, 1289–1296. [Google Scholar] [CrossRef]
- Asner, G.P.; Knapp, D.E.; Anderson, C.B.; Martin, R.E.; Vaughn, N. Large-scale climatic and geophysical controls on the leaf economics spectrum. Proc. Natl. Acad. Sci. USA 2016, 113, E4043–E4051. [Google Scholar] [CrossRef]
- Stacy, E.A.; Johansen, J.B.; Sakishima, T.; Price, D.K.; Pillon, Y. Incipient radiation within the dominant Hawaiian tree Metrosideros polymorpha. Heredity 2014, 113, 334–342. [Google Scholar] [CrossRef]
- Stacy, E.A.; Johansen, J.B.; Sakishima, T.; Price, D.K. Genetic analysis of an ephemeral intraspecific hybrid zone in the hypervariable tree, Metrosideros polymorpha, on Hawai’i Island. Heredity 2016, 117, 173–183. [Google Scholar] [CrossRef]
- Seeley, M.M.; Vaughn, N.R.; Shanks, B.L.; Martin, R.E.; König, M.; Asner, G.P. Classifying a highly polymorphic tree species across landscapes using airborne imaging spectroscopy. Remote Sens. 2023, 15, 4365. [Google Scholar] [CrossRef]
- Asner, G.P.; Knapp, D.E.; Boardman, J.; Green, R.O.; Kennedy-Bowdoin, T.; Eastwood, M.; Martin, R.E.; Anderson, C.; Field, C.B. Carnegie Airborne Observatory-2: Increasing science data dimensionality via high-fidelity multi-sensor fusion. Remote Sens. Environ. 2012, 124, 454–465. [Google Scholar] [CrossRef]
- Seeley, M.M.; Martin, R.E.; Vaughn, N.R.; Thompson, D.R.; Dai, J.; Asner, G.P. Quantifying the Variation in Reflectance Spectra of Metrosideros polymorpha Canopies across Environmental Gradients. Remote Sens. 2023, 15, 1614. [Google Scholar] [CrossRef]
- Farr, T.G.; Rosen, P.A.; Caro, E.; Crippen, R.; Duren, R.; Hensley, S.; Kobrick, M.; Paller, M.; Rodriguez, E.; Roth, L.; et al. The Shuttle Radar Topography Mission. Rev. Geophys. 2007, 45, 1–33. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1029/2005RG000183 (accessed on 21 June 2023). [CrossRef]
- PRISM Climate Group, O.S.U. PRISM Gridded Climate Data 2014. Available online: https://prism.oregonstate.edu (accessed on 20 May 2023).
- Sherrod, D.R.; Sinton, J.M.; Watkins, S.E.; Brunt, K.M. Geologic Map of the State of Hawai’i; Open-File Report 2007-1089; U.S. Geological Survey: Reston, VA, USA, 2007. Available online: http://pubs.usgs.gov/of/2007/1089/ (accessed on 20 May 2023).
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V.; et al. Scikit-learn: Machine Learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]
- Cordell, S.; Goldstein, G.; Mueller-Dombois, D.; Webb, D.; Vitousek, P.M. Physiological and morphological variation in Metrosideros polymorpha, a dominant Hawaiian tree species, along an altitudinal gradient: The role of phenotypic plasticity. Oecologia 1998, 113, 188–196. [Google Scholar] [CrossRef]
- Joel, G.; Aplet, G.; Vitousek, P.M. Leaf Morphology Along Environmental Gradients in Hawaiian Metrosideros Polymorpha. Biotropica 1994, 26, 17–22. [Google Scholar] [CrossRef]
- Martin, R.E.; Asner, G.P. Leaf Chemical and Optical Properties of Metrosideros polymorpha across Environmental Gradients in Hawaii. Biotropica 2009, 41, 292–301. [Google Scholar] [CrossRef]
- Waskom, M.L. seaborn: Statistical data visualization. J. Open Source Softw. 2021, 6, 3021. [Google Scholar] [CrossRef]
- Hayes, F.J.; Buchanan, S.W.; Coleman, B.; Gordon, A.M.; Reich, P.B.; Thevathasan, N.V.; Wright, I.J.; Martin, A.R. Intraspecific variation in soy across the leaf economics spectrum. Ann. Bot. 2019, 123, 107–120. [Google Scholar] [CrossRef]
- Vasseur, F.; Violle, C.; Enquist, B.J.; Granier, C.; Vile, D. A common genetic basis to the origin of the leaf economics spectrum and metabolic scaling allometry. Ecol. Lett. 2012, 15, 1149–1157. [Google Scholar] [CrossRef] [PubMed]
- Cordell, S.; Goldstein, G.; Meinzer, F.C.; Handley, L.L. Allocation of nitrogen and carbon in leaves of Metrosideros polymorpha regulates carboxylation capacity and δ13C along an altitudinal gradient. Funct. Ecol. 1999, 13, 811–818. [Google Scholar] [CrossRef]
- Corn, C.A.; Hiesey, W.M. Altitudinal Variation in Hawaiian Metrosideros. Am. J. Bot. 1973, 60, 991–1002. [Google Scholar] [CrossRef]
- Drake, D.R.; Mueller-Dombois, D. Population Development of Rain Forest Trees on a Chronosequence of Hawaiian Lava Flows. Ecology 1993, 74, 1012–1019. [Google Scholar] [CrossRef]
- Kitayama, K. Ecological and Genetic Implications of Foliar Polymorphism inMetrosideros polymorphaGaud. (Myrtaceae) in a Habitat Matrix on Mauna Loa, Hawaii. Ann. Bot. 1997, 80, 491–497. [Google Scholar] [CrossRef]
- Mueller-Dombois, D. Vegetation dynamics and the evolution of Metrosideros polymorpha in Hawaii. Phytocoenologia 1994, 24, 609–614. [Google Scholar] [CrossRef]
- Stemmermann, L. Ecological studies of Hawaiian Metrosideros in a successional context. Pac. Sci. 1983, 37, 361–373. [Google Scholar]
- Stemmermann, L.; Ihsle, T. Replacement of Metrosideros polymorpha, `Ohi`a, in Hawaiian Dry Forest Succession. Biotropica 1993, 25, 36–45. [Google Scholar] [CrossRef]
- Cordell, S.; Goldstein, G.; Meinzer, F.C.; Vitousek, P.M. Regulation of leaf life-span and nutrient-use efficiency of Metrosideros polymorpha trees at two extremes of a long chronosequence in Hawaii. Oecologia 2001, 127, 198–206. [Google Scholar] [CrossRef]
- Seeley, M.M.; Stacy, E.A.; Martin, R.E.; Asner, G.P. Foliar functional and genetic variation in a keystone Hawaiian tree species estimated through spectroscopy. Oecologia 2023, 202, 15–28. [Google Scholar] [CrossRef]
- Izuno, A.; Kitayama, K.; Onoda, Y.; Tsujii, Y.; Hatakeyama, M.; Nagano, A.J.; Honjo, M.N.; Shimizu-Inatsugi, R.; Kudoh, H.; Shimizu, K.K.; et al. The population genomic signature of environmental association and gene flow in an ecologically divergent tree species Metrosideros polymorpha (Myrtaceae). Mol. Ecol. 2017, 26, 1515–1532. [Google Scholar] [CrossRef]
- Gerdol, R.; Iacumin, P.; Tonin, R. Bedrock geology affects foliar nutrient status but has minor influence on leaf carbon isotope discrimination across altitudinal gradients. PLoS ONE 2018, 13, e0202810. [Google Scholar] [CrossRef]
- Midolo, G.; De Frenne, P.; Hölzel, N.; Wellstein, C. Global patterns of intraspecific leaf trait responses to elevation. Glob. Chang. Biol. 2019, 25, 2485–2498. [Google Scholar] [CrossRef]
- Körner, C. Alpine Treelines: Functional Ecology of the Global High Elevation Tree Limits; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012; ISBN 978-3-0348-0396-0. [Google Scholar]
- Körner, C. The nutritional status of plants from high altitudes. Oecologia 1989, 81, 379–391. [Google Scholar] [CrossRef]
- Albert, C.H.; de Bello, F.; Boulangeat, I.; Pellet, G.; Lavorel, S.; Thuiller, W. On the importance of intraspecific variability for the quantification of functional diversity. Oikos 2012, 121, 116–126. [Google Scholar] [CrossRef]
- Crutsinger, G.M.; Souza, L.; Sanders, N.J. Intraspecific diversity and dominant genotypes resist plant invasions. Ecol. Lett. 2008, 11, 16–23. [Google Scholar] [CrossRef]
- Des Roches, S.; Post, D.M.; Turley, N.E.; Bailey, J.K.; Hendry, A.P.; Kinnison, M.T.; Schweitzer, J.A.; Palkovacs, E.P. The ecological importance of intraspecific variation. Nat. Ecol. Evol. 2018, 2, 57–64. [Google Scholar] [CrossRef]
- Baldeck, C.A.; Asner, G.P.; Martin, R.E.; Anderson, C.B.; Knapp, D.E.; Kellner, J.R.; Wright, S.J. Operational Tree Species Mapping in a Diverse Tropical Forest with Airborne Imaging Spectroscopy. PLoS ONE 2015, 10, e0118403. [Google Scholar] [CrossRef]
- Balzotti, C.S.; Asner, G.P.; Adkins, E.D.; Parsons, E.W. Spatial drivers of composition and connectivity across endangered tropical dry forests. J. Appl. Ecol. 2020, 57, 1593–1604. [Google Scholar] [CrossRef]
- Chakravortty, S.; Shah, E.; Chowdhury, A.S. Application of Spectral Unmixing Algorithm on Hyperspectral Data for Mangrove Species Classification. In Proceedings of the Applied Algorithms: First International Conference, ICAA 2014, Kolkata, India, 13–15 January 2014; Gupta, P., Zaroliagis, C., Eds.; Springer International Publishing: Cham, Switzerland, 2014; pp. 223–236, (Lecture Notes in Computer Science). [Google Scholar]
- Pontius, J.; Hanavan, R.P.; Hallett, R.A.; Cook, B.D.; Corp, L.A. High spatial resolution spectral unmixing for mapping ash species across a complex urban environment. Remote Sens. Environ. 2017, 199, 360–369. [Google Scholar] [CrossRef]
- Shang, X.; Chisholm, L.A. Classification of Australian Native Forest Species Using Hyperspectral Remote Sensing and Machine-Learning Classification Algorithms. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2014, 7, 2481–2489. [Google Scholar] [CrossRef]
- Somers, B.; Asner, G.P. Tree species mapping in tropical forests using multi-temporal imaging spectroscopy: Wavelength adaptive spectral mixture analysis. Int. J. Appl. Earth Obs. Geoinf. 2014, 31, 57–66. [Google Scholar] [CrossRef]
- Torabzadeh, H.; Leiterer, R.; Hueni, A.; Schaepman, M.E.; Morsdorf, F. Tree species classification in a temperate mixed forest using a combination of imaging spectroscopy and airborne laser scanning. Agric. For. Meteorol. 2019, 279, 107744. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Leaf Mass per Area | Chlorophyll a+b | Carotenoids | Total Carbon | Nitrogen | Phosphorus | Phenols | Cellulose | Top of Canopy Height | ||
|---|---|---|---|---|---|---|---|---|---|---|
| Elevation | Slope | 16.972 | −0.118 | −0.095 | −2.256 | −0.074 | −0.037 | −5.179 | −0.500 | 0.121 |
| R2 | 0.282 | 0.162 | 0.166 | 0.205 | 0.157 | 0.039 | 0.055 | 0.160 | 0.003 | |
| Soil Substrate Age | Slope | −6.908 | 0.098 | 0.083 | 1.702 | 0.073 | 0.027 | 6.934 | 0.443 | 0.437 |
| r2 | 0.021 | 0.051 | 0.057 | 0.053 | 0.070 | 0.010 | 0.045 | 0.057 | 0.015 | |
| Elevation Ranges: | ||||||||||
| 0–150 | Slope | −6.950 | 0.132 | 0.090 | 2.291 | 0.056 | 0.097 | 4.563 | 0.162 | 0.190 |
| r2 | 0.032 | 0.086 | 0.061 | 0.131 | 0.042 | 0.108 | 0.028 | 0.014 | 0.003 | |
| 150–300 | Slope | −15.161 | 0.222 | 0.178 | 2.047 | 0.129 | 0.021 | 6.740 | 0.587 | −0.110 |
| r2 | 0.133 | 0.198 | 0.197 | 0.107 | 0.191 | 0.008 | 0.059 | 0.160 | 0.001 | |
| 300–600 | Slope | −8.200 | 0.141 | 0.116 | 1.254 | 0.097 | 0.000 | 4.973 | 0.449 | 0.576 |
| r2 | 0.051 | 0.108 | 0.116 | 0.043 | 0.131 | 0.000 | 0.028 | 0.084 | 0.026 | |
| 600–900 | Slope | −7.346 | 0.095 | 0.082 | 2.063 | 0.069 | 0.033 | 9.178 | 0.552 | 0.397 |
| r2 | 0.042 | 0.069 | 0.082 | 0.108 | 0.084 | 0.016 | 0.086 | 0.104 | 0.013 | |
| 900–1200 | Slope | −9.884 | 0.060 | 0.065 | 1.780 | 0.065 | 0.012 | 9.219 | 0.556 | 0.595 |
| r2 | 0.074 | 0.034 | 0.058 | 0.097 | 0.076 | 0.002 | 0.099 | 0.119 | 0.024 | |
| 1200–1500 | Slope | −5.906 | 0.098 | 0.091 | 2.133 | 0.097 | 0.020 | 11.209 | 0.708 | 0.883 |
| r2 | 0.028 | 0.091 | 0.123 | 0.137 | 0.176 | 0.008 | 0.151 | 0.196 | 0.068 | |
| 1500–1800 | Slope | −7.701 | 0.096 | 0.076 | 1.620 | 0.066 | 0.065 | 5.534 | 0.314 | 0.879 |
| r2 | 0.047 | 0.091 | 0.092 | 0.074 | 0.098 | 0.066 | 0.034 | 0.041 | 0.067 | |
| 1800–2100 | Slope | −5.669 | 0.099 | 0.083 | 2.295 | 0.088 | 0.076 | 10.107 | 0.659 | 0.482 |
| r2 | 0.013 | 0.060 | 0.068 | 0.118 | 0.113 | 0.068 | 0.085 | 0.117 | 0.020 | |
| 2100–3000 | Slope | −11.126 | 0.009 | 0.025 | 1.380 | 0.044 | -0.062 | 3.900 | 0.357 | 0.186 |
| r2 | 0.056 | 0.001 | 0.010 | 0.037 | 0.039 | 0.040 | 0.010 | 0.030 | 0.007 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seeley, M.M.; Asner, G.P. Large-Scale Controls on the Leaf Economic Spectrum of the Overstory Tree Species Metrosideros polymorpha. Remote Sens. 2023, 15, 4707. https://doi.org/10.3390/rs15194707
Seeley MM, Asner GP. Large-Scale Controls on the Leaf Economic Spectrum of the Overstory Tree Species Metrosideros polymorpha. Remote Sensing. 2023; 15(19):4707. https://doi.org/10.3390/rs15194707
Chicago/Turabian StyleSeeley, Megan M., and Gregory P. Asner. 2023. "Large-Scale Controls on the Leaf Economic Spectrum of the Overstory Tree Species Metrosideros polymorpha" Remote Sensing 15, no. 19: 4707. https://doi.org/10.3390/rs15194707
APA StyleSeeley, M. M., & Asner, G. P. (2023). Large-Scale Controls on the Leaf Economic Spectrum of the Overstory Tree Species Metrosideros polymorpha. Remote Sensing, 15(19), 4707. https://doi.org/10.3390/rs15194707