
Environmental Performance of Regional Protected Area Network: Typological Diversity and Fragmentation of Forests


 ,
, 
Abstract
1. Introduction
- Assessment of the composition and spatial distribution of forest cover in PAs;
- Assessment of the structure (fragmentation) of forest cover in PAs;
- Identification of the main factors that determine the composition and structure of PAs forests, taking into account natural conditions and anthropogenic pressure.
2. Materials and Methods
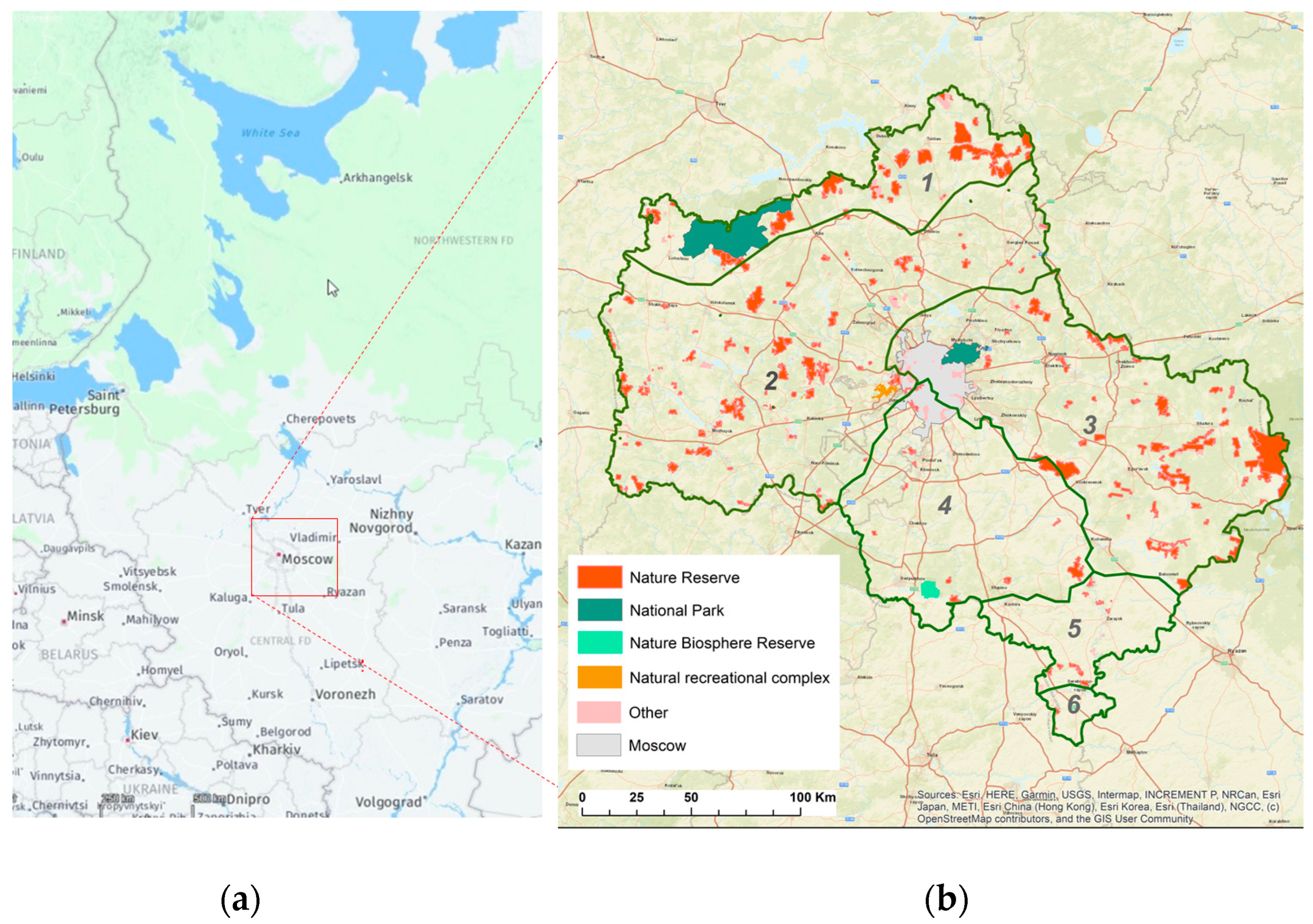
2.1. Study Area
2.2. History and Structure of PAs Network in the MR
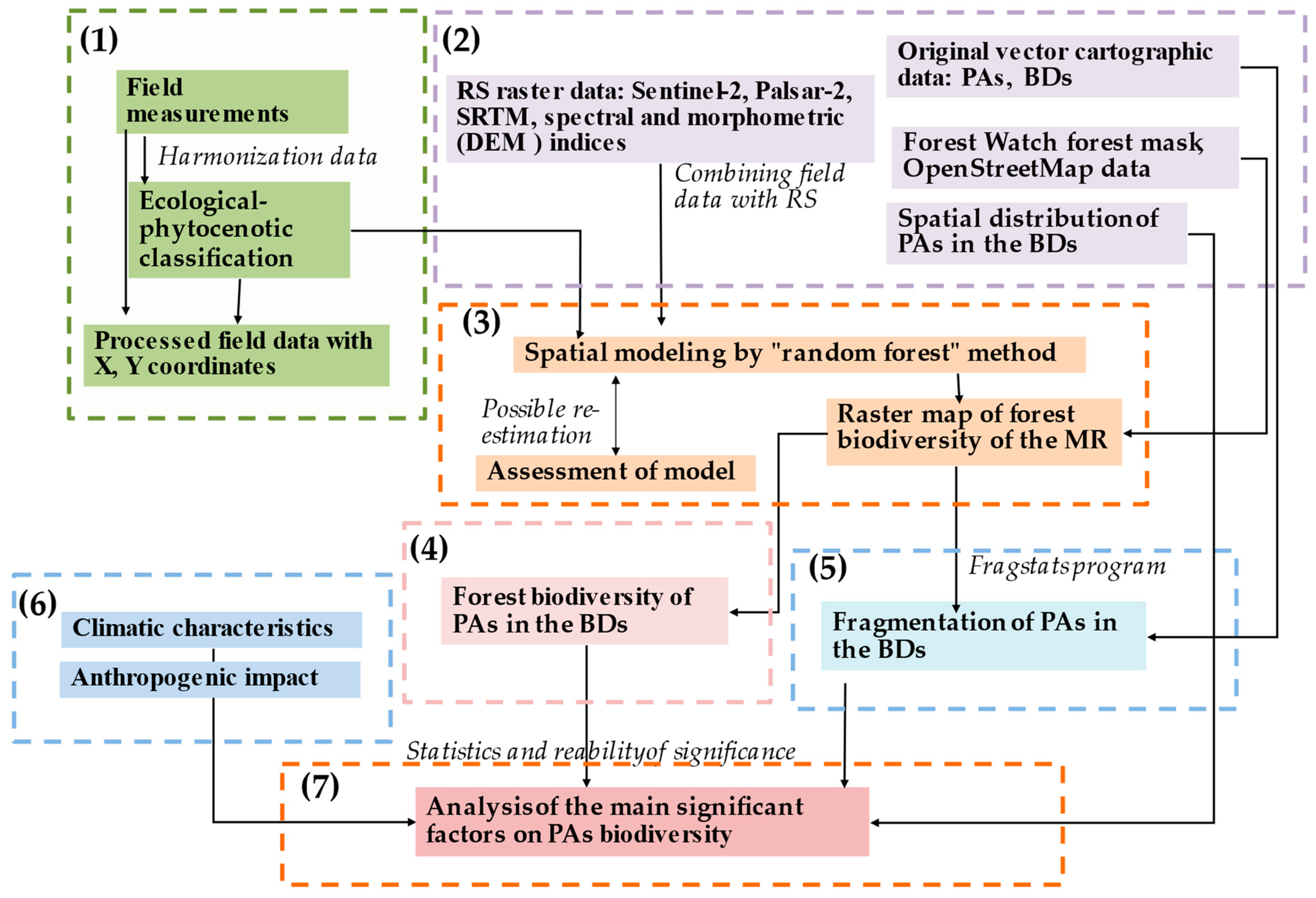
2.3. Study Design
- The territory of the historical part of Moscow within the boundaries until 2012 is excluded from the total area of the MR. Accordingly, such categories of PAs as the Dendrological park and Botanical Garden and the Natural Historical Park were not included in the analysis.
- The protection of forest areas that characterize terrestrial vegetation covers most of the entire list of PAs. In this regard, Strict protected water object was excluded from consideration.
- PAs have various shapes and sizes and are distinguished by a variety of management systems and origins. To analyze the typical combination of types of plant communities, PAs were filtered by the minimum area. The previous results of our research allowed us to reveal the average size of a forest patch in the near zone of Moscow (14 ± 4 ha) and on the periphery (37 ± 26 ha) (unpublished data). Therefore, PAs with an area of less than 36 ha, equaling 10 × 10 pixels with a resolution of 60 m (Sentinel-2 resolution) were excluded from the analysis. Most of the small PAs are mainly represented by the Natural Monument category or local PAs. As a result, the analyzed PAs account for 5.8% of the study area (265.2 thousand ha).
- Area of PAs;
- Climate variables;
- Establishment date of PAs (existence time of PAs);
- Percentage of forests;
- Fragmentation;
- Category of PAs;
- Anthropogenic pressure (distance from Moscow and other large settlements).
2.4. Field Data and Classification
2.5. RS Data, Cartographic Modeling, and Fragmentation Evaluation
2.6. Assessment of the Environmental Variables of PAs
2.7. Analysis of Correlation between the Main Factors and Forest Biodiversity of PAs
3. Results
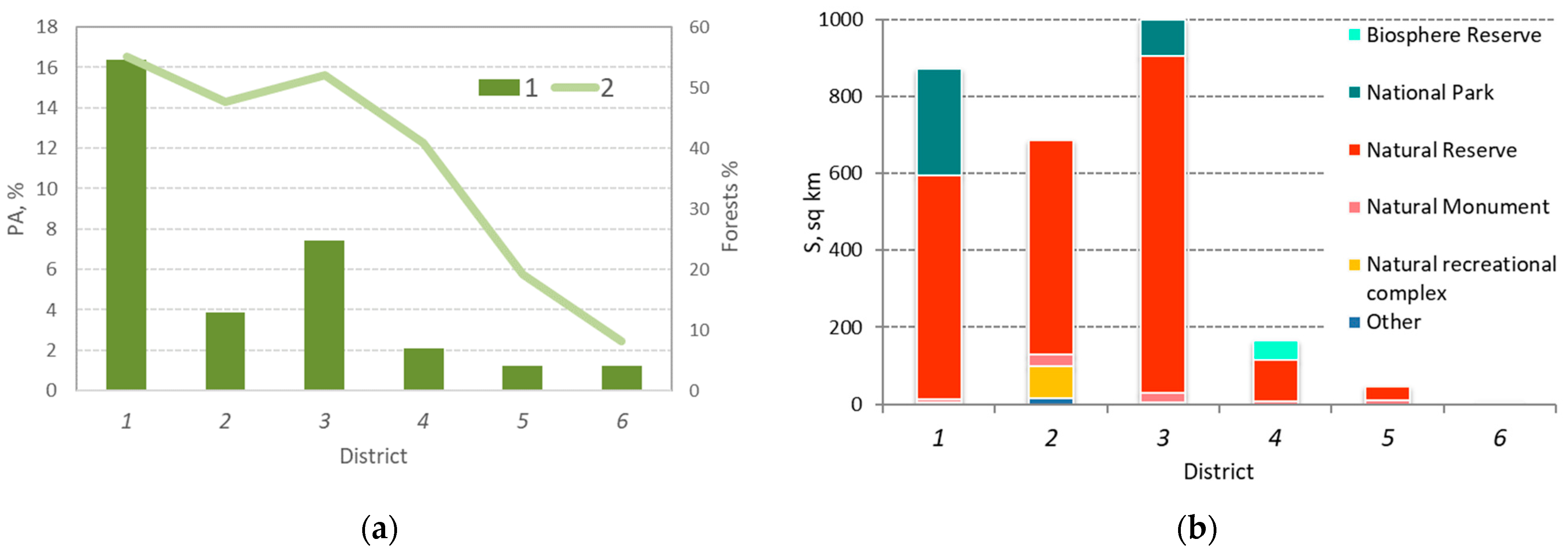
3.1. Analysis of the Spatial Distribution of PAs
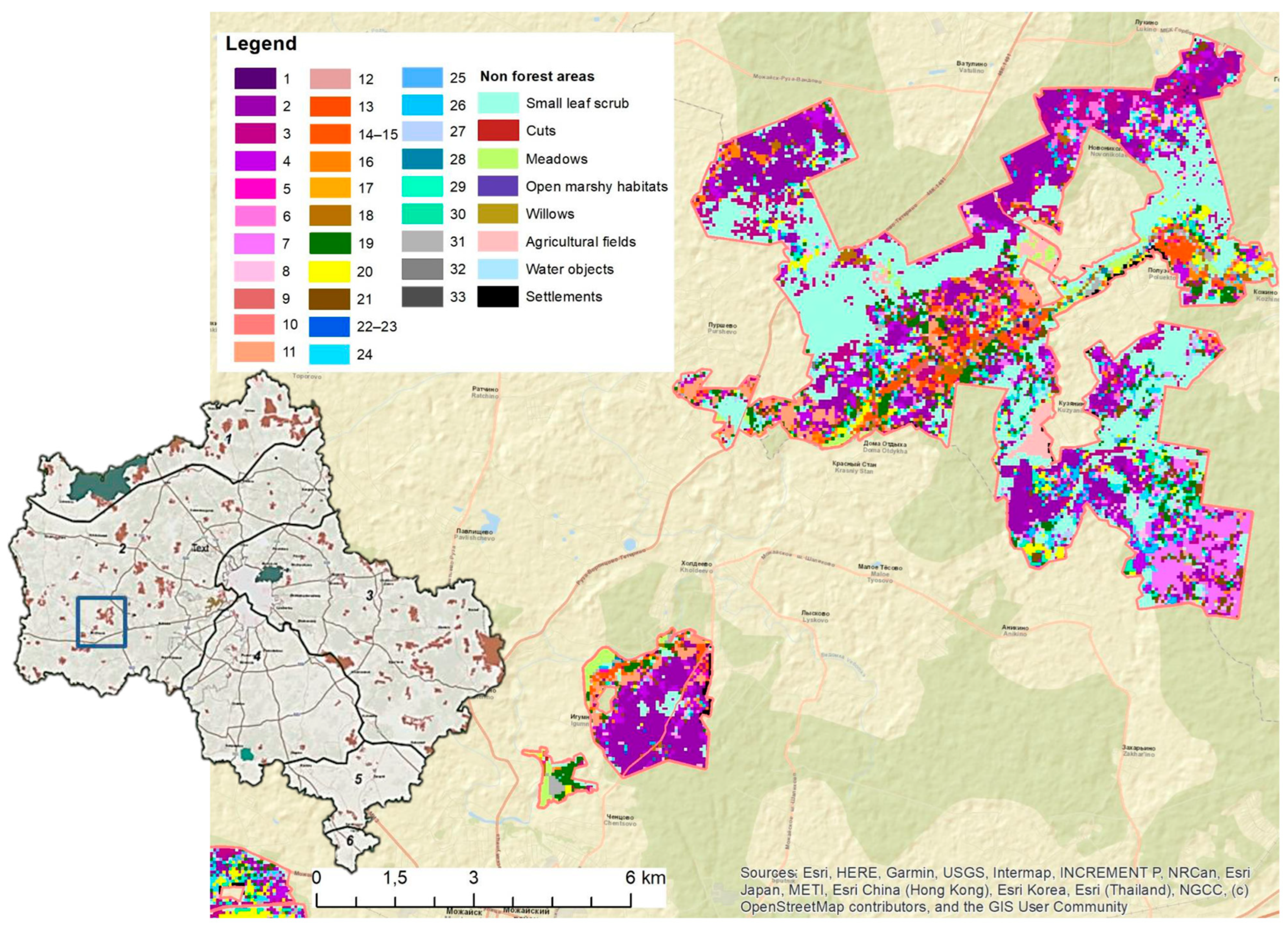
3.2. Results of Ecological-Phytocenotic Classification
3.3. Cartographic Mapping of Forest Typological Diversity
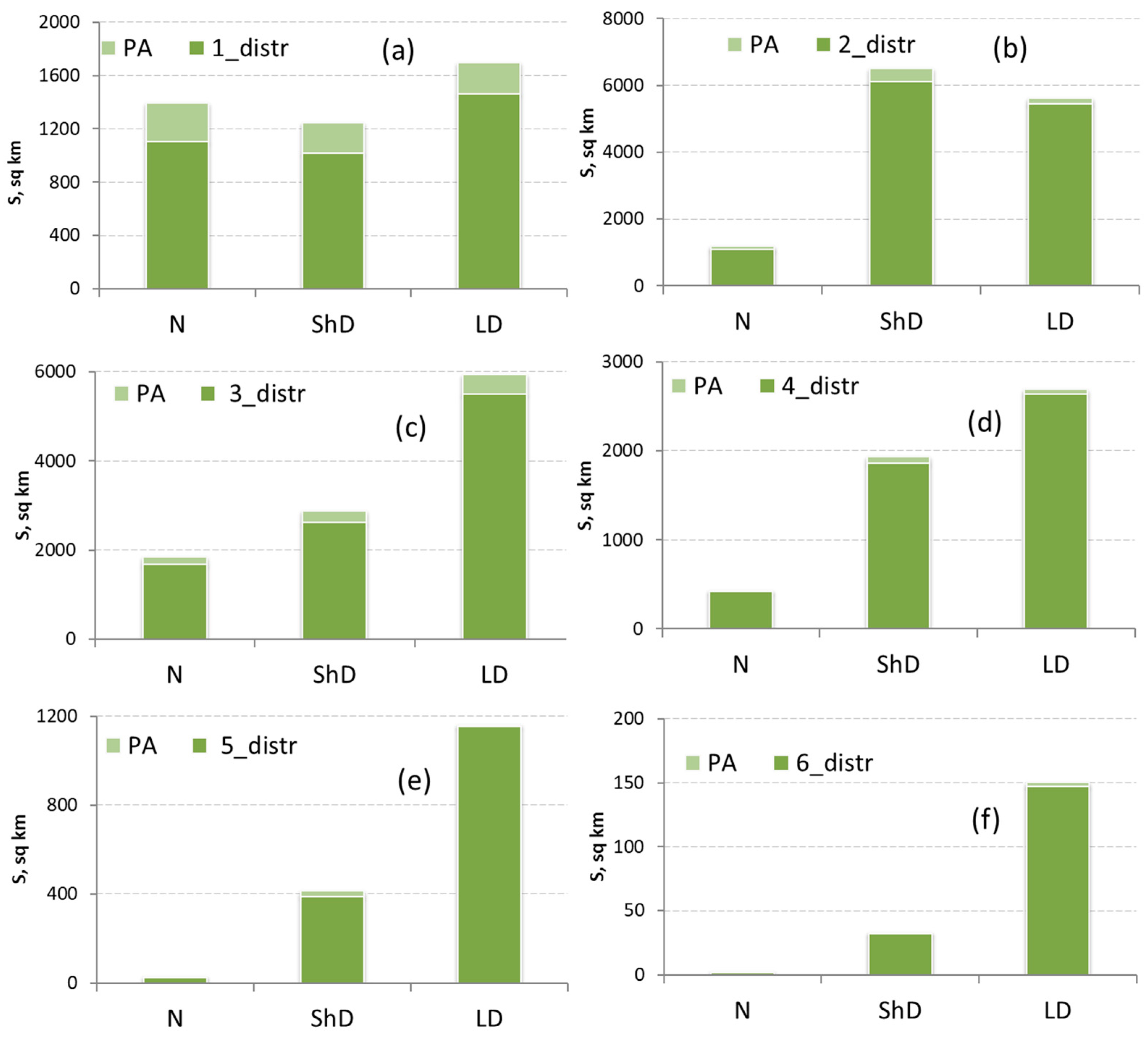
3.4. Analysis of the Spatial Distribution of PAs by Forest Composition
3.5. Analysis of the Spatial Distribution of PAs Fragmentation Metrics
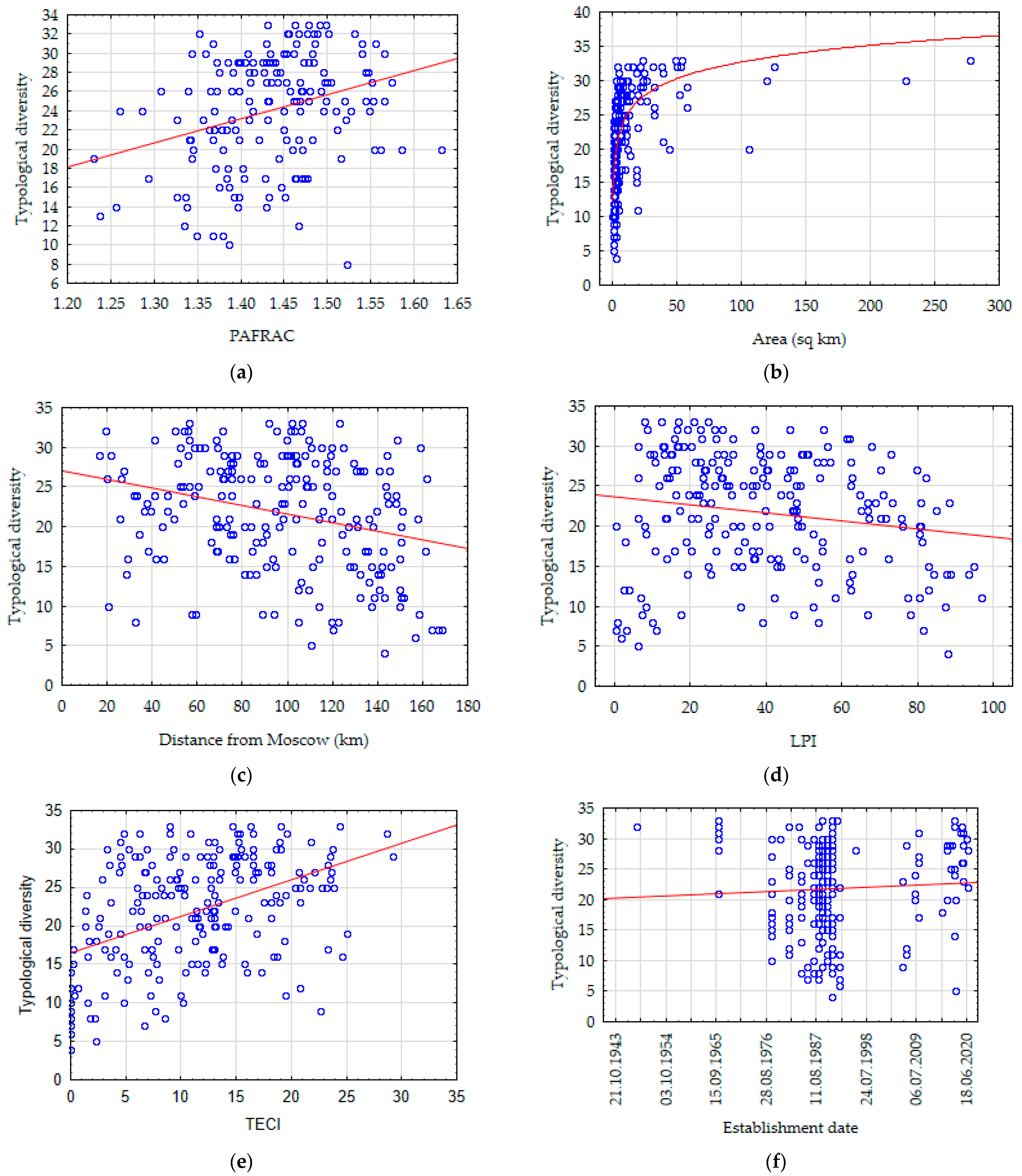
3.6. Assessment of the Main Environmental Variables and Parameters of PAs
3.7. Analysis of Correlation between the Main Factors and the Forest Biodiversity of PAs
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Name | Description | Characteristics, Formula |
|---|---|---|---|
| 1 | B02—Blue | Sensitivity to plant aging, carotenoids, browning and soil background; atmospheric correction (aerosol scattering) | 458–522 нм |
| 2 | B06—Red Edge | Red edge position, atmospheric correction (aerosol scattering) | 733–747 нм |
| 3 | NDWI2 | Normalized differential water index. Emphasizes the humidity of habitats [45,78] | |
| 4 | BNDWI | Normalized difference index of blue and infrared. Relationship with leaf area index and dry biomass volume [45,79] | |
| 5 | GLI | Green leaf index. Chlorophyll and leaf surface characteristics based on visible bands [45,80] | |
| 6 | DEM SRTM (elevation) | Position relative to watersheds and stream valleys [81] | meters |
| 7 | HH Palsar | Textural heterogeneity of the crown surface, tree layer height, biomass [46] | conventional units |
| Metrics | Formula | Units |
|---|---|---|
| PD Patch density | N—number of patches in PAs A—area of PAs | Number per 100 ha |
| LPI Largest patch index | aij—area (m2) of patch ij A—area of PA | Percentage |
| PAFRAC Perimeter-area fractal dimension | PAFRAC = aij—area (m2) of patch ij pij—perimeter (m) of patch ij N—number of patches in PA | None |
| TECI Total edge contrast index | eik—total length of boundaries between formations i and k in the SPNA, including the outer boundaries of the PAs belonging to the formation i. E*—total length of boundaries in PAs, including outer boundaries. dik—contrast of boundaries between formations i and k. | Percentage |
| IJI interspersion/juxtaposition index | eik—total length of boundaries between formations i and k in PAs. E—total length of boundaries in PAs, excluding outer boundaries. m—number of formations in PAs | Percentage |
| SPLIT Splitting index | aij—area (m2) of patch ij A—protected area | none |
| Formation | Association Group | Legend Index | Type of Dynamics | ||
|---|---|---|---|---|---|
| 1 | Spruce forests (Sp) (Picea abies) | 1 | Spruce forests with birch, aspen and pine dwarf shrubs–small herb–green moss (Vaccinium myrtillus, V. vitis-idaea, Oxalis acetosella, Calamagrostis arundinacea, Luzula pilosa, Pleurozium schreberi, Hylocomium splendens) | Sp_DshShG | N |
| 2 | Spruce forests with birch and aspen small herb (Oxalis acetosella) | Sp_Sh | ShD | ||
| 3 | Spruce forests with birch, aspen and pine small herb–broad herbs (Oxalis acetosella, Carex pilosa, Galeobdolon luteum) | Sp_ShBh | ShD | ||
| 4 | Spruce forests with birch, aspen, pine, oak and linden broad herbs (Galeobdolon luteum, Aegopodium podagraria, Carex pilosa, Anemonoides nemorosa, Oxalis acetosella) | Sp_Bh | ShD | ||
| 2 | Spruce—aspen/birch forests (Sp-As/B) (Picea abies—Populus tremula—Betula pendula, B. pubescens) | 5 | Spruce—aspen/birch dwarf shrubs–small herbs–green mosses (Vaccinium myrtillus, Oxalis acetosella, Calamagrostis arundinacea, Orthilia secunda, Pleurozium schreberi, Hylocomium splendens) | Sp-As/B_DshShG | ShD |
| 6 | Spruce—aspen/birch small herbs (Oxalis acetosella, Dryopteris carthusiana, Rubus saxatilis, Plagiomnium affine) | Sp-As/B_Sh | ShD | ||
| 7 | Spruce—aspen and spruce—small herbs–broad herbs (Corylus avellana, Athyrium filix-femina, Dryopteris carthusiana, D. filix-mas, Oxalis acetosella, Rubus saxatilis, Convallaria majalis, Galeobdolon luteum, Atrichum undulatum, Hylocomoim splendens) | Sp-As/B_ShBh | ShD | ||
| 8 | Spruce—birch with oak and linden broad herbs (Aegopodium podagraria, Carex pilosa, Pulmonaria obscura, Dryopteris filix-mas, Galeobdolon luteum, Eurhynchium angustirete) | Sp-As/B_Bh | ShD | ||
| 3 | Pine—spruce forests (P-Sp) (Pinus sylvestris—Picea abies) | 9 | Pine—spruce with birch dwarf shrubs–small herbs–green mosses (Vaccinium myrtillus, Oxalis acetosella, Dryopteris carthusisna, Pleurozium schreberi, Hylocomium splendens) | Sp-As/B_DshShG | ShD |
| 10 | Pine—spruce small herbs (Oxalis acetosella) | Sp-As/B_Sh | ShD | ||
| 11 | Pine—spruce small herbs–broad herbs (Corylus avellana, Oxalis acetosella, Galeobdolon luteum, Dryopteris carthusiana) | Sp-As/B_ShBh | ShD | ||
| 12 | Pine—spruce with birch broad herbs (Athyrium filix-femina, Galeobdolon luteum, Carex pilosa, Oxalis acetosella) | Sp-As/B_Bh | ShD | ||
| 4 | Pine forests (P) (Pinus sylvestris) | 13 | Pine with spruce and birch dwarf shrubs–small herbs–green mosses (Vaccinium yrtillus, V. vit1s-idaea, Pteridium aquilinum, Calamagrostis arundinacea, Convallaria majalis, Luzula pilosa, Maianthemum bifolium, Hylocomium splendens, Pleurozium schreberi, Dicranum scoparium) | P_DshShG | N |
| 14 | Pine with spruce and birch small herbs (Corylus avellana, Oxalis acetosella, Vaccinium myrtillus, Calamagrostis arundinacea) | P_Sh | ShD | ||
| 15 | Pine with spruce and birch partly with oak and linden small herbs–broad herbs (Oxalis acetosella, Gymnocarpium dryopteris, Galeobdolon luteum, Dryopteris carthusiana, Athyrium filix-femina, Aegopodium podagraria) | P_ShBh | ShD | ||
| 16 | Pine with spruce, birch, oak, and linden broad herbs (Carex pilosa, Convallaria majalis, Galeobdolon luteum, Ranunculus cassubicus, Oxalis acetosella) | P_Bh | ShD | ||
| 17 | Pine with spruce and birch meadow herbs (Calamagrostis arundinacea, Poa angustifolia, Convallaria majalis, Fragaria vesca) | P_Mh | ShD | ||
| 18 | Pine with birch (Betula pubescens) dwarf shrubs–herbal-sphagnum (Chamaedaphne calyculata, Ledum palustre, Vaccinium myrtillus, V. uliginosum, Oxycoccus palustris, Eriophorum vaginatum, Sphagnum angustifolium, S. magellanicum) | P_DshHSh | N | ||
| 5 | Oak forests (O) (Quercus robur) | 19 | Oak with linden, spruce, and birch broad herbs (Aegopodium podagraria, Carex pilosa, Galeobdolon luteum) | O_Bh | N |
| 6 | Linden forests (L) (Tilia cordata) | 20 | Linden broad herbs (Carex pilosa, Aegopodium podagraria, Mercurialis perennis, Pulmonaria obscura) | L_Bh | N |
| 7 | Broad leaf—spruce forests (Bl-Sp) (Quercus robur—Tilia cordata—Picea abies) | 21 | Oak—linden—spruce broad herbs (Carex pilosa, Galeobdolon luteum, Aegopodium podagraria, Asarum europaeum, Pulmonaria obscura, Ranunculus cassubicus, Stellaria nemorum) | Bl-Sp_Bh | N |
| 8 | Birch forests (B) (Betula pendula, B. pubescens) | 22 | Birch with spruce and aspen small herbs (Oxalis acetosella, Pyrola rotundifolia, Luzula pilosa) | B_Sh | LD |
| 23 | Birch with spruce and aspen small herbs–broad herbs (Oxalis acetosella, Athyrium filix-femina, Calamagrostis arundinacea, Rubus saxatilis, Galeobdolon luteum, Aegopodium podagraria, Pyrola rotundifolia, Cirriphyllum piliferum) | B_ShBh | LD | ||
| 24 | Birch with spruce and grey alder broad herb (Aegopodium podagraria, Carex pilosa, Galeobdolon luteum, Pulmonaria obscura, Stellaria nemoru) | B_Bh | LD | ||
| 25 | Birch moist herb—broad herb (Salix caprea, Filipendula ulmaria, Athyrium filix-femina, Urtica dioica, Calamagrostis arundinacea, Impatiens noli-tangere, Pulmonaria obscura, Geum rivale, Atrichum undulatum) | B_MihBh | LD | ||
| 26 | Birch with spruce and aspen grass-marsh (Filipendula ulmaria, Calamagrostis canescens, Phragmites australis, Carex acuta, C. vesicaria, Scirpus sylvaticus, Aulacomnium palustre, Climacium dendroides) | B_Gm | N | ||
| 27 | Birch with spruce, aspen, and willow meadow herbs (Bromopsis inermis, Calamagrostis arundinacea, C. epigeios, Fragaria vesca, Lysimachia nummularia, Veronica chamaedrys, Deschampsia cespitosa) | B_Mh | LD | ||
| 28 | Birch with spruce dwarf shrubs–herbal-sphagnum (Chamaedaphne calyculata, Vaccinium uliginosum, Eriophorum vaginatum, Carex lasiocarpa, Sphagnum spp., Polytrichum commune) | B_DshHSh | ShD | ||
| 9 | Aspen forests (As) (Populus tremula) | 29 | Aspen with birch, spruce, oak, and linden broad herbs (Corylus avellana, Aegopodium podagraria, Galeobdolon luteum, Carex pilosa, Mercurialis perennis) | As_Bh | ShD |
| 30 | Aspen with birch, spruce, oak, and bird cherry moist herbs—broad herbs (Padus avium, Athyrium filix-femina, Crepis paludosa, Filipendula ulmaria, Urtica dioica, Pulmonaria obscura, Equisetum pratense, Stellaria nemorum, Impatiens noli-tangere, Atrichum undulatum, Plagiomnium cuspidatum) | As_MihBh | ShD | ||
| 10 | Grey alder forests (GAl) (Alnus incana) | 31 | Grey alder moist herbs—broad herbs (Urtica dioica, Campanula latifolia, Filipendula ulmaria, Rubus idaeus, Aegopodium podagraria, Chrysosplenium alternifolium, Myosoton aquaticum, Stellaria nemorum, Plagiomnium undulatum) | GAl_MihBh | N |
| 11 | Black alder forests (BAl) (Alnus glutinosa) | 32 | Black alder moist herbs—broad herbs (Impatiens noli-tangere, Urtica dioica, Milium effusum, Paris quadrifolia, Ranunculus cassubicus) | BAl_MihBh | N |
| 33 | Black alder grass-marsh (Urtica dioica, Filipendula ulmaria, Phragmites australis, Carex appropinquata, C. vesicaria, Calla palustris, Humulus lupulus) | BAl_Gm | N |
| Ass. Gr. Number | Number of Points | Average Polygon Area, ha | Total Area of Polygons, ha | Number of Pixels for the Training Sample | Convergence Ass. Gr. by Test Sample | Percentage of Forest Area, % |
|---|---|---|---|---|---|---|
| 1 | 32 | 0.57 | 7.37 | 84 | 60 | 5.3 |
| 2 | 35 | 5.28 | 58.06 | 197 | 55 | 5.5 |
| 3 | 124 | 0.97 | 10.63 | 155 | 23 | 6.8 |
| 4 | 132 | 0.60 | 6.60 | 166 | 18 | 4.8 |
| 5 | 22 | 0.73 | 8.05 | 45 | 30 | 1.0 |
| 6 | 13 | 0.41 | 4.90 | 30 | 67 | 0.7 |
| 7 | 62 | 0.35 | 3.14 | 71 | 13 | 1.4 |
| 8 | 91 | 0.52 | 5.23 | 104 | 21 | 3.4 |
| 9 | 31 | 1.30 | 14.28 | 69 | 41 | 2.0 |
| 10 | 16 | 1.88 | 18.76 | 65 | 63 | 1.0 |
| 11 | 41 | 0.78 | 8.58 | 132 | 56 | 1.5 |
| 12 | 38 | 1.65 | 16.52 | 88 | 41 | 0.7 |
| 13 | 46 | 2.44 | 29.24 | 134 | 69 | 2.8 |
| 14 + 15 | 56 | 2.45 | 23.75 | 182 | 54 | 3.8 |
| 16 | 63 | 2.12 | 23.31 | 148 | 46 | 1.9 |
| 17 | 14 | 0.95 | 7.62 | 85 | 81 | 2.6 |
| 18 | 46 | 11.21 | 123.28 | 422 | 89 | 4.6 |
| 19 | 52 | 6.42 | 64.19 | 233 | 71 | 3.7 |
| 20 | 109 | 4.43 | 48.74 | 470 | 77 | 12.7 |
| 21 | 35 | 1.76 | 17.56 | 82 | 53 | 1.0 |
| 22 + 23 | 26 | 0.87 | 11.24 | 58 | 80 | 0.8 |
| 24 | 131 | 2.01 | 24.07 | 212 | 55 | 13.7 |
| 25 | 13 | 1.33 | 13.27 | 59 | 46 | 0.7 |
| 26 | 18 | 0.90 | 9.85 | 73 | 46 | 1.7 |
| 27 | 22 | 0.38 | 3.81 | 50 | 44 | 1.0 |
| 28 | 10 | 1.86 | 18.63 | 66 | 54 | 1.0 |
| 29 | 65 | 0.89 | 8.90 | 109 | 46 | 3.5 |
| 30 | 7 | 0.83 | 5.82 | 19 | 100 | 0.1 |
| 31 | 28 | 0.37 | 3.69 | 65 | 36 | 1.6 |
| 32 | 22 | 1.21 | 12.06 | 55 | 71 | 2.0 |
| 33 | 31 | 0.98 | 10.81 | 68 | 77 | 6.6 |
| 34 | 26 | 2.59 | 25.86 | 102 | 72 | * |
| 35 | 3 | 0.78 | 1.55 | 7 | 50 | * |
| 36 | 8 | 2.26 | 13.56 | 47 | 93 | * |
| 37 | 10 | 2.27 | 4.54 | 29 | 50 | * |
| 38 | 6 | 2.31 | 3.98 | 30 | 67 | * |
References
- Joppa, L.N.; Pfaff, A. Global Protected Area Impacts. Proc. R. Soc. B Biol. Sci. 2011, 278, 1633–1638. [Google Scholar] [CrossRef] [PubMed]
- Joseph, G. Fundamentals of Remote Sensing; Universities Press: Delhi, India, 2005. [Google Scholar]
- MCPFE—Environment—European Commission. Available online: https://ec.europa.eu/environment/forests/mcpfe.htm (accessed on 19 October 2022).
- Spangenberg, J. Reconciling Sustainability and Growth: Criteria, Indicators, Policies. Sustain. Dev. 2004, 12, 74–86. [Google Scholar] [CrossRef]
- Hansen, A.J.; DeFries, R. Ecological Mechanisms Linking Protected Areas to Surrounding Lands. Ecol. Appl. 2007, 17, 974–988. [Google Scholar] [CrossRef]
- D’Andrea, E.; Ferretti, F.; Zapponi, L.; Badano, D.; Balestrieri, R.; Basile, M.; Becagli, C.; Bertini, G.; Bertollotto, P.; Birtele, D.; et al. Indicators of Sustainable Forest Management: Application and Assessment. Ann. Silvic. Res. 2016, 40. [Google Scholar] [CrossRef]
- Defries, R.; Karanth, K.; Pareeth, S. Interactions between Protected Areas and Their Surroundings in Human-Dominated Tropical Landscape. Biol. Conserv. 2010, 143, 2870–2880. [Google Scholar] [CrossRef]
- Bellón, B.; Blanco, J.; De Vos, A.; de O. Roque, F.; Pays, O.; Renaud, P.-C. Integrated Landscape Change Analysis of Protected Areas and Their Surrounding Landscapes: Application in the Brazilian Cerrado. Remote Sens. 2020, 12, 1413. [Google Scholar] [CrossRef]
- Nagendra, H.; Lucas, R.; Honrado, J.P.; Jongman, R.H.G.; Tarantino, C.; Adamo, M.; Mairota, P. Remote Sensing for Conservation Monitoring: Assessing Protected Areas, Habitat Extent, Habitat Condition, Species Diversity, and Threats. Ecol. Indic. 2013, 33, 45–59. [Google Scholar] [CrossRef]
- Corbane, C.; Lang, S.; Pipkins, K.; Alleaume, S.; Deshayes, M.; García Millán, V.E.; Strasser, T.; Vanden Borre, J.; Toon, S.; Michael, F. Remote Sensing for Mapping Natural Habitats and Their Conservation Status—New Opportunities and Challenges. Int. J. Appl. Earth Obs. Geoinf. 2015, 37, 7–16. [Google Scholar] [CrossRef]
- Forstmaier, A.; Shekhar, A.; Chen, J. Mapping of Eucalyptus in Natura 2000 Areas Using Sentinel 2 Imagery and Artificial Neural Networks. Remote Sens. 2020, 12, 2176. [Google Scholar] [CrossRef]
- Wang, R.; Gamon, J.A. Remote Sensing of Terrestrial Plant Biodiversity. Remote Sens. Environ. 2019, 231, 111218. [Google Scholar] [CrossRef]
- Hansen, M.; Potapov, P.; Margono, B.; Stehman, S.; Turubanova, S.; Tyukavina, A. Response to Comment on “High-Resolution Global Maps of 21st-Century Forest Cover Change”. Science 2014, 344, 981. [Google Scholar] [CrossRef] [PubMed]
- Potapov, P.; Hansen, M.C.; Laestadius, L.; Turubanova, S.; Yaroshenko, A.; Thies, C.; Smith, W.; Zhuravleva, I.; Komarova, A.; Minnemeyer, S.; et al. The Last Frontiers of Wilderness: Tracking Loss of Intact Forest Landscapes from 2000 to 2013. Sci. Adv. 2017, 3, e1600821. [Google Scholar] [CrossRef]
- Scharsich, V.; Mtata, K.; Hauhs, M.; Lange, H.; Bogner, C. Analysing Land Cover and Land Use Change in the Matobo National Park and Surroundings in Zimbabwe. Remote Sens. Environ. 2017, 194, 278–286. [Google Scholar] [CrossRef]
- Leidner, A.; Brink, A.; Szantoi, Z. Leveraging Remote Sensing for Conservation Decision Making. Eos Trans. Am. Geophys. Union 2013, 94, 508. [Google Scholar] [CrossRef]
- Szantoi, Z.; Brink, A.; Buchanan, G.; Bastin, L.; Lupi, A.; Simonetti, D.; Mayaux, P.; Peedell, S.; Davy, J. A Simple Remote Sensing Based Information System for Monitoring Sites of Conservation Importance. Remote Sens. Ecol. Conserv. 2016, 2, 16–24. [Google Scholar] [CrossRef]
- Mikula, K.; Kollar, M.; Ožvat, A.A.; Ambroz, M.; Čahojová, L.; Jarolimek, I.; Šibík, J.; Šibíková, M. Natural Numerical Networks for Natura 2000 Habitats Classification by Satellite Images. Appl. Math. Model. 2023, 116, 209–235. [Google Scholar] [CrossRef]
- Alcaraz-Segura, D.; Cabello, J.; Paruelo, J.; Delibes, M. Use of Descriptors of Ecosystem Functioning for Monitoring a National Park Network: A Remote Sensing Approach. Environ. Manag. 2009, 43, 38–48. [Google Scholar] [CrossRef]
- Zeng, J.; Chen, T.; Yao, X.; Chen, W. Do Protected Areas Improve Ecosystem Services? A Case Study of Hoh Xil Nature Reserve in Qinghai-Tibetan Plateau. Remote Sens. 2020, 12, 471. [Google Scholar] [CrossRef]
- Liu, M.; Dries, L.; Heijman, W.; Huang, J.; Zhu, X.; Hu, Y.; Chen, H. The Impact of Ecological Construction Programs on Grassland Conservation in Inner Mongolia, China—Liu—2018—and Degradation & Development—Wiley Online Library. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1002/ldr.2692 (accessed on 17 October 2022).
- Imbrenda, V.; Lanfredi, M.; Coluzzi, R.; Simoniello, T. A Smart Procedure for Assessing the Health Status of Terrestrial Habitats in Protected Areas: The Case of the Natura 2000 Ecological Network in Basilicata (Southern Italy). Remote Sens. 2022, 14, 2699. [Google Scholar] [CrossRef]
- Rose, R.A.; Byler, D.; Eastman, J.R.; Fleishman, E.; Geller, G.; Goetz, S.; Guild, L.; Hamilton, H.; Hansen, M.; Headley, R.; et al. Ten Ways Remote Sensing Can Contribute to Conservation. Conserv. Biol. 2015, 29, 350–359. [Google Scholar] [CrossRef]
- Wegmann, M.; Santini, L.; Leutner, B.; Safi, K.; Rocchini, D.; Bevanda, M.; Latifi, H.; Dech, S.; Rondinini, C. Role of African Protected Areas in Maintaining Connectivity for Large Mammals. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 2014, 369, 20130193. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Xun, B.; Shi, P.; Shao, H.; Liu, Y. Ecological Restoration Planning Based on Connectivity in an Urban Area. Ecol. Eng. 2012, 46, 24–33. [Google Scholar] [CrossRef]
- Girardet, H. Creating Regenerative Cities; Routledge: London, UK, 2014. [Google Scholar]
- Noss, R.F. Assessing and Monitoring Forest Biodiversity: A Suggested Framework and Indicators. For. Ecol. Manag. 1999, 115, 135–146. [Google Scholar] [CrossRef]
- Baines, O.; Wilkes, P.; Disney, M. Quantifying Urban Forest Structure with Open-Access Remote Sensing Data Sets. Urban For. Urban Green. 2020, 50, 126653. [Google Scholar] [CrossRef]
- Wang, K.; Wang, T.; Liu, X. A Review: Individual Tree Species Classification Using Integrated Airborne LiDAR and Optical Imagery with a Focus on the Urban Environment. Forests 2019, 10, 1. [Google Scholar] [CrossRef]
- Torbick, N.; Ledoux, L.; Salas, W.; Zhao, M. Regional Mapping of Plantation Extent Using Multisensor Imagery. Remote Sens. 2016, 8, 236. [Google Scholar] [CrossRef]
- Ghosh, S.M.; Behera, M.D. Aboveground Biomass Estimation Using Multi-Sensor Data Synergy and Machine Learning Algorithms in a Dense Tropical Forest. Appl. Geogr. 2018, 96, 29–40. [Google Scholar] [CrossRef]
- Kotlov, I.P.; Chernenkova, T.V. Modeling of Forest Communities Spatial Structure at the Regional Level through Remote Sensing and Field Sampling: Constraints and Solutions. Forests 2020, 11, 1088. [Google Scholar] [CrossRef]
- Bobrovskii, M.V. Forest Soils of European Russia: Biotic and Anthropogenic Factors of Formation; Tovarishchestvo Nauchnykh Izdanii KMK: Moscow, Russia, 2010. [Google Scholar]
- Tsvetkov, M.A. Changes in the Forest Cover of European Russia from the End of the 17th Century to 1914; SSSR AS: Moscow, Russia, 1957. [Google Scholar]
- Potapov, P.V.; Turubanova, S.A.; Tyukavina, A.; Krylov, A.M.; McCarty, J.L.; Radeloff, V.C. Eastern Europe’s Forest Cover Dynamics from 1985 to 2012 Quantified from the Full Landsat Archive. Remote Sens. Environ. 2015, 28–43. [Google Scholar] [CrossRef]
- Information Issue “on the State of Natural Resources and the Environment of the Moscow Region in 2021”. Available online: https://mep.mosreg.ru/dokumenty/informaciya-i-statistika/12-08-2022-11-35-53-informatsionnyy-vypusk-o-sostoyanii-prirodnykh-res (accessed on 19 October 2022).
- Gribova, S.A.; Isachenko, T.I.; Lavrenko, E.M. Vegetation of European Part of the USSR; Nauka: Leningrad, USSR, 1980. [Google Scholar]
- Chernenkova, T.V.; Kotlov, I.P.; Belyaeva, N.G.; Suslova, E.G.; Morozova, O.V.; Pesterova, O.; Arkhipova, M.V. Role of Silviculture in the Formation of Norway Spruce Forests along the Southern Edge of Their Range in the Central Russian Plain. Forests 2020, 11, 778. [Google Scholar] [CrossRef]
- Petrov, V.V. New Scheme of Moscow Region Geobotanical Zoning. Vestn. Mosk. Univ. 1968, 44–49. [Google Scholar]
- Sobolev, N.; Shvarts, E.; Kreindlin, M.; Mokievsky, V.; Zubakin, V. Russia’s Protected Areas: A Survey and Identification of Development Problems. Biodivers. Conserv. 1995, 4, 964–983. [Google Scholar] [CrossRef]
- World Conservation Monitoring Centre; IUCN Commission on National Parks; Protected Areas. 1990 United Nations List of National Parks and Protected Areas; Iucn: Gland, Switzerland; Cambridge, UK, 1990. [Google Scholar]
- Chernen’kova, T.V.; Kotlov, I.P.; Belyaeva, N.G.; Suslova, E.G.; Morozova, O.V. Assessment and mapping of the cenotic diversity of the Moscow region’s forest. Лecoвeдeниe 2022, 6. [Google Scholar]
- Chernenkova, T.; Kotlov, I.; Belyaeva, N.; Suslova, E. Spatiotemporal Modeling of Coniferous Forests Dynamics along the Southern Edge of Their Range in the Central Russian Plain. Remote Sens. 2021, 13, 1886. [Google Scholar] [CrossRef]
- Chernenkova, T.V.; Morozova, O.V. Classification and Mapping of Coenotic Diversity of Forests. Contemp. Probl. Ecol. 2017, 10, 738–747. [Google Scholar] [CrossRef]
- Abdullah, H.; Skidmore, A.K.; Darvishzadeh, R.; Heurich, M. Sentinel-2 Accurately Maps Green-attack Stage of European Spruce Bark Beetle (Ips Typographus, L.) Compared with Landsat-8. Remote Sens. Ecol. Conserv. 2019, 5, 87–106. [Google Scholar] [CrossRef]
- Shimada, M.; Itoh, T.; Motooka, T.; Watanabe, M.; Shiraishi, T.; Thapa, R.; Lucas, R. New Global Forest/Non-Forest Maps from ALOS PALSAR Data (2007–2010). Remote Sens. Environ. 2014, 155, 13–31. [Google Scholar] [CrossRef]
- Graham, M.H. Confronting Multicollinearity in Ecological Multiple Regression. Ecology 2003, 84, 2809–2815. [Google Scholar] [CrossRef]
- Grabska, E.; Frantz, D.; Ostapowicz, K. Evaluation of Machine Learning Algorithms for Forest Stand Species Mapping Using Sentinel-2 Imagery and Environmental Data in the Polish Carpathians. Remote Sens. Environ. 2020, 251. [Google Scholar] [CrossRef]
- Gislason, P.O.; Benediktsson, J.A.; Sveinsson, J.R. Random Forest Classification of Multisource Remote Sensing and Geographic Data. In Proceedings of the 2004 IEEE International Geoscience and Remote Sensing Symposium, Anchorage, AK, USA, 20–24 September 2004; IEEE: Piscataway, NJ, USA, 2004; Volume 2, pp. 1049–1052. [Google Scholar]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Inglada, J.; Christophe, E. The Orfeo Toolbox Remote Sensing Image Processing Software. In Proceedings of the 2009 IEEE International Geoscience and Remote Sensing Symposium, Cape Town, South Africa, 12–17 July 2009; IEEE: Piscataway, NJ, USA, 2009; Volume 4, p. IV-733-IV–736. [Google Scholar]
- Cochran, W.G. Sampling Techniques, 3rd ed.; Wiley Series in Probability and Mathematical Statistics; Wiley: New York, NY, USA, 1977; ISBN 978-0-471-16240-7. [Google Scholar]
- Lyons, M.B.; Keith, D.A.; Phinn, S.R.; Mason, T.J.; Elith, J. A Comparison of Resampling Methods for Remote Sensing Classification and Accuracy Assessment. Remote Sens. Environ. 2018, 208, 145–153. [Google Scholar] [CrossRef]
- Joelsson, S.R.; Benediktsson, J.A.; Sveinsson, J.R. Random Forest Classification of Remote Sensing Data. In Signal and Image Processing for Remote Sensing; CRC Press: Boca Raton, FL, USA, 2006; pp. 344–361. ISBN 0-429-12312-4. [Google Scholar]
- Hansen, M.C.; Potapov, P.V.; Moore, R.; Hancher, M.; Turubanova, S.A.; Tyukavina, A.; Thau, D.; Stehman, S.V.; Goetz, S.J.; Loveland, T.R. High-Resolution Global Maps of 21st-Century Forest Cover Change. Science 2013, 342, 850–853. [Google Scholar] [CrossRef] [PubMed]
- Haklay, M.; Weber, P. Openstreetmap: User-Generated Street Maps. IEEE Pervasive Comput. 2008, 7, 12–18. [Google Scholar] [CrossRef]
- McGarigal, K. FRAGSTATS: Spatial Pattern Analysis Program for Quantifying Landscape Structure; US Department of Agriculture, Forest Service, Pacific Northwest Research Station: Washington, DC, USA, 1995.
- Wagner, J.M.; Shimshak, D.G. Stepwise Selection of Variables in Data Envelopment Analysis: Procedures and Managerial Perspectives. Eur. J. Oper. Res. 2007, 180, 57–67. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-Km Spatial Resolution Climate Surfaces for Global Land Areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Wang, Z.; Shrestha, R.; Yao, T.; Kalb, V. Black Marble User Guide (Version 1.2). Available online: https://ladsweb.modaps.eosdis.nasa.gov/missions-and-measurements/viirs/VIIRS_Black_Marble_UG_v1.2_April_2021.pdf (accessed on 4 March 2022).
- Tronin, A.; Gornyy, V.; Kritsuk, S.; Latypov, I.S. Nighttime Lights as a Quantitative Indicator of Anthropogenic Load on Ecosystems. Sovrem. Probl. Distantsionnogo Zondirovaniya Zemli Iz Kosm. 2014, 11, 237–244. [Google Scholar]
- Puzachenko, E. Mathematical Methods in Ecological and Geographical Studies; Academia: Moscow, Russia, 2004. [Google Scholar]
- Chernen’kova, T.V.; Suslova, E.G.; Morozova, O.V.; Belyaeva, N.G.; Kotlov, I.P. Biodiversity of Forests in the Moscow Region. Aкaдeмия Mocкca 2020, 4, 60–144. (In Russian) [Google Scholar] [CrossRef]
- Explore the World’s Protected Areas. Available online: https://www.protectedplanet.net/en (accessed on 20 October 2022).
- Leverington, F.; Costa, K.; Pavese, H.; Lisle, A.; Hockings, M. A Global Analysis of Protected Area Management Effectiveness. Environ. Manag. 2010, 46, 685–698. [Google Scholar] [CrossRef]
- Laurance, W.F.; Carolina Useche, D.; Rendeiro, J.; Kalka, M.; Bradshaw, C.J.; Sloan, S.P.; Laurance, S.G.; Campbell, M.; Abernethy, K.; Alvarez, P.; et al. Averting Biodiversity Collapse in Tropical Forest Protected Areas. Nature 2012, 489, 290–294. [Google Scholar] [CrossRef]
- Ammer, C.; Fichtner, A.; Fischer, A.; Gossner, M.M.; Meyer, P.; Seidl, R.; Thomas, F.M.; Annighöfer, P.; Kreyling, J.; Ohse, B.; et al. Key Ecological Research Questions for Central European Forests. Basic Appl. Ecol. 2018, 32, 3–25. [Google Scholar] [CrossRef]
- Koskikala, J.; Kukkonen, M.; Käyhkö, N. Mapping Natural Forest Remnants with Multi-Source and Multi-Temporal Remote Sensing Data for More Informed Management of Global Biodiversity Hotspots. Remote Sens. 2020, 12, 1429. [Google Scholar] [CrossRef]
- Du, Z.; Yang, J.; Ou, C.; Zhang, T. Smallholder Crop Area Mapped with a Semantic Segmentation Deep Learning Method. Remote Sens. 2019, 11, 888. [Google Scholar] [CrossRef]
- Alban, J.D.T.D.; Jamaludin, J.; de Wen, D.W.; Than, M.M.; Webb, E.L. Improved Estimates of Mangrove Cover and Change Reveal Catastrophic Deforestation in Myanmar. Environ. Res. Lett. 2020, 15, 034034. [Google Scholar] [CrossRef]
- Guirado, E.; Alcaraz-Segura, D.; Cabello, J.; Puertas-Ruíz, S.; Herrera, F.; Tabik, S. Tree Cover Estimation in Global Drylands from Space Using Deep Learning. Remote Sens. 2020, 12, 343. [Google Scholar] [CrossRef]
- Leopold, A. Game Management; University of Wisconsin Press: Madison, WI, USA, 1987. [Google Scholar]
- Harper, K.A.; Macdonald, S.E.; Burton, P.J.; Chen, J.; Brosofske, K.D.; Saunders, S.C.; Euskirchen, E.S.; Roberts, D.; Jaiteh, M.S.; Esseen, P.-A. Edge Influence on Forest Structure and Composition in Fragmented Landscapes. Conserv. Biol. 2005, 19, 768–782. [Google Scholar] [CrossRef]
- De Matos, T.P.V.; De Matos, V.P.V.; De Mello, K.; Valente, R.A. Protected Areas and Forest Fragmentation: Sustainability Index for Prioritizing Fragments for Landscape Restoration. Geol. Ecol. Landsc. 2021, 5, 19–31. [Google Scholar] [CrossRef]
- Liu, J.; Coomes, D.A.; Gibson, L.; Hu, G.; Liu, J.; Luo, Y.; Wu, C.; Yu, M. Forest Fragmentation in China and Its Effect on Biodiversity. Biol. Rev. 2019, 94, 1636–1657. [Google Scholar] [CrossRef]
- Zuidema, P.A.; Sayer, J.A.; Dijkman, W. Forest Fragmentation and Biodiversity: The Case for Intermediate-Sized Conservation Areas. Environ. Conserv. 1996, 23, 290–297. [Google Scholar] [CrossRef]
- Townsend, P.A.; Lookingbill, T.R.; Kingdon, C.C.; Gardner, R.H. Spatial Pattern Analysis for Monitoring Protected Areas. Remote Sens. Environ. 2009, 113, 1410–1420. [Google Scholar] [CrossRef]
- McFeeters, S.K. The Use of the Normalized Difference Water Index (NDWI) in the Delineation of Open Water Features. Int. J. Remote Sens. 1996, 17, 1425–1432. [Google Scholar] [CrossRef]
- Hancock, D.W.; Dougherty, C.T. Relationships between Blue- and Red-based Vegetation Indices and Leaf Area and Yield of Alfalfa. Crop Sci. 2007, 47, 2547–2556. [Google Scholar] [CrossRef]
- Gobron, N.; Pinty, B.; Verstraete, M.M.; Widlowski, J.-L. Advanced Vegetation Indices Optimized for Up-Coming Sensors: Design, Performance, and Applications. IEEE Trans. Geosci. Remote Sens. 2000, 38, 2489–2505. [Google Scholar]
- Puzachenko, Y.G.; Sandlersky, R.B.; Krenke, A.N.; Puzachenko, Y.M. Multispectral Remote Information in Forest Research. Contemp. Probl. Ecol. 2014, 7, 838–854. [Google Scholar] [CrossRef]







| Category (IUCN) | Number | Total Area, ha |
|---|---|---|
| Nature Biosphere Reserve (I) 1 | 1 | 4957 |
| National Park (II) | 2 | 71,796 |
| Natural Reserve (IV) | 182 | 237,061 |
| Natural Monument (III) | 164 | 10,305 |
| Coastal recreation area (V) | 5 | 7254 |
| Strictly protected water object (V) | 1 | 7658 |
| Regional Natural Reserve (V) | 7 | 21.97 |
| Natural recreational complex (V) | 5 | 28.66 |
| Dendrological park and Botanical Garden (V) | 2 | 0.6 |
| Natural History Park (V) | 14 | 1.77 |
| Other | >100 | >4000 |
| Total | >380 | > |
| Indicator | LT#1 | MZ#2 | NSh#3 | PK#4 | KZ#5 | S#6 |
| BD area, thousand ha | 431.4 | 1670.7 | 1232.1 | 795.9 | 274.8 | 49.9 |
| Forest area of the district, thousand ha | 286.8 | 849.7 | 668.2 | 328.4 | 50.7 | 3.3 |
| Area of PAs within the BD, thousand ha | 82.7 | 67.6 | 97.4 | 16.2 | 35.1 | 0.6 |
| Number of PAs | 34 | 95 | 70 | 18 | 14 | 6 |
| PAs percentage of the district area, % | 19.17 | 4.04 | 7.90 | 2.03 | 1.28 | 1.22 |
| Average area of PAs, ha | 2432.39 | 711.34 | 1371.32 | 897.84 | 251.05 | 101.10 |
| PAs Category | Average Number of Ass. Gr. | Number of PAs | Std. Dev. |
|---|---|---|---|
| Nature Biosphere Reserve | 31.00 | 1.00 | |
| National Park | 30.50 | 2.00 | 0.71 |
| Natural Reserve | 21.04 | 156.00 | 6.68 |
| Natural Monument | 14.02 | 44.00 | 6.54 |
| Coastal recreation area | 17.7 | 3.00 | 12.74 |
| Regional Natural Reserve | 20.5 | 6.00 | 3.76 |
| Natural recreational complex | 17.3 | 3.00 | 10.07 |
| For all categories | 19.6 | 215.00 | 7.22 |
| Indicators, Units | LT#1 | MZ#2 | NSh#3 | PK#4 | KZ#5 | S#6 | |
|---|---|---|---|---|---|---|---|
| Average patch area, ha | BD | 16.31 | 13.34 | 15.27 | 16.54 | 10.10 | 8.05 |
| PAs | 17.88 | 12.89 | 12.54 | 14.50 | 60.33 | 3.73 | |
| PD, n/100 ha | BD | 3.17 | 3.55 | 3.14 | 2.41 | 1.71 | 0.78 |
| PAs | 16.87 | 14.59 | 8.68 | 7.62 | 11.57 | 3.17 | |
| LPI, % | BD | 2.80 | 0.60 | 1.54 | 1.81 | 0.44 | 1.29 |
| PAs | 42.67 | 43.95 | 38.35 | 38.51 | 57.17 | 6.33 | |
| PAFRAC, none | BD | 1.48 | 1.55 | 1.51 | 1.51 | 1.45 | 1.46 |
| PAs | 1.41 | 1.45 | 1.44 | 1.40 | 1.39 | 1.49 | |
| TECI, % | BD | 18.24 | 13.47 | 14.26 | 17.53 | 7.20 | 1.58 |
| PAs | 14.20 | 9.72 | 11.30 | 12.20 | 4.79 | 2.75 | |
| IJI,% | BD | 65.50 | 61.63 | 62.63 | 66.47 | 50.32 | 49.14 |
| PAs | 60.08 | 60.08 | 61.54 | 57.92 | 57.88 | 58.93 |
| Area, ha | Distance to Moscow, km | Perimeter, km | Pavg Feb | Pavg April | Tavg Jan | Tavg March | Date of Creation PAs | NTL | PD | LPI | PAFRAC | TECI | IJI | SPLIT | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | |
| 1 | 0.08 | −0.06 | −0.06 | −0.21 | 0.04 | −0.07 | −0.03 | −0.35 | −0.22 | 0.22 | 0.22 | −0.07 | 0.22 | ||
| 2 | 0.00 | 0.35 | −0.39 | 0.03 | −0.10 | −0.10 | 0.07 | 0.02 | 0.17 | −0.12 | −0.06 | ||||
| 3 | 0.02 | −0.04 | −0.11 | 0.07 | −0.07 | 0.10 | −0.39 | −0.30 | 0.25 | 0.13 | −0.06 | 0.18 | |||
| 4 | −0.10 | 0.29 | −0.04 | 0.06 | 0.05 | −0.01 | 0.00 | −0.21 | 0.13 | 0.04 | |||||
| 5 | −0.35 | 0.15 | −0.01 | −0.39 | 0.11 | 0.02 | 0.25 | −0.15 | −0.02 | 0.00 | |||||
| 6 | −0.10 | 0.07 | 0.38 | 0.17 | −0.09 | −0.07 | −0.11 | 0.14 | 0.04 | ||||||
| 7 | 0.06 | 0.16 | 0.08 | 0.07 | −0.02 | 0.06 | 0.01 | −0.10 | |||||||
| 8 | 0.13 | 0.10 | −0.08 | −0.03 | −0.06 | 0.06 | −0.03 | ||||||||
| 9 | 0.08 | −0.11 | −0.17 | −0.25 | 0.12 | 0.00 | |||||||||
| 10 | −0.27 | −0.12 | −0.21 | 0.46 | −0.14 | ||||||||||
| 11 | −0.22 | 0.10 | −0.28 | −0.14 | |||||||||||
| 12 | 0.23 | −0.11 | 0.22 | ||||||||||||
| 13 | −0.13 | −0.16 | |||||||||||||
| 14 | −0.08 | ||||||||||||||
| 15 |
| Area, ha | Distance to Moscow, km | P_avg Feb | T_avg Jan | T_avg March | Date of PAs Establishment | PD | LPI | PAFRAC | TECI | IJI | SPLIT |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 0.32 | −0.30 | −0.03 | 0.11 | 0.10 | 0.01 | −0.18 | −0.19 | 0.34 | 0.19 | 0.03 | −0.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chernenkova, T.; Kotlov, I.; Belyaeva, N.; Suslova, E.; Lebedeva, N. Environmental Performance of Regional Protected Area Network: Typological Diversity and Fragmentation of Forests. Remote Sens. 2023, 15, 276. https://doi.org/10.3390/rs15010276
Chernenkova T, Kotlov I, Belyaeva N, Suslova E, Lebedeva N. Environmental Performance of Regional Protected Area Network: Typological Diversity and Fragmentation of Forests. Remote Sensing. 2023; 15(1):276. https://doi.org/10.3390/rs15010276
Chicago/Turabian StyleChernenkova, Tatiana, Ivan Kotlov, Nadezhda Belyaeva, Elena Suslova, and Natalia Lebedeva. 2023. "Environmental Performance of Regional Protected Area Network: Typological Diversity and Fragmentation of Forests" Remote Sensing 15, no. 1: 276. https://doi.org/10.3390/rs15010276
APA StyleChernenkova, T., Kotlov, I., Belyaeva, N., Suslova, E., & Lebedeva, N. (2023). Environmental Performance of Regional Protected Area Network: Typological Diversity and Fragmentation of Forests. Remote Sensing, 15(1), 276. https://doi.org/10.3390/rs15010276