
First Form, Then Function: 3D Reconstruction of Cucumber Plants (Cucumis sativus L.) Allows Early Detection of Stress Effects through Leaf Dimensions



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Measurements
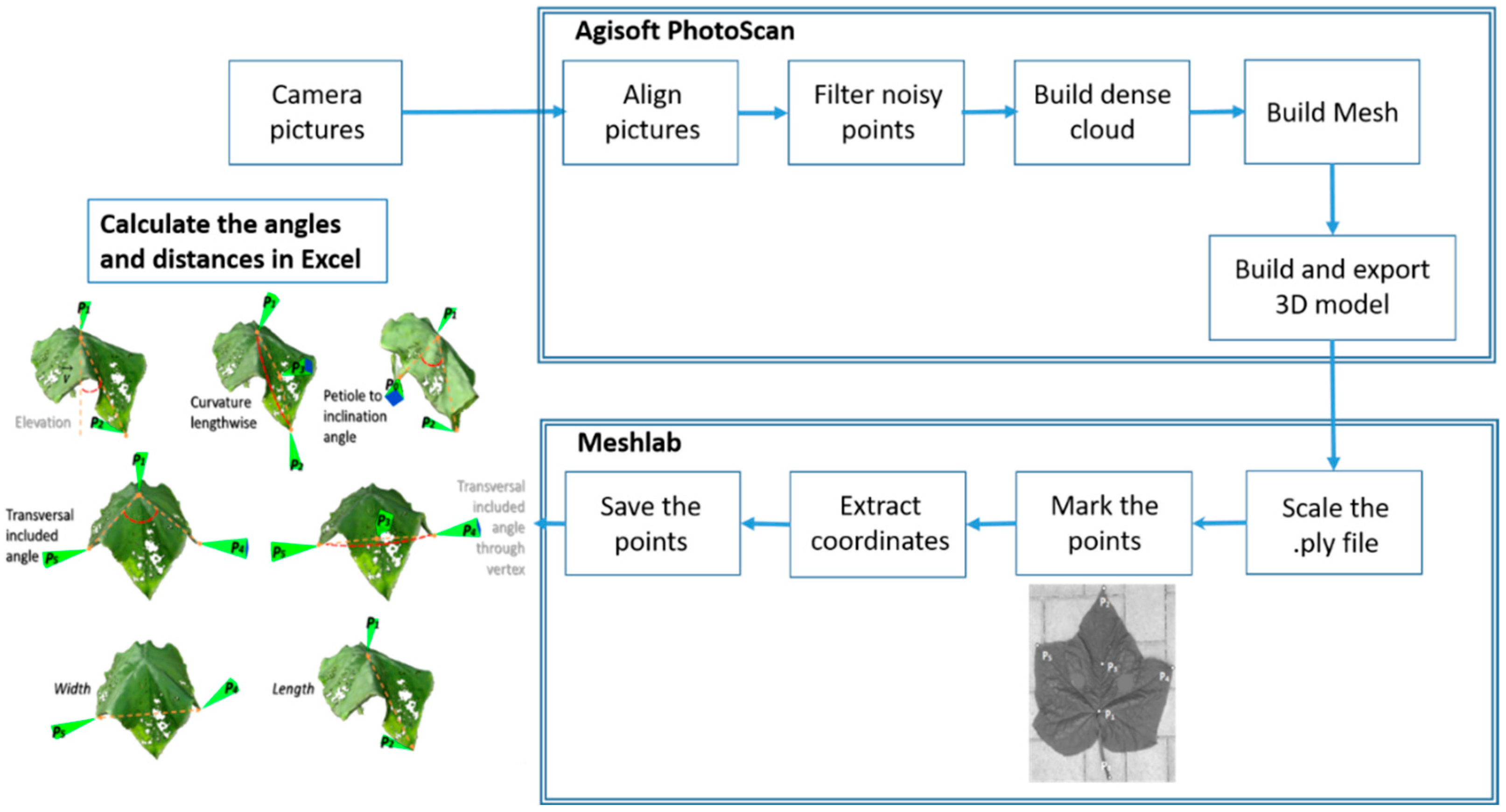
2.2.1. Non-Invasive Measurements
2.2.2. Invasive Measurements
2.3. Statistical Analysis
3. Results
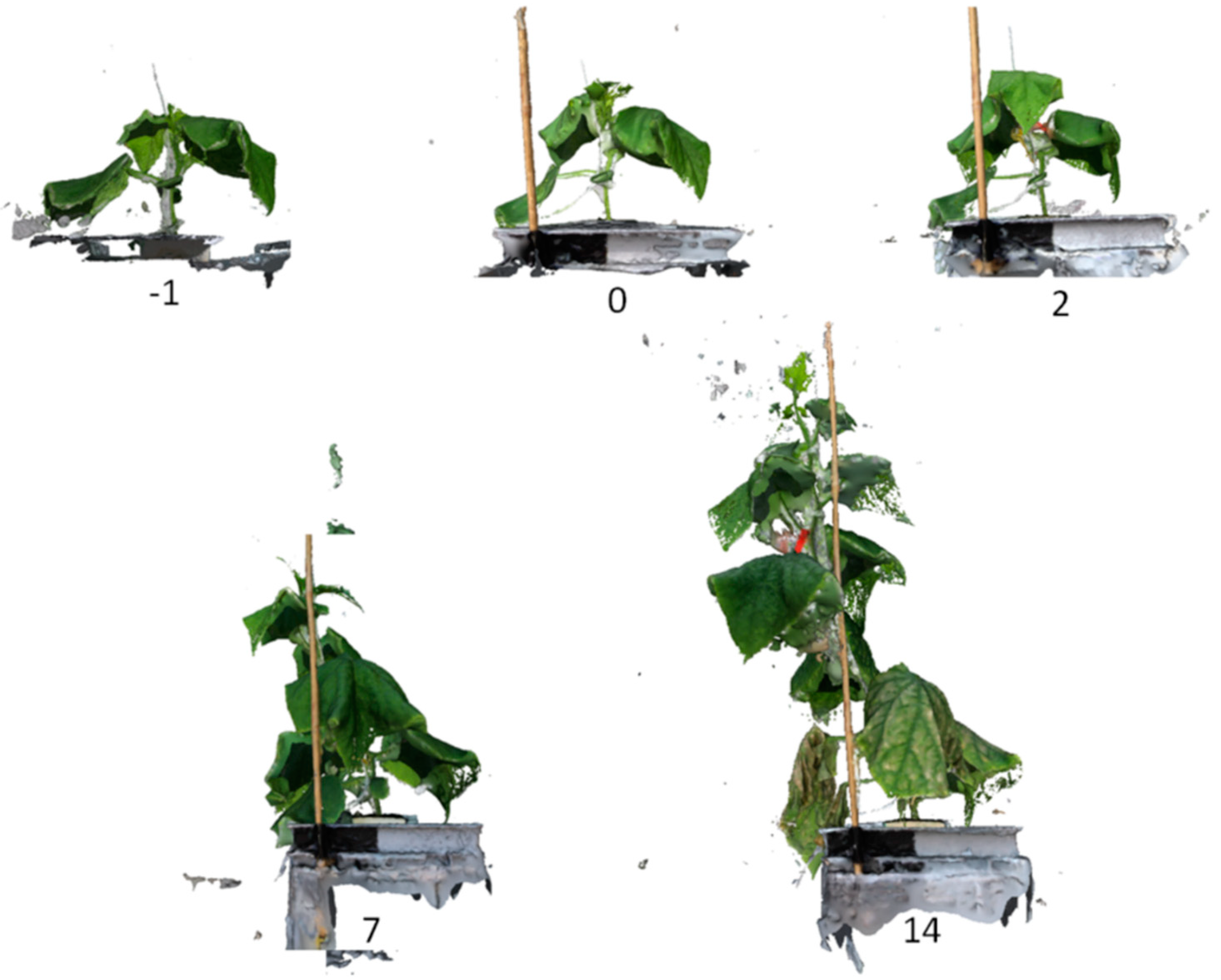
3.1. 3D Reconstruction of Plants
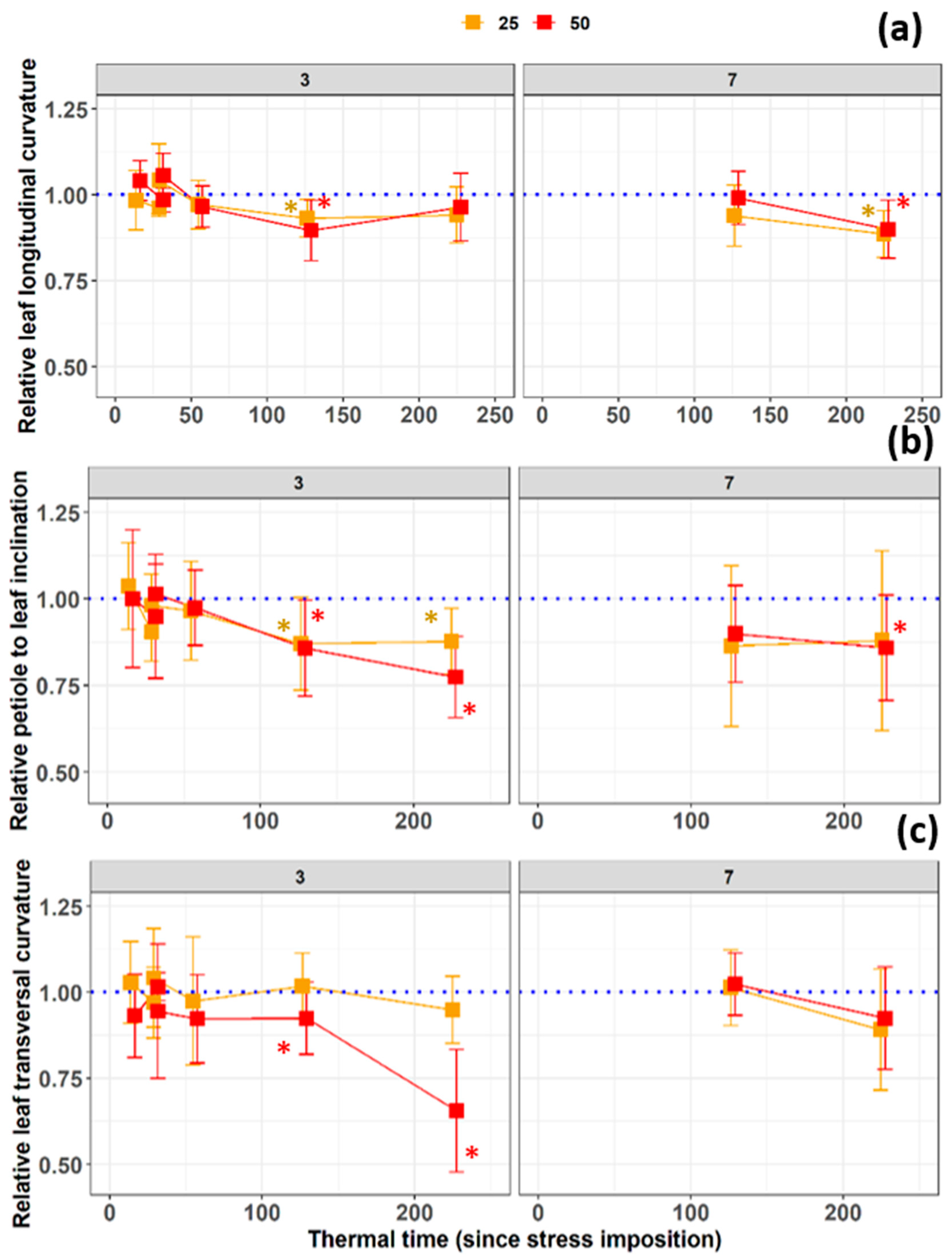
3.2. Early Detection of Salinity Stress from Leaf Longitudinal and Transversal Dimensions after 60 °Cd
3.3. Stress Response of the Transversal Leaf Included Angle
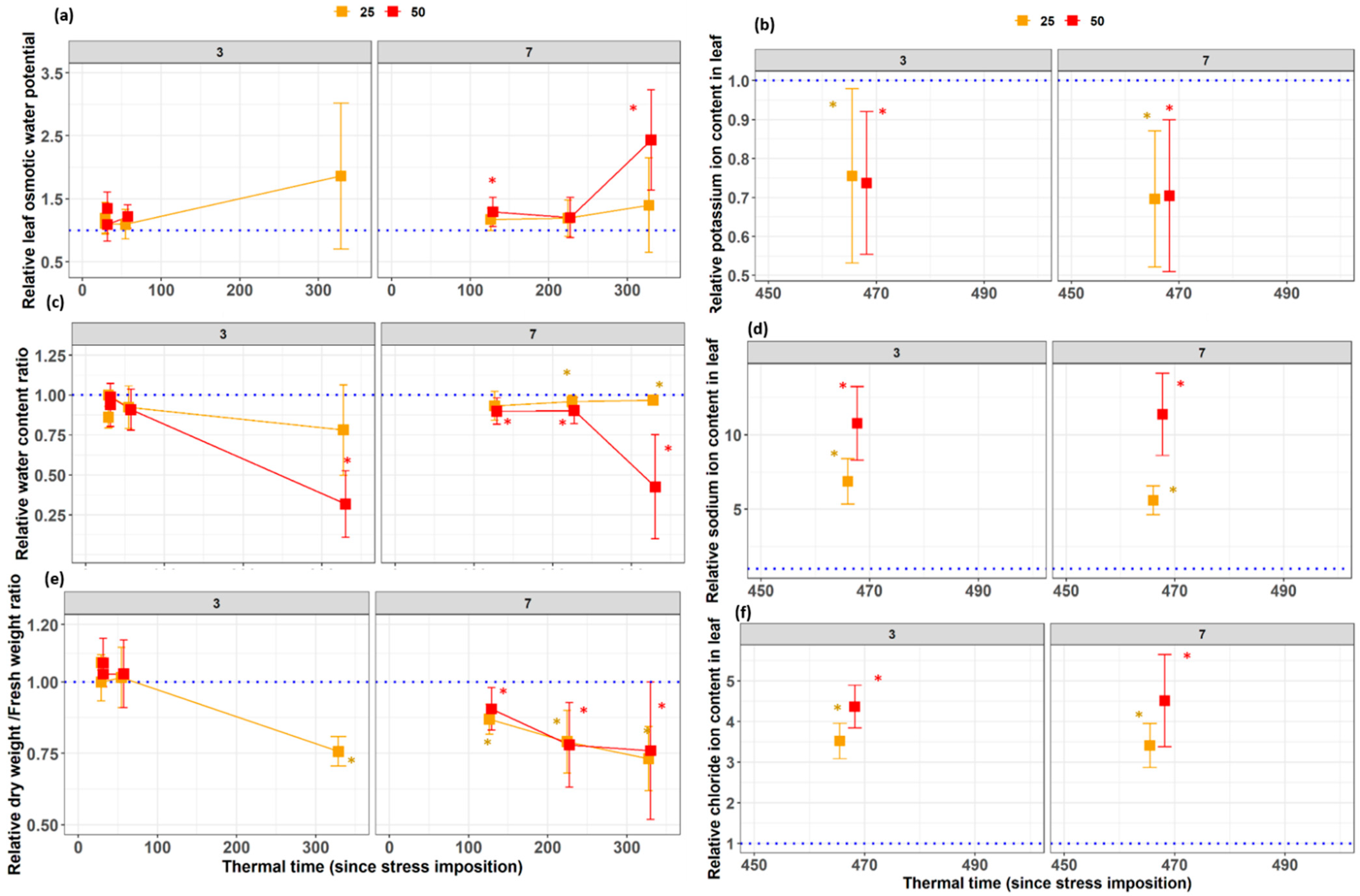
3.4. Physiological Traits
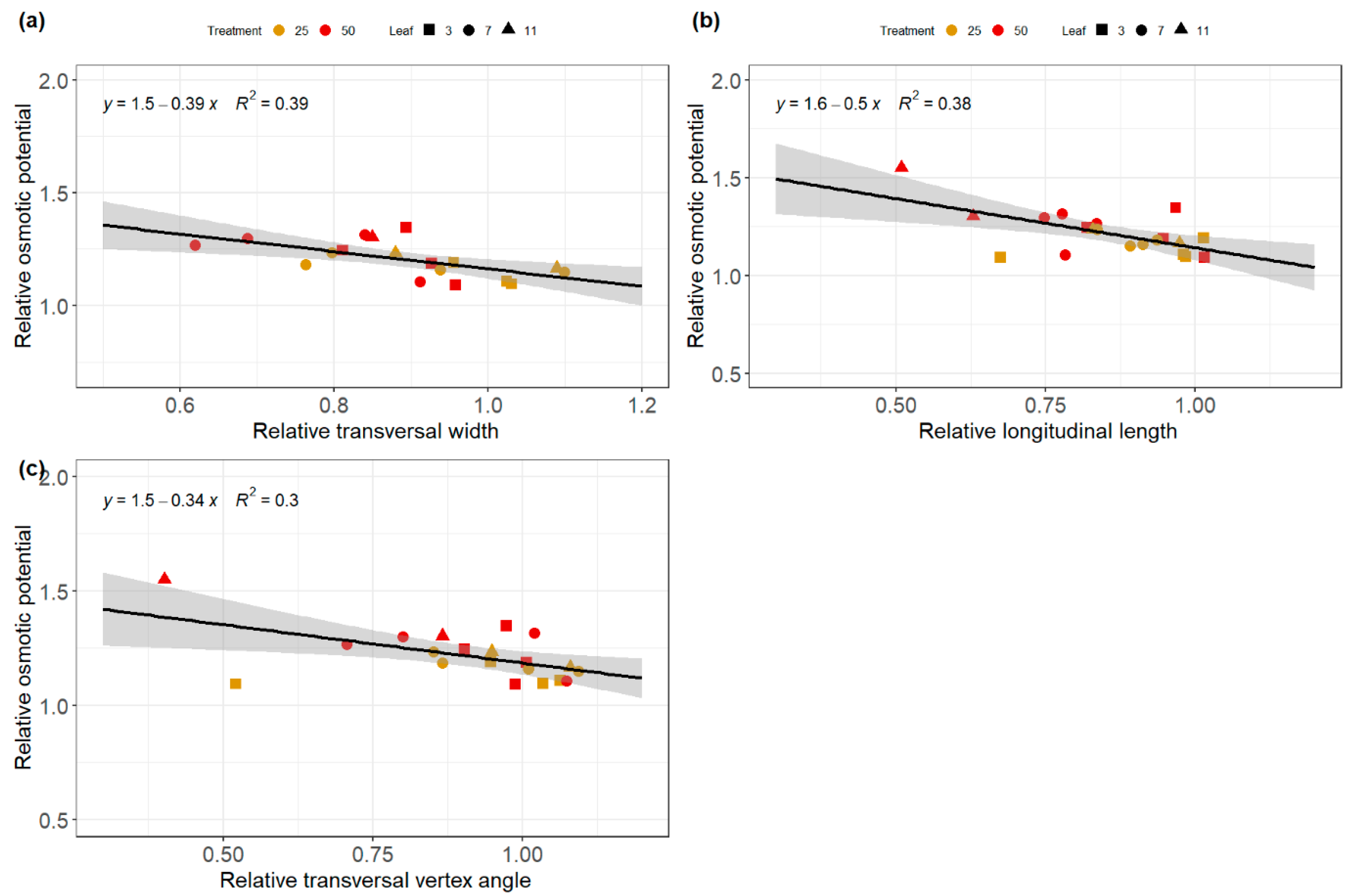
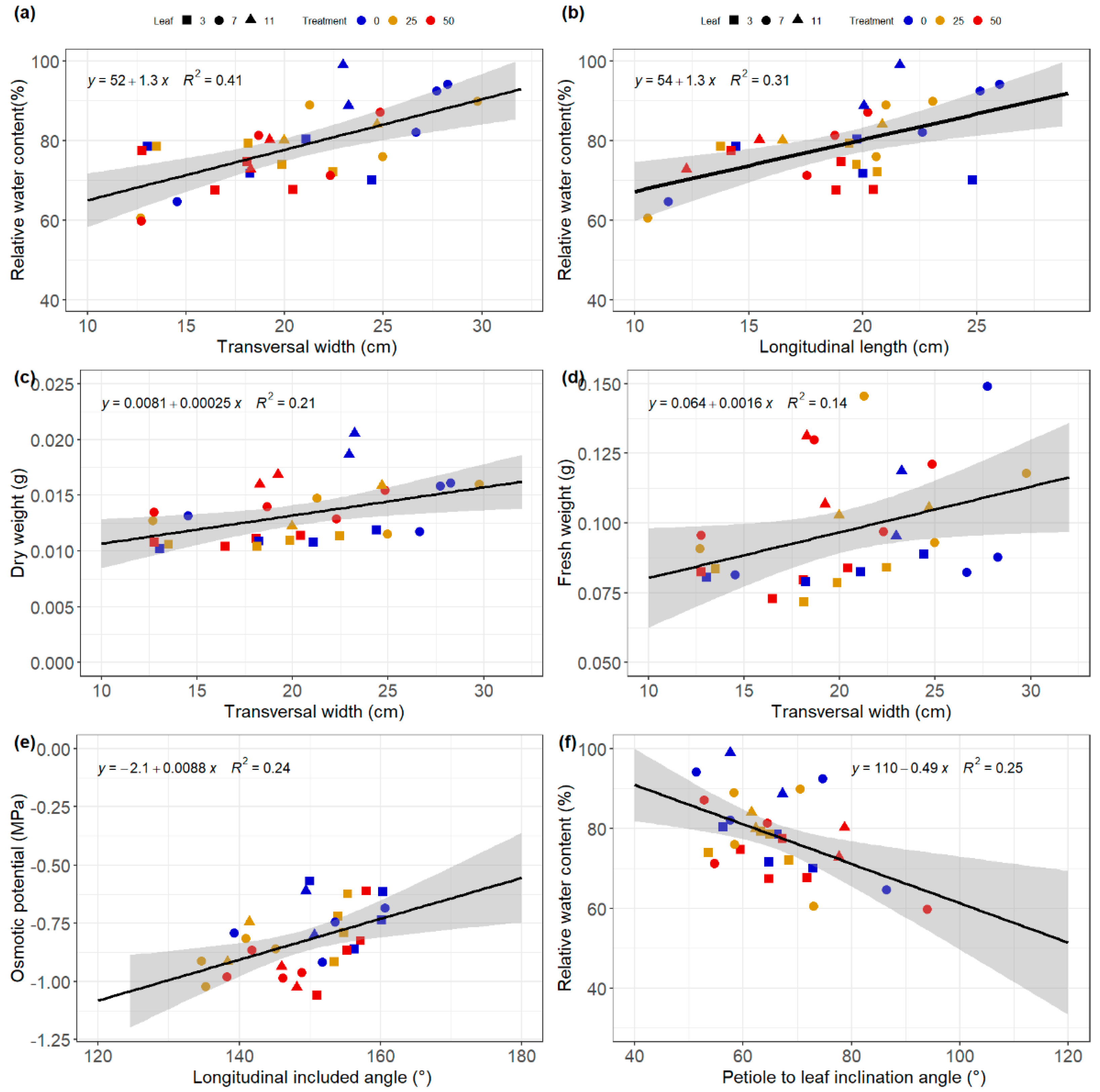
3.5. Relationships between Architectural and Physiological Traits
4. Discussion
4.1. Later Significant Differences between Stress and Control for Invasive Measurements
4.2. Earlier Response of Non-Invasive Morphological Traits
4.3. Relationship between Non-Invasive and Invasive Data
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- de Micco, V.; Aronne, G. morpho-anatomical traits for plant adaptation to drought. In Plant Responses to Drought Stress: From Morphological to Molecular Features; Aroca, R., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 37–61. ISBN 978-3-642-32653-0. [Google Scholar]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]
- Isayenkov, S.V.; Maathuis, F.J.M. Plant salinity stress: Many unanswered questions remain. Front. Plant Sci. 2019, 10, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drew, M.C.; Hold, P.S.; Picchioni, G.A. Inhibition by NaCl of Net CO2 Fixation and Yield of Cucumber. J. Am. Soc. Hortic. Sci. 1990, 115, 472–477. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.-W.; Kahlen, K.; Stützel, H. Disentangling the contributions of osmotic and ionic effects of salinity on stomatal, mesophyll, biochemical and light limitations to photosynthesis. Plant Cell Environ. 2015, 38, 1528–1542. [Google Scholar] [CrossRef] [PubMed]
- Stępień, P.; Kłbus, G. Water relations and photosynthesis in Cucumis sativus L. leaves under salt stress. Biol. Plant 2006, 50, 610–616. [Google Scholar] [CrossRef]
- Al-Karaki, G.N. Growth, water use efficiency, and sodium and potassium acquisition by tomato cultivars grown under salt stress. J. Plant Nutr. 2000, 23, 1–8. [Google Scholar] [CrossRef]
- Kahlen, K.; Stützel, H. Modelling photo-modulated internode elongation in growing glasshouse cucumber canopies. New Phytol. 2011, 190, 697–708. [Google Scholar] [CrossRef]
- Dassot, M.; Colin, A.; Santenoise, P.; Fournier, M.; Constant, T. Terrestrial laser scanning for measuring the solid wood volume, including branches, of adult standing trees in the forest environment. Comput. Electron. Agric. 2012, 89, 86–93. [Google Scholar] [CrossRef]
- Llop, J.; Gil, E.; Llorens, J.; Miranda-Fuentes, A.; Gallart, M. Testing the Suitability of a Terrestrial 2D LiDAR Scanner for Canopy Characterization of Greenhouse Tomato Crops. Sensors 2016, 16, 1435. [Google Scholar] [CrossRef] [Green Version]
- Biskup, B.; Scharr, H.; Schurr, U.; Rascher, U. A stereo imaging system for measuring structural parameters of plant canopies. Plant Cell Environ. 2007, 30, 1299–1308. [Google Scholar] [CrossRef]
- Alenyà, G.; Dellen, B.; Torras, C. 3D modelling of leaves from color and ToF data for robotized plant measuring. In Proceedings of the IEEE International Conference on Robotics and Automation, Shanghai, China, 9–13 May 2011; pp. 3408–3414, ISBN 9781612843865. [Google Scholar]
- Chéné, Y.; Rousseau, D.; Lucidarme, P.; Bertheloot, J.; Caffier, V.; Morel, P.; Belin, É.; Chapeau-Blondeau, F. On the use of depth camera for 3D phenotyping of entire plants. Comput. Electron. Agric. 2012, 82, 122–127. [Google Scholar] [CrossRef]
- Santos, T.T.; Koenigkan, L.V.; Barbedo, J.G.; Rodrigues, G.C. 3d plant modeling: Localization, mapping and segmentation for plant phenotyping using a single hand-held camera. In Lecture Notes in Computer Science (Including Subseries Lecture Notes in Artificial Intelligence and Lecture Notes in Bioinformatics); Springer: Berlin/Heidelberg, Germany, 2015; pp. 247–263. ISBN 9783319162195. [Google Scholar]
- Behmann, J.; Steinrücken, J.; Plümer, L. Detection of early plant stress responses in hyperspectral images. ISPRS J. Photogramm. Remote Sens. 2014, 93, 98–111. [Google Scholar] [CrossRef]
- Roscher, R.; Behmann, J.; Mahlein, A.K.; Dupuis, J.; Kuhlmann, H.; Plümer, L. Detection of disease symptoms on hyperspectral 3D plant models. ISPRS Ann. Photogramm. Remote Sens. Spat. Inf. Sci. 2016, 3, 89–96. [Google Scholar] [CrossRef] [Green Version]
- Mohd Asaari, M.S.; Mishra, P.; Mertens, S.; Dhondt, S.; Inzé, D.; Wuyts, N.; Scheunders, P. Close-range hyperspectral image analysis for the early detection of stress responses in individual plants in a high-throughput phenotyping platform. ISPRS J. Photogramm. Remote Sens. 2018, 138, 121–138. [Google Scholar] [CrossRef]
- Carter, G.A. Responses of Leaf Spectral Reflectance to Plant Stress. Am. J. Bot. 1993, 80, 239. [Google Scholar] [CrossRef]
- Hoque, E.; Hutzler, P.J.S. Spectral blue-shift of red edge monitors damage class of beech trees. Remote Sens. Environ. 1992, 39, 81–84. [Google Scholar] [CrossRef]
- Rock, B.N.; Hoshizaki, T.; Miller, J.R. Comparison of in situ and airborne spectral measurements of the blue shift associated with forest decline. Remote Sens. Environ. 1988, 24, 109–127. [Google Scholar] [CrossRef]
- Horler, D.N.H. The Red Edge of Plant Leaf Reflectance; Taylor and Francis: London, UK, 1983. [Google Scholar]
- Maimaitiyiming, M.; Ghulam, A.; Bozzolo, A.; Wilkins, J.L.; Kwasniewski, M.T. Early Detection of Plant Physiological Responses to Different Levels of Water Stress Using Reflectance Spectroscopy. Remote Sens. 2017, 9, 745. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, E.; Heuvelink, E.; Gelder, A.d.; Stanghellini, C. New Non-invasive Tools for Early Plant Stress Detection. Procedia Environ. Sci. 2015, 29, 249–250. [Google Scholar] [CrossRef] [Green Version]
- Azzari, G.; Goulden, M.L.; Rusu, R.B. Rapid characterization of vegetation structure with a Microsoft Kinect sensor. Sensors 2013, 13, 2384–2398. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Zhang, Q. Three-Dimensional Reconstruction of a Dormant Tree Using RGB-D Cameras. In Proceedings of the American Society of Agricultural and Biological Engineers, Kansas City, MO, USA, 21–24 July 2013. [Google Scholar]
- Cignoni, P.; Callieri, M.; Corsini, M.; Dellepiane, M.; Ganovelli, F.; Ranzuglia, G. MeshLab: An Open-Source Mesh Processing Tool. In Proceedings of the Sixth Eurographics Italian Chapter Conference, Salerno, Italy, 2–4 July 2008; Scarano, V., Ed.; Eurographics Assoc: Aire-la-Ville, Switzerland, 2008; pp. 129–136, ISBN 9783905673685. [Google Scholar]
- Álvarez-Aragón, R.; Haro, R.; Benito, B.; Rodríguez-Navarro, A. Salt intolerance in Arabidopsis: Shoot and root sodium toxicity, and inhibition by sodium-plus-potassium overaccumulation. Planta 2016, 243, 97–114. [Google Scholar] [CrossRef] [PubMed]
- Wolf, O.; Munns, R.; Tonnet, M.L.; Jeschke, W.D. The role of the stem in the partitioning of Na+ and K+ in salt-treated barley. J. Exp. Bot. 1991, 42, 697–704. [Google Scholar] [CrossRef]
- Zhang, L.; Ma, H.; Chen, T.; Pen, J.; Yu, S.; Zhao, X. Morphological and physiological responses of cotton (Gossypium hirsutum L.) plants to salinity. PLoS ONE 2014, 9, e112807. [Google Scholar] [CrossRef] [PubMed] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moualeu-Ngangué, D.; Bötzl, M.; Stützel, H. First Form, Then Function: 3D Reconstruction of Cucumber Plants (Cucumis sativus L.) Allows Early Detection of Stress Effects through Leaf Dimensions. Remote Sens. 2022, 14, 1094. https://doi.org/10.3390/rs14051094
Moualeu-Ngangué D, Bötzl M, Stützel H. First Form, Then Function: 3D Reconstruction of Cucumber Plants (Cucumis sativus L.) Allows Early Detection of Stress Effects through Leaf Dimensions. Remote Sensing. 2022; 14(5):1094. https://doi.org/10.3390/rs14051094
Chicago/Turabian StyleMoualeu-Ngangué, Dany, Maria Bötzl, and Hartmut Stützel. 2022. "First Form, Then Function: 3D Reconstruction of Cucumber Plants (Cucumis sativus L.) Allows Early Detection of Stress Effects through Leaf Dimensions" Remote Sensing 14, no. 5: 1094. https://doi.org/10.3390/rs14051094
APA StyleMoualeu-Ngangué, D., Bötzl, M., & Stützel, H. (2022). First Form, Then Function: 3D Reconstruction of Cucumber Plants (Cucumis sativus L.) Allows Early Detection of Stress Effects through Leaf Dimensions. Remote Sensing, 14(5), 1094. https://doi.org/10.3390/rs14051094