




Vegetation Greenness Sensitivity to Precipitation and Its Oceanic and Terrestrial Component in Selected Biomes and Ecoregions of the World

 ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
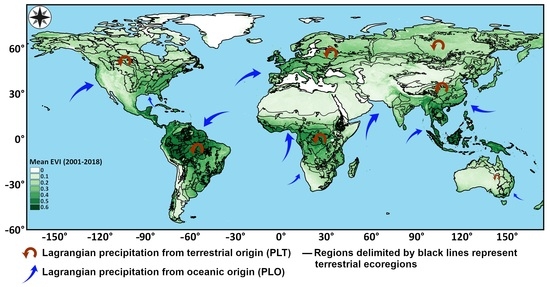
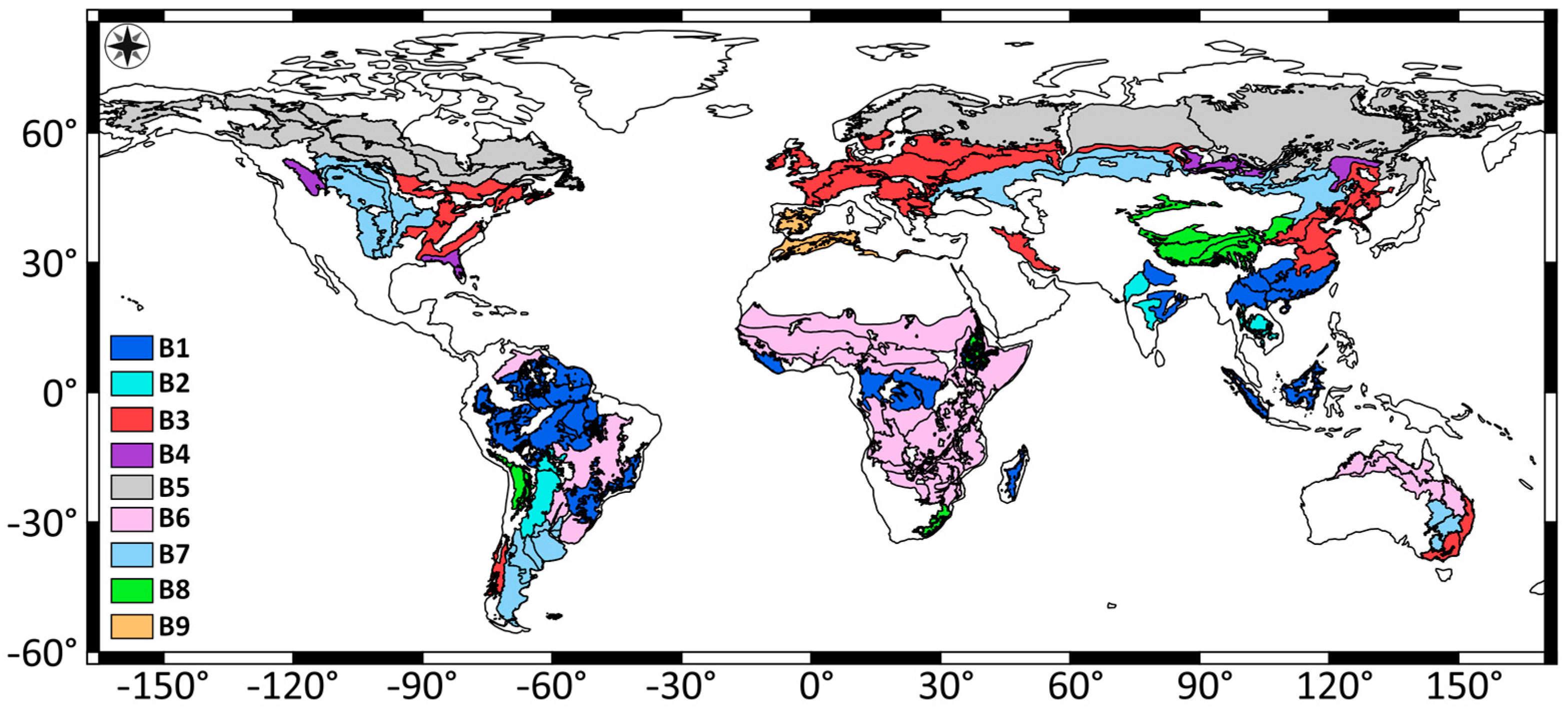
2.1. Study Regions
2.2. Data
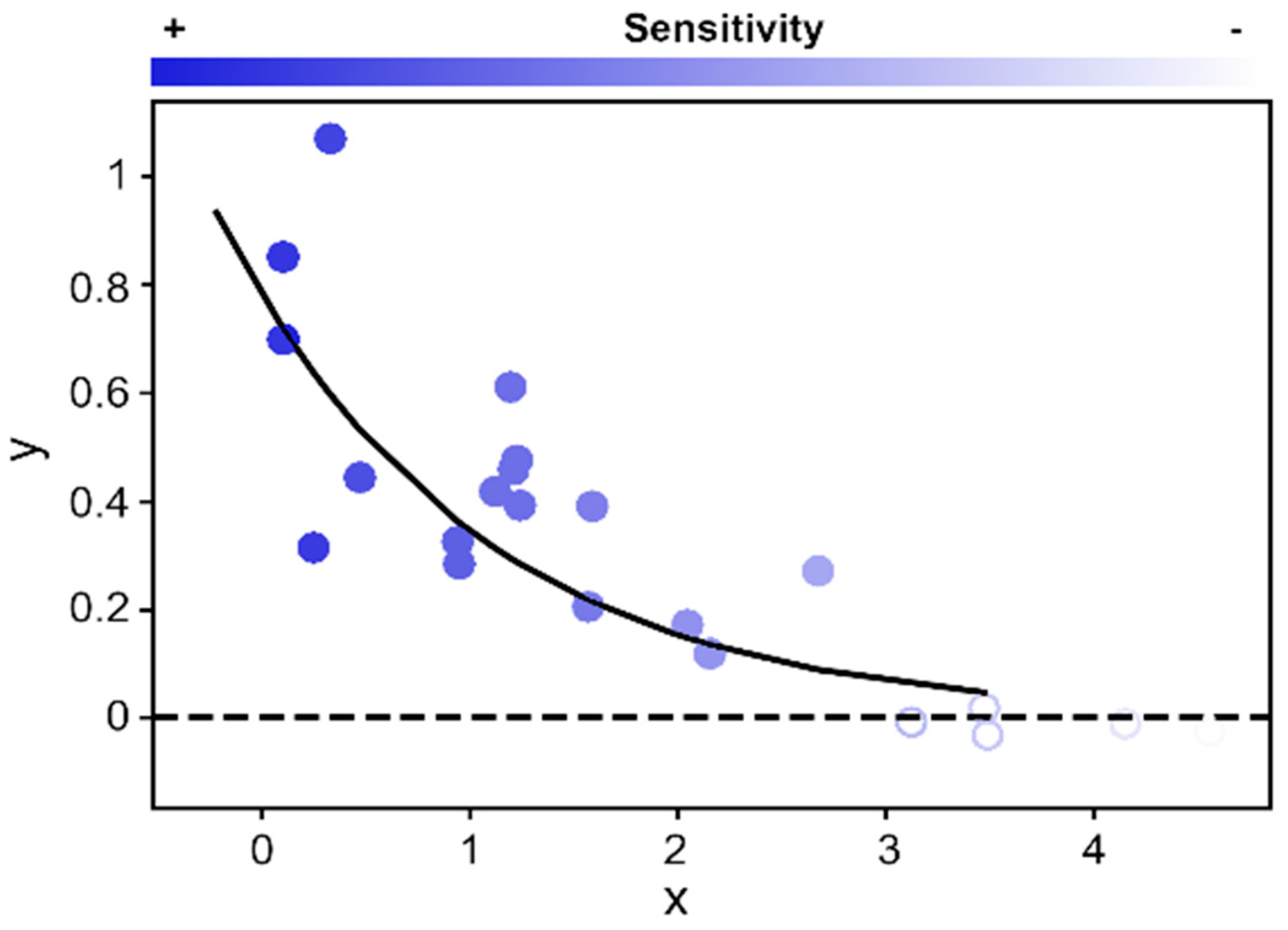
2.3. Vegetation Greenness Sensitivity (VGS) Metric and Statistical Analyses
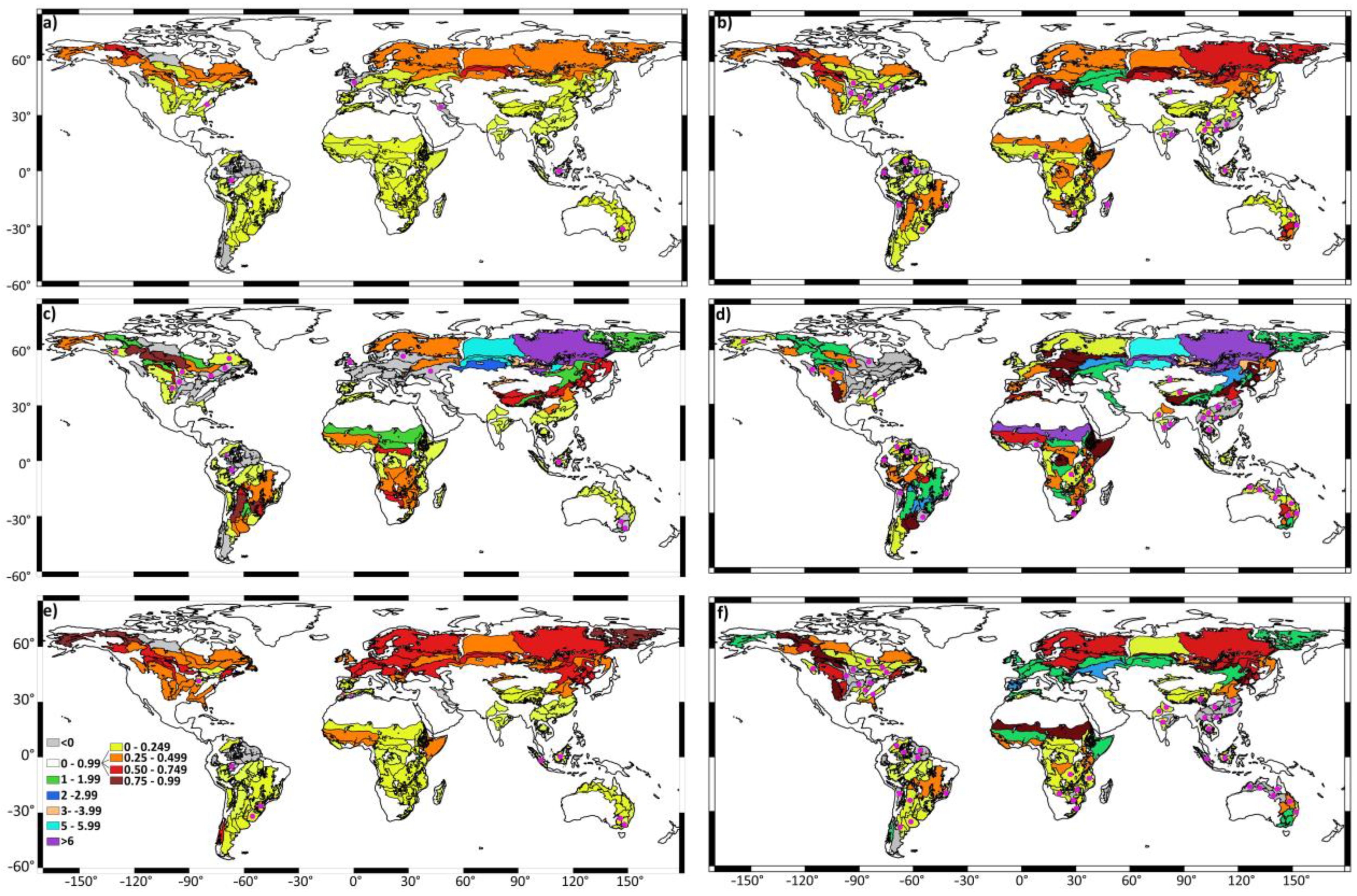
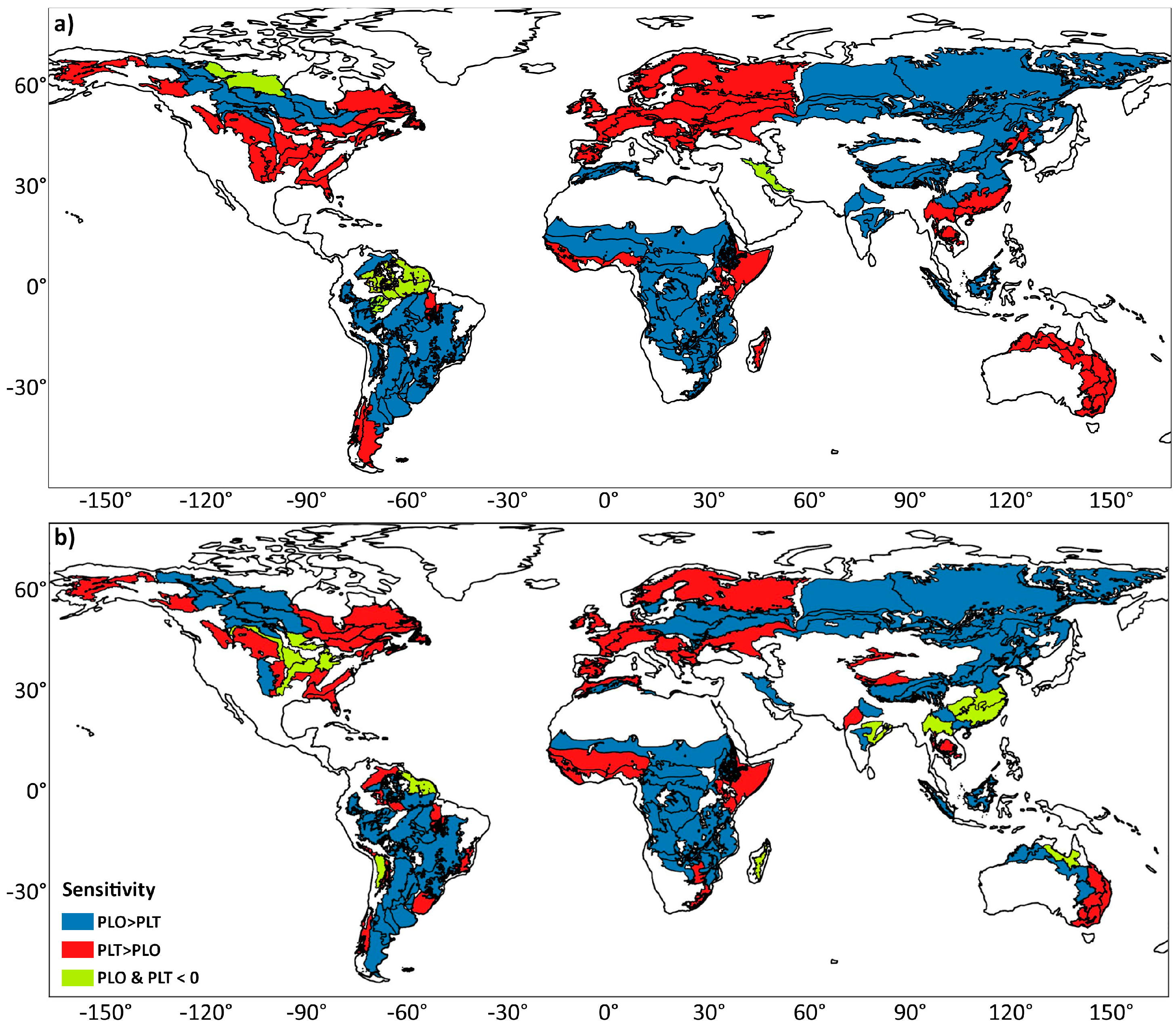
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Han, F.; Yan, J.; Ling, H.B. Variance of vegetation coverage and its sensitivity to climatic factors in the Irtysh River basin. PeerJ 2021, 9, e11334. [Google Scholar] [CrossRef]
- Li, M.; Song, Y.; Huang, X.; Li, J.; Mao, Y.; Zhu, T.; Cai, X.; Liu, B. Improving mesoscale modeling using satellite-derived land surface parameters in the Pearl River Delta region, China. J. Geophys. Res. Atmos. 2014, 119, 6325–6346. [Google Scholar] [CrossRef]
- Zhang, W.; Luo, G.; Chen, C.; Ochege, F.U.; Hellwich, O.; Zheng, H.; Hamdi, R.; Wu, S. Quantifying the Contribution of Climate Change and Human Activities to Biophysical Parameters in an Arid Region. Ecol. Indic. 2021, 129, 107996. [Google Scholar] [CrossRef]
- Wang, W.L.; Anderson, B.T.; Phillips, N.; Kaufmann, R.K.; Potter, C.; Myneni, R.B. Feedbacks of vegetation on summertime climate variability over the North American grasslands. Part I: Statistical analysis. Earth Interact. 2006, 10, 1–5. [Google Scholar] [CrossRef]
- Fay, P.A.; Kaufman, D.M.; Nippert, J.B.; Carlisle, J.D.; Harper, C.W. Changes in grassland ecosystem function due to extreme rainfall events: Implications for responses to climate change. Glob. Chang. Biol. 2008, 14, 1600–1608. [Google Scholar] [CrossRef]
- Kong, D.; Zhang, Q.; Singh, V.P.; Shi, P. Seasonal vegetation response to climate change in the Northern Hemi-sphere (1982–2013). Glob. Planet. Chang. 2017, 148, 1–8. [Google Scholar] [CrossRef]
- Weltzin, J.F.; Loik, M.E.; Schwinning, S.; Williams, D.G.; Fay, P.A.; Haddad, B.M.; Harte, J.; Huxman, T.E.; Knapp, A.K.; Lin, G.; et al. Assessing the response of terrestrial ecosystems to potential changes in precipitation. Bioscience 2003, 53, 941–952. [Google Scholar] [CrossRef]
- Zhao, F.; Wu, Y.; Sivakumar, B.; Long, A.; Qiu, L.; Chen, J.; Wang, L.; Liu, S.; Hu, H. Climatic and Hydrologic Controls on Net Primary Production in a Semiarid Loess Watershed. J. Hydrol. 2019, 568, 803–815. [Google Scholar] [CrossRef]
- Piao, S.; Mohammat, A.; Fang, J.; Cai, Q.; Feng, J. NDVI-based increase in growth of temperate grasslands and its responses to climate changes in China. Glob. Environ. Chang. 2006, 16, 340–348. [Google Scholar] [CrossRef]
- Wang, J.; Wang, K.; Zhang, M.; Zhang, C. Impacts of climate change and human activities on vegetation cover in hilly southern China. Ecol. Eng. 2015, 81, 451–461. [Google Scholar] [CrossRef]
- Zheng, K.; Wei, J.; Pei, J.; Cheng, H.; Zhang, X.; Huang, F.; Li, F.; Ye, J. Impacts of climate change and human activities on grassland vegetation variation in the Chinese Loess Plateau. Sci. Total Environ. 2019, 660, 236–244. [Google Scholar] [CrossRef]
- Papagiannopoulou, C.; Miralles, D.G.; Dorigo, W.A.; Verhoest, N.E.C.; Depoorter, M.; Waegeman, W. Vegetation anomalies caused by antecedent precipitation in most of the world. Environ. Res. Lett. 2017, 12, 074016. [Google Scholar] [CrossRef]
- Maurer, G.E.; Hallmark, A.J.; Brown, R.F.; Sala, O.E.; Collins, S.L. Sensitivity of primary production to precipitation across the United States. Ecol. Lett. 2020, 23, 527–536. [Google Scholar] [CrossRef] [PubMed]
- Ayanlade, A.; Jeje, O.D.; Nwaezeigwe, J.O.; Orimoogunje, O.O.I.; Olokeogun, O.S. Rainfall seasonality effects on vegetation greenness in different ecological zones. Environ. Chall. 2021, 4, 100144. [Google Scholar] [CrossRef]
- Soomro, S.; Hu, C.; Jian, S.; Wu, Q.; Boota, M.W.; Soomro, M.H.A.A. Precipitation changes and their relationships with vegetation responses during 1982–2015 in Kunhar River basin, Pakistan. Water Supply 2021, 21, 3657–3671. [Google Scholar] [CrossRef]
- Wang, J.; Rich, P.M.; Price, K.P. Temporal responses of NDVI to precipitation and temperature in the central Great Plains, USA. Int. J. Remote Sens. 2003, 24, 2345–2364. [Google Scholar] [CrossRef]
- Hawinkel, P.; Thiery, W.; Hermitte, S.L.; Swinnen, E.; Verbist, B.; Van Orshoven, J.; Muys, B. Vegetation response to precipitation variability in East Africa controlled by biogeographical factors. J. Geophys. Res. Biogeosci. 2016, 121, 2422–2444. [Google Scholar] [CrossRef]
- Jiang, L.; Jiapaer, G.; Bao, A.; Guo, H.; Ndayisaba, F. Vegetation dynamics and responses to climate change and human activities in Central Asia. Sci. Total Environ. 2017, 599–600, 967–980. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, W.; Fu, J. Vegetation response to precipitation anomalies under different climatic and biogeographical conditions in China. Sci. Rep. 2020, 10, 830. [Google Scholar] [CrossRef]
- Nightingale, J.M.; Phinn, S.R. Assessment of relationships between precipitation and satellite derived vegetation condition within South Australia. Aust. Geogr. Stud. 2003, 41, 180–195. [Google Scholar] [CrossRef]
- Nemani, R.R.; Keeling, C.D.; Hashimoto, H.; Jolly, W.M.; Piper, S.C.; Tucker, C.J.; Myneni, R.B.; Running, S.W. Climate-driven increases in global terrestrial net primary production from 1982 to 1999. Science 2003, 300, 1560–1563. [Google Scholar] [CrossRef] [PubMed]
- Piao, S.; Fang, J.; Zhou, L.; Guo, Q.; Henderson, M.; Ji, W.; Li, Y.; Tao, S. Interannual variations of monthly and seasonal normalized difference vegetation index (NDVI) in China from 1982 to 1999. J. Geophys. Res. 2003, 108, 4401. [Google Scholar] [CrossRef]
- Wu, D.; Zhao, X.; Liang, S.; Zhou, T.; Huang, K.; Tang, B.; Zhao, W. Time-lag effects of global vegetation responses to climate change. Glob. Chang. Biol. 2015, 21, 3520–3531. [Google Scholar] [CrossRef] [PubMed]
- Zhe, M.; Zhang, X. Time-lag effects of NDVI responses to climate change in the Yamzhog Yumco Basin, South Tibet. Ecol. Indic. 2021, 124, 107431. [Google Scholar] [CrossRef]
- Huete, A.R.; Liu, H.Q.; Batchily, K.; van Leeuwen, W. A comparison of vegetation indices over a global set of TM images for EOS-MODIS. Remote Sens. Environ. 1997, 59, 440–451. [Google Scholar] [CrossRef]
- Wardlow, B.D.; Egbert, S.L.; Kastens, J.H. Analysis of time-series MODIS 250m vegetation index data for crop classification in the U.S. Central Great Plains. Remote Sens. Environ. 2007, 108, 290–310. [Google Scholar] [CrossRef]
- Chuai, X.W.; Huang, X.J.; Wang, W.J.; Bao, G. NDVI, temperature and precipitation changes and their relationships with different vegetation types during 1998–2007 in Inner Mongolia, China. Int. J. Climatol. 2013, 33, 1696–1706. [Google Scholar] [CrossRef]
- Nielsen, U.N.; Ball, B.A. Impacts of altered precipitation regimes on soil communities and biogeochemistry in arid and semi-arid ecosystems. Glob. Chang. Biol. 2015, 21, 1407–1421. [Google Scholar] [CrossRef]
- Gao, J.; Jiao, K.; Wu, S.; Ma, D.; Zhao, D.; Yin, Y.; Dai, E. Past and future effects of climate change on spatially heterogeneous vegetation activity in China. Earth’s Future 2017, 5, 679–692. [Google Scholar] [CrossRef]
- Weiss, J.L.; Gutzler, D.S.; Coonrod, J.E.A.; Dahm, C.N. Long-term vegetation monitoring with NDVI in a diverse semi-arid setting, central New Mexico, USA. J. Arid Environ. 2004, 58, 249–272. [Google Scholar] [CrossRef]
- Wang, J.; Price, K.P.; Rich, P.M. Spatial patterns of NDVI in response to precipitation and temperature in the central Great Plains. Int. J. Remote Sens. 2001, 22, 3827–3844. [Google Scholar] [CrossRef]
- Piao, S.; Friedlingstein, P.; Ciais, P.; Viovy, N.; Demarty, J. Growing season extension and its impact on terrestrial carbon cycle in the Northern Hemisphere over the past 2 decades. Glob. Biogeochem. Cycles 2007, 21, GB3018. [Google Scholar] [CrossRef]
- Gaughan, A.; Stevens, F. Linking vegetation response to seasonal precipitation in the Okavango–Kwando–Zambezi catchment of southern Africa. Int. J. Remote Sens. 2012, 33, 37–41. [Google Scholar] [CrossRef]
- Wong, W.F.J. Spatial and temporal analysis of MODIS EVI and TRMM 3B43 rainfall retrievals in Australia. In Proceedings of the 19th International Conference on Geoinformatics, Shanghai, China, 24–26 June 2006; pp. 1–7. [Google Scholar] [CrossRef]
- Zhang, Q.; Kong, D.; Singh, V.P.; Shi, P. Response of vegetation to diferent time-scales drought across China: Spatiotemporal patterns, causes and implications. Glob. Planet. Chang. 2017, 152, 1–11. [Google Scholar] [CrossRef]
- Lee, J.; Frankenberg, C.; Berry, J.A.; Boyce, C.K.; Fisher, J.B.; Morrow, R.; Worden, J.R.; Asefi, S.; Badgley, G.; Saatchi, S. Forest productivity and water stress in Amazonia: Observations from GOSAT chlorophyll fluorescence. Proc. R. Soc. B Biol. Sci. 2013, 280, 10823–10827. [Google Scholar] [CrossRef]
- Restrepo-Coupe, N.; da Rocha, H.R.; Hutyra, L.R.; da Araujo, A.C.; Borma, L.S.; Christoffersen, B.; Cabral, O.M.R.; de Camargo, P.B.; Cardoso, F.L.; Saleska, S.R.; et al. What drives the seasonality of photosynthesis across the Amazon basin? A cross-site analysis of eddy flux tower measurements from the Brasil flux network. Agric. For. Meteorol. 2013, 182–183, 128–144. [Google Scholar] [CrossRef]
- Hof, A.R.; Dymond, C.C.; Mladenoff, D.J. Climate change mitigation through adaptation: The effectiveness of forest diversification by novel tree planting regimes. Ecosphere 2017, 8, e01981. [Google Scholar] [CrossRef]
- Gallagher, R.V.; Allen, S.; Wright, I.J. Safety margins and adaptive capacity of vegetation to climate change. Sci. Rep. 2019, 9, 8241. [Google Scholar] [CrossRef]
- Gimeno, L.; Nieto, R.; Sorí, R. The growing importance of oceanic moisture sources for continental precipitation. NPJ Clim. Atmos. Sci. 2020, 3, 27. [Google Scholar] [CrossRef]
- Tietjen, B.; Schlaepfer, D.R.; Bradford, J.B.; Lauenroth, W.K.; Hall, S.A.; Duniway, M.C.; Hochstrasser, T.; Jia, G.; Munson, S.M.; Pyke, D.A.; et al. Climate change-induced vegetation shifts lead to more ecological droughts despite projected rainfall increases in many global temperate drylands. Glob. Chang. Biol. 2017, 23, 2743–2754. [Google Scholar] [CrossRef]
- Keys, P.W.; Wang-Erlandsson, L.; Gordon, L.J. Revealing Invisible Water: Moisture Recycling as an Ecosystem Service. PLoS ONE 2016, 11, e0151993. [Google Scholar] [CrossRef]
- van der Ent, R.J.; Savenije, H.H.G.; Schaefli, B.; Steele-Dunne, S.C. Origin and fate of atmospheric moisture over continents. Water Resour. Res. 2010, 46, W09525. [Google Scholar] [CrossRef]
- Drumond, A.; Marengo, J.; Ambrizzi, T.; Nieto, R.; Moreira, L.; Gimeno, L. The role of the Amazon Basin moisture in the atmospheric branch of the hydrological cycle: A Lagrangian analysis. Hydrol. Earth Syst. Sci. 2014, 18, 2577–2598. [Google Scholar] [CrossRef]
- Chug, D.; Dominguez, F.; Yang, Z. The Amazon and La Plata river basins as moisture sources of South America: Climatology and intraseasonal variability. J. Geophys. Res. Atmos. 2022, 127, e2021JD035455. [Google Scholar] [CrossRef]
- Sorí, R.; Nieto, R.; Vicente-Serrano, S.M.; Drumond, A.; Gimeno, L. A Lagrangian perspective of the hydrological cycle in the Congo River basin. Earth Syst. Dynam. 2017, 8, 653–675. [Google Scholar] [CrossRef]
- Zemp, D.C.; Schleussner, C.F.; Barbosa, H.M.J.; van der Ent, R.J.; Donges, J.F.; Heinke, J.; Sampaio, G.; Rammig, A. On the importance of cascading moisture recycling in South America. Atmos. Chem. Phys. 2014, 14, 13337–13359. [Google Scholar] [CrossRef]
- Liu, N.; Harper, R.J.; Dell, B.; Liu, S.; Yu, Z. Vegetation dynamics and rainfall sensitivity for different vegetation types of the Australian continent in the dry period 2002–2010. Ecohydrology 2017, 10, e1811. [Google Scholar] [CrossRef]
- Claessen, J.; Martens, B.; Verhoest, N.E.C.; Molini, A.; Miralles, D.G. Global climatic drivers of vegetation based on wavelet analysis. In Proceedings of the 9th International Workshop on the Analysis of Multitemporal Remote Sensing Images (MultiTemp), Brugge, Belgium, 27–29 June 2017; pp. 1–3. [Google Scholar] [CrossRef]
- Sharma, M.; Bangotra, P.; Gautam, A.S.; Gautam, S. Sensitivity of normalized difference vegetation index (NDVI) to land surface temperature, soil moisture and precipitation over district Gautam Buddh Nagar, UP, India. Stoch. Environ. Res. Risk Assess 2022, 36, 1779–1789. [Google Scholar] [CrossRef]
- Zhang, Y.; Gentine, P.; Luo, X.; Lian, X.; Liu, Y.; Zhou, S.; Michalak, A.M.; Sun, W.; Fisher, J.B.; Piao, S.; et al. Increasing sensitivity of dryland vegetation greenness to precipitation due to rising atmospheric CO2. Nat. Commun. 2022, 13, 4875. [Google Scholar] [CrossRef]
- Drumond, A.; Stojanovic, M.; Nieto, R.; Vicente-Serrano, S.M.; Gimeno, L. Linking Anomalous Moisture Transport and Drought Episodes in the IPCC Reference Regions. Bull. Am. Meteorol. Soc. 2019, 100, 1481–1498. [Google Scholar] [CrossRef]
- Herrera-Estrada, J.E.; Diffenbaugh, N.S. Landfalling droughts: Global tracking of moisture deficits from the oceans onto land. Water Resour. Res. 2020, 56, e2019WR026877. [Google Scholar] [CrossRef]
- Sorí, R.; Gimeno-Sotelo, L.; Nieto, R.; Liberato, M.L.R.; Stojanovic, M.; Pérez-Alarcón, A.; Fernández-Alvarez, J.C.; Gimeno, L. Oceanic and terrestrial origin of precipitation over 50 major world river basins: Implications for the occurrence of drought. Sci. Total Environ. 2023, 859, 160288. [Google Scholar] [CrossRef] [PubMed]
- Erlingis, J.M.; Gourley, J.J.; Basara, J.B. Diagnosing Moisture Sources for Flash Floods in the United States. Part II: Terrestrial and Oceanic Sources of Moisture. J. Hydrometeor. 2019, 20, 1511–1531. [Google Scholar] [CrossRef]
- Vázquez, M.; Nieto, R.; Liberato, M.L.R.; Gimeno, L. A data base of contributions of major oceanic and terrestrial moisture sources on continental daily extreme precipitation. Data Brief. 2021, 35, 106830. [Google Scholar] [CrossRef] [PubMed]
- Krug, A.; Aemisegger, F.; Sprenger, M.; Ahrens, B. Moisture sources of heavy precipitation in Central Europe in synoptic situations with Vb-cyclones. Clim. Dyn. 2022, 59, 3227–3245. [Google Scholar] [CrossRef]
- IPCC. 2023: Climate Change 2023: Synthesis Report. A Report of the Intergovernmental Panel on Climate Change. In Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Core Writing Team, Lee, H., Romero, J., Eds.; IPCC: Geneva, Switzerland, 2023; in press. [Google Scholar]
- Hsu, J.S.; Powell, J.; Adler, P.B. Sensitivity of mean annual primary production to precipitation. Glob. Chang. Biol. 2012, 18, 2246–2255. [Google Scholar] [CrossRef]
- Gang, C.; Wang, Z.; Zhou, W.; Chen, Y.; Li, J.; Cheng, J.; Guo, L.; Odeh, I.; Chen, C. Projecting the dynamics of terrestrial net primary productivity in response to future climate change under the RCP2.6 scenarios. Environ. Earth Sci. 2015, 74, 5949–5959. [Google Scholar] [CrossRef]
- Cao, D.; Zhang, J.; Han, J.; Zhang, T.; Yang, S.; Wang, J.; Prodhan, F.A.; Yao, F. Projected increases in global terrestrial net primary productivity loss caused by drought under climate change. Earth’s Future 2022, 10, e2022EF002681. [Google Scholar] [CrossRef]
- Findell, K.L.; Keys, P.W.; van der Ent, R.J.; Lintner, B.R.; Berg, A.; Krasting, J.P. Rising Temperatures Increase Importance of Oceanic Evaporation as a Source for Continental Precipitation. J. Clim. 2019, 32, 7713–7726. [Google Scholar] [CrossRef]
- Zeng, Z.; Piao, S.; Lin, X.; Yin, G.; Peng, S.; Ciais, P.; Myneni, R.B. Global evapotranspiration over the past three decades: Estimation based on the water balance equation combined with empirical models. Environ. Res. Lett. 2012, 7, 014026. [Google Scholar] [CrossRef]
- Wang, R.; Li, L.; Gentine, P.; Zhang, Y.; Chen, J.; Chen, X.; Chen, L.; Ning, L.; Yuan, L.; Lü, G. Recent increase in the observation-derived land evapotranspiration due to global warming. Environ. Res. Lett. 2022, 17, 024020. [Google Scholar] [CrossRef]
- Eltahir, E.A.B.; Bras, R.L. Precipitation recycling in the Amazon basin. Q. J. R. Meteorol. Soc. 1994, 120, 861–880. [Google Scholar] [CrossRef]
- Sorí, R.; Marengo, J.; Nieto, R.; Drumond, A.; Gimeno, L. The Atmospheric Branch of the Hydrological Cycle over the Negro and Madeira River Basins in the Amazon Region. Water 2018, 10, 738. [Google Scholar] [CrossRef]
- Worden, S.; Fu, R.; Chakraborty, S.; Liu, J.; Worden, J. Where does moisture come from over the Congo Basin? J. Geophys. Res. Biogeosci. 2021, 126, e2020JG006024. [Google Scholar] [CrossRef]
- Hasnat, G.N.T.; Hossain, M.K. Global overview of tropical dry forests. In Handbook of Research on the Conservation and Restoration of Tropical Dry Forests; Bhadouria, R., Tripathi, S., Srivastava, P., Singh, P., Eds.; IGI Global: Hershey, PA, USA, 2020; pp. 1–23. [Google Scholar] [CrossRef]
- Olson, D.M.; Dinerstein, E.; Wikramanayake, E.D.; Burgess, N.D.; Powell, G.V.N.; Underwood, E.C.; D’amico, J.A.; Itoua, I.; Strand, H.E.; Morrison, J.C. Terrestrial Ecoregions of the World: A New Map of Life on Earth: A new global map of terrestrial ecoregions provides an innovative tool for conserving biodiversity. BioScience 2001, 51, 933–938. [Google Scholar] [CrossRef]
- Keith, H.; Mackey, B.G.; Lindenmayer, D.B. Re-evaluation of forest biomass carbon stocks and lessons from the world’s most carbon-dense forests. Proc. Natl. Acad. Sci. USA 2009, 106, 11635–11640. [Google Scholar] [CrossRef]
- Olson, D.M.; Dinerstein, E. The Global 200: Priority ecoregions for global conservation. Ann. Mo. Bot. Gard. 2002, 89, 199–224. [Google Scholar] [CrossRef]
- Harris, I.; Osborn, T.J.; Jones, P.; Lister, D. Version 4 of the CRU TS monthly high-resolution gridded multivariate climate dataset. Sci. Data 2020, 7, 109. [Google Scholar] [CrossRef]
- Dee, D.P.; Uppala, S.M.; Simmons, A.J.; Berrisford, P.; Poli, P.; Kobayashi, S.; Andrae, U.; Balmaseda, M.A.; Balsamo, G.; Bauer, P.; et al. The ERA-interim reanalysis: Configuration and performance of the data assimilation system. Q. J. R. Meteorol. Soc. 2011, 137, 553–597. [Google Scholar] [CrossRef]
- Hernandez-Stefanoni, J.L.; Ponce-Hernandez, R. Mapping the Spatial Variability of Plant Diversity in a Tropical Forest: Comparison of Spatial Interpolation Methods. Environ. Monit. Assess 2006, 117, 307–334. [Google Scholar] [CrossRef]
- Munyati, C.; Sinthumule, N.I. Comparative suitability of ordinary kriging and Inverse Distance Weighted interpolation for indicating intactness gradients on threatened savannah woodland and forest stands. Environ. Sustain. Indic. 2021, 12, 100151. [Google Scholar] [CrossRef]
- Hersbach, H.; Bell, B.; Berrisford, P.; Hirahara, S.; Horányi, A.; Muñoz-Sabater, J.; Thépaut, J.N. The ERA5 global reanalysis. Q. J. R. Meteorol. Soc. 2020, 146, 1999–2049. [Google Scholar] [CrossRef]
- Nieto, R.; Gimeno, L. A database of optimal integration times for Lagrangian studies of atmospheric moisture sources and sinks. Sci. Data 2019, 6, 59. [Google Scholar] [CrossRef] [PubMed]
- Nieto, R.; Gimeno, L. Addendum: A database of optimal integration times for Lagrangian studies of atmospheric moisture sources and sinks. Sci. Data 2021, 8, 130. [Google Scholar] [CrossRef] [PubMed]
- Stohl, A.; Forster, C.; Frank, A.; Seibert, P.; Wotawa, G. Technical note: The Lagrangian particle dispersion model FLEXPART version 6.2. Atmos. Chem. Phys. 2005, 5, 2461–2474. [Google Scholar] [CrossRef]
- Stohl, A.; James, P. A Lagrangian analysis of the atmospheric branch of the global water cycle. Part I: Method description, validation, and demonstration for the August 2002 flooding in central Europe. J. Hydrometeorol. 2004, 5, 656–678. [Google Scholar] [CrossRef]
- Didan, K.; Munoz, A.B. MODIS Vegetation Index User’s Guide (MOD13 Series) Version 3.10, September 2019. (Collection 6.1). Available online: https://lpdaac.usgs.gov/documents/621/MOD13_User_Guide_V61.pdf (accessed on 16 June 2023).
- Camberlin, P.; Martiny, N.; Phillipon, N.; Richard, Y. Determinants of the interannual relationships between remote sensed photosynthetic activity and rainfall in tropical Africa. Remote Sens. Environ. 2007, 106, 199–216. [Google Scholar] [CrossRef]
- Forzieri, G.; Feyen, L.; Cescatti, A.; Vivoni, E.R. Spatial and temporal variations in ecosystem response to monsoon precipitation variability in southwestern North America. J. Geophys. Res. 2014, 119, 1999–2017. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Jiang, Z.; Huete, A.; Didan, K.; Miura, T. Development of a two-band enhanced vegetation index without a blue band. Remote Sens. Environ. 2008, 112, 3833–3845. [Google Scholar] [CrossRef]
- Bari, E.; Nipa, N.J.; Roy, B. Association of vegetation indices with atmospheric & biological factors using MODIS time series products. Environ. Chall. 2021, 5, 100376. [Google Scholar] [CrossRef]
- Chen, X.L.; Zhao, H.M.; Li, P.X.; Yin, Z.Y. Remote sensing image-based analysis of the relationship between urban heat island and land use/cover changes. Remote Sens. Environ. 2006, 104, 133–146. [Google Scholar] [CrossRef]
- Ayanlade, A. Remote sensing vegetation dynamics analytical methods: A review of vegetation indices techniques. Geoinform. Pol. 2017, 16, 7–17. [Google Scholar] [CrossRef]
- Alexandridis, T.K.; Ovakoglou, G.; Clevers, J.G.P.W. Relationship between MODIS EVI and LAI across time and space. Geocarto Int. 2020, 35, 1385–1399. [Google Scholar] [CrossRef]
- Guo, N.; Wang, X.; Cai, D.; Yang, J. Comparison and evaluation between MODIS vegetation indices in Northwest China. In Proceedings of the IEEE International Geoscience and Remote Sensing Symposium (IGARSS), Barcelona, Spain, 23–27 July 2007; p. 3366. [Google Scholar] [CrossRef]
- Huxman, T.E.; Snyder, K.A.; Tissue, D.; Joshua Leffler, A.; Ogle, K.; Pockman, W.T.; Sandquist, D.R.; Potts, D.L.; Schwinning, S. Precipitation pulses and carbon fluxes in semiarid and arid ecosystems. Oecologia 2004, 141, 254–268. [Google Scholar] [CrossRef]
- Chamaillé-Jammes, S.; Fritz, H. Precipitation-NDVI relationships in eastern and southern African savannas vary along a precipitation gradient. Int. J. Remote Sens. 2009, 30, 3409–3422. [Google Scholar] [CrossRef]
- Wu, Z.; Ahlström, A.; Smith, B.; Ardö, J.; Eklundh, L.; Fensholt, R.; Lehsten, V. Climate data induced uncertainty in model-based estimations of terrestrial primary productivity. Environ. Res. Lett. 2017, 12, 064013. [Google Scholar] [CrossRef]
- Guo, L.; van der Ent, R.J.; Klingaman, N.P.; Demory, M.; Vidale, P.L.; Turner, A.G.; Stephan, C.C.; Chevuturi, A. Moisture Sources for East Asian Precipitation: Mean Seasonal Cycle and Interannual Variability. J. Hydrometeor. 2019, 20, 657–672. [Google Scholar] [CrossRef]
- Felton, A.J.; Zavislan-Pullaro, S.; Smith, M.D. Semiarid ecosystem sensitivity to precipitation extremes: Weak evidence for vegetation constraints. Ecology 2019, 100, e02572. [Google Scholar] [CrossRef]
- Hilker, T.; Lyapustin, A.I.; Tucker, C.J.; Sellers, C.J. Vegetation dynamics and rainfall sensitivity of the Amazon. Biol. Sci. 2014, 111, 16041–16046. [Google Scholar] [CrossRef]
- Giardina, F.; Konings, A.G.; Kennedy, D.; Alemohammad, S.H.; Oliveira, R.S.; Uriarte, M.; Gentine, P. Tall Amazonian forests are less sensitive to precipitation variability. Nat. Geosci. 2018, 11, 405–409. [Google Scholar] [CrossRef]
- Chen, Z.; Liu, H.; Xu, C.; Wu, X.; Liang, B.; Cao, J.; Chen, D. Modeling vegetation greenness and its climate sensitivity with deep-learning technology. Ecol. Evol. 2021, 11, 7335–7345. [Google Scholar] [CrossRef]
- McVicar, T.R.; Roderick, M.L.; Donohue, R.J.; Li, L.T.; Van Niel, T.G.; Thomas, A.; Grierer, J.; Jhajharia, D.; Himri, Y.; Mahowald, N.M.; et al. Global review and synthesis of trends in observedterrestrial near-surface wind speeds: Implications for evaporation. J. Hydrol. 2012, 416–417, 182–205. [Google Scholar] [CrossRef]
- Bao, Z.; Zhang, J.; Wang, G.; Guan, T.; Jin, J.; Liu, Y.; Li, M.; Ma, T. The sensitivity of vegetation cover to climate change in multiple climatic zones using machine learning algorithms. Ecol. Indic. 2021, 124, 107443. [Google Scholar] [CrossRef]
- Seddon, A.W.R.; Macias-Fauria, M.; Long, P.R.; Benz, D.; Willis, K.J. Sensitivity of global terrestrial ecosystems to climate variability. Nature 2016, 531, 229–232. [Google Scholar] [CrossRef]
- Jiang, P.; Ding, W.; Yuan, Y.; Ye, W.; Mu, Y. Interannual variability of vegetation sensitivity to climate in China. J. Environ. Manag. 2022, 301, 113768. [Google Scholar] [CrossRef]
- Koirala, S.; Jung, M.; Reichstein, M.; de Graaf, I.E.; Camps-Valls, G.; Ichii, K.; Carvalhais, N. Global distribution of groundwater-vegetation spatial covariation. Geophys. Res. Lett. 2017, 44, 4134–4142. [Google Scholar] [CrossRef]
- Zhang, W.; Li, H.; Puepkke, S.G.; Diao, Y.; Nie, X.; Geng, J.; Chen, D.; Pang, J. Nutrient loss is sensitive to land cover changes and slope gradients of agricultural hillsides: Evidence from four contrasting pond systems in a hilly catchment. Agric. Water Manag. 2020, 237, 106165. [Google Scholar] [CrossRef]
- Ilvonen, L.; López-Sáez, J.A.; Holmström, L.; Alba-Sánchez, F.; Pérez-Díaz, S.; Carrión, J.S.; Ramos-Román, M.J.; Camuera, J.; Jiménez-Moreno, G.; Ruha, L.; et al. Spatial and temporal patterns of Holocene precipitation change in the Iberian Peninsula. Boreas 2022, 51, 776–792. [Google Scholar] [CrossRef]
- Lélé, M.I.; Leslie, L.M.; Lamb, P.J. Analysis of Low-Level Atmospheric Moisture Transport Associated with the West African Monsoon. J. Clim. 2015, 28, 4414–4430. [Google Scholar] [CrossRef]
- Niang, C.; Mancho, A.M.; García-Garrido, V.J.; Mohino, E.; Rodriguez-Fonseca, B.; Curbelo, J. Transport pathways across the West African Monsoon as revealed by Lagrangian Coherent Structures. Sci. Rep. 2020, 10, 12543. [Google Scholar] [CrossRef]
- Viste, E.; Sorteberg, A. Moisture transport into the Ethiopian highlands. Int. J. Climatol. 2013, 33, 249–263. [Google Scholar] [CrossRef]
- Stojanovic, M.; Mehabie Mulualem, G.; Sorí, R.; Vázquez, M.; Nieto, R.; Gimeno, L. Precipitation Moisture Sources of Ethiopian River Basins and Their Role During Drought Conditions. Front. Earth Sci. 2022, 10, 1–17. [Google Scholar] [CrossRef]
- Keys, P.W.; Barnes, E.A.; van der Ent, R.J.; Gordon, L.J. Variability of moisture recycling using a precipitationshed framework. Hydrol. Earth Syst. Sci. 2014, 18, 3937–3950. [Google Scholar] [CrossRef]
- Batibeniz, F.; Ashfaq, M.; Önol, B.; Utku Turuncoglu, U.; Mehmood, S.; Evans, K.J. Identification of major moisture sources across the Mediterranean Basin. Clim. Dyn. 2020, 54, 4109–4127. [Google Scholar] [CrossRef]
- Papritz, L.; Hauswirth, D.; Hartmuth, K. Moisture origin, transport path-ways, and driving processes of intense wintertime moisture transport into the Arctic. Weather Clim. Dynam. 2022, 3, 1–20. [Google Scholar] [CrossRef]
- Quetin, G.R.; Swann, A.L.S. Empirically Derived Sensitivity of Vegetation to Climate across Global Gradients of Temperature and Precipitation. J. Clim. 2017, 30, 5835–5849. [Google Scholar] [CrossRef]
- Rathore, S.; Bindoff, N.L.; Ummenhofer, C.C.; Phillips, H.E.; Feng, M. Near-Surface Salinity Reveals the Oceanic Sources of Moisture for Australian Precipitation through Atmospheric Moisture Transport. J. Clim. 2020, 33, 6707–6730. [Google Scholar] [CrossRef]
- Winkler, A.J.; Myneni, R.B.; Hannart, A.; Sitch, A.; Haverd, V.; Lombardozzi, D.; Arora, V.K.; Pongratz, J.; Nabel, J.E.M.S.; Goll, D.S.; et al. Slowdown of the greening trend in natural vegetation with further rise in atmospheric CO2. Biogeosciences 2021, 18, 4985–5010. [Google Scholar] [CrossRef]
- Theeuwen, J.J.E.; Staal, A.; Tuinenburg, O.A.; Hamelers, B.V.M.; Dekker, S.C. Local moisture recycling across the globe. Hydrol. Earth Syst. Sci. 2023, 27, 1457–1476. [Google Scholar] [CrossRef]
- Benestad, R.E.; Lussana, C.; Lutz, J.; Dobler, A.; Landgren, O.; Haugen, J.E.; Mezghani, A.; Casati, B.; Parding, K.M. Global hydroclimatological indicators and changes in the global hydrological cycle and rainfall patterns. PLoS Clim. 2022, 1, e0000029. [Google Scholar] [CrossRef]
- Chen, Z.; Zhou, T.; Zhang, W.; Li, P.; Zhao, S. Projected changes in the annual range of precipitation under stabilized 1.5 °C and 2.0 °C warming futures. Earth’s Future 2020, 8, e2019EF001435. [Google Scholar] [CrossRef]
- Li, W.; Migliavacca, M.; Forkel, M.; Denissen, J.M.C.; Reichstein, M.; Yang, H.; Duveiller, G.; Weber, U.; Orth, R. Widespread increasing vegetation sensitivity to soil moisture. Nat. Commun. 2022, 13, 3959. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stojanovic, M.; Sorí, R.; Guerova, G.; Vázquez, M.; Nieto, R.; Gimeno, L. Vegetation Greenness Sensitivity to Precipitation and Its Oceanic and Terrestrial Component in Selected Biomes and Ecoregions of the World. Remote Sens. 2023, 15, 4706. https://doi.org/10.3390/rs15194706
Stojanovic M, Sorí R, Guerova G, Vázquez M, Nieto R, Gimeno L. Vegetation Greenness Sensitivity to Precipitation and Its Oceanic and Terrestrial Component in Selected Biomes and Ecoregions of the World. Remote Sensing. 2023; 15(19):4706. https://doi.org/10.3390/rs15194706
Chicago/Turabian StyleStojanovic, Milica, Rogert Sorí, Guergana Guerova, Marta Vázquez, Raquel Nieto, and Luis Gimeno. 2023. "Vegetation Greenness Sensitivity to Precipitation and Its Oceanic and Terrestrial Component in Selected Biomes and Ecoregions of the World" Remote Sensing 15, no. 19: 4706. https://doi.org/10.3390/rs15194706
APA StyleStojanovic, M., Sorí, R., Guerova, G., Vázquez, M., Nieto, R., & Gimeno, L. (2023). Vegetation Greenness Sensitivity to Precipitation and Its Oceanic and Terrestrial Component in Selected Biomes and Ecoregions of the World. Remote Sensing, 15(19), 4706. https://doi.org/10.3390/rs15194706