
Precipitation Dominates the Distribution of Species Richness on the Kunlun–Pamir Plateau


 , ,
, ,  ,
,  and
and
Abstract
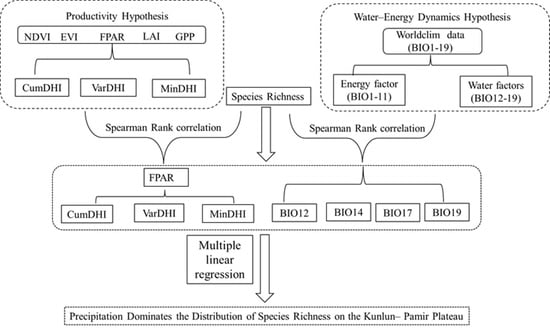
1. Introduction
2. Data and Methods
2.1. Data
2.1.1. MODIS Data
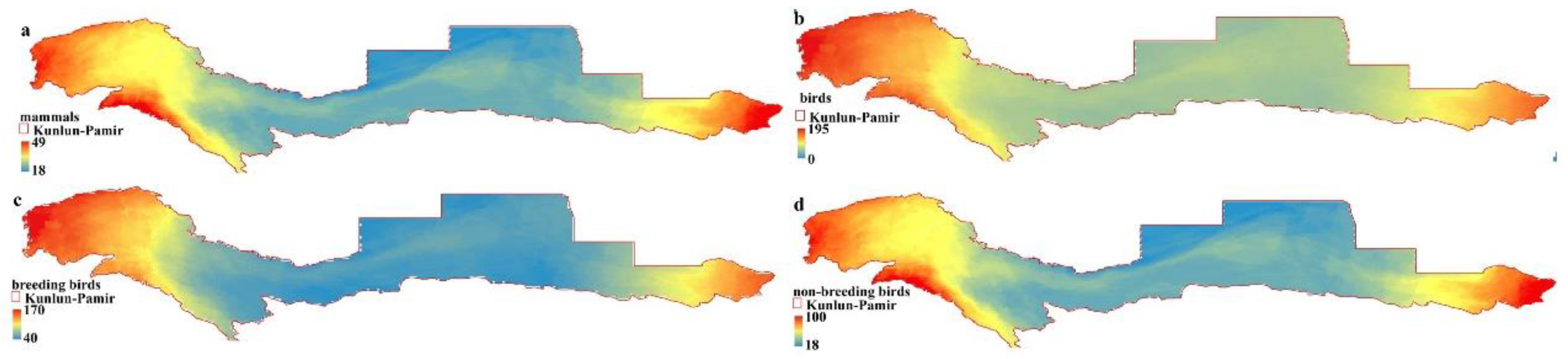
2.1.2. Species Richness
2.1.3. Climatic Data
2.2. Methods
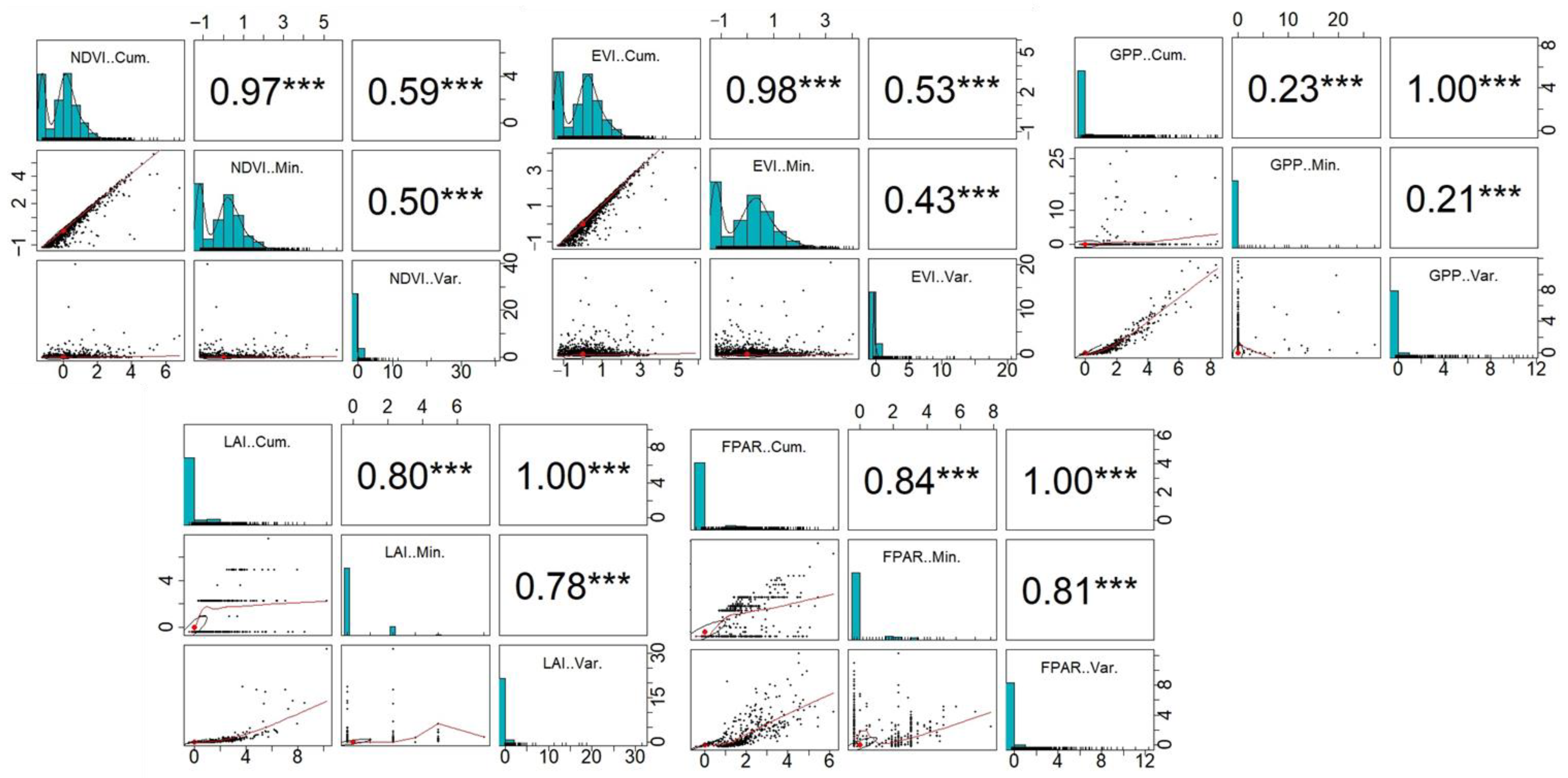
2.2.1. Calculation of the DHIs
2.2.2. Statistical Analysis
3. Results
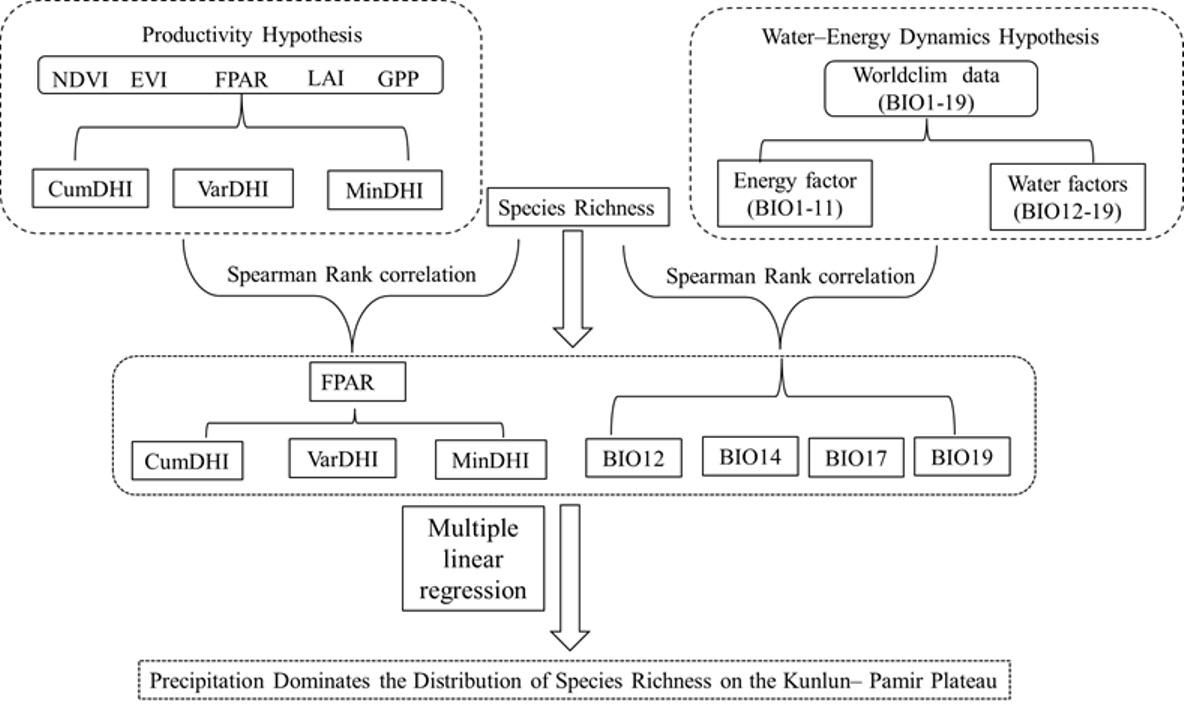
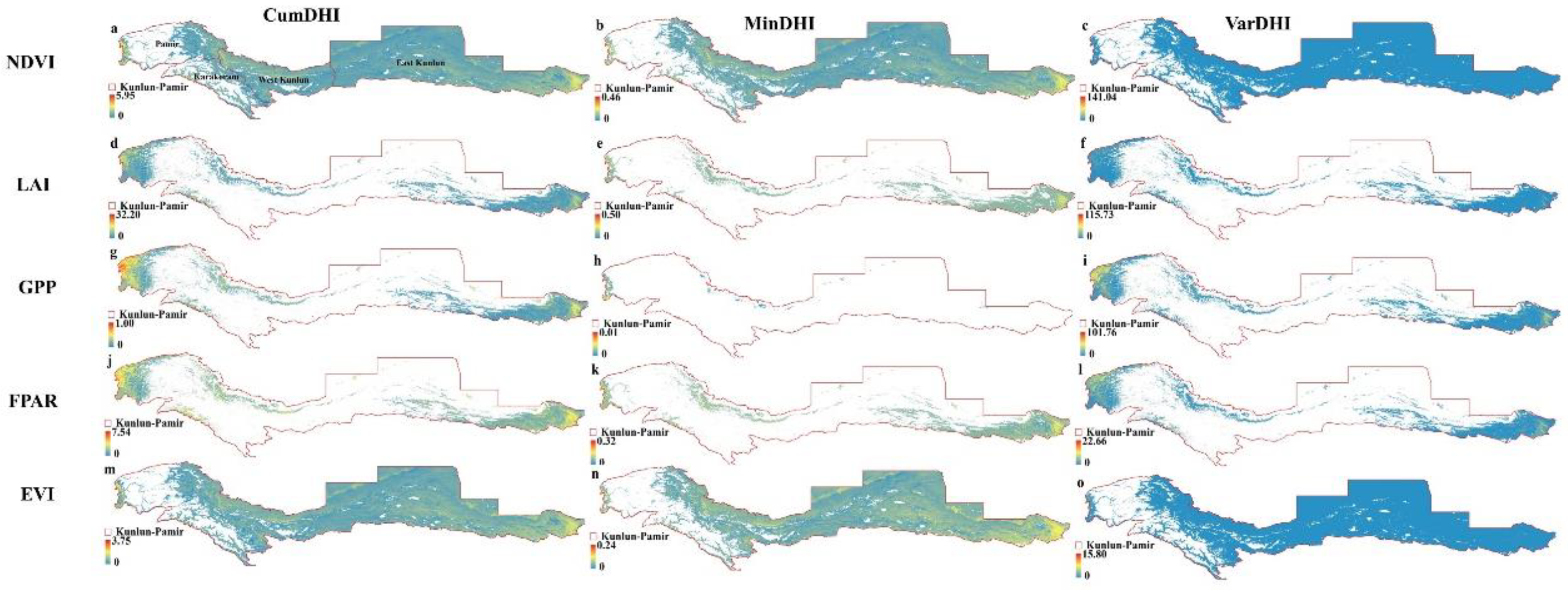
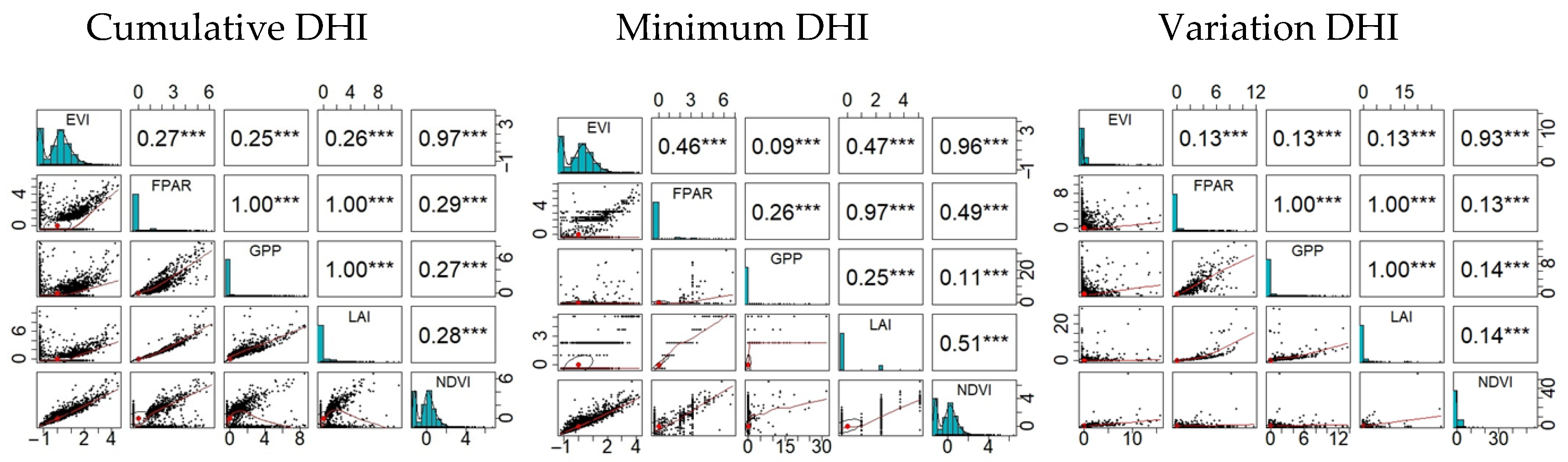
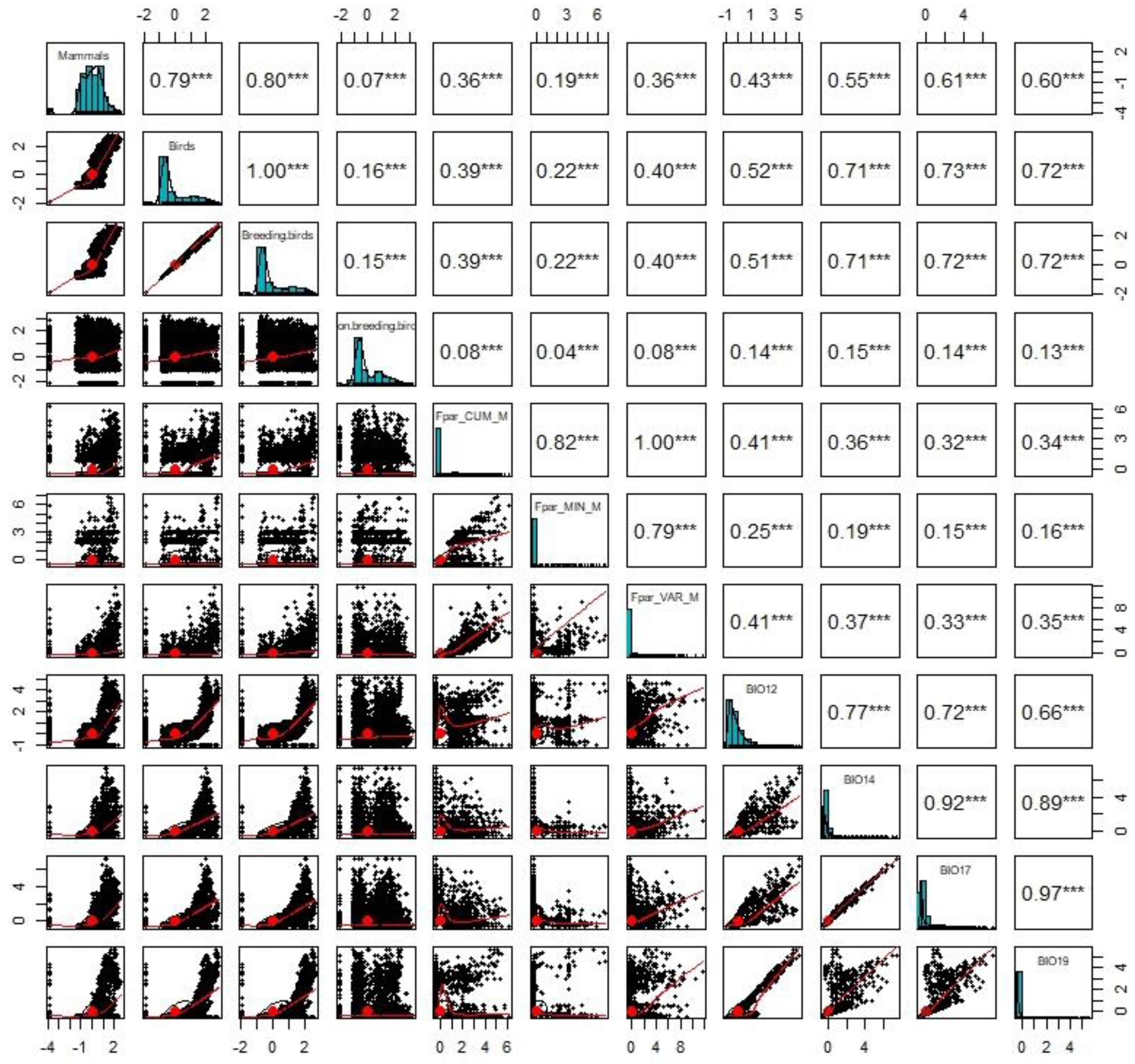
3.1. Distribution Patterns of DHIs
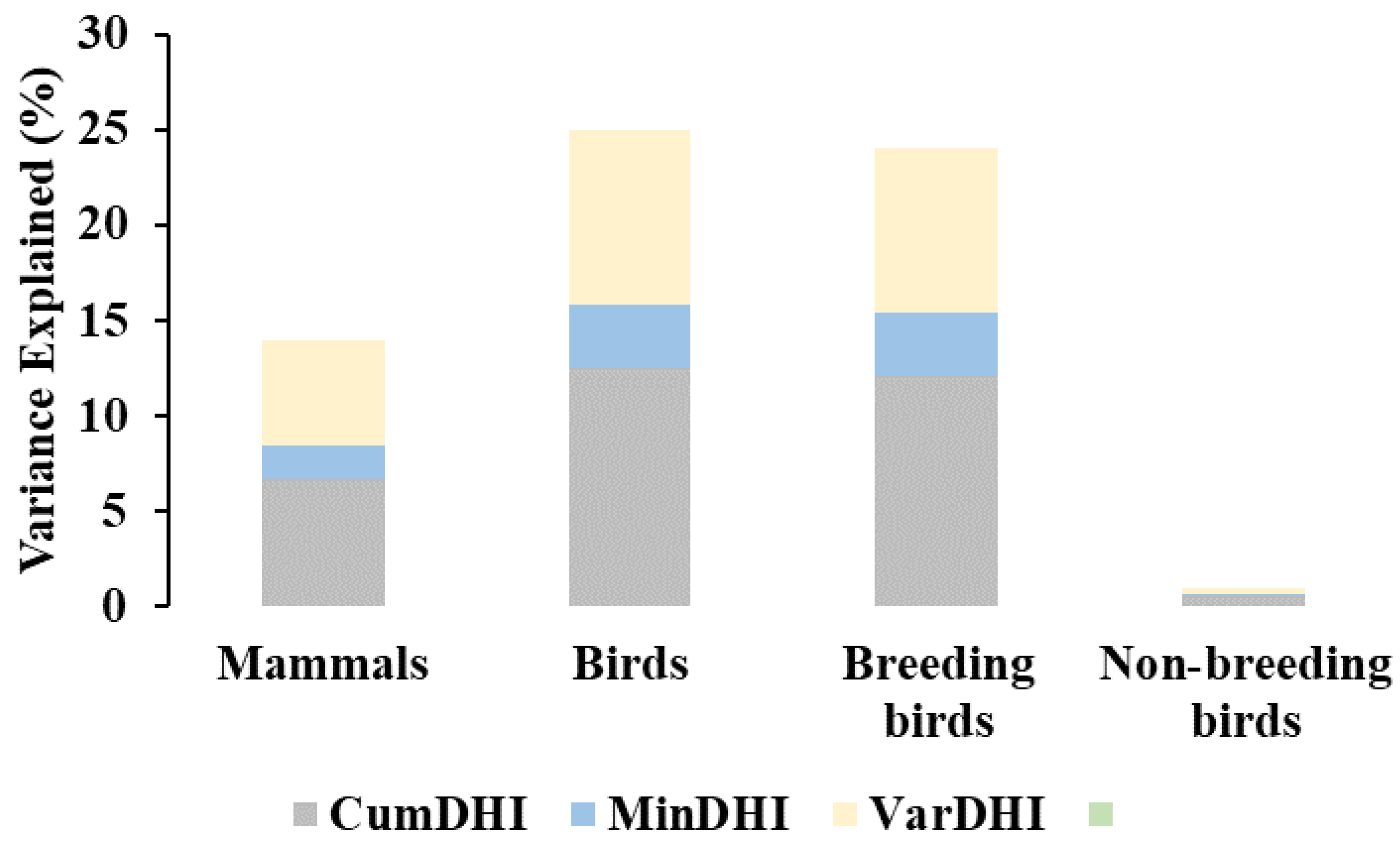
3.2. Species Richness and Productivity Hypothesis
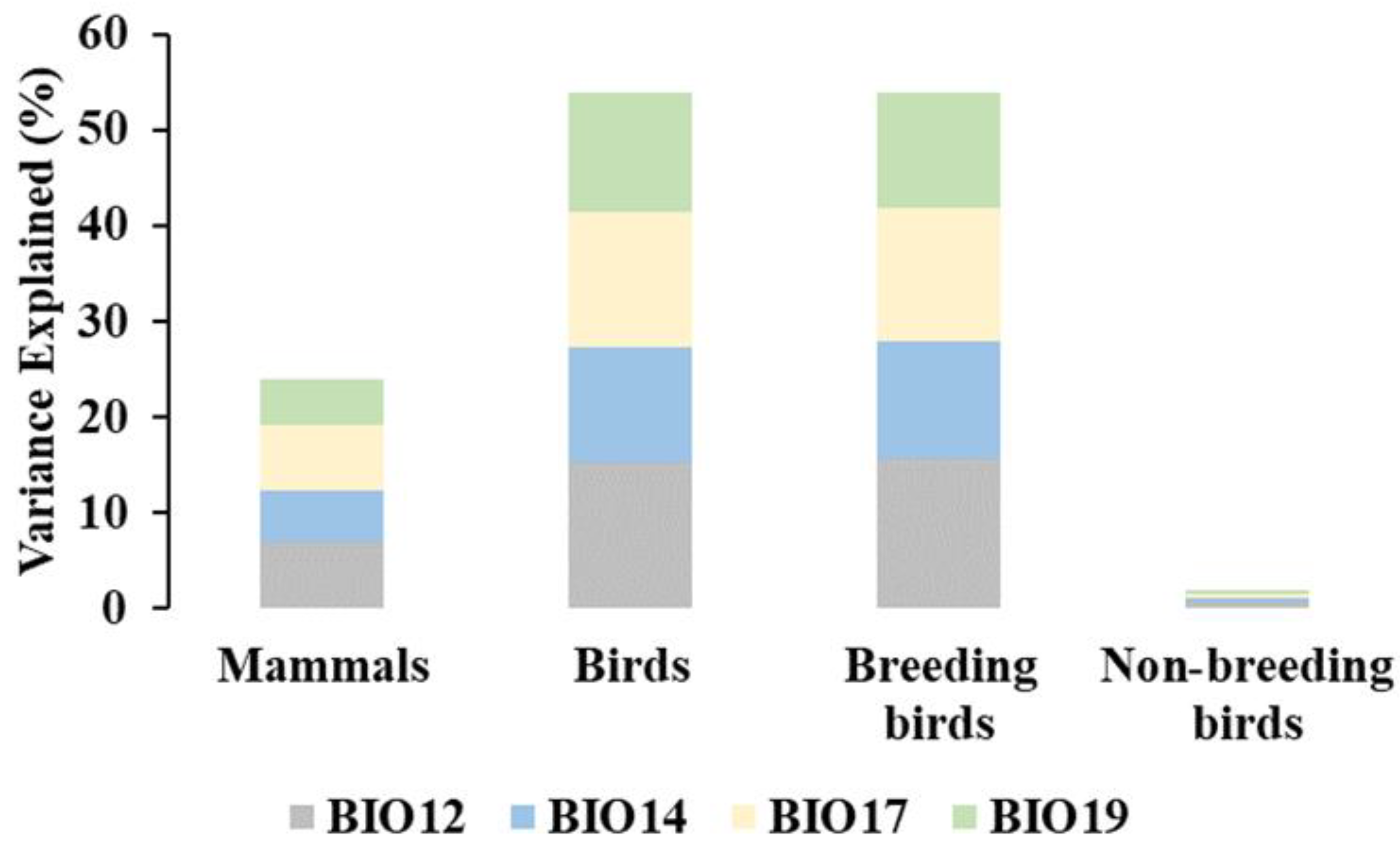
3.3. Species Richness and the Water–Energy Dynamics Hypothesis
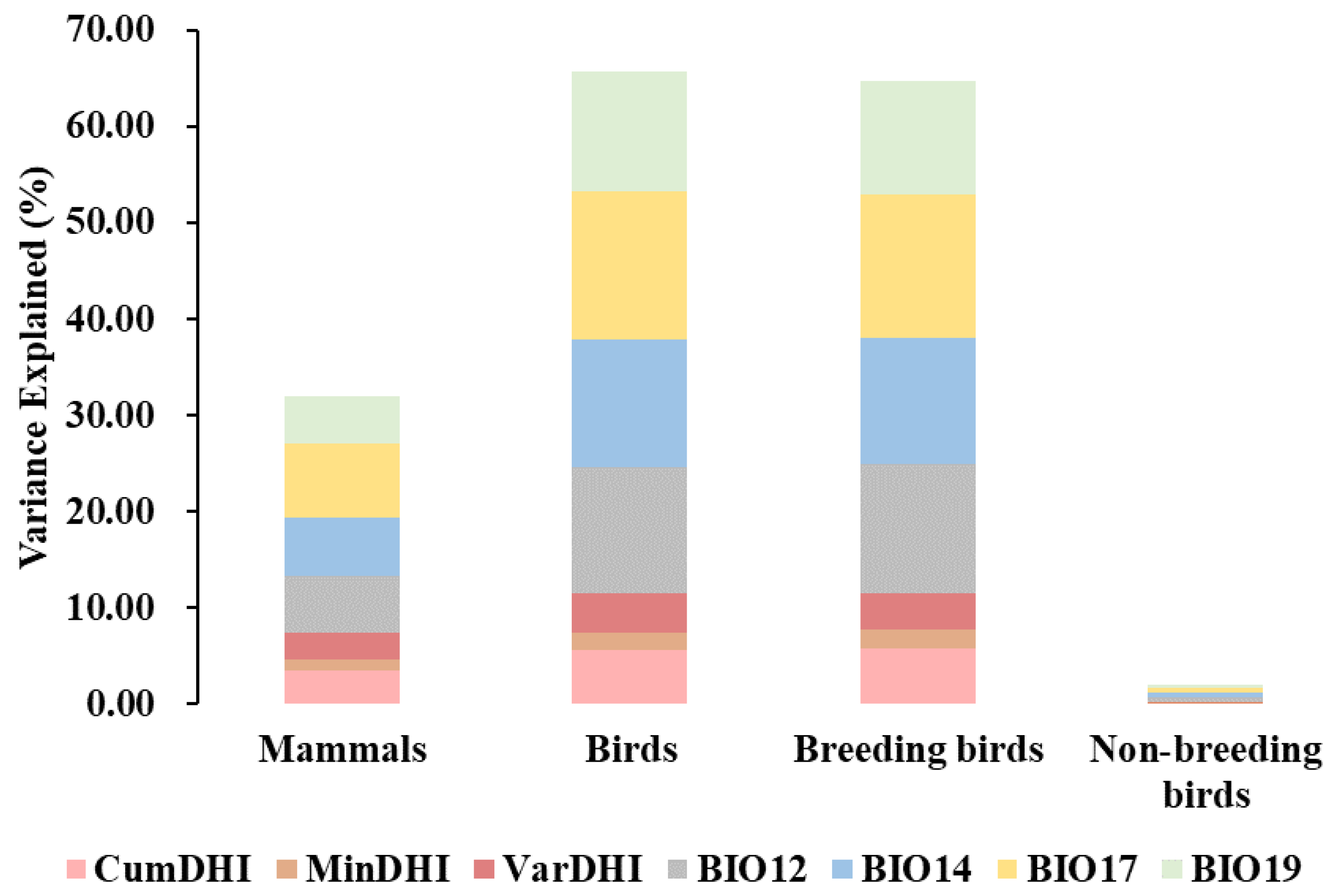
3.4. The Combined Effect of the Productivity Hypothesis and Water–Energy Dynamics Hypothesis on Species Richness
4. Discussion
4.1. The Application of the Productivity Hypothesis
4.2. The Application of the Water–Energy Dynamic Hypothesis
4.3. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Gotelli, N.J.; Colwell, R.K. Quantifying biodiversity: Procedures and pitfalls in the measurement and comparison of species richness. Ecol. Lett. 2001, 4, 379–391. [Google Scholar] [CrossRef]
- D’Antraccoli, M.; Roma-Marzio, F.; Carta, A.; Landi, S.; Bedini, G.; Chiarucci, A.; Peruzzi, L. Drivers of floristic richness in the Mediterranean: A case study from Tuscany. Biodivers. Conserv. 2019, 28, 1411–1429. [Google Scholar] [CrossRef]
- Balmford, A.; Green, R.E.; Jenkins, M. Measuring the changing state of nature. Trends Ecol. Evol. 2003, 18, 326–330. [Google Scholar] [CrossRef]
- Butchart, S.H.M.; Walpole, M.; Collen, B.; van Strien, A.; Scharlemann, J.P.W.; Almond, R.E.A.; Baillie, J.E.M.; Bomhard, B.; Brown, C.; Bruno, J.; et al. Global Biodiversity: Indicators of Recent Declines. Science 2010, 328, 1164–1168. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.H. Two Decades of Homage to Santa Rosalia: Toward a General Theory of Diversity. Am. Zool. 1981, 21, 877–888. [Google Scholar] [CrossRef]
- Wright, D.H. Species-Energy Theory: An Extension of Species-Area Theory. Oikos 1983, 41, 496. [Google Scholar] [CrossRef]
- O’Brien, E.M. Climatic Gradients in Woody Plant Species Richness: Towards an Explanation Based on an Analysis of Southern Africa’s Woody Flora. J. Biogeogr. 1993, 20, 181–198. [Google Scholar] [CrossRef]
- Hawkins, B.A.; Porter, E.E. Water-energy balance and the geographic pattern of speciesrichness of western Palearctic butterflies. Ecol. Entomol. 2010, 28, 678–686. [Google Scholar] [CrossRef]
- Currie, D.J. Energy and large-scale patterns of animal- andplant-species richness. Am. Nat. 1991, 137, 27–49. [Google Scholar] [CrossRef]
- Turner, J.R. Explaining the global biodiversity gradient: Energy, area, history and natural selection. Basic Appl. Ecol. 2004, 5, 435–448. [Google Scholar] [CrossRef]
- Hawkins, B.A.; Field, R.; Cornell, H.V.; Currie, D.J.; Guégan, J.; Kaufman, D.M.; Kerr, J.T.; Mittelbach, G.G.; Oberdorff, T.; O’Brien, E.M.; et al. Energy, water, and broad-scale geographic patternsof species richness. Ecology 2003, 84, 3105–3117. [Google Scholar] [CrossRef]
- Gillooly, J.F.; Brown, J.H.; West, G.B.; Savage, V.M.; Charnov, E.L. Effects of size and temperature on metabolic rate. Science 2001, 293, 2248–2251. [Google Scholar] [CrossRef] [PubMed]
- Allen, A.P.; Brown, J.H.; Gillooly, J.F. Global biodiversity, biochemical kinetics, and the energetic-equivalence rule. Science 2002, 297, 1545–1548. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.H.; Gillooly, J.F.; Allen, A.P.; Savage, V.M.; West, G.B. Toward a metabolic theory of ecology. Ecology 2004, 85, 1771–1789. [Google Scholar] [CrossRef]
- Gaston, K.J. Global patterns in biodiversity. Nature 2000, 405, 220–227. [Google Scholar] [CrossRef]
- Evans, K.L.; James, N.; Gaston, K.J. Abundance, species richness and energy availability in the North American avifauna. Glob. Ecol. Biogeogr. 2006, 15, 372–385. [Google Scholar] [CrossRef]
- O’Brien, E.M. Biological relativity to water?energy dynamics. J. Biogeogr. 2010, 33, 1868–1888. [Google Scholar] [CrossRef]
- Qian, H.; Wang, X.; Wang, S.; Li, Y. Environmental determinants of amphibian and reptile species richness in China. Ecography 2007, 30, 471–482. [Google Scholar] [CrossRef]
- Huang, C.-y. Using Geographical Variation of Water and Energy to Explainthe Spatial Distribution Patterns of Pteridophyta of China; Shanxi University: Taiyuan, China, 2018. [Google Scholar]
- Hurlbert, A.H.; Haskell, J.P. The effect of energy and seasonality on avian species richness and community composition. Am. Nat. 2003, 161, 83–97. [Google Scholar] [CrossRef]
- St-Louis, V.; Pidgeon, A.M.; Clayton, M.K.; Locke, B.A.; Bash, D.; Radeloff, V.C. Satellite image texture and a vegetation index predict avian biodiversity in the Chihuahuan Desert of New Mexico. Ecography 2009, 32, 468–480. [Google Scholar] [CrossRef]
- Hobi, M.L.; Dubinin, M.; Graham, C.H.; Coops, N.C.; Clayton, M.K.; Pidgeon, A.M.; Radeloff, V.C. A comparison of Dynamic Habitat Indices derived from different MODIS products as predictors of avian species richness. Remote Sens. Environ. 2017, 195, 142–152. [Google Scholar] [CrossRef]
- Scholes, R.J.; Walters, M.; Turak, E.; Saarenmaa, H.; Heip, C.H.; Tuama, É.Ó.; Faith, D.P.; Mooney, H.A.; Ferrier, S.; Jongman, R.H.; et al. Building a global observing system for biodiversity. Curr. Opin. Environ. Sustain. 2012, 4, 139–146. [Google Scholar] [CrossRef]
- Pereira, H.M.; Ferrier, S.; Walters, M.; Geller, G.N.; Jongman, R.H.G.; Scholes, R.J.; Bruford, M.W.; Brummitt, N.; Butchart, S.H.M.; Cardoso, A.C.; et al. Essential Biodiversity Variables. Science 2013, 339, 277–278. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Franklin, S.E.; Guo, X.; He, Y.; McDermid, G.J. Problems in remote sensing of landscapes and habitats. Prog. Phys. Geogr. Earth Environ. 2009, 33, 747–768. [Google Scholar] [CrossRef]
- Radeloff, V.; Dubinin, M.; Coops, N.; Allen, A.; Brooks, T.; Clayton, M.; Costa, G.; Graham, C.; Helmers, D.; Ives, A.; et al. The Dynamic Habitat Indices (DHIs) from MODIS and global biodiversity. Remote Sens. Environ. 2019, 222, 204–214. [Google Scholar] [CrossRef]
- Coops, N.C.; Waring, R.H.; Wulder, M.A.; Pidgeon, A.M.; Radeloff, V.C. Bird diversity: A predictable function of satellite-derived estimates of seasonal variation in canopy light absorbance across the United States. J. Biogeogr. 2009, 36, 905–918. [Google Scholar] [CrossRef]
- Coops, N.C.; Wulder, M.; Iwanicka, D. Demonstration of a satellite-based index to monitor habitat at continental-scales. Ecol. Indic. 2009, 9, 948–958. [Google Scholar] [CrossRef]
- Andrew, M.E.; Wulder, M.A.; Coops, N.C.; Baillargeon, G. Beta-diversity gradients of butterflies along productivity axes. Glob. Ecol. Biogeogr. 2012, 21, 352–364. [Google Scholar] [CrossRef]
- Michaud, J.-S.; Coops, N.C.; Andrew, M.E.; Wulder, M.A.; Brown, G.S.; Rickbeil, G.J. Estimating moose (Alces alces ) occurrence and abundance from remotely derived environmental indicators. Remote Sens. Environ. 2014, 152, 190–201. [Google Scholar] [CrossRef]
- Zhang, C.; Cai, D.; Guo, S.; Guan, Y.; Fraedrich, K.; Nie, Y.; Liu, X.; Bian, X. Spatial-Temporal Dynamics of China’s Terrestrial Biodiversity: A Dynamic Habitat Index Diagnostic. Remote Sens. 2016, 8, 227. [Google Scholar] [CrossRef]
- Razenkova, E.; Radeloff, V.C.; Dubinin, M.; Bragina, E.V.; Allen, A.M.; Clayton, M.K.; Pidgeon, A.M.; Baskin, L.M.; Coops, N.C.; Hobi, M. Vegetation productivity summarized by the Dynamic Habitat Indices explains broad-scale patterns of moose abundance across Russia. Sci. Rep. 2020, 10, 836. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Li, Z.; Zhou, P. Mass balance reconstruction for Shiyi Glacier in the Qilian Mountains, Northeastern Tibetan Plateau, and its climatic drivers. Clim. Dyn. 2020, 56, 969–984. [Google Scholar] [CrossRef]
- Jenkins, C.N.; Pimm, S.L.; Joppa, L.N. Global patterns of terrestrial vertebrate diversity and conservation. Proc. Natl. Acad. Sci. USA 2013, 110, E2602–E2610. [Google Scholar] [CrossRef] [PubMed]
- Pimm, S.L.; Jenkins, C.N.; Abell, R.; Brooks, T.M.; Gittleman, J.L.; Joppa, L.N.; Raven, P.H.; Roberts, C.M.; Sexton, J.O. The biodiversity of species and their rates of extinction, distribution, and protection. Science 2014, 344, 1246752. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, C.N.; Van Houtan, K.S. Global and regional priorities for marine biodiversity protection. Biol. Conserv. 2016, 204, 333–339. [Google Scholar] [CrossRef]
- IUCN. IUCN Red List of Threatened Species; Version 2018-1. Available online: http://www.iucnredlist.org (accessed on 7 August 2022).
- BirdLife International and Handbook of the Birds of the World. Bird Species Distribution Maps of the World. Version 7.0. 2018. Available online: http://datazone.birdlife.org/species/requestdis (accessed on 7 August 2022).
- Zhang Chunyan, G.S.; Yanning, G.; Danlu, C.; Lei, Y.W.W.; Han, X. A global dataset of monthly maximum fractions of photosyntheticallyactive radiation of the continents (2001–2010). China Sci. Data 2017, 2, 52–61. [Google Scholar]
- Coops, N.C.; Wulder, M.A.; Duro, D.C.; Han, T.; Berry, S. The development of a Canadian dynamic habitat index using multi–temporal satellite estimates of canopy light absorbance. Ecol. Indic. 2008, 8, 754–766. [Google Scholar] [CrossRef]
- Mac Nally, R.; Walsh, C.J. Hierarchical partitioning public-domain software. Biodivers. Conserv. 2004, 13, 659. [Google Scholar] [CrossRef]
- Li, L.; Wang, Z.; Zerbe, S.; Abdusalih, N.; Tang, Z.; Ma, M.; Yin, L.; Mohammat, A.; Han, W.; Fang, J. Species richness patterns and water-energy dynamics in the drylands of northwest China. PLoS ONE 2013, 8, e66450. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, E.M. Water-energy dynamics, climate, and prediction of woody plant species richness: An interim general model. J. Biogeogr. 1998, 25, 379–398. [Google Scholar] [CrossRef]
- Dai Kui, S. Estimation and impact factor of wild ungulates population in west Kunlun mountain area. J. Green Sci. Technol. 2012, 4, 3–5. [Google Scholar]
- Sutherland, W.J.; Butchart, S.H.M.; Connor, B.; Culshaw, C.; Dicks, L.V.; Dinsdale, J.; Doran, H.; Entwistle, A.C.; Fleishman, E.; Gibbons, D.W. A 2018 Horizon Scan of Emerging Issues for Global Conservation and Biological Diversity. Trends Ecol. Evol. 2018, 33, 47–58. [Google Scholar] [CrossRef]
- Ferrer-Castán, D.; Morales-Barbero, J.; Vetaas, O.R. Water-energy dynamics, habitat heterogeneity, history, and broad-scale patterns of mammal diversity. Acta Oecologica 2016, 77, 176–186. [Google Scholar] [CrossRef]
- Newbold, T.; Hudson, L.N.; Hill, S.L.L.; Contu, S.; Lysenko, I.; Senior, R.A.; Börger, L.; Bennett, D.J.; Choimes, A.; Collen, B.; et al. Global effects of land use on local terrestrial biodiversity. Nature 2015, 520, 45. [Google Scholar] [CrossRef]
- Cahill, A.E.; Aiello-Lammens, M.; Fisher-Reid, M.C.; Hua, X.; Karanewsky, C.J.; Ryu, H.Y.; Sbeglia, G.C.; Spagnolo, F.; Waldron, J.B.; Warsi, O.; et al. How does climate change cause extinction? Proc. R. Soc. B Boil. Sci. 2013, 280, 20121890. [Google Scholar] [CrossRef]
- Wang, X.; Liu, S.; Ding, Y.; Guo, W.; Jiang, Z.; Lin, J.; Han, Y. An approach for estimating the breach probabilities of moraine-dammed lakes in the Chinese Himalayas using remote-sensing data. Nat. Hazards Earth Syst. Sci. 2012, 12, 3109–3122. [Google Scholar] [CrossRef]
- Larsen, L.G.; Harvey, J. Modeling of hydroecological feedbacks predicts distinct classes of landscape pattern, process, and restoration potential in shallow aquatic ecosystems. Geomorphology 2010, 126, 279–296. [Google Scholar] [CrossRef]
- Chen, S.; Mao, L.; Zhang, J.; Zhou, K.; Gao, J. Environmental determinants of geographic butterflyrichness pattern in eastern China. Biodivers. Conserv. 2014, 23, 1453–1467. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Name | Platform | Temporal Resolution (Day) | Spatial Resolution (m) |
|---|---|---|---|---|
| NDVI | MOD13A1 | Terra | 16 | 500 |
| EVI | MOD13A1 | Terra | 16 | 500 |
| FPAR | MOD15A2H | Terra | 8 | 500 |
| LAI | MOD15A2H | Terra | 8 | 500 |
| GPP | MOD17A2HGF | Terra | 8 | 500 |
| Variables | Description | |
|---|---|---|
| Energy factor | BIO1 | Annual Mean Temperature |
| BIO2 | Mean Diurnal Range | |
| BIO3 | Isothermality | |
| BIO4 | Temperature Seasonality | |
| BIO5 | Max Temperature of Warmest Month | |
| BIO6 | Min Temperature of Coldest Month | |
| BIO7 | Temperature Annual Range | |
| BIO8 | Mean Temperature of Wettest Quarter | |
| BIO9 | Mean Temperature of Driest Quarter | |
| BIO10 | Mean Temperature of Warmest Quarter | |
| BIO11 | Mean Temperature of Coldest Quarter | |
| Water factors | BIO12 | Annual Precipitation |
| BIO13 | Precipitation of Wettest Month | |
| BIO14 | Precipitation of Driest Month | |
| BIO15 | Precipitation Seasonality | |
| BIO16 | Precipitation of Wettest Quarter | |
| BIO17 | Precipitation of Driest Quarter | |
| BIO18 | Precipitation of Warmest Quarter | |
| BIO19 | Precipitation of Coldest Quarter |
| Cumulative DHI R2 Adj | RMSE | Minimum DHI R2 Adj | RMSE | Variation DHI R2 Adj | RMSE | All DHIs R2 Adj | RMSE | |
|---|---|---|---|---|---|---|---|---|
| Mammals | 0.12 *** | 7.03 | 0.03 *** | 7.38 | 0.12 *** | 7.04 | 0.14 *** | 6.98 |
| Birds | 0.20 *** | 37.10 | 0.04 *** | 40.59 | 0.19 *** | 37.42 | 0.25 *** | 36.73 |
| Breeding birds | 0.20 *** | 34.95 | 0.05 *** | 38.17 | 0.18 *** | 35.40 | 0.24 *** | 34.66 |
| Non-breeding birds | 0.00 *** | 18.97 | 0.00 ** | 19.01 | 0.00 *** | 18.98 | 0.01 *** | 18.97 |
| BIO12 R2 Adj | RMSE | BIO14 R2 Adj | RMSE | BIO17 R2 Adj | RMSE | BIO19 R2 Adj | RMSE | The Four BIOs R2 Adj | RMSE | |
|---|---|---|---|---|---|---|---|---|---|---|
| Mammals | 0.24 *** | 6.54 | 0.20 *** | 6.70 | 0.23 *** | 6.59 | 0.21 *** | 6.65 | 0.29 *** | 6.32 |
| Birds | 0.54 *** | 28.12 | 0.49 *** | 29.60 | 0.53 *** | 28.48 | 0.52 *** | 28.91 | 0.61 *** | 25.82 |
| Breeding birds | 0.54 *** | 26.55 | 0.49 *** | 27.92 | 0.52 *** | 27.07 | 0.50 *** | 18.85 | 0.60 *** | 24.69 |
| Non-breeding birds | 0.02 *** | 18.84 | 0.02 *** | 18.86 | 0.02 *** | 18.85 | 0.02 * | 18.85 | 0.02 *** | 18.82 |
| Four BIOs and Three FPAR-Based DHIs R2 Adj | RMSE | |
|---|---|---|
| Mammals | 0.32 *** | 6.18 |
| Birds | 0.66 *** | 24.34 |
| Breeding birds | 0.65 *** | 23.28 |
| Non-breeding birds | 0.02 *** | 18.82 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, X.; Bao, A.; Zhang, J.; Yu, T.; Zheng, G.; Yuan, Y.; Wang, T.; Nzabarinda, V.; De Maeyer, P.; Van de Voorde, T. Precipitation Dominates the Distribution of Species Richness on the Kunlun–Pamir Plateau. Remote Sens. 2022, 14, 6187. https://doi.org/10.3390/rs14246187
Huang X, Bao A, Zhang J, Yu T, Zheng G, Yuan Y, Wang T, Nzabarinda V, De Maeyer P, Van de Voorde T. Precipitation Dominates the Distribution of Species Richness on the Kunlun–Pamir Plateau. Remote Sensing. 2022; 14(24):6187. https://doi.org/10.3390/rs14246187
Chicago/Turabian StyleHuang, Xiaoran, Anming Bao, Junfeng Zhang, Tao Yu, Guoxiong Zheng, Ye Yuan, Ting Wang, Vincent Nzabarinda, Philippe De Maeyer, and Tim Van de Voorde. 2022. "Precipitation Dominates the Distribution of Species Richness on the Kunlun–Pamir Plateau" Remote Sensing 14, no. 24: 6187. https://doi.org/10.3390/rs14246187
APA StyleHuang, X., Bao, A., Zhang, J., Yu, T., Zheng, G., Yuan, Y., Wang, T., Nzabarinda, V., De Maeyer, P., & Van de Voorde, T. (2022). Precipitation Dominates the Distribution of Species Richness on the Kunlun–Pamir Plateau. Remote Sensing, 14(24), 6187. https://doi.org/10.3390/rs14246187