Abstract
Determining the driving climatic factors at critical periods and potential legacy effects is crucial for grassland productivity predictions on the Qinghai–Tibet Plateau (QTP). However, studies with limited and ex situ ground samples from highly heterogeneous alpine meadows brought great uncertainties. This study determined the key climatic factors at critical plant developmental stages and the impact of previous plant growth status for interannual aboveground net primary productivity (ANPP) variations in different QTP grassland types. We hypothesize that the impact of climatic factors on grassland productivity varies in different periods and different vegetation types, while its legacy effects are not great. Pixel-based partial least squares regression was used to associate interannual ANPP with precipitation and air temperature at different developmental stages and prior-year ANPP from 2000 to 2019 using remote sensing techniques. Results indicated different findings from previous studies. Precipitation at the reproductive stage (July–August) was the most prominent controlling factor for ANPP which was also significantly affected by precipitation and temperature at the withering (September–October) and dormant stage (November–February), respectively. The influence of precipitation was more significant in alpine meadows than in alpine steppes, while the differentiated responses to climatic factors were attributed to differences in water consumption at different developmental stages induced by leaf area changes, bud sprouting, growth, and protection from frost damage. The prior-year ANPP showed a non-significant impact on ANPP of current year, except for alpine steppes, and this impact was much less than that of current-year climatic factors, which may be attributed to the reduced annual ANPP variations related to the inter-annual carbon circulation of alpine perennial herbaceous plants and diverse root/shoot ratios in different vegetation types. These findings can assist in improving the interannual ANPP predictions on the QTP under global climate change.
1. Introduction
Global climate change is altering existing climatic conditions, including dramatic changes in temperature and precipitation affecting terrestrial ecosystem processes [1,2,3]. Alpine grasslands, one of the most vulnerable ecosystems, are extremely sensitive to climate change and have complex feedback processes with the environment [4,5]. Alterations in alpine grassland productivity driven by climate change affect forage resource sustainability, pastoral livelihood profitability and the global carbon balance [6]. However, spatiotemporal variations in alpine grassland productivity and its interactions with global climate change are not well understood [6,7]. Accordingly, clarifying the sensitive climatic factors affecting grasslands and the corresponding periods can assist in predicting the growth responses of alpine plants under global climate change, leading to more rational and effective pasture management recommendations for decision-makers [4,8].
The current understanding of the relationship between climatic factors and grassland productivity based on field observation is derived from temporal models using long-term datasets from one location, or spatial models using datasets that describe spatial patterns across a climatic gradient [9,10]. These two approaches seem to provide substantially different predictions of ecological responses to future environmental changes [9,11,12]. Spatial models reflect the chronic ecological responses of plants related to species composition and their corresponding traits to long-term changes in climatic conditions [9,13]. Temporal models predict changes in plant growth within communities with altered climatic factors and indicate relatively short-term ecological responses to climatic variations related to plant density, leaf size, and stem thickness per plant [9,10]. Compared to spatial models, temporal models can better predict relatively short-term forage yield variations for livestock production. Therefore, exploring the responses of alpine grassland aboveground net primary productivity (ANPP) to climatic variables using a temporal model is necessary.
Alpine grasslands on the Qinghai–Tibet Plateau (QTP), known as the global “third pole” [14], are extremely sensitive to climate change [15]. Under the dual pressures of global climate change and rapid population increase, alpine grasslands of the QTP have been severely degraded, greatly affecting the animal husbandry industry and herder’s livelihoods [16]. To predict grassland productivity and arrange pastoral production, many studies on vegetation productivity–climate relationships from a temporal perspective have been conducted based on current-year climatic factors [15,17,18,19] and possible potential lagged or legacy effects [20,21].
The impact of the current-year climatic factors on grassland productivity on the QTP has been studied from annual and growing season [17,18,19] to seasonal and monthly scales [15,22,23]. Plant resource requirements vary in different vegetation types and plant developmental stages owing to different life activities [24,25,26]. Thus, ANPP variations may be more relevant to climatic factors during periods pertinent to the specific phenology or life history of plants in an ecosystem [27,28]. More studies at seasonal and monthly scales have demonstrated that temperature was the dominant climatic factor for alpine meadows, shrubs in spring, summer, and fall (Table S1) [19,22,29]. For alpine steppes, vegetation growth was affected by both precipitation and temperature, but climatic factors were more prominent in summer than in spring and fall (Table S1) [15,19,29]. However, the seasonal time division is inconsistent with that of the developmental stage.
Recently, a developmental stage-based study of ANPP and climatic factors in alpine meadows was conducted by Li et al. [4] with long-term in situ observations. However, limited vegetation types and ex situ ground samples under the high heterogeneity of grasslands may induce great uncertainties in determining the responses of interannual ANPP variations to climatic changes. Satellite-derived vegetation indices may be ideal surrogates for field-harvested biomass/ANPP in environmentally harsh or large areas [30,31]. Therefore, it is necessary to determine the vegetation productivity–climate relationship of different vegetation types at the developmental stage scale based on remote sensing images on the QTP. In this study, we hypothesized that the impact of climatic factors on plant growth varies at different developmental stages and different vegetation types have diverse responses (Hypothesis 1).
Prior-year or earlier climatic conditions can significantly affect plant growth (lagged or legacy effects) and are necessary for the prediction of grassland ANPP [32,33,34]. However, relevant studies on the QTP have focused on the impact of relatively short-term (several months) legacy effects within the current year, and a 0–2-month lag has been detected in the vegetation productivity–climate relationship [20,21,35]. Therefore, confirming the impact of long-term (at least 1 year) legacy effects on grassland ANPP is also significant for the QTP.
The currently proposed legacy effects can be divided into two categories: biotic and abiotic legacies [36]. Abiotic legacies indicate that prior-year climatic conditions may influence current-year ANPP through soil moisture carryover [37,38,39,40], or by modifying soil nutrient availability [41,42,43]. Studies showed that precipitation can contribute the legacy impact on plants if their rooting depth allows them to access the stored water from earlier wet years [44]. Precipitation in the previous year may also influence the inorganic nitrogen pool by affecting the microbial mineralization, immobilization, and nitrogen leaching, further promoting or suppressing vegetation growth [41]. Biotic legacies can be mediated by changes in individual plants or shifts in community structure [45,46,47]. Such individual plant changes include tiller [48], stolon [49], and meristem [25] dynamics. Variations in tiller and stolon density regulate the vegetation productivity [48,50]. However, long-lasting (at least 1 year) abiotic legacies hardly exist on the QTP because the soil wetting duration usually lasts less than 2 days even after heavy rain on the QTP [51]. In addition, soil inorganic nitrogen availability is affected by rainfall events and only lasts for a short phase (less than 3 months) [52].
Accordingly, the long-lasting legacy in plant growth responses to climate change on the QTP, if any, is more likely to be a biotic legacy. Biotic legacies generally shape the current-year ANPP by affecting the plant growth status of previous year through tiller, stolon, and meristem dynamics [25,48,50]. However, the inter-annual carbon circulation of plants may be more pronounced in the prior-year climatic legacy effect on the current-year ANPP variations under harsh environments. Plant interannual carbon allocation is a widespread strategy that allows perennial herbaceous plants to seasonally accumulate non-structural carbohydrates (NSCs) in underground organs before the onset of cold or dry periods, and to transfer NSCs to aboveground organs the following year, serving as a vital resource for life activities in the following year [53,54,55]. This strategy can guarantee that plants complete their life circle in harsh environments and is particularly significant for the development of alpine plants [56], and it may also reduce the interannual ANPP fluctuation in alpine grasslands and decrease the impact of prior-year legacy from climatic factors. In addition, the root/shoot (R/S) ratios of different grassland types varied enormously with different vegetation types [57,58,59,60,61]. Species with higher R/S ratios have developed root systems to store more NSCs in winter to alleviate the effects of legacies. Consequently, we also hypothesized that alpine grasslands may not indicate a great prior-year legacy impact, and vegetation types dominated by species with higher R/S ratios have lower legacy effects and vice versa (Hypothesis 2). In this study, we tested the abovementioned two hypotheses and answered the following two questions: (i) What are the main climatic factors and critical developmental stages driving ANPP for different vegetation types in the QTP grassland ecosystem; (ii) Is ANPP of different vegetation types on the QTP affected by the antecedent plant growth status, and if it is, how much are the impacts?
2. Materials and Methods
2.1. Study Area
The study area is located in the Three-River Headwaters (TRH) region (31°39′–37°17′N, 89°45′–102°23′E), referring to the source area of the Yangtze, Yellow, and Lancang Rivers. It is also called the “Chinese Water Tower”, with a total area of approximately 3.95 × 105 km2 (Figure 1). This region is dominated by mountain and canyon landforms, and the overall terrain gradually rises from southeast to northwest, with altitudes ranging from 3200 to 4700 m [62]. The annual precipitation ranges from 262 to 723 mm, and the annual average temperature ranges from −5.6 to 3.8 °C, from the northwest to southeast TRH region, with climatic zones changing from semiarid (dominant climatic zone) to humid subtropical [63,64]. The vegetation types in the TRH region are primarily grasslands (approximately 68%), including alpine meadows and steppes which predominately consist of alpine steppes, except a small area of temperate steppes. Alpine meadows are the most widely distributed and largest natural grassland in the TRH region, accounting for approximately 50% of the total grassland area. They can be divided into typical alpine meadow and alpine swamp meadow, which are dominated by Kobresia, with some mixed forbs and grasses.

Figure 1.
Study area, including the Three-River Headwaters region: (a) location in relation to China, and (b) vegetation types.
2.2. Data Collection and Preprocessing
In this study, the Terra Moderate-Resolution Imaging Spectroradiometer (MODIS) normalized difference vegetation index (NDVI, MOD13A1) was used for indicating ANPP between 2000 and 2019, with a spatial resolution of 500 m and a temporal resolution of 16 days. The dataset was confirmed to be of high quality and suitable for expressing vegetation dynamics [30,65]. The NDVI dataset was obtained from Google Earth Engine (http://earthengine.google.com, accessed on 20 March 2021) [66]. Monthly NDVI values were calculated using the maximum value composites method, which further eliminated errors caused by clouds, atmosphere, sun angle, and other interferences [67]. NDVI cannot indicate vegetation status accurately under low vegetation coverage with low greenness due to the impact of soil background [68]. Thus, we excluded pixels with low NDVI value for analysis to reduce uncertainties. We selected 0.1 as threshold of low NDVI according to the suggestion from Piao et al. [69,70].
The meteorological dataset included the monthly average temperature and monthly total precipitation from January 1982 to December 2019 at a spatial resolution of 1 km, and was downloaded from the National Earth System Science Data Center (https://gre.geodata.cn/, accessed on 20 March 2021). These meteorological data were downscaled in China using the Delta spatial downscaling scheme and based on the global 0.5° climate dataset published by the Climatic Research Unit and the global high-resolution climate dataset published by WorldClim [71]. The resulting dataset was verified to be credible by 496 independent meteorological observation stations [71,72].
Vegetation types and their distributions were obtained from a 1:1,000,000 digitalized vegetation map of China [73]. The alpine grasslands in the TRH region were extracted from the vegetation map, including the typical alpine meadow (TAM), alpine swamp meadow (ASM), alpine steppes (AST), and alpine shrubs (ASH). Other vegetation types were not considered because of their small proportions. Finally, we obtained an approximate distribution of the five vegetation types in the study area (Figure 1).
2.3. Methods
2.3.1. Developmental Stages of Grasslands on QTP
In addition to the annual and growing season precipitation/temperature, this study also concentrated on climatic factors at plant developmental stages. Considering the plant germination time and months that do not influence the estimated annual ANPP, the developmental stages of alpine grassland were determined based on a plant-centric year instead of the calendar year. The measured annual ANPP of grasslands on the QTP is commonly determined by approximating the maximum aboveground biomass during the growing season [74,75]. In a TRH ecosystem with a spring–summer growing season, the maximum biomass typically occurs in mid–late August [76,77,78]. Most herbaceous species on the QTP begin the germination after the maximum biomass is reached, and then enter the growth stage after experiencing winter dormancy [62,79,80]. Precipitation or temperature that occurs after the maximum biomass is reached should not influence the estimated annual ANPP. Therefore, this study took a plant-centric year from September to August instead of January to December of the calendar year (Figure 2).
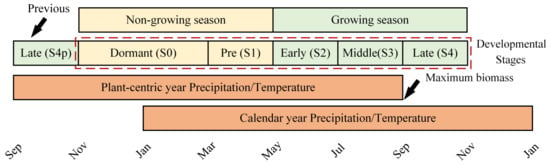
Figure 2.
Plant-centric year was defined starting from September to the end of August, and the major growing season was from early May to October. The dotted line represents the complete developmental cycle divided into five stages, including the dormant (S0) and pre- (S1) to late- (S4) growing season stages [27]. However, we used the S4p instead of S4 for a complete plant-centric year because the developmental process of most herbaceous species begins with germination after the maximum biomass occurring in mid–late August.
We defined the non-growing and growing seasons as November to April and May to October, respectively [4]. The non-growing season was sub-divided into two periods: the dormant stage (S0, from November to February) and the pre-growing season stage (S1, two months from the end of S0 to the end of the non-growing season). The growing season was divided into three equal stages, following the method by Robinson et al. [27], each of which lasted for two months: the early-growing season, also called the vegetative stage (S2); the mid-growing season, also known as the reproductive stage (S3); and the late-growing season, also known as the withering stage (S4). Considering the plant germination after the maximum biomass occurred from mid–late August, we used a previous withering stage (S4p) instead of S4 to fill the plant-centric year (Figure 2).
2.3.2. Indices of Input Variables
Grassland biomass is commonly obtained through field harvesting, and annual ANPP can be estimated using aboveground biomass (AGB). However, intense field sampling and sample processing [81,82] incur high costs for large spatial areas [83], and it is particularly difficult to collect samples from the QTP under harsh natural conditions. In addition, studies conducted with limited and ex situ samples in highly heterogeneous grassland may induce great uncertainties in determining the responses of ANPP to climatic changes. Thus, vegetation indices, indicating a close relationship with AGB or annual ANPP, have commonly been used as ideal surrogates of AGB or ANPP from field-harvested biomass in environmentally harsh or large areas [30,31,84]. Accordingly, we used NDVI at the peak of the growing season as a proxy for the net photosynthetic activity to determine the impacts of climatic changes on grassland annual ANPP, which was considered as a suitable approximation for these ecosystems [74,75].
The total precipitation and mean air temperature during a study period, including annual, growing season, and developmental stage scales were used as the climatic factor indices. The annual period was from September to August, and the growing season was from May to October. The developmental stages used in this study (S4p–S3), as discussed in Section 2.3.1, were from September to October, November to February, March to April, May to June, and July to August, respectively (Figure 2). The precipitation and temperature in the five developmental stages are abbreviated to P4p–P3 and T4p–T3, respectively.
To study the effects of the prior-year plant growth status on that of the current year, the maximum NDVI of the previous year (Np), representing the ANPP of that year, was added as an input variable. The long-lasting legacy in the responses of plant growth to climate change on the QTP is more likely to be a biotic legacy. However, prior-year vegetation growth status may be affected by precipitation, temperature, and radiation, etc. from the previous year, and it may also influence the bud growth and ability to resist freezing damage, thereby affecting grassland productivity next year. Thus, linking the prior-year climatic factors to the current-year vegetation productivity introduces large uncertainties owing to complex impact paths (prior-year climatic factors → prior-year plant growth status → current year grassland ANPP). The antecedent vegetation growth status is easy to obtain with the advancement of remote sensing technology. Consequently, directly considering the prior-year plant growth status as an impact factor instead of the antecedent climatic factors to detect the ANPP drivers may be a better option [85,86].
2.3.3. Impact of Driving Factors on ANPP
The impact of driving factors on ANPP was evaluated by partial least squares (PLS) regression, which integrates the strengths of multiple linear regression, principal component analysis and canonical correlation analysis [87]. Unlike many other regression methods, PLS can be used effectively when the number of observations is close to or lower than that of the independent variables; these variables are also highly correlated [88,89].
Standardized model coefficients (SMC) and variable importance in projection (VIP) are the two main outputs in PLS regression [90]. SMC values represent the direction and strength of each variable impact in PLS regression [88]. VIP values indicate the importance of independent variables that explain the variations of dependent variables based on the weighted sum of squares of PLS loadings [88,90]. For interpretation purposes, only predictors with a VIP of more than 0.8 are considered important [91]. Centering and scaling of the dependent and independent variables were also conducted to allow comparisons between different variables [90].
The independent variables included the total precipitation, mean temperature at five developmental stages, and the prior year maximum NDVI; the dependent variable was the current year maximum NDVI. The correct number of components for the PLS model was selected based on the root mean squared error (RMSE) of the PLS model [87]. In the PLS regression, variables with a VIP > 0.8 and large absolute SMC values represent the climatic factors in relevant phases or Np significantly influencing vegetation ANPP. Positive SMC values indicate that increasing prior-year NDVI or current-year climatic factors in the relevant period should promote ANPP, whereas negative SMC values imply negative effects on grassland productivity [88].
To determine the impact of driving factors on the ANPP of a grassland type or the entire area, SMC and VIP values in each pixel were calculated with PLS regression and then we averaged all pixels within a given extent. Predictors with an average VIP of more than 0.8 were considered important.
3. Results
3.1. Impact of Current-Year Climatic Factors
Grassland ANPP was significantly affected by precipitation at the annual and growing season scales, but not in response to air temperature. The SMC values of the growing season and annual precipitation in the whole region were 0.316 and 0.313, clearly higher than the 0.105 (VIP = 0.726) and 0.079 (VIP = 0.696) of the temperature in the growing season and plant-centric year, respectively (Figure 3). The VIP values of the growing season/annual precipitation were >0.8, whereas those of temperature were <0.8 for the entire area and four vegetation types (Figure 4). This indicated that precipitation, rather than temperature, was the main driving factor of ANPP variations on the QTP.
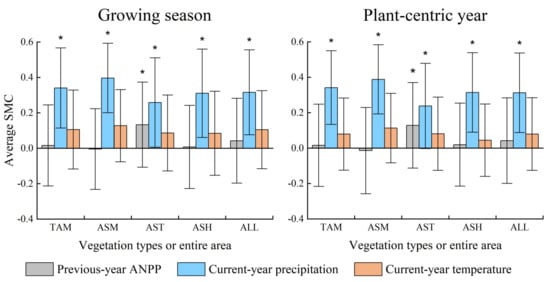
Figure 3.
Responses of aboveground net primary productivity (ANPP) to antecedent plant growth status and climatic factors for different vegetation types and the entire area at the growing season and annual scales. ‘TAM’, ‘ASM’, ‘AST’, ‘ASH’, and ‘ALL’ represents typical alpine meadow, alpine swamp meadow, alpine steppes, alpine shrubs, and the entire area; ‘*’ represents average VIP > 0.8; error bar represents the standard deviation of SMCs for the entire area or a vegetation type.
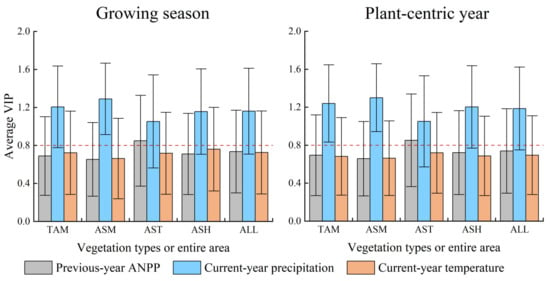
Figure 4.
Average VIP values of PLS regression in the responses of ANPP to antecedent plant growth status and climatic factors for different vegetation types and the entire area at the growing season and annual scales. The error bar represents the standard deviation of VIPs for the entire area or a vegetation type. ‘TAM’ to ‘ALL’ were the same as that in Figure 3.
ANPP variations showed divergent responses to climatic factors at different developmental stages. The SMC value of P3 was 0.284 (VIP = 1.441) in the whole region, 6.03 times the average coefficient for all the impact factors. For different vegetation types, the SMC value was 6.77, 7.98, 4.33, and 6.38 times its average coefficient of all impact factors, from TAM, ASM, AST, to ASH, respectively (Figure 5). P4p and T0 also affected ANPP significantly, but were much less than P3. The SMC value of P4p was 0.087 (VIP = 0.836) in the entire area, and 0.070 (VIP = 0.808), 0.086 (VIP = 0.811), 0.124 (VIP = 0.893), and 0.054 (VIP = 0.806), respectively, for TAM, ASM, AST, and ASH; the SMC value of T0 was 0.054 (VIP = 0.837) in the entire area, and 0.065 (VIP = 0.851), 0.039 (VIP = 0.861), 0.067 (VIP = 0.837), and 0.066 (VIP = 0.816), respectively, for TAM, ASM, AST, and ASH (Figure 5). In other developmental stages, ANPP did not show a consistently pronounced response to climatic factors for different vegetation types. This indicated that the critical climatic factors for alpine grassland were P3, P4p, and T0, with the corresponding key developmental stages of S3, S4p, and S0, respectively; P3 was the most prominent controlling factor.

Figure 5.
Responses of ANPP to antecedent plant growth status and climatic factors for different vegetation types and the entire area at the developmental stage scale. ‘*’ represents average VIP > 0.8, while the error bar represents the standard deviation of SMCs for the entire area or a vegetation type. S4p to S3 are five plant developmental stages in a plant-centric year, representing the withering, dormant, pre-growing season, vegetative, and reproductive stage, respectively. ‘Np’ indicates previous-year annual ANPP.
ANPP variations showed differentiated responses to the dominant climatic factor for different vegetation types. ANPP variations for ASM exerted the strongest response to growing season/annual precipitation with SMC of 0.397/0.388, followed by TAM and ASH, with an SMC of 0.341/0.342 and 0.311/0.314, respectively, and AST with an SMC of 0.259/0.238 (Figure 3). The impact of P3 was also more pronounced in alpine meadows (TAM and ASM) than in alpine steppes, and this impact in ASM was higher than in TAM. This indicated that the influence of precipitation on interannual ANPP was more significant in alpine meadows than in alpine steppes on the QTP.
3.2. Impact of Prior-Year Plant Growth Status
The effect of prior-year ANPP on current-year ANPP was not significant. The VIP value of Np at the growing season and annual scales in the entire area was 0.736 and 0.740, respectively, all <0.8 (Figure 4), indicating a non-significant effect on ANPP variations. The VIP value of Np at the developmental stage scale was 0.822, while the corresponding SMC value was 0.017, only 6% of P3 (Figure 5). This implied that the prior-year legacy impact on current-year ANPP was limited in alpine grasslands on the QTP.
The effect of prior-year ANPP on current-year ANPP varied with vegetation types. The SMC of Np was 0.133 (VIP = 0.850), 0.129 (VIP = 0.852), and 0.079 (VIP = 0.944) for the AST at annual, growing season, and developmental stage scales, respectively (Figure 3 and Figure 5). In the other three vegetation types, SMCs of Np ranged from −0.02 to 0.02, and VIPs were <0.8, with no significant effects on grassland ANPP variations (Figure 4 and Figure 6). This implied that the effect of antecedent plant growth status on ANPP was non-significant, except for alpine steppes.
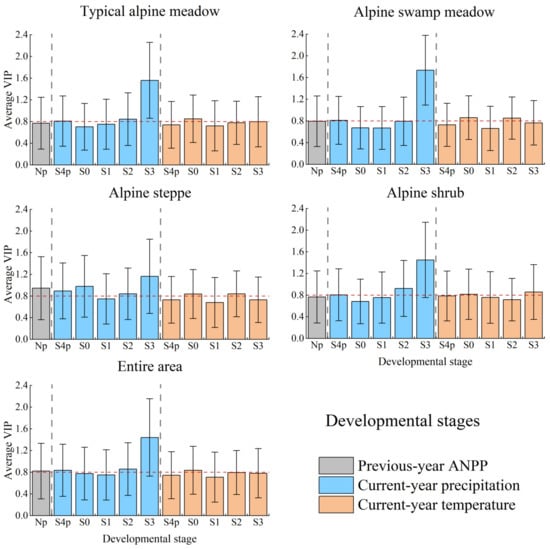
Figure 6.
Average VIP values of PLS regression in the responses of ANPP to antecedent plant growth status and climatic factors for different vegetation types and the entire area at the developmental stage scale. The error bar represents the standard deviation of VIPs for the entire area or a vegetation type. S4p to S3 and ‘Np’ were the same as that in Figure 5.
4. Discussion
4.1. Comparison with Previous Studies
4.1.1. Comparison with Results of Previous Studies on QTP
More studies had indicated that temperature was the dominant climatic factor for vegetation growth on the QTP (Table S1) [17,92,93,94]. However, our results showed that the responses of ANPP variations to precipitation rather than temperature were more pronounced whether in the entire area or in different vegetation types (Figure 3 and Figure 4), which supported the findings of a small number of studies [18,20,95,96]. These results were consistent with the findings of previous controlled experiments on the QTP [74,97,98,99,100]. A global meta-analysis also indicated that cold ecosystems were more responsive to precipitation variations than other ecosystems [74,101].
Our results also showed that the S3 stage precipitation (P3) was the main controlling factor, and the S4p stage precipitation (P4p) and the S0 stage temperature (T0) also significantly affected vegetation productivity, which was inconsistent with the findings of Li et al. [4], in which the annual ANPP variations were determined by the onset and end of the growing season temperature (T1 and T4p). However, the limited and ex situ ground samples under the high heterogeneity of grasslands may induce uncertainties in uncovering the vegetation–climate relationship and the ground samples in this study were only collected in alpine meadows. Our results also showed a diverse response of inter-annual ANPP to climatic factors in different vegetation types. The influence of P3 was more significant in alpine meadows (TAM and ASM) than in alpine steppes, and its influence in ASM was higher than in TAM (Figure 5 and Figure 6).
Research on the legacy effects of climate on grassland productivity in the QTP ecosystem has been scant to date [102]. This study showed that the impact of prior year plant growth status on ANPP was non-significant except for the alpine steppes, even though the significant impact of the alpine steppes was much less than that of current-year climatic factors. This difference in the legacy effect between alpine meadows and steppes is consistent with reports from Li et al. [102] on the QTP along an elevation gradient.
4.1.2. Possible Reasons for Discrepancies from Other Studies
First, the differences in the surrogates were used as vegetation productivity. In this study, the annual maximum NDVI was used as a surrogate of annual ANPP [4,17,70]. However, other studies often used the mean or cumulative value of NDVI/biomass within one period (month, season, growing season, or year) as a surrogate of vegetation productivity, but these indices should be an approximation of the average biomass state over the study period [15,17,18]. Therefore, these studies may not reflect the relationships between annual vegetation ANPP and interannual climatic factors during different periods.
Second, different study phases were chosen. Studies have shown inter-decadal variation in the responses of vegetation productivity to interannual variations in precipitation or temperature, and such responses are strongly affected by the interactions between these two climatic factors [19]. Zhang et al. [94] suggested that changes in climatic conditions before and after 2000 had directly resulted in the restricting factors of plant growth being converted from temperature and radiation to precipitation in the TRH region.
Third, differences in the NDVI products were identified. There are two NDVI products that were mainly used in the remote sensing-based research on the vegetation–climate relationship. One is MODIS NDVI, and the other is the advanced very high-resolution radiometer (AVHRR) NDVI. However, discrepancies have been found between these two NDVI datasets in some arid and Arctic areas with sparse herbaceous and shrub covers [103]. Recent research has revealed an opposite trend of vegetation greenness on the QTP since the beginning of 21st century based on the two NDVI products [104]. This was probably because the AVHRR sensor suffered from some well-known shortcomings, such as orbit drift in the satellite overpass time and post-launch degradation in sensor calibrations [105]. Problems with AVHRR satellite imagery with coarse spatial resolution have been reported in many previous studies on the QTP [7,106], which failed to reveal the detailed growth curves of alpine grasslands.
4.2. Low-Temperature Acclimation in the Alpine Ecosystem May Introduce a Pronounced Impact of Precipitation on ANPP Rather than Temperature
Lower temperatures are not the limiting factor in the interannual variations of alpine grassland productivity because of physiological thermal acclimation in alpine ecosystems [56,107,108]. Thermal acclimation to low temperatures primarily results from changes in the photosynthetic apparatus and is associated with electron transport in the thylakoid membrane, particularly photosystem II [109,110]. The optimum temperature for photosynthesis correlates with active quantum flux density (QFD) so that the optimum is at low/high temperatures when QFD is low/high [56]. Thus, the photosynthetic temperature response curve is so wide [111,112] that photosynthesis can operate at 95% of the maximum rate over a range of 8 °C [56]. At the same time, re-adjustment from the optimum temperatures to the prevailing temperatures is very fast (only a few days) [113,114]. Therefore, changes in vegetation ANPP did not demonstrate a significant correlation with annual or growing season air temperatures (Figure 3).
Our temporal model-based results indicate that alpine plant growth is significantly affected by precipitation (Figure 3). Effective control of alpine plants to potential moisture shortages can be conducted through the reduction of leaf area (or increase in root production) per plant and/or the reduction of coverage per unit of land area in the short term, and replacement of species from less to more water-condition-adapted ones in the long term [56]. In addition, mineral nutrients are highly correlated with soil water content which mainly comes from precipitation. Lower soil water content may reduce nutrient supply for alpine plants by limiting nutrient cycling and microbial activity [56,115]. Temporal models of vegetation productivity–climate relationships depict a relatively short-term impact of water conditions on plant growth [9]. Thus, to reduce the physiological water restriction, the adaptive community responses of alpine plants to potential moisture shortage will operate via low ground cover and high R/S ratios. Consequently, the responses of alpine plant productivity to precipitation were more significant than those to temperature (Figure 3).
4.3. Possible Mechanisms for ANPP Responses to Climatic Factors at the Critical Developmental Stage
4.3.1. More Water Consumption from the Largest Plant Leaf Area May Lead to the Most Pronounced Impacts of S3 Stage Precipitation
The vegetation–precipitation relationship is discussed in Section 4.2. The short-term regulation of alpine plants to moisture is achieved by changing the leaf area or coverage (e.g., LAI) and R/S ratios (morphology). Leaf area index (LAI), defined as “the one-sided green leaf area per unit ground surface area”, is an important indicator reflecting plant productivity. Alpine plants in the reproductive period (S3 stage) had the largest LAI, and the absolute growth rate of dominant species was approximately 2 times that of plants in the vegetative period (S2 stage) on the QTP [62]. Plants with higher LAI or coverage consume more soil moisture and are more sensitive to precipitation changes, which explains why alpine plant productivity is highly responsive to P3.
Periodic moisture shortages commonly occurs in alpine plants in a semi-arid climate [56], and they also appear on the QTP, which is characterized as a semi-arid or arid region. Thus, vegetation types dominated with species with a higher leaf area are more likely to be affected by water availability because these plants consume more soil moisture, as mentioned above. The peak LAI in the S3 stage varied among different vegetation types within the study area [116,117,118]. According to observations, LAI in ASM, TAM, and AST is approximately 2.5~4.0 [118,119], 1~2.5 [118,119], and 0.2~1.9 [117,118,119] m2m−2, respectively. Therefore, the influence of P3 was more significant in alpine meadows (TAM and ASM) than in alpine steppes, and its influence in ASM was higher than that in TAM.
4.3.2. Better Water Availability Promotes Bud Bank Size and Status at the S4 Stage Leading to a Higher ANPP
Changes in soil water availability can influence grassland productivity by altering the number of active buds in bud banks [120]. Many studies have indicated that plant population regeneration and maintenance are principally regulated by belowground bud bank dynamics and vegetative reproduction in perennial herbaceous plants [121,122,123]. Rainfall or drought events that occur during the peak periods of bud production and outgrowth significantly affect bud bank size [24,25] since bud primordium production and its differentiation and development are particularly sensitive to water stress [124].
Overwintering buds are common in most alpine plants on the Tibetan Plateau. Alpine meadows and steppes are the two main vegetation types on the QTP. Kobresia, the dominant species of alpine meadows, is a rhizomatous perennial sedge that is widely distributed in the eastern region of the QTP. Overwintering buds of a typical tiller of Kobresia (Kobresia humilis, K. pygmaea, and K. tibetica) generally emerge near September [62,79,80,125]. Stipa purpurea, one of the dominant species in alpine steppes, is a perennial bunchgrass species. Given the lack of relevant materials on the growth cycle of S. purpurea, we used two perennial bunchgrasses, Leymus chinensis [120] and Agropyron desertorum [124], as references, which are widely distributed in the Eastern Eurasian steppes and the Great Basin, USA, respectively. The two bunchgrasses grew overwintering buds in the fall. Based on the behaviors of the two bunchgrasses, we postulate that overwintering buds of S. purpurea may also shoot out in fall. Consequently, water availability in the S4 stage greatly influenced sprout and bud bank sizes, which subsequently affected vegetation growth (Figure 5).
4.3.3. Higher Temperature at the S0 Stage Benefits Overwintering Buds Facilitating Plant Growth Next Year
For perennial herbaceous plants, the belowground meristem population related with rhizomes and other perennial organs (bud bank) plays an essential role in local plant structure, population persistence, and dynamics [122,126]. Overwintering buds (or meristem in general) commonly exist in most alpine plants on the QTP, but they are more easily damaged by low temperatures during winter period (e.g., frost damage) [127,128,129]. Although buds of perennial herbaceous plants can be protected via insulation from snow cover, fewer and infrequent snowfalls lead to very thin snow cover on the QTP (winter snowfalls at 55 stations from 1971 to 2011 were <30 mm [130]), which is much lower than that in the high-latitude and Arctic regions of Europe and North America, where the low temperature environment is similar to that of the QTP [131,132]. Additionally, strong winds in winter will further hamper the formation of extensive snow cover [133]. Therefore, warmer winter can protect overwintering buds from freezing damage and promote the bud survival rate, which explains why vegetation growth exerts a significant positive response to T0 (Figure 5).
4.4. Interannual Carbon Circulation May Decrease the ANPP Responses to Antecedent Vegetation Growth Status
Our results illustrated that the antecedent vegetation growth status had non-significant impacts on the current year’s ANPP, except for alpine steppe plants (Figure 3 and Figure 5). This may be related to the interannual carbon circulation for perennial plants in the alpine ecosystem. Plants accumulate NSCs seasonally in underground organs before the onset of cold or dry periods and transfer them to aboveground organs the following year, serving as a vital resource for life activities next year [53,54,55]. This type of carbon allocation strategy ensures that plants grow and reproduce securely even in years with extreme climatic conditions, and this is particularly critical for the development of alpine vegetation in harsh environments [56]. Therefore, this strategy will reduce annual ANPP fluctuations, leading to a low legacy impact of prior-year plant growth status.
The interannual carbon circulation of perennial herbs is commonly observed on the QTP. Many reports have demonstrated that the underground biomass of plants in the alpine meadow on the QTP presented an N-shaped change from May to October during the growing season, and seasonal valley and peak values were found in July–August and September–October, respectively [134,135,136]. This is because most plants in alpine meadows on the QTP enter the reproductive period from July to August, particularly with flowering and fruiting, and consume plentiful NSCs stored underground in the previous fall [62,136]. Thus, plants can grow and reproduce safely even under extreme climatic conditions, ensuring a stabilized ANPP.
Compared with sedges (the dominant species in alpine meadows), grasses (the dominant species in alpine steppes) have relatively lower R/S ratios on the QTP [57,58,59,60,61]. R/S ratios can be interpreted as manifesting the differential photosynthate investment between the above- and belowground organs in plants [137]. Consequently, ANPP fluctuations were magnified in alpine steppe plants with lower R/S ratios because of relatively fewer stored NSCs in underground organs each year, resulting in a higher correlation with antecedent vegetation growth status. Although ANPP variations in alpine steppes are affected by the antecedent plant growth status, this impact is significantly weaker than that of precipitation because of the generally high R/S ratios of alpine plants (Figure 3 and Figure 5).
4.5. Research Implications and Uncertainties
This study demonstrated that alpine plants were more sensitive to precipitation (Figure 3) and exerted divergent responses to climatic factors during different developmental stages (Figure 5), which differed from the antecedent understanding based on remote sensing images and in situ observations on the QTP. This study also showed that the antecedent plant growth status has no significant impact on ANPP variations, except for AST (Figure 3 and Figure 5). These results shed further light on the mechanisms underlying the short-term responses of alpine ecosystems to climate variation and legacy effects. Given the divergent responses of different-stage climatic factors and the divergent impacts of antecedent plant growth status for different vegetation types, short-term grassland productivity can be predicted more accurately, which can help predict forage yield and be used as guidelines for alpine grassland management on the QTP.
This study also involved uncertainties in its results. First, the satellite-based NDVI was taken as a proxy for ANPP and we did not use the observed ANPP directly. This may introduce uncertainties. NDVI may also have some limitations in identifying the ANPP of plants in ASM with open water because water bodies can affect the reflection albedo of vegetation from the mixed pixels of the used images [138]. Second, although we aimed to reveal the impact of climatic factors on vegetation ANPP on the QTP, anthropogenic activities, such as grazing, can also influence grassland ANPP. However, complete separation of these factors remains challenging [18,93,139]. Third, the poor research foundation of plant physiology and ecology on the QTP brought additional uncertainties in ascertaining the responsive mechanisms of ANPP to climatic change. In this study, we only presented the possible responsive mechanisms of ANPP to climatic change, and more fieldwork and investigation should be conducted to verify and improve the understanding of these mechanisms.
5. Conclusions
We detected the responses of alpine grassland ANPP to the current-year climatic variables at the developmental stage scale and the prior-year plant growth status using a temporal model on the QTP. The results indicated that precipitation, rather than temperature, was the main driving factor for all vegetation types. The reproductive-stage (July–August) precipitation was the most prominent controlling factor, and the withering stage (September–October) precipitation and dormant stage temperature (November–February) also significantly affected grassland productivity. For different vegetation types, the influence of precipitation was more significant in alpine meadows (TAM and ASM) than in alpine steppes, and the influence in ASM was higher than in TAM. This study also showed that the impact of prior-year plant growth status on ANPP was non-significant, except for the alpine steppes, and even the conspicuous impact from the alpine steppes was much less than that of current-year climatic factors. Given the divergent responses of climatic factors at different developmental stages and the divergent impacts of antecedent plant growth status for different vegetation types, more accurate predictions of grassland productivity and forage yield can be provided on the QTP.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/rs14071564/s1, Table S1: Previous studies on the dominant climatic factors of vegetation productivity variations at different time scales on the Qinghai–Tibet Plateau [15,17,18,19,20,22,23,29,70,92,93,94,95,96,140,141,142,143,144,145,146].
Author Contributions
Conceptualization, D.Z., B.L. and X.G.; formal analysis, D.Z. and B.L.; methodology, D.Z., B.L., Y.Y., W.L. and J.X.; investigation, Y.L. (Yan Liu), Y.L. (Ying Li), R.L. and W.L.; resources, X.G., B.L. and Y.Y.; data curation, Y.J.; writing—original draft preparation, D.Z.; writing—review and editing, D.Z. and B.L.; visualization, D.Z., W.L. and J.X.; supervision, X.G.; project administration, B.L.; funding acquisition, B.L. and X.G. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (grant no. U2243206) and the National Key Research and Development Plan of China (grant no. 2016YFC0500205). This research was sponsored by the Ministry of Science and Technology of China.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Guo, Q.; Hu, Z.M.; Li, S.G.; Li, X.R.; Sun, X.M.; Yu, G.R. Spatial variations in aboveground net primary productivity along a climate gradient in Eurasian temperate grassland: Effects of mean annual precipitation and its seasonal distribution. Glob. Chang. Biol. 2012, 18, 3624–3631. [Google Scholar] [CrossRef]
- Craine, J.M.; Nippert, J.B.; Elmore, A.J.; Skibbe, A.M.; Hutchinson, S.L.; Brunsell, N.A. Timing of climate variability and grassland productivity. Proc. Natl. Acad. Sci. USA 2012, 109, 3401–3405. [Google Scholar] [CrossRef] [PubMed]
- Easterling, D.R.; Meehl, G.A.; Parmesan, C.; Changnon, S.A.; Karl, T.R.; Mearns, L.O. Climate extremes: Observations, modeling, and impacts. Science 2000, 289, 2068–2074. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhang, F.; Li, Y.; Zhao, X.; Cao, G. Thirty-year variations of above-ground net primary production and precipitation-use efficiency of an alpine meadow in the north-eastern Qinghai-Tibetan Plateau. Grass Forage Sci. 2016, 71, 208–218. [Google Scholar] [CrossRef]
- Elmendorf, S.C.; Henry, G.H.R.; Hollister, R.D.; Bjork, R.G.; Bjorkman, A.D.; Callaghan, T.V.; Collier, L.S.; Cooper, E.J.; Cornelissen, J.H.; Day, T.A. Global assessment of experimental climate warming on tundra vegetation: Heterogeneity over space and time. Ecol. Lett. 2012, 15, 164–175. [Google Scholar] [CrossRef]
- Harris, R.B. Rangeland degradation on the Qinghai-Tibetan plateau: A review of the evidence of its magnitude and causes. J. Arid. Environ. 2010, 74, 1–12. [Google Scholar] [CrossRef]
- Wang, C.Z. A remote sensing perspective of alpine grasslands on the Tibetan Plateau: Better or worse under “Tibet Warming”? Remote Sens. Appl. Soc. Environ. 2016, 3, 36–44. [Google Scholar] [CrossRef]
- Dong, S.K.; Wang, X.X.; Liu, S.L.; Li, Y.Y.; Su, X.K.; Wen, L.; Zhu, L. Reproductive responses of alpine plants to grassland degradation and artificial restoration in the Qinghai-Tibetan Plateau. Grass Forage Sci. 2015, 70, 229–238. [Google Scholar] [CrossRef]
- Wilcox, K.R.; Blair, J.M.; Smith, M.D.; Knapp, A.K. Does ecosystem sensitivity to precipitation at the site-level conform to regional-scale predictions? Ecology 2016, 97, 561–568. [Google Scholar] [CrossRef]
- Adler, P.B.; White, E.P.; Cortez, M.H. Matching the forecast horizon with the relevant spatial and temporal processes and data sources. Ecography 2020, 43, 1729–1739. [Google Scholar] [CrossRef]
- Knapp, A.K.; Ciais, P.; Smith, M.D. Reconciling inconsistencies in precipitation-productivity relationships: Implications for climate change. New Phytol. 2017, 214, 41–47. [Google Scholar] [CrossRef]
- Huxman, T.E.; Smith, M.D.; Fay, P.A.; Knapp, A.K.; Shaw, M.R.; Loik, M.E.; Smith, S.D.; Tissue, D.T.; Zak, J.C.; Weltzin, J.F.; et al. Convergence across biomes to a common rain-use efficiency. Nature 2004, 429, 651–654. [Google Scholar] [CrossRef]
- Munson, S.M.; Bunting, E.L.; Bradford, J.B.; Butterfield, B.J.; Gremer, J.R. Plant Production Responses to Precipitation Differ Along an Elevation Gradient and Are Enhanced Under Extremes. Ecosystems 2018, 22, 699–708. [Google Scholar] [CrossRef]
- Zhang, G.L.; Zhang, Y.J.; Dong, J.W.; Xiao, X.M. Green-up dates in the Tibetan Plateau have continuously advanced from 1982 to 2011. Proc. Natl. Acad. Sci. USA 2013, 110, 4309–4314. [Google Scholar] [CrossRef]
- Zheng, Z.T.; Zhu, W.Q.; Zhang, Y.J. Seasonally and spatially varied controls of climatic factors on net primary productivity in alpine grasslands on the Tibetan Plateau. Glob. Ecol. Conserv. 2020, 21, e00814. [Google Scholar] [CrossRef]
- Gao, Q.Z.; Guo, Y.Q.; Xu, H.M.; Ganjurjav, H.; Li, Y.; Wan, Y.F.; Qin, X.B.; Ma, X.; Liu, S. Climate change and its impacts on vegetation distribution and net primary productivity of the alpine ecosystem in the Qinghai-Tibetan Plateau. Sci. Total Environ. 2016, 554–555, 34–41. [Google Scholar] [CrossRef]
- Gao, Y.H.; Zhou, X.; Wang, Q.; Wang, C.Z.; Zhan, Z.M.; Chen, L.F.; Yan, J.X.; Qu, R. Vegetation net primary productivity and its response to climate change during 2001–2008 in the Tibetan Plateau. Sci. Total Environ. 2013, 444, 356–362. [Google Scholar] [CrossRef]
- Chen, B.X.; Zhang, X.Z.; Tao, J.; Wu, J.S.; Wang, J.S.; Shi, P.L.; Zhang, Y.J.; Yu, C.Q. The impact of climate change and anthropogenic activities on alpine grassland over the Qinghai-Tibet Plateau. Agric. For. Meteorol. 2014, 189–190, 11–18. [Google Scholar] [CrossRef]
- Cong, N.; Shen, M.G.; Yang, W.; Yang, Z.Y.; Zhang, G.X.; Piao, S.L. Varying responses of vegetation activity to climate changes on the Tibetan Plateau grassland. Int. J. Biometeorol. 2017, 61, 1433–1444. [Google Scholar] [CrossRef]
- Li, L.H.; Zhang, Y.L.; Wu, J.S.; Li, S.C.; Zhang, B.H.; Zu, J.X.; Zhang, H.M.; Ding, M.J.; Paudel, B. Increasing sensitivity of alpine grasslands to climate variability along an elevational gradient on the Qinghai-Tibet Plateau. Sci. Total Environ. 2019, 678, 21–29. [Google Scholar] [CrossRef]
- Zhe, M.; Zhang, X.Q. Time-lag effects of NDVI responses to climate change in the Yamzhog Yumco Basin, South Tibet. Ecol. Indic. 2021, 124, 107431. [Google Scholar] [CrossRef]
- Pang, G.J.; Wang, X.J.; Yang, M.X. Using the NDVI to identify variations in, and responses of, vegetation to climate change on the Tibetan Plateau from 1982 to 2012. Quat. Int. 2017, 444, 87–96. [Google Scholar] [CrossRef]
- Xu, W.X.; Gu, S.; Zhao, X.Q.; Xiao, J.S.; Tang, Y.H.; Fang, J.Y.; Zhang, J.; Jiang, S. High positive correlation between soil temperature and NDVI from 1982 to 2006 in alpine meadow of the Three-River Source Region on the Qinghai-Tibetan Plateau. Int. J. Appl. Earth Obs. Geoinf. 2011, 13, 528–535. [Google Scholar] [CrossRef]
- Van der Weide, B.L.; Hartnett, D.C.; Carter, D.L. Belowground bud banks of tallgrass prairie are insensitive to multi-year, growing-season drought. Ecosphere 2014, 5, 103. [Google Scholar] [CrossRef]
- Ott, J.P.; Klimesova, J.; Hartnett, D.C. The ecology and significance of below-ground bud banks in plants. Ann. Bot. 2019, 123, 1099–1118. [Google Scholar] [CrossRef]
- Chapin III, F.S.; Bret-Harte, M.S.; Hobbie, S.E.; Zhong, H. Plant functional types as predictors of transient responses of arctic vegetation to global change. J. Veg. Sci. 1996, 7, 347–358. [Google Scholar] [CrossRef]
- Robinson, T.M.P.; La Pierre, K.J.; Vadeboncoeur, M.A.; Byrne, K.M.; Thomey, M.L.; Colby, S.E. Seasonal, not annual precipitation drives community productivity across ecosystems. Oikos 2013, 122, 727–738. [Google Scholar] [CrossRef]
- La Pierre, K.J.; Yuan, S.; Chang, C.C.; Avolio, M.L.; Hallett, L.M.; Schreck, T.; Smith, M.D. Explaining temporal variation in above-ground productivity in a mesic grassland: The role of climate and flowering. J. Ecol. 2011, 99, 1250–1262. [Google Scholar] [CrossRef][Green Version]
- Zhang, L.; Guo, H.D.; Wang, C.Z.; Ji, L.; Li, J.; Wang, K.; Dai, L. The long-term trends (1982–2006) in vegetation greenness of the alpine ecosystem in the Qinghai-Tibetan Plateau. Environ. Earth Sci. 2014, 72, 1827–1841. [Google Scholar] [CrossRef]
- Jiang, W.G.; Yuan, L.H.; Wang, W.J.; Cao, R.; Zhang, Y.F.; Shen, W.M. Spatio-temporal analysis of vegetation variation in the Yellow River Basin. Ecol. Indic. 2015, 51, 117–126. [Google Scholar] [CrossRef]
- Paruelo, J.M.; Epstein, H.E.; Lauenroth, W.K.; Burke, I.C. ANPP estimates from NDVI for the central grassland region of the United States. Ecology 1997, 78, 953–958. [Google Scholar] [CrossRef]
- Dudney, J.; Hallett, L.M.; Larios, L.; Farrer, E.C.; Spotswood, E.N.; Stein, C.; Suding, K.N. Lagging behind: Have we overlooked previous-year rainfall effects in annual grasslands? J. Ecol. 2017, 105, 484–495. [Google Scholar] [CrossRef]
- Sherry, R.A.; Arnone, J.A.; Johnson, D.W.; Schimel, D.S.; Verburg, P.S.; Luo, Y. Carry over from previous year environmental conditions alters dominance hierarchy in a prairie plant community. J. Plant Ecol. 2012, 5, 134–146. [Google Scholar] [CrossRef]
- Tenhumberg, B.; Crone, E.E.; Ramula, S.; Tyre, A.J. Time-lagged effects of weather on plant demography: Drought and Astragalus scaphoides. Ecology 2018, 99, 915–925. [Google Scholar] [CrossRef]
- Zhong, L.; Ma, Y.M.; Xue, Y.K.; Piao, S.L. Climate Change Trends and Impacts on Vegetation Greening Over the Tibetan Plateau. J. Geophys. Res. Atmos. 2019, 124, 7540–7552. [Google Scholar] [CrossRef]
- Hoover, D.L.; Lauenroth, W.K.; Milchunas, D.G.; Porensky, L.M.; Augustine, D.J.; Derner, J.D. Sensitivity of productivity to precipitation amount and pattern varies by topographic position in a semiarid grassland. Ecosphere 2021, 12, e03376. [Google Scholar] [CrossRef]
- Sherry, R.A.; Weng, E.; Arnone Iii, J.A.; Johnson, D.W.; Schimel, D.S.; Verburg, P.S.; Wallace, L.L.; Luo, Y.Q. Lagged effects of experimental warming and doubled precipitation on annual and seasonal aboveground biomass production in a tallgrass prairie. Glob. Chang. Biol. 2008, 14, 2923–2936. [Google Scholar] [CrossRef]
- Sun, Q.Q.; Meyer, W.S.; Marschner, P. Direct and carry-over effects of summer rainfall on ecosystem carbon uptake and water use efficiency in a semi-arid woodland. Agric. For. Meteorol. 2018, 263, 15–24. [Google Scholar] [CrossRef]
- Chen, T.; de Jeu, R.A.M.; Liu, Y.Y.; van der Werf, G.R.; Dolman, A.J. Using satellite based soil moisture to quantify the water driven variability in NDVI: A case study over mainland Australia. Remote Sens. Environ. 2014, 140, 330–338. [Google Scholar] [CrossRef]
- Saatchi, S.; Asefi-Najafabady, S.; Malhi, Y.; Aragao, L.E.; Anderson, L.O.; Myneni, R.B.; Nemani, R. Persistent effects of a severe drought on Amazonian forest canopy. Proc. Natl. Acad. Sci. USA. 2013, 110, 565–570. [Google Scholar] [CrossRef]
- Evans, S.E.; Burke, I.C. Carbon and nitrogen decoupling under an 11-year drought in the shortgrass steppe. Ecosystems 2013, 16, 20–33. [Google Scholar] [CrossRef]
- Giese, M.; Gao, Y.Z.; Lin, S.; Brueck, H. Nitrogen availability in a grazed semi-arid grassland is dominated by seasonal rainfall. Plant Soil 2010, 340, 157–167. [Google Scholar] [CrossRef]
- Shen, W.; Jenerette, G.D.; Hui, D.; Scott, R.L. Precipitation legacy effects on dryland ecosystem carbon fluxes: Direction, magnitude and biogeochemical carryovers. Biogeosciences 2016, 13, 425–439. [Google Scholar] [CrossRef]
- Wiegand, T.; Snyman, H.A.; Kellner, K.; Paruelo, J.M. Do grasslands have a memory: Modeling phytomass production of a semiarid South African grassland. Ecosystems 2004, 7, 243–258. [Google Scholar] [CrossRef]
- Gong, Y.H.; Zhao, D.M.; Ke, W.B.; Fang, C.; Pei, J.Y.; Sun, G.J.; Ye, J.S. Legacy effects of precipitation amount and frequency on the aboveground plant biomass of a semi-arid grassland. Sci. Total Environ. 2020, 705, 135899. [Google Scholar] [CrossRef]
- Han, J.J.; Chen, J.Q.; Shi, W.Y.; Song, J.; Hui, D.F.; Ru, J.Y.; Wan, S.Q. Asymmetric responses of resource use efficiency to previous-year precipitation in a semi-arid grassland. Funct. Ecol. 2021, 35, 807–814. [Google Scholar] [CrossRef]
- Shi, Z.; Lin, Y.; Wilcox, K.R.; Souza, L.; Jiang, L.F.; Jiang, J.; Jung, C.G.; Xu, X.; Yuan, M.T.; Guo, X. Successional change in species composition alters climate sensitivity of grassland productivity. Glob. Chang. Biol. 2018, 24, 4993–5003. [Google Scholar] [CrossRef]
- Reichmann, L.G.; Sala, O.E.; Peters, D.P.C. Precipitation legacies in desert grassland primary production occur through previous-year tiller density. Ecology 2013, 94, 435–443. [Google Scholar] [CrossRef]
- Reichmann, L.G.; Sala, O.E.; Whitehead, D. Differential sensitivities of grassland structural components to changes in precipitation mediate productivity response in a desert ecosystem. Funct. Ecol. 2014, 28, 1292–1298. [Google Scholar] [CrossRef]
- Reichstein, M.; Bahn, M.; Ciais, P.; Frank, D.; Mahecha, M.D.; Seneviratne, S.I.; Zscheischler, J.; Beer, C.; Buchmann, N.; Frank, D.C.; et al. Climate extremes and the carbon cycle. Nature 2013, 500, 287–295. [Google Scholar] [CrossRef]
- Gao, Z.Y.; Lin, Z.J.; Niu, F.J.; Luo, J. Soil water dynamics in the active layers under different land-cover types in the permafrost regions of the Qinghai–Tibet Plateau, China. Geoderma 2020, 364, 114176. [Google Scholar] [CrossRef]
- Yahdjian, L.; Sala, O.E. Size of Precipitation Pulses Controls Nitrogen Transformation and Losses in an Arid Patagonian Ecosystem. Ecosystems 2010, 13, 575–585. [Google Scholar] [CrossRef]
- Diaz-Toribio, M.H.; Putz, F.E. Underground carbohydrate stores and storage organs in fire-maintained longleaf pine savannas in Florida, USA. Am. J. Bot. 2021, 108, 432–442. [Google Scholar] [CrossRef]
- De Moraes, M.G.; de Carvalho, M.A.M.; Franco, A.C.; Pollock, C.J.; Figueiredo-Ribeiro, R.; de Cássia, L. Fire and Drought: Soluble Carbohydrate Storage and Survival Mechanisms in Herbaceous Plants from the Cerrado. BioScience 2016, 66, 107–117. [Google Scholar] [CrossRef]
- Wyka, T. Carbohydrate storage and use in an alpine population of the perennial herb, Oxytropis sericea. Oecologia 1999, 120, 198–208. [Google Scholar] [CrossRef]
- Körner, C. Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2003; ISBN 978-3642189692. [Google Scholar]
- Mou, X.M.; Li, X.G.; Zhao, N.N.; Yu, Y.W.; Kuzyakov, Y. Tibetan sedges sequester more carbon belowground than grasses: A 13C labeling study. Plant Soil 2018, 426, 287–298. [Google Scholar] [CrossRef]
- Song, M.H.; Hu, Q.W.; Tian, Y.Q.; Ouyang, H. Seasonal patterns of root and shoot interactions in an alpine meadow on the Tibetan Plateau. J. Plant Ecol. 2010, 5, 182–190. [Google Scholar] [CrossRef]
- You, Q.G.; Xue, X.; Peng, F.; Xu, M.H.; Duan, H.C.; Dong, S.Y. Comparison of ecosystem characteristics between degraded and intact alpine meadow in the Qinghai-Tibetan Plateau, China. Ecol. Eng. 2014, 71, 133–143. [Google Scholar] [CrossRef]
- Li, J.; Zhang, F.W.; Lin, L.; Li, H.Q.; Du, Y.G.; Li, Y.K.; Cao, G.M. Response of the plant community and soil water status to alpine Kobresia meadow degradation gradients on the Qinghai–Tibetan Plateau, China. Ecol. Res. 2015, 30, 589–596. [Google Scholar] [CrossRef]
- Zeng, C.X.; Wu, J.S.; Zhang, X.Z. Effects of Grazing on Above-vs. Below-Ground Biomass Allocation of Alpine Grasslands on the Northern Tibetan Plateau. PLoS ONE 2015, 10, e0135173. [Google Scholar] [CrossRef]
- Zhou, X.M. Chinese Kobresia Meadow; Science Press: Beijing, China, 2001; ISBN 9787030085702. (In Chinese) [Google Scholar]
- Zhang, T.; Li, B.L.; Yuan, Y.C.; Gao, X.Z.; Sun, Q.L.; Xu, L.L.; Jiang, Y.H. Spatial downscaling of TRMM precipitation data considering the impacts of macro-geographical factors and local elevation in the Three-River Headwaters Region. Remote Sens. Environ. 2018, 215, 109–127. [Google Scholar] [CrossRef]
- Qin, D.H. Ecological Protection and Sustainable Development in the Three-River Headwater; Regin Science Press: Beijing, China, 2014; ISBN 9787030387189. (In Chinese) [Google Scholar]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-scale geospatial analysis for everyone. Remote Sens. Environ. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- Stow, D.; Petersen, A.; Hope, A.; Engstrom, R.; Coulter, L. Greenness trends of Arctic tundra vegetation in the 1990s: Comparison of two NDVI data sets from NOAA AVHRR systems. Int. J. Remote Sens. 2010, 28, 4807–4822. [Google Scholar] [CrossRef]
- Montandon, L.M.; Small, E.E. The impact of soil reflectance on the quantification of the green vegetation fraction from NDVI. Remote Sens. Environ. 2008, 112, 1835–1845. [Google Scholar] [CrossRef]
- Piao, S.L.; Mohammat, A.; Fang, J.Y.; Cai, Q.; Feng, J.M. NDVI-based increase in growth of temperate grasslands and its responses to climate changes in China. Glob. Environ. Chang. 2006, 16, 340–348. [Google Scholar] [CrossRef]
- Piao, S.L.; Fang, J.Y.; He, J.S. Variations in Vegetation Net Primary Production in the Qinghai-Xizang Plateau, China, from 1982 to 1999. Clim. Chang. 2006, 74, 253–267. [Google Scholar] [CrossRef]
- Peng, S.Z.; Ding, Y.X.; Wen, Z.M.; Chen, Y.M.; Cao, Y.; Ren, J.Y. Spatiotemporal change and trend analysis of potential evapotranspiration over the Loess Plateau of China during 2011–2100. Agric. For. Meteorol. 2017, 233, 183–194. [Google Scholar] [CrossRef]
- Peng, S.Z.; Ding, Y.X.; Liu, W.Z.; Li, Z. 1 km monthly temperature and precipitation dataset for China from 1901 to 2017. Earth Syst. Sci. Data 2019, 11, 1931–1946. [Google Scholar] [CrossRef]
- Zhang, X.S. 1:1 Million Vegetation Map of CHINA; Geological Publishing House: Beijing, China, 2008; ISBN 978-7-116-05146-1. [Google Scholar]
- Xu, W.; Zhu, M.Y.; Zhang, Z.H.; Ma, Z.Y.; Liu, H.Y.; Chen, L.T.; Cao, G.M.; Zhao, X.Q.; Schmid, B.; He, J.S. Experimentally simulating warmer and wetter climate additively improves rangeland quality on the Tibetan Plateau. J. Appl. Ecol. 2018, 55, 1486–1497. [Google Scholar] [CrossRef]
- Klein, J.A.; Harte, J.; Zhao, X.Q. Experimental warming, not grazing, decreases rangeland quality on the Tibetan Plateau. Ecol. Appl. 2007, 17, 541–557. [Google Scholar] [CrossRef]
- Hu, Z.M.; Yu, G.R.; Fan, J.W.; Zhong, H.P.; Wang, S.Q.; Li, S.G. Precipitation-use efficiency along a 4500-km grassland transect. Glob. Ecol. Biogeogr. 2010, 19, 842–851. [Google Scholar] [CrossRef]
- Wang, S.P.; Duan, J.C.; Xu, G.P.; Wang, Y.F.; Zhang, Z.H.; Rui, Y.C.; Luo, C.Y.; Xu, B.; Zhu, X.X.; Chang, X.F. Effects of warming and grazing on soil N availability, species composition, and ANPP in an alpine meadow. Ecology 2012, 93, 2365–2376. [Google Scholar] [CrossRef]
- Qiu, B.; Li, W.K.; Wang, X.Q.; Shang, L.Y.; Song, C.Q.; Guo, W.D.; Zhang, Y.G. Satellite-observed solar-induced chlorophyll fluorescence reveals higher sensitivity of alpine ecosystems to snow cover on the Tibetan Plateau. Agric. For. Meteorol. 2019, 271, 126–134. [Google Scholar] [CrossRef]
- Bell, K.L.; Bliss, L.C. Autecology of Kobresia bellardii: Why winter snow accumulation limits local distribution. Ecol. Monogr. 1979, 49, 377–402. [Google Scholar] [CrossRef]
- Yang, Y.W.; Li, X.L. Pilot study of clonal growth and reproduction of Kobresia humilis. J. Qinghai Univ. 2008, 26, 31–34. (In Chinese) [Google Scholar] [CrossRef]
- Lauenroth, W.K.; Hunt, H.W.; Swift, D.M.; Singh, J.S. Estimating aboveground net primary production in grasslands: A simulation approach. Ecol. Model. 1986, 33, 297–314. [Google Scholar] [CrossRef]
- Singh, J.S.; Lauenroth, W.K.; Steinhorst, R.K. Review and assessment of various techniques for estimating net aerial primary production in grasslands from harvest data. Bot. Rev. 1975, 41, 181–232. [Google Scholar] [CrossRef]
- Jobbágy, E.G.; Sala, O.E.; Paruelo, J.M. Patterns and controls of primary production in the Patagonian steppe: A remote sensing approach. Ecology 2002, 83, 307–319. [Google Scholar] [CrossRef][Green Version]
- Piao, S.L.; Friedlingstein, P.; Ciais, P.; Viovy, N.; Demarty, J. Growing season extension and its impact on terrestrial carbon cycle in the Northern Hemisphere over the past 2 decades. Glob. Biogeochem. Cycles 2007, 21, GB3018. [Google Scholar] [CrossRef]
- Oesterheld, M.; Loreti, J.; Semmartin, M.; Sala, O.E. Inter-annual variation in primary production of a semi-arid grassland related to previous-year production. J. Veg. Sci. 2001, 12, 137–142. [Google Scholar] [CrossRef]
- Griffin-Nolan, R.J.; Carroll, C.J.W.; Denton, E.M.; Johnston, M.K.; Collins, S.L.; Smith, M.D.; Knapp, A.K. Legacy effects of a regional drought on aboveground net primary production in six central US grasslands. Plant Ecol. 2018, 219, 505–515. [Google Scholar] [CrossRef]
- Zhang, Q.; Kong, D.D.; Shi, P.J.; Singh, V.P.; Sun, P. Vegetation phenology on the Qinghai-Tibetan Plateau and its response to climate change (1982–2013). Agric. For. Meteorol. 2018, 248, 408–417. [Google Scholar] [CrossRef]
- Guo, L.; Cheng, J.M.; Luedeling, E.; Koerner, S.E.; He, J.S.; Xu, J.C.; Gang, C.C.; Li, W.; Luo, R.M.; Peng, C.H. Critical climate periods for grassland productivity on China’s Loess Plateau. Agric. For. Meteorol. 2017, 233, 101–109. [Google Scholar] [CrossRef]
- Wang, L.; Yu, H.Y.; Zhang, Q.; Xu, Y.J.; Tao, Z.X.; Alatalo, J.H.; Dai, J.H. Responses of aboveground biomass of alpine grasslands to climate changes on the Qinghai-Tibet Plateau. J. Geogr. Sci. 2018, 28, 1953–1964. [Google Scholar] [CrossRef]
- Luedeling, E.; Gassner, A. Partial Least Squares Regression for analyzing walnut phenology in California. Agric. For. Meteorol. 2012, 158–159, 43–52. [Google Scholar] [CrossRef]
- Wold, S. PLS for multivariate linear modeling. In Chemometric Methods in Molecular Design: Methods and Principles in Medicinal Chemistry; VCH Verlagsgesellschaft mbH: Weinheim, Germany, 1995; pp. 195–218. [Google Scholar]
- Bai, Y.; Guo, C.; Degen, A.A.; Ahmad, A.A.; Wang, W.; Zhang, T.; Li, W.; Ma, L.; Huang, M.; Zeng, H.; et al. Climate warming benefits alpine vegetation growth in Three-River Headwater Region, China. Sci. Total Environ. 2020, 742, 140574. [Google Scholar] [CrossRef]
- Xu, H.J.; Wang, X.P.; Zhang, X.X. Alpine grasslands response to climatic factors and anthropogenic activities on the Tibetan Plateau from 2000 to 2012. Ecol. Eng. 2016, 92, 251–259. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, C.B.; Wang, Z.Q.; Chen, Y.Z.; Gang, C.C.; An, R.; Li, J.L. Vegetation dynamics and its driving forces from climate change and human activities in the Three-River Source Region, China from 1982 to 2012. Sci. Total Environ. 2016, 563–564, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Wang, Y.; Ma, Y.; Ma, W.; Liang, C.; Flynn, D.F.B.; Schmid, B.; Fang, J.; He, J.S. Field-based observations of regional-scale, temporal variation in net primary production in Tibetan alpine grasslands. Biogeosciences 2014, 11, 2003–2016. [Google Scholar] [CrossRef]
- Li, P.L.; Hu, Z.M.; Liu, Y.W. Shift in the trend of browning in Southwestern Tibetan Plateau in the past two decades. Agric. For. Meteorol. 2020, 287, 107950. [Google Scholar] [CrossRef]
- Fu, G.; Shen, Z.X.; Zhang, X.Z. Increased precipitation has stronger effects on plant production of an alpine meadow than does experimental warming in the Northern Tibetan Plateau. Agric. For. Meteorol. 2018, 249, 11–21. [Google Scholar] [CrossRef]
- Yuan, X.; Chen, Y.; Qin, W.K.; Xu, T.L.; Mao, Y.H.; Wang, Q.; Chen, K.L.; Zhu, B. Plant and microbial regulations of soil carbon dynamics under warming in two alpine swamp meadow ecosystems on the Tibetan Plateau. Sci. Total Environ. 2021, 790, 148072. [Google Scholar] [CrossRef]
- Yang, Y.; Hopping, K.A.; Wang, G.; Chen, J.; Peng, A.; Klein, J.A. Permafrost and drought regulate vulnerability of Tibetan Plateau grasslands to warming. Ecosphere 2018, 9, e02233. [Google Scholar] [CrossRef]
- Zhang, T.; Zhang, Y.J.; Xu, M.J.; Zhu, J.T.; Chen, N.; Jiang, Y.B.; Huang, K.; Zu, J.X.; Liu, Y.J.; Yu, G.R. Water availability is more important than temperature in driving the carbon fluxes of an alpine meadow on the Tibetan Plateau. Agric. For. Meteorol. 2018, 256–257, 22–31. [Google Scholar] [CrossRef]
- Wu, Z.T.; Dijkstra, P.; Koch, G.W.; PeÑUelas, J.; Hungate, B.A. Responses of terrestrial ecosystems to temperature and precipitation change: A meta-analysis of experimental manipulation. Glob. Chang. Biol. 2011, 17, 927–942. [Google Scholar] [CrossRef]
- Li, P.L.; Zhu, D.; Wang, Y.L.; Liu, D. Elevation dependence of drought legacy effects on vegetation greenness over the Tibetan Plateau. Agric. For. Meteorol. 2020, 295, 108190. [Google Scholar] [CrossRef]
- Fensholt, R.; Proud, S.R. Evaluation of earth observation based global long term vegetation trends—Comparing GIMMS and MODIS global NDVI time series. Remote Sens. Environ. 2012, 119, 131–147. [Google Scholar] [CrossRef]
- Liu, Y.C.; Li, Z.; Chen, Y.N.; Li, Y.P.; Li, H.W.; Xia, Q.Q.; Kayumba, P.M. Evaluation of consistency among three NDVI products applied to High Mountain Asia in 2000–2015. Remote Sens. Environ. 2022, 269, 112821. [Google Scholar] [CrossRef]
- Tian, F.; Fensholt, R.; Verbesselt, J.; Grogan, K.; Horion, S.; Wang, Y.J. Evaluating temporal consistency of long-term global NDVI datasets for trend analysis. Remote Sens. Environ. 2015, 163, 326–340. [Google Scholar] [CrossRef]
- Wang, H.; Liu, H.Y.; Huang, N.; Bi, J.; Ma, X.L.; Ma, Z.Y.; Shangguan, Z.J.; Zhao, H.F.; Feng, Q.S.; Liang, T.G. Satellite-derived NDVI underestimates the advancement of alpine vegetation growth over the past three decades. Ecology 2021, 102, e03518. [Google Scholar] [CrossRef] [PubMed]
- Way, D.A.; Yamori, W. Thermal acclimation of photosynthesis: On the importance of adjusting our definitions and accounting for thermal acclimation of respiration. Photosynth. Res. 2014, 119, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Elmendorf, S.C.; Henry, G.H.R.; Hollister, R.D.; Björk, R.G.; Boulanger-Lapointe, N.; Cooper, E.J.; Cornelissen, J.H.C.; Day, T.A.; Dorrepaal, E.; Elumeeva, T.G. Plot-scale evidence of tundra vegetation change and links to recent summer warming. Nat. Clim. Chang. 2012, 2, 453–457. [Google Scholar] [CrossRef]
- Yamasaki, T.; Yamakawa, T.; Yamane, Y.; Koike, H.; Satoh, K.; Katoh, S. Temperature acclimation of photosynthesis and related changes in photosystem II electron transport in winter wheat. Plant Physiol. 2002, 128, 1087–1097. [Google Scholar] [CrossRef] [PubMed]
- Mawson, B.T.; Cummins, W.R. Thermal acclimation of photosynthetic electron transport activity by thylakoids of Saxifraga cernua. Plant Physiol. 1989, 89, 325–332. [Google Scholar] [CrossRef]
- Erschbamer, B.; Kiebacher, T.; Mallaun, M.; Unterluggauer, P. Short-term signals of climate change along an altitudinal gradient in the South Alps. Plant Ecol. 2009, 202, 79–89. [Google Scholar] [CrossRef]
- Theurillat, J.P.; Guisan, A. Potential impact of climate change on vegetation in the European Alps: A review. Clim. Chang. 2001, 50, 77–109. [Google Scholar] [CrossRef]
- Billings, W.D.; Mooney, H.A. The ecology of arctic and alpine plants. Biol. Rev. 1968, 43, 481–529. [Google Scholar] [CrossRef]
- Pisek, A.; Larcher, W.; Vegis, A.; Napp-Zinn, K. The normal temperature range. In Temperature and Life; Springer: Berlin/Heidelberg, Germany, 1973; pp. 102–194. [Google Scholar]
- Kammer, P.M.; Schöb, C.; Eberhard, G.; Gallina, R.; Meyer, R.; Tschanz, C. The relationship between soil water storage capacity and plant species diversity in high alpine vegetation. Plant Ecol. Divers. 2013, 6, 457–466. [Google Scholar] [CrossRef]
- Liu, Y.W.; Geng, X.D.; Tenzintarchen; Wei, D.; Dai, D.X.; Xu, R. Divergence in ecosystem carbon fluxes and soil nitrogen characteristics across alpine steppe, alpine meadow and alpine swamp ecosystems in a biome transition zone. Sci. Total Environ. 2020, 748, 142453. [Google Scholar] [CrossRef]
- Hu, Z.M.; Yu, G.R.; Zhou, Y.L.; Sun, X.M.; Li, Y.N.; Shi, P.L.; Wang, Y.F.; Song, X.; Zheng, Z.M.; Zhang, L.; et al. Partitioning of evapotranspiration and its controls in four grassland ecosystems: Application of a two-source model. Agric. For. Meteorol. 2009, 149, 1410–1420. [Google Scholar] [CrossRef]
- Li, H.Q.; Wang, C.Y.; Zhang, F.W.; He, Y.T.; Shi, P.L.; Guo, X.W.; Wang, J.B.; Zhang, L.M.; Li, Y.N.; Cao, G.M.; et al. Atmospheric water vapor and soil moisture jointly determine the spatiotemporal variations of CO2 fluxes and evapotranspiration across the Qinghai-Tibetan Plateau grasslands. Sci. Total Environ. 2021, 791, 148379. [Google Scholar] [CrossRef]
- Qin, Y.; Yi, S.; Ren, S.; Li, N.; Chen, J. Responses of typical grasslands in a semi-arid basin on the Qinghai-Tibetan Plateau to climate change and disturbances. Environ. Earth Sci. 2013, 71, 1421–1431. [Google Scholar] [CrossRef]
- Wang, J.F.; Shi, Y.J.; Ao, Y.N.; Yu, D.F.; Wang, J.; Gao, S.; Knops, J.M.H.; Mu, C.S.; Li, Z.J. Summer drought decreases Leymus chinensis productivity through constraining the bud, tiller and shoot production. J. Agron. Crop Sci. 2019, 205, 554–561. [Google Scholar] [CrossRef]
- Benson, E.J.; Hartnett, D.C. The role of seed and vegetative reproduction in plant recruitment and demography in tallgrass prairie. Plant Ecol. 2006, 187, 163–178. [Google Scholar] [CrossRef]
- Dalgleish, H.J.; Hartnett, D.C. Below-ground bud banks increase along a precipitation gradient of the North American Great Plains: A test of the meristem limitation hypothesis. New Phytol. 2006, 171, 81–89. [Google Scholar] [CrossRef]
- Knapp, A.K.; Smith, M.D. Variation among biomes in temporal dynamics of aboveground primary production. Science 2001, 291, 481–484. [Google Scholar] [CrossRef]
- Busso, C.A.; Mueller, R.J.; Richards, J.H. Effects of drought and defoliation on bud viability in two caespitose grasses. Ann. Bot. 1989, 63, 477–485. [Google Scholar] [CrossRef]
- Deng, Z.F.; Xie, X.L.; Zhou, X.M. Primary Study on Reproductive Strategies of Kobresia humilis Population in Alpine Meadow. Chin. J. Ecol. 2001, 20, 68–70. (In Chinese) [Google Scholar] [CrossRef]
- Ott, J.P.; Hartnett, D.C. Contrasting bud bank dynamics of two co-occurring grasses in tallgrass prairie: Implications for grassland dynamics. Plant Ecol. 2012, 213, 1437–1448. [Google Scholar] [CrossRef]
- Malyshev, A.V.; Henry, H.A.L. Frost damage and winter nitrogen uptake by the grass Poa pratensis L.: Consequences for vegetative versus reproductive growth. Plant Ecol. 2012, 213, 1739–1747. [Google Scholar] [CrossRef]
- Rixen, C.; Dawes, M.A.; Wipf, S.; Hagedorn, F. Evidence of enhanced freezing damage in treeline plants during six years of CO2 enrichment and soil warming. Oikos 2012, 121, 1532–1543. [Google Scholar] [CrossRef]
- Wheeler, J.A.; Hoch, G.; Cortes, A.J.; Sedlacek, J.; Wipf, S.; Rixen, C. Increased spring freezing vulnerability for alpine shrubs under early snowmelt. Oecologia 2014, 175, 219–229. [Google Scholar] [CrossRef]
- Jiang, Y.H.; Li, B.L.; Yuan, Y.C.; Sun, Q.L.; Zhang, T.; Liu, Y.; Li, Y.; Li, R. Divergent shifts in flowering phenology of herbaceous plants on the warming Qinghai–Tibetan plateau. Agric. For. Meteorol. 2021, 307, 108502. [Google Scholar] [CrossRef]
- Bjorkman, A.D.; Elmendorf, S.C.; Beamish, A.L.; Vellend, M.; Henry, G.H. Contrasting effects of warming and increased snowfall on Arctic tundra plant phenology over the past two decades. Glob. Chang. Biol. 2015, 21, 4651–4661. [Google Scholar] [CrossRef]
- Esteban, P.; Jones, P.D.; Martín-Vide, J.; Mases, M. Atmospheric circulation patterns related to heavy snowfall days in Andorra, Pyrenees. Int. J. Climatol. 2005, 25, 319–329. [Google Scholar] [CrossRef]
- Ma, N.; Yu, K.L.; Zhang, Y.S.; Zhai, J.Q.; Zhang, Y.Q.; Zhang, H.B. Ground observed climatology and trend in snow cover phenology across China with consideration of snow-free breaks. Clim. Dyn. 2020, 55, 2867–2887. [Google Scholar] [CrossRef]
- Li, G.D. The study of underground phytomass and its seasonal changes in alpine grass—Kobresia meadow of Tianzhu area, Gansu. Cao Ye Xue Bao 1990, 1, 42–46. (In Chinese) [Google Scholar]
- Pu, J.Y.; Li, Y.N.; Zhao, L.; Yang, S.H. The relationship between seasonal changes of Kobresia humilis meadow biomass and the meteorological factors. Acta Agrestia Sin. 2005, 13, 238–241. (In Chinese) [Google Scholar] [CrossRef]
- Zhu, B.W.; Zhou, H.K.; Xu, Y.X.; Li, Y.N.; Tang, K. Study on seasonal dynamics of biomass in meadow grassland of north shore of Qinghai lake. Pratacult. Sci. 2008, 25, 62–66. (In Chinese) [Google Scholar] [CrossRef]
- Wu, Y.B.; Tan, H.C.; Deng, Y.C.; Wu, J.; Xu, X.L.; Wang, Y.F.; Tang, Y.H.; Higashi, T.R.; Cui, X.Y. Partitioning pattern of carbon flux in a Kobresia grassland on the Qinghai-Tibetan Plateau revealed by field 13C pulse-labeling. Glob. Chang. Biol. 2009, 16, 2322–2333. [Google Scholar] [CrossRef]
- Lenney, M.P.; Woodcock, C.E.; Collins, J.B.; Hamdi, H. The status of agricultural lands in Egypt: The use of multitemporal NDVI features derived from Landsat TM. Remote Sens. Environ. 1996, 56, 8–20. [Google Scholar] [CrossRef]
- Li, L.H.; Zhang, Y.L.; Liu, L.S.; Wu, J.S.; Li, S.C.; Zhang, H.Y.; Zhang, B.H.; Ding, M.J.; Wang, Z.F.; Paudel, B. Current challenges in distinguishing climatic and anthropogenic contributions to alpine grassland variation on the Tibetan Plateau. Ecol. Evol. 2018, 8, 5949–5963. [Google Scholar] [CrossRef]
- Guo, B.; Zang, W.Q.; Yang, F.; Han, B.M.; Chen, S.T.; Liu, Y.; Yang, X.; He, T.L.; Chen, X.; Liu, C.T.; et al. Spatial and temporal change patterns of net primary productivity and its response to climate change in the Qinghai-Tibet Plateau of China from 2000 to 2015. J. Arid. Land 2019, 12, 1–17. [Google Scholar] [CrossRef]
- Zhang, L.; Guo, H.D.; Ji, L.; Lei, L.P.; Wang, C.Z.; Yan, D.M.; Li, B.; Li, J. Vegetation greenness trend (2000 to 2009) and the climate controls in the Qinghai-Tibetan Plateau. J. Appl. Remote Sens. 2013, 7, 73572. [Google Scholar] [CrossRef]
- Yang, Y.H.; Piao, S.L. Variations in grassland vegetation cover in relation to climatic factors on the Tibetan Plateau. J. Plant Ecol. 2006, 30, 1–8. (In Chinese) [Google Scholar] [CrossRef]
- Du, J.Q.; Zhao, C.X.; Shu, J.M.; Jiaerheng, A.; Yuan, X.J.; Yin, J.Q.; Fang, S.F.; He, P. Spatiotemporal changes of vegetation on the Tibetan Plateau and relationship to climatic variables during multiyear periods from 1982–2012. Environ. Earth Sci. 2015, 75. [Google Scholar] [CrossRef]
- Xu, W.X.; Liu, X.D. Response of vegetation in the Qinghai-Tibet Plateau to global warming. Chin. Geogr. Sci. 2007, 17, 151–159. [Google Scholar] [CrossRef]
- Ding, M.J.; Zhang, Y.L.; Liu, L.S.; Zhang, W.; Wang, Z.F.; Bai, W.Q. The relationship between NDVI and precipitation on the Tibetan Plateau. J. Geogr. Sci. 2007, 17, 259–268. [Google Scholar] [CrossRef]
- Gao, Q.Z.; Li, Y.; Wan, Y.F.; Qin, X.B.; Jiangcun, W.Z.; Liu, Y.H. Dynamics of alpine grassland NPP and its response to climate change in Northern Tibet. Clim. Change 2009, 97, 515–528. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).