1. Introduction
Planting cover crops is recognized as an important conservation practice to promote agricultural sustainability via the provisioning of agro-ecosystem services [
1]. Some of these well-documented agro-ecosystem services include the protection of soil from erosion, increased nutrient retention, reduction in nutrient leaching losses, reduction in disease and pest infestations, increased weed suppression, increased infiltration, reduced overland water flow, soil health improvements via carbon sequestration, and developing resilient cropping systems that better adapt and mitigate climate change [
2,
3,
4,
5,
6]. Cover crop performance estimations, such as percent ground cover and biomass accumulation, are directly related to the magnitude of agro-ecosystem services that cover crops can provide [
4,
7,
8]. Despite the well-recognized relationship between cover crop performance and the associated benefits to cropping systems, few studies to date have assessed the ability of remote sensing to estimate cover crop performance, especially at the landscape scale.
Remote sensing is a powerful tool capable of producing landscape-level assessments of cover crop presence and absence [
9,
10], emergence and termination [
11,
12], and performance [
13,
14,
15]. To date, remote sensing in cover crop research has primarily employed optical sensing approaches using proximal (hand-held) sensors [
14], unoccupied aerial vehicles (UAVs) [
16,
17], high-resolution aerial images [
18], and Earth-observing satellites [
8,
10,
13,
19]. Optical vegetation indices (VIs) used to estimate plant biomass usually exploit the relationship between visible light with low reflectance (linked to plant pigment concentration) and high reflectance observed in near-infrared (NIR) light (influenced by internal leaf structure) [
20]. This visible-NIR reflectance difference is enhanced during plant growth and manifests as increased fractional cover, which is linked to plant biomass. Both the triangular vegetation index (TVI) [
21] and the normalized difference vegetation index (NDVI) [
22] exploit the visible-NIR difference and have been used to successfully estimate biomass in cereal grass cover crop species [
13,
14].
VIs derived from optical imagery, particularly NDVI, tend to saturate at a leaf area index (LAI) greater than three in numerous plant species [
23,
24]. Similarly, previous work observed NDVI saturation at approximately 1500 kg ha
−1 biomass in cereal grass species [
14]. This substantially limits our ability to estimate biomass values at the higher ranges (i.e., >1500 kg ha
−1) that are frequently observed in the springtime, particularly in late-terminated cover crop fields under warmer growing conditions. Although red-edge bands were shown to be more resistant to saturation [
17,
24,
25,
26] they have only recently been incorporated into earth-observing platforms such as the European Space Agency’s (ESA) Sentinel-2 satellites (launched in June 2015 and March 2017). Sentinel-2 satellites contain three red-edge bands and have a 20-m spatial resolution and a five-day revisit time, resulting in a greater spatial and temporal resolution than Landsat satellites. Red-edge indices such as the normalized difference red-edge index (NDVI_RE) and the simple ratio red-edge (SR_RE) have shown strong relationships with chlorophyll A [
27], which, in turn, is closely linked with crop biophysical characteristics such as LAI and fractional ground cover [
28,
29]. Additionally, previous work demonstrated that the visibly atmospherically resistant index using a red-edge band (VARI_RE) was better able to predict the LAI of corn (
Zea mays L.) past the point of saturation (i.e., up to LAI values of six) [
30]. In contrast, other studies found that red-edge-based indices did not improve biomass estimation in corn and soybeans (
Glycine max L.) when compared to traditional VIs such as NDVI [
31]. In addition to red-edge based indices, new VIs, such as the NIR reflectance of vegetation index (NIRv) [
32] and kernel NDVI (kNDVI) [
33], have also been developed to address saturation issues with optical remote sensing. NIRv has shown good agreement with gross primary production (GPP) in corn and soybeans [
32]. Finally, kNDVI is a nonlinear generalization of NDVI that is more resistant to saturation [
33] and, therefore, may also be useful in estimating higher cover crop biomass levels beyond the point of saturation.
Active remote sensing approaches such as synthetic aperture radar (SAR) also have emerged as a viable technique for monitoring and estimating crop biophysical characteristics [
34,
35,
36], although most studies focus on the use of SAR data in crop classification. SARs operate at microwave wavelengths (0.18–30 cm), emit coherent signals, and measure the backscattered intensity (backscatter) and phase of the return signals. The intensity and phase of the return signal at a given frequency and polarization can be used to elucidate the ‘polarimetric signatures’ of landscape features based on the type of scattering that is present [
37]. Agricultural studies have most commonly utilized SAR imagery in either the L-band (15–30 cm) or C-band (3.75–7.5 cm) wavelength ranges, often with two linear polarizations (dual-pol) or four linear polarizations (quad-pol). The unique information contained in SAR polarimetric channels, such as a change from transmitted horizontal (H) to return vertical (V) backscatter or from V to H (i.e., cross-polarization), provides information on the presence of volume scattering targets. Vegetation exhibits strong volume scattering; thus, cross-polarization backscatter has been shown to be well correlated with total biomass in wheat (
Triticum aestivum L.) and soybeans [
38,
39]. Similarly, the radar vegetation index (RVI), which is produced using co- and cross-polarization backscatter, has been shown to be sensitive to vegetation water content and LAI [
40].
In addition to the backscatter intensity, SAR signals also contain phase information. Combining polarimetric backscatter and polarimetric phase information provides additional information, allowing for more refined classifications of ground targets with the use of polarimetric decompositions [
36]. For instance, the polarimetric parameters decomposed from SAR data—such as entropy, anisotropy, and alpha angle—can successfully predict crop biophysical characteristics such as biomass and height under certain conditions [
41,
42,
43]. Finally, the cross-channel correlation coefficient known as the interferometric SAR (InSAR) coherence compares the phase information from two repeat-pass SAR images. When phase differences are similar for a neighborhood of pixels, InSAR coherence values will be high, and when phase differences are random, InSAR coherence values will be low. Reductions in InSAR coherence are often indicative of changes in target properties between two images, such as changes in soil roughness, soil moisture, vegetation water content, and vegetation structure [
44,
45]. However, InSAR coherence may also be useful in biomass estimation, as it quantifies phase change between two images and can, therefore, be used to estimate crop height [
46,
47] and growth [
48]. Although the use of SAR to estimate crop biophysical characteristics is on the rise, few studies to date have integrated both optical and SAR data [
49]. Moreover, to our knowledge, none focus specifically on small-stature winter cover crops that undergo a unique phenological development that includes fall establishment followed by a winter dormancy period and later springtime growth.
The ESA’s publicly available multispectral (Sentinel-2) and SAR (Sentinel-1; dual polarization, C-band) satellite imagery provides a unique opportunity to assess the efficacy of linking contemporary VIs using red-edge bands and SAR-derived metrics such as backscatter intensities, polarimetric parameters, and InSAR coherence. When integrated, these two data streams may enable a more accurate prediction of winter cover crop biomass beyond the previously identified saturation points. Thus, our first objective was to determine which optical index from Sentinel-2 was best-suited to estimating the cover crop biomass. Secondly, we evaluated whether the addition of SAR backscatter intensities, polarimetric parameters, and InSAR coherence to optical data increased our ability to estimate cover crop biomass, especially over 1500 kg ha
−1 [
14]. We hypothesized that: (1) non-normalized, red-edge indices would outperform other VIs for estimating winter cover crop biomass and (2) that the integration of optical and SAR metrics would result in better biomass estimation in winter cover crops. To test these hypotheses, we collected destructive biomass samples over three winter–spring cover crop growing seasons from cover crop fields planted with cereal grass species at the U.S. Department of Agriculture (USDA), Beltsville Agricultural Research Center (BARC) facility located in Beltsville, Maryland, United States, and regressed a suite of remote sensing measures to biomass.
2. Methods
2.1. Study Area and Sampling Plan
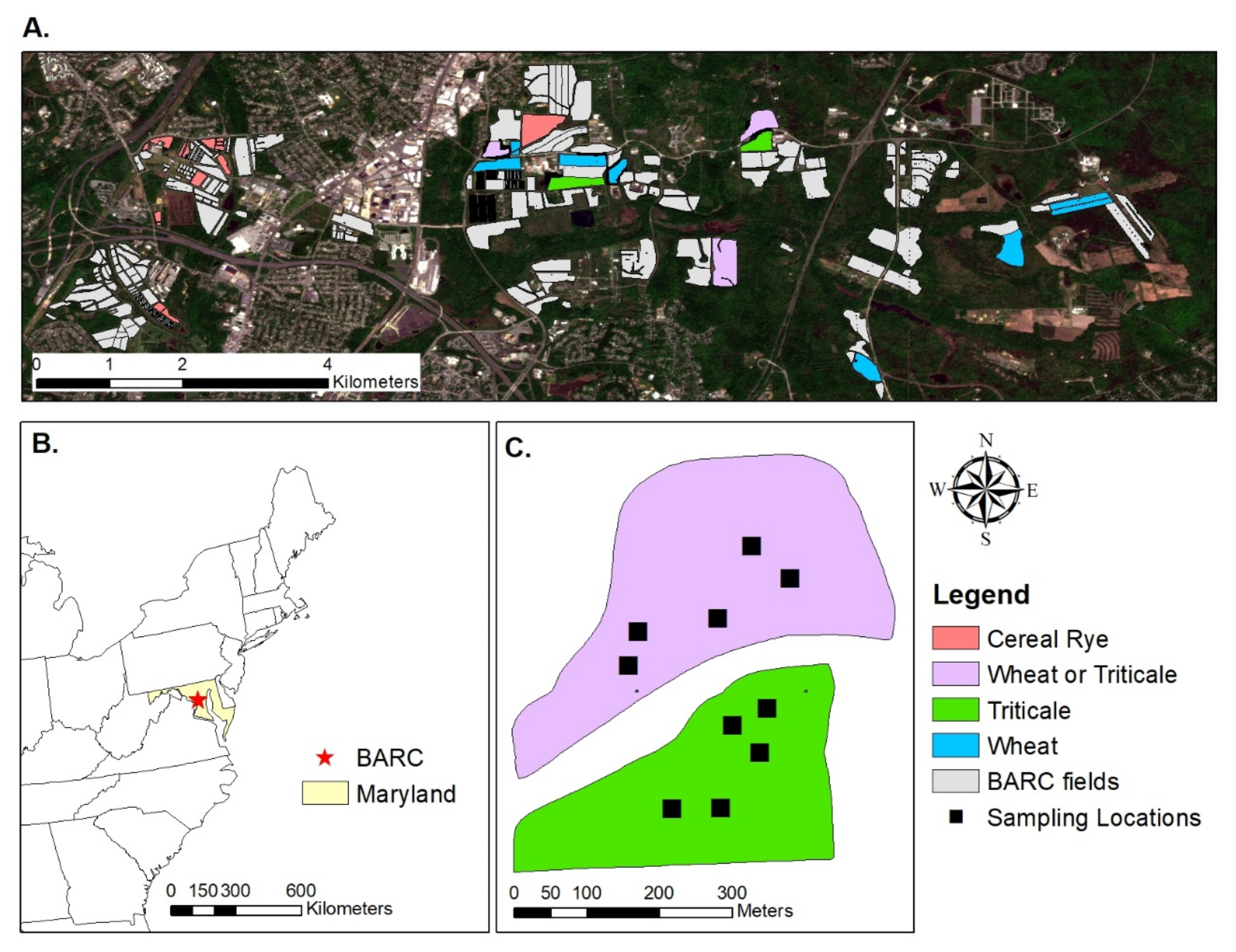
We sampled 27 cover crop fields (~5 sampling locations per field) over three winter-to-spring seasons (2018–2019, 2019–2020, and 2020–2021) at the USDA-BARC research facility (
Figure 1). These fields were planted with one of the three cereal grass cover crop species: wheat (
n = 431 across 10 fields), ‘Aroostook’ cereal rye (
Secale cereale;
n = 153 across 14 fields), and triticale (
Triticale hexaploide Lart.;
n = 185 across six fields). Cover crops were planted between 26 September and 18 October of a given year, after harvest of the previous summer crop (usually corn, soybeans, or alfalfa (
Medicago sativa)). Some of the fields—those in triticale and some wheat—were fertilized in the fall, as these fields were harvested later in the spring of the following year (typically in April) for dairy silage production.
During the 2019–2020 and 2020–2021 seasons, we sampled approximately five locations per field at each sampling time. The sampling dates were distributed throughout the cover crop growth period: once per month from October–February, twice per month from March–April, and once in May. We performed more frequent sampling in the springtime due to the faster cover crop growth and development. Sampling locations were kept consistent within seasons. During the spring of 2019, we collected cover crop biomass twice (late April–early May). We flagged five sampling locations per field with the goal of capturing spatial variability in cover crop performance (i.e., capturing areas of low and high biomass). Across all seasons, we selected sampling locations with locally uniform growth towards the center of each field to avoid edge effects and mixed pixels. Samples were spaced at least 30 m apart to avoid collections within the same pixel.
Destructive biomass samples were collected near flagged locations within each field. At each flag, we sampled aboveground cover crop biomass from ~0.57 m−2 areas in paper bags (i.e., harvested 1-m length cover crop from three adjacent drilled rows that were 19 cm apart). Paper bags containing fresh biomass samples were then dried in an oven for approximately one week at 60 °C and the dry weights were recorded. We then scaled these values from g m−2 to kg ha−1.
During the 2019–2020 and 2020–2021 growing seasons, we also collected a series of nadir photographs from a red, green, blue (RGB) Canon PowerShot G16 camera (12.1 megapixels). In previous work, nadir photographs were used to estimate fractional vegetative cover [
14]. However, as we did not have complete time series photographs associated with every sampling event, we could not include fractional vegetative cover as a covariate in our analysis. Instead, we used photographs and estimates of fractional vegetative cover where possible to interpret the relationship between the remotely sensed data and the destructively sampled biomass. We calculated fractional vegetative cover for each photograph by applying threshold values to the digital numbers for red/green ratio [
50] and excess green [
51] VIs.
2.2. Growing Degree Days
We calculated growing degree days (GDD) to assess inter-annual weather differences in cover crop growth across our study years. We used a combination of weather station data from 2019 to 2020 at BARC and the daily Daymet [
52,
53] estimations from a local weather station in Beltsville in 2021 to calculate GDD using the following equation:
where
Tmin and
Tmax were the daily minimum and maximum temperatures. We used 4 °C for
Tbase, based on prior studies for small-grain crops [
14,
54].
2.3. Soil Moisture Collections and Analyses
Sentinel-1 transmits C-band signals, which do not penetrate deeply into the soil column, and instead interact primarily with the soil surface. Because C-band signals respond strongly to moisture content, we calculated the correlation coefficients of four Sentinel-1 backscatter indices (VV, VH, VV/VH, and RVI) relative to measured values of soil volumetric water content (VWC) collected using a handheld sensor. Handheld collections were taken with an Acclima SDI-12 sensor in conjunction with Sentinel-1 overpasses at each of the fields on five dates in 2021 (March: 2nd, 15th, 26th, and April: 19th and 29th). Seven surface level measurements at 2 cm depth were taken near each plot, with six being taken within a 0.5 m radius of the center of the plot and the seventh in the center.
2.4. Sentinel-2 Imagery Acquisition and Processing
We procured level 2A Sentinel-2 reflectance products from ESA’s Copernicus service and extracted the band reflectance for nine bands at the 20-m pixel resolution, including blue (B2), green (B3), red (B4), three red edge bands (B5, B6, B7), narrow band NIR (B8A), and two shortwave infrared bands (SWIR, B11 and B12). We selected the image acquisition that corresponded most closely to each destructive biomass sampling date. We applied the provided 20-m resolution cloud mask at the strictest level (i.e., 0) to remove clouds and cloud shadows. In total, we used 21 scenes and out of 769 plot-date measurements 180 samples were removed from the analysis due to the presence of clouds or cloud shadows. We also removed 16 observations that had a gap of >6 days between destructive sampling and the satellite image acquisition (mean image gap = ~3 days, standard deviation of image gap = ~2 days). This resulted in a total of 573 observations [
55] for three cereal grass species (cereal rye:
n = 94, triticale:
n = 158, and wheat:
n = 321). Next, we extracted band reflectance data to calculate 27 VIs (
Table 1). These VIs were selected with a focus on (1) red-edge indices (
n = 14) and (2) previously identified indices sensitive to biomass or similar plant characteristics, such as LAI, height, or leaf/stem density (
n = 13). We processed the acquired images and extracted band reflectance using the ‘raster’ [
56] and ‘sf’ [
57] packages in the R statistical environment [
58].
2.5. Sentinel-1 Data Acquisition and Processing
We used Google Earth Engine (GEE) [
70] to process Sentinel-1 backscatter imagery with VH and VV polarizations. The Sentinel-1 ground range detected (GRD) scenes on GEE had previously been processed with ESA’s Sentinel Application Platform (SNAP) toolbox (SNAP, 2021) using the processing sequence of: updating image orbit file, GRD image border noise removal, thermal noise removal, radiometric calibration, and geometric terrain correction (orthorectification). In addition to using the VH and VV backscatter imagery, we also calculated RVI by modifying the original quad-pol equation [
40] to be suited for dual-pol as per [
66]. We also further adapted the equation because the dual-pol modifications described in [
66] included polarizations that are not present in Sentinel-1 (
Table 2).
To perform polarimetric decompositions on the same Sentinel-1 scenes that are processed to backscatter in GEE, we downloaded these scenes in single-look-complex (SLC) format from ESA to the calculate polarimetric parameters of entropy, anisotropy, and alpha. Polarimetric decompositions were performed in SNAP toolbox using the following sequence: image calibration, C2 matrix computation, polarimetric speckle filtering (5 × 5 Refined Lee), H-Alpha dual-pol decomposition (window size = 5), and geometric terrain correction. To generate Sentinel-1 InSAR coherence imagery, we used the Alaska Satellite Facility (ASF) Hybrid Pluggable Processing Pipeline (HyP3) tool [
71], which contains modified Copernicus Sentinel data 2021, processed by ESA). Sentinel-1 InSAR pairs were selected using the ASF SAR baseline tool to maintain the shortest possible temporal and perpendicular baselines (i.e., the greatest degree of temporal and spatial similarity between image acquisitions). We developed a selection protocol for the InSAR pairs ordered through HyP3, where the image with the later acquisition time in the InSAR pair was matched most closely to ground biomass sampling. Once image pairs were selected, we used the HyP3 tool to generate interferograms with embedded Gamma software, performing an interferogram generation with a 10 × 2 window size (smallest possible window size). The Sentinel-1 interferogram and associated InSAR coherence images output from this process resulted in a pixel size of approximately 40 m, meaning that the destructive biomass sampling of small plots (0.57 m
−2) was unlikely to significantly impact coherence estimation. As the backscatter imagery produced using GEE, polarimetric decomposition imagery, and InSAR coherence imagery were derived from the same Sentinel-1 images, their incidence angles were identical for a single image, and nearly identical for an InSAR pair. It is also important to note that the incidence angle of observation for the Sentinel-1 image time series was consistent over our study sites, exhibiting a spatiotemporal mean incidence angle of 41.31 degrees, with temporal standard deviation of 0.08 degrees, and a spatial standard deviation of 0.15 degrees.
2.6. Statistical Analyses
All analyses were performed in the R statistical environment [
58]. As the relationship between cover crop biomass and VIs is often non-linear [
14], we fit three regression models to estimate cover crop biomass from optical data: (1) log-transformed, (2) polynomial, and (3) segmented regression. Regression models were evaluated using various measures: goodness-of-fit statistic (adjusted R
2), error measures such as the root mean square error (RMSE) and the mean absolute error (MAE), and model fit using Akaike’s Information Criterion for small sample sizes (AICc) [
74]. We opted to use AICc over the traditional AIC to avoid overfitting our data, as the number of observations increases AICc and AIC converge, whereas, for smaller sample sizes, AICc are penalized more heavily. AICc were calculated using the following equation:
where
denotes the number of predictors in a model,
represents the maximum likelihood, and
n is the sample size.
Finally, we calculated AIC weights (AICw) for the log-linear and polynomial models. AICw quantifies the probability that a given model was the best model in the set of compared models—in our case, the best model among the 27 VIs compared for a given model type (e.g., log-linear). Note that AICc and AICw cannot be used to compare different model sets (i.e., log-linear vs. polynomial). We also checked for the normality and homoscedasticity of the residuals for each model. After identifying the best-performing optical index using the above steps (objective 1), we assessed whether there were seasonal- or species-level differences using analysis of variance (ANOVA) and slope and intercept contrasts in the R ‘emmeans’ package [
75].
To assess whether the integration of optical and SAR data could improve our estimations of cover crop biomass (objective 2), we paired the best-performing optical index with each SAR metric (n = 8) individually. As with objective 1, these models were then evaluated using adjusted R2, RMSE, MAE, AICc, and AICw. If more than one SAR metric improved model fit (i.e., AICc < −2) and these metrics were not collinear (i.e., r < 0.7), we evaluated combined models with the optical index identified in objective 1 and multiple SAR variables. We used ANOVAs to assess whether models containing both optical and SAR data were significantly better than models with only optical predictors. Finally, we conducted cross-validation using a 70–30% train–test split iterated 100 times, and used adjusted R2, RMSE, and MAE to summarize these results.
3. Results
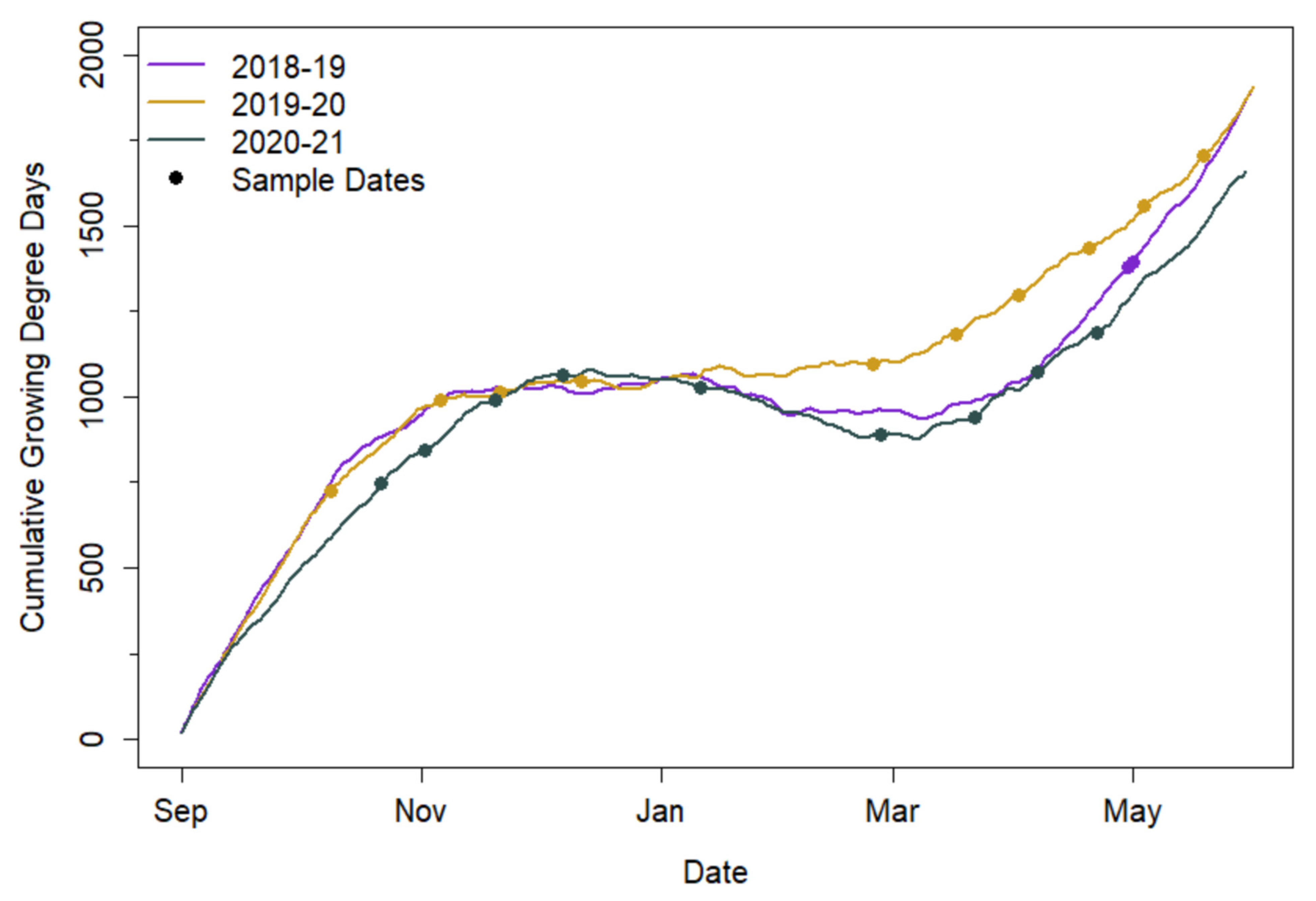
Cumulative GDD did not vary substantially among study years (
Figure 2). Despite similarities in cumulative GDD during the cover crop growth period over the years, cover crop biomass production varied substantially. Destructively sampled cover crop biomass ranged from 141 to 4498 kg ha
−1 in spring 2019, from 3.5 to 8770 kg ha
−1 during 2019–2020, and from 30 to 5675 kg ha
−1 during 2020–2021.
We did find evidence of species-level differences in fractional vegetative cover at both low, medium, and high biomass levels (
Figure 3). However, we did not have sufficient observations to conduct a formal analysis to determine whether these differences were statistically significant. At low biomass levels (~1000 kg ha
−1), triticale appears to have a higher fractional vegetative cover than cereal rye and wheat. As no cereal rye biomass levels exceeded 4000 kg ha
−1, we were not able to compare the highest biomass ranges between all three species. However, our in situ photographic data indicated that cereal rye was patchier than wheat and triticale at all biomass levels, with the caveat that we obtained limited cereal rye photographs and a gap of >7 days from photographs and biomass harvests in 2019–2020.
We also examined the spectral response from Sentinel-2 for each species at a low and high biomass (
Figure 4). As anticipated, we observed a greater NIR reflectance and decreased red reflectance at high biomass (>3000 kg ha
−1) compared to low biomass (<1000 kg ha
−1), which is, in part, related to the exposed soil present at low ground cover. Similarly, the reflectance of the first red-edge band (B5) decreased, while the second and third red-edge bands (B6 and B7) increased reflectance with increasing cover crop biomass levels. Interestingly, in the SWIR portion of the spectrum, reflectance from low biomass exceeded thoseof high biomass for all species, but the magnitude of this difference was smallest in cereal rye.
Finally, we examined the relationship between handheld soil moisture collections and SAR metrics. The results showed unexpectedly poor relationships (R
2 = 0–0.08) between soil moisture and backscatter intensities, polarimetric parameters, and InSAR coherence (
Appendix A). Hence, we did not further employ remote sensing estimates of soil moisture in our analyses due to the high uncertainty in calibration.
3.1. Optical Index Results
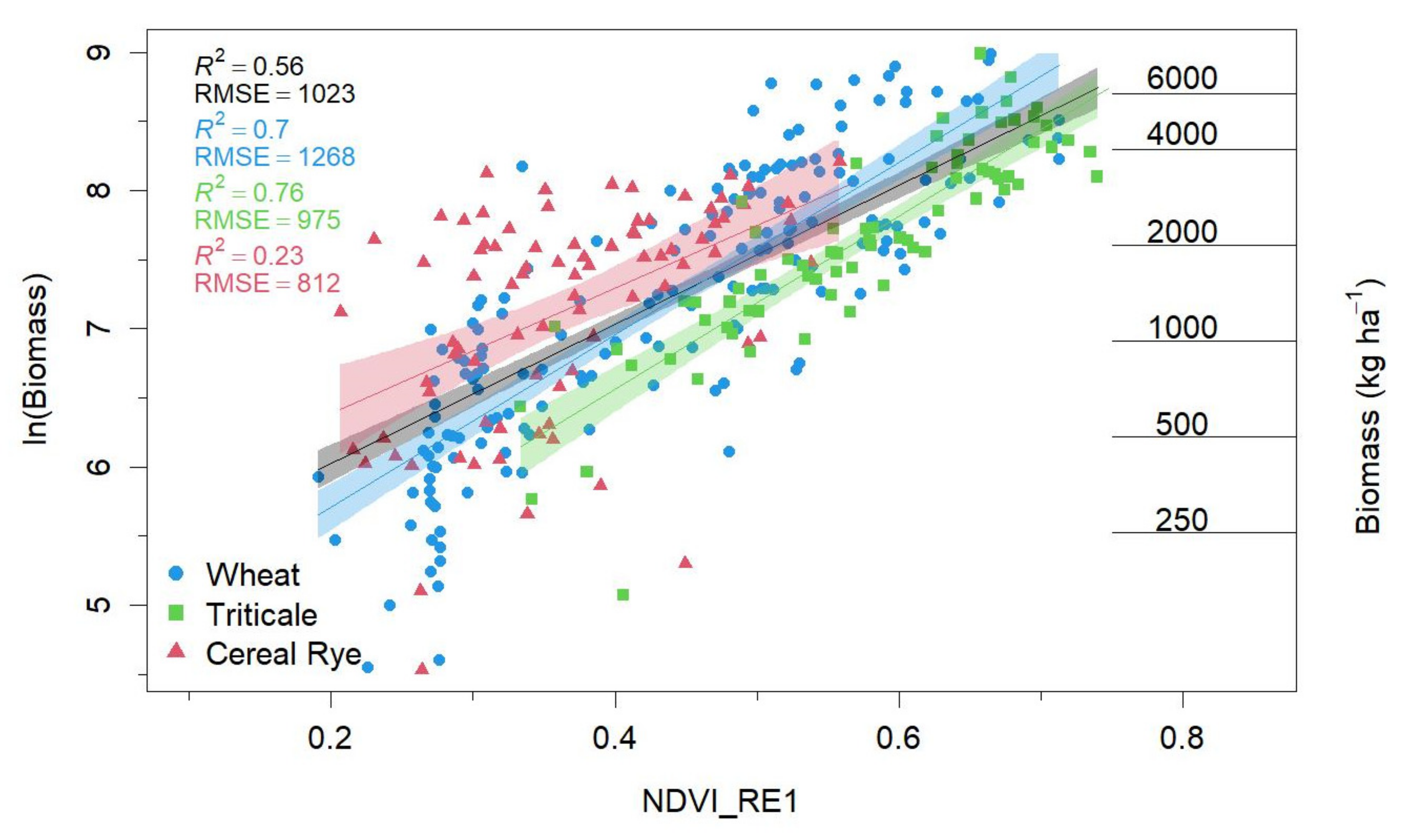
Log-linear models best met regression assumptions and performed similarly to polynomial and segmented models. Thus, we only present results from log-linear models in the main text. Among all the considered VIs, NDVI_RE1 proved to be best at predicting cover crop biomass (R
2 = 0.61; RMSE = 1019; MAE = 639; AICw = 0.98;
Table 3;
Figure 5). Although NDRE and NDWI1 had similar coefficients of determination and error values (
Table 3), their AICc were ≥ +8 when compared to NDVI_RE1. Our findings were further supported by AICw values, where we found a 98% probability that the model using NDVI_RE1 to predict cover crop biomass was the best when compared to the other VIs included in our evaluation.
After identifying NDVI_RE1 as most suitable for estimating cover crop biomass, we evaluated whether seasonal- or species-level differences were present using ANOVAs and slope and intercept contrasts. We found that significant effects (i.e., a relationship between NDVI_RE1 vs. biomass) varied between winter (October–February) and spring (March–May) samples (
Appendix B;
Table A1). Therefore, we opted to move forward with modeling the relationship between cover crop biomass and NDVI_RE1 with only spring data (
n = 332;
Figure 5), as there were only two observations in winter that exceeded 1500 kg ha
−1, which is the existing saturation point identified for cereal grass cover crops [
14]. Furthermore, we found species-level differences between cereal rye, triticale, and wheat (
Appendix B;
Table A2). In our study, cereal rye had lower observed biomass ranges than triticale and wheat. Therefore, we further examined whether these species-level differences remained when we limited our analysis to similar biomass ranges for these three cereal grass species. Our results suggest that species-level differences were still present within this sub-analysis (results not shown). Therefore, as we found significant differences between species (primarily between cereal rye and the other two species) in our ANOVAs and slope-intercept contrasts, we also modeled species separately (
Figure 5). With these season*species interactions, the results of our optical index selection (NDVI_RE1) showed substantial increases in goodness-of-fit wheat (R
2 = 0.70) and triticale (R
2 = 0.76). However, we observed that the relationship between NDVI_RE1 and cereal rye biomass performed poorly (R
2 = 0.23).
3.2. Optical—Synthetic Aperture Radar (SAR) Results
Results from objective 1 indicated that NDVI_RE1 was our best-performing optical index, but also that seasonal and species level effects were present in our data. Therefore, our analyses integrating optical and SAR data focused solely on spring samples (n = 332) and modeled species separately in objective 2. Each SAR metric (backscatter intensities and indices, polarimetric parameters, and InSAR coherence) was individually added to NDVI_RE1 regression models to predict cover crop biomass and assess how model statistics changed with each additional SAR metric.
The addition of most SAR metrics did not substantially improve model fits or reduce errors compared to models that had only NDVI_RE1 (
Table 4) but were species-specific. However, combining InSAR coherence and NDVI_RE1 did increase our ability to predict the biomass of all three cereal grass species (
Table 4), with improved model fits (i.e., R
2 increased by 4 to 11% and AICc decreased by 15 to 24 among species). Wheat, triticale, and cereal rye models with InSAR coherence also demonstrated reductions in error that ranged from small to moderate: RMSE decreased by 42, 182, and 36 kg ha
−1 and MAE decreased by 95, 84, and 52 kg ha
−1, respectively.
Models including both NDVI_RE1 and InSAR coherence were significantly better than the model using NDVI_RE1 alone in all three species (F = 15.75–26.86,
p < 0.001;
Table 5). Similarly, models with both NDVI_RE1 and InSAR coherence demonstrated an increased predictive ability in terms of cross-validation compared to models with just NDVI_RE1 (
R
2 = +3–9%;
Table 6). We also observed small-to-moderate reductions in errors in wheat, triticale, and rye cross-validation (
RMSE = 37, 188, and 23 kg ha
−1;
MAE = 74, 92, and 37 kg ha
−1, respectively).
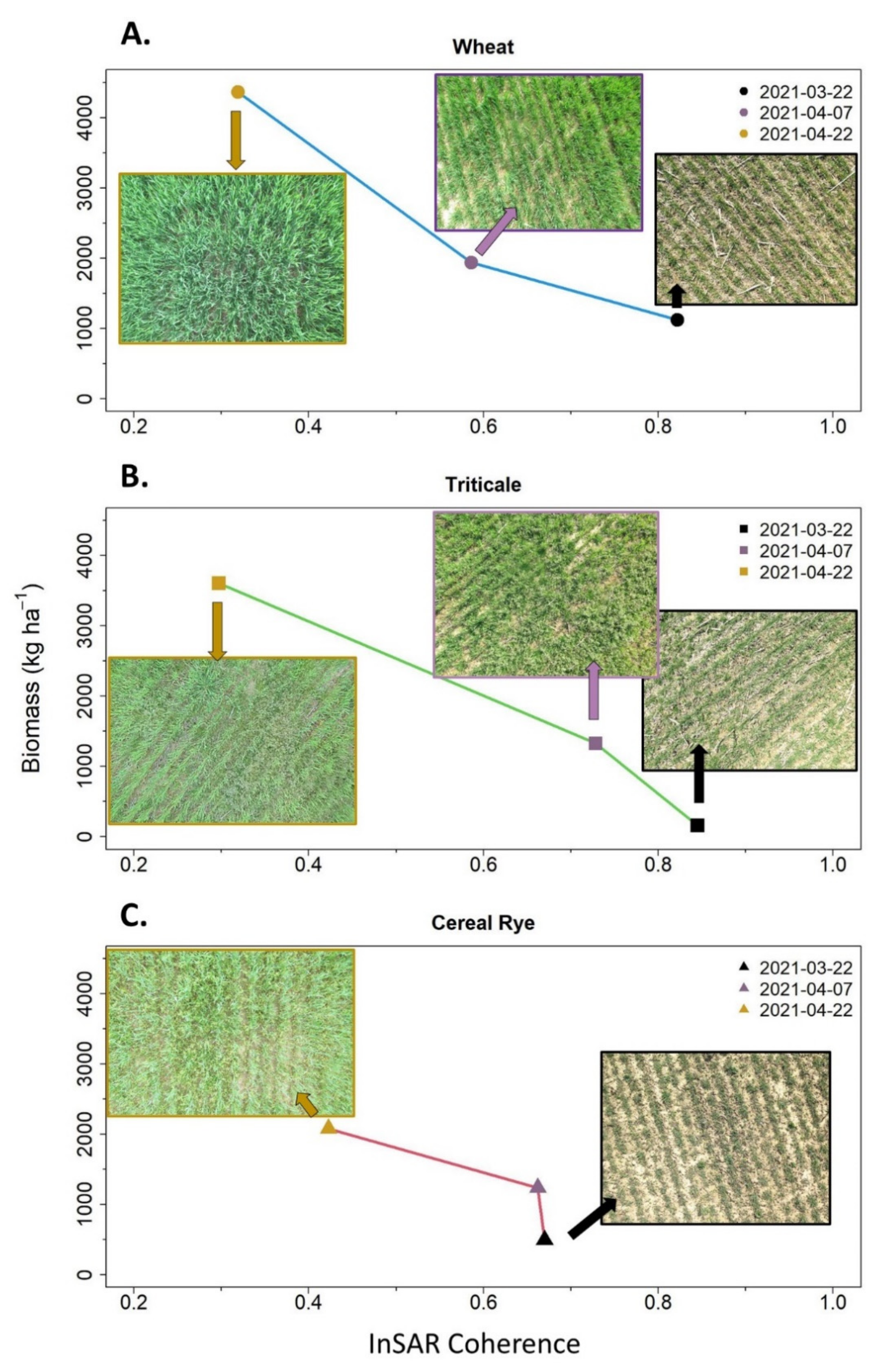
We observed that, as biomass increased, InSAR coherence decreased, a pattern that results from increased volume scattering producing return signals with a random phase (
Figure 6). However, in 47 out of 573 datapoints (i.e., two April sampling dates: cereal rye = 0, triticale = 26, wheat = 21), cover crop termination events occurred between the acquisition of the InSAR image pairs. Termination events may result in additional InSAR coherence decreases as they significantly alter the landscape structure between images in the InSAR pair. For this reason, we performed a sensitivity analysis to assess whether model statistics changed between models with or without including these 47 observations and found no substantial differences (
Appendix C).
As the addition of multiple SAR metrics produced statistically improved biomass predictions compared to models with NDVI_RE1 alone (
Table 5), we also explored the use of multiple SAR parameters in addition to NDVI_RE1 for each of the three cover crop species, which we refer to as “combined models.” All combined models were screened for collinearity and no predictor variables correlated above the predefined threshold (r > 0.7). For wheat and triticale, the best-performing combined model contained NDVI_RE1, InSAR coherence, and entropy (wheat) or anisotropy (triticale) as predictor variables based on a combination of R
2, error measures, AICc and AICw (
Table 3 and
Table 5). In contrast, the best-performing model for the prediction of cereal rye biomass included NDVI_RE1, InSAR coherence, as well as VV and RVI derived from backscatter intensities (
R
2 = +15%;
Table 4). Despite these gains in SAR metrics in the cereal rye models, they remained the poorest-performing models compared to wheat and triticale. Combined models for each of the three species were also statistically more significant than models with NDVI_RE1 alone (
Table 5), but not when compared to models that included only InSAR coherence and NDVI_RE1 except for cereal rye (not shown). Similarly, we found no substantial changes between combined models and those with NDVI_RE1 and InSAR coherence alone during cross-validation (
Table 6).


 ,
, 



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}