
Dynamic Forecast of Desert Locust Presence Using Machine Learning with a Multivariate Time Lag Sliding Window Technique

 ,
,  ,
,  ,
,  ,
,  , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Locust Historical Field Survey Dataset
2.1.1. Study Area
2.1.2. Locust Dataset and Data Cleaning
2.1.3. Pseudo-Absence Generation
2.2. Extraction of Environmental Indicators of Desert Locust Presence
2.2.1. Multiple Indicators from Multisource Data
2.2.2. Extraction of Time Series of Dynamic Indicators
2.2.3. Extraction of Static Indicators
2.3. Time Lag Variable Importance Ranking for Dynamic Indicators
2.4. Model for Forecast Based on a Temporal Sliding Window
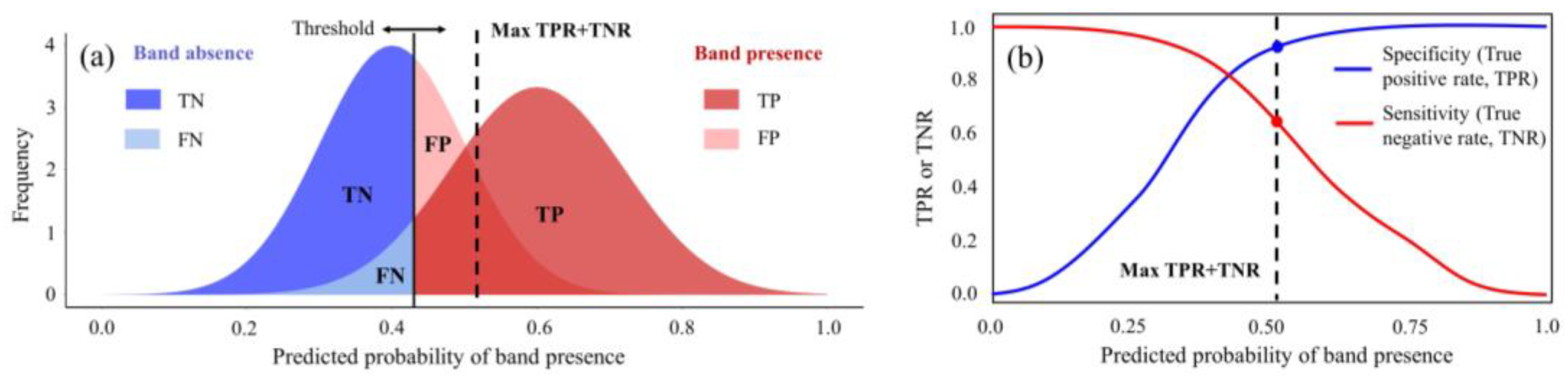
2.5. Model Evaluation and Accuracy Assessment
3. Results
3.1. Lagging Response of Hopper Band Occurrence to Environmental Drivers
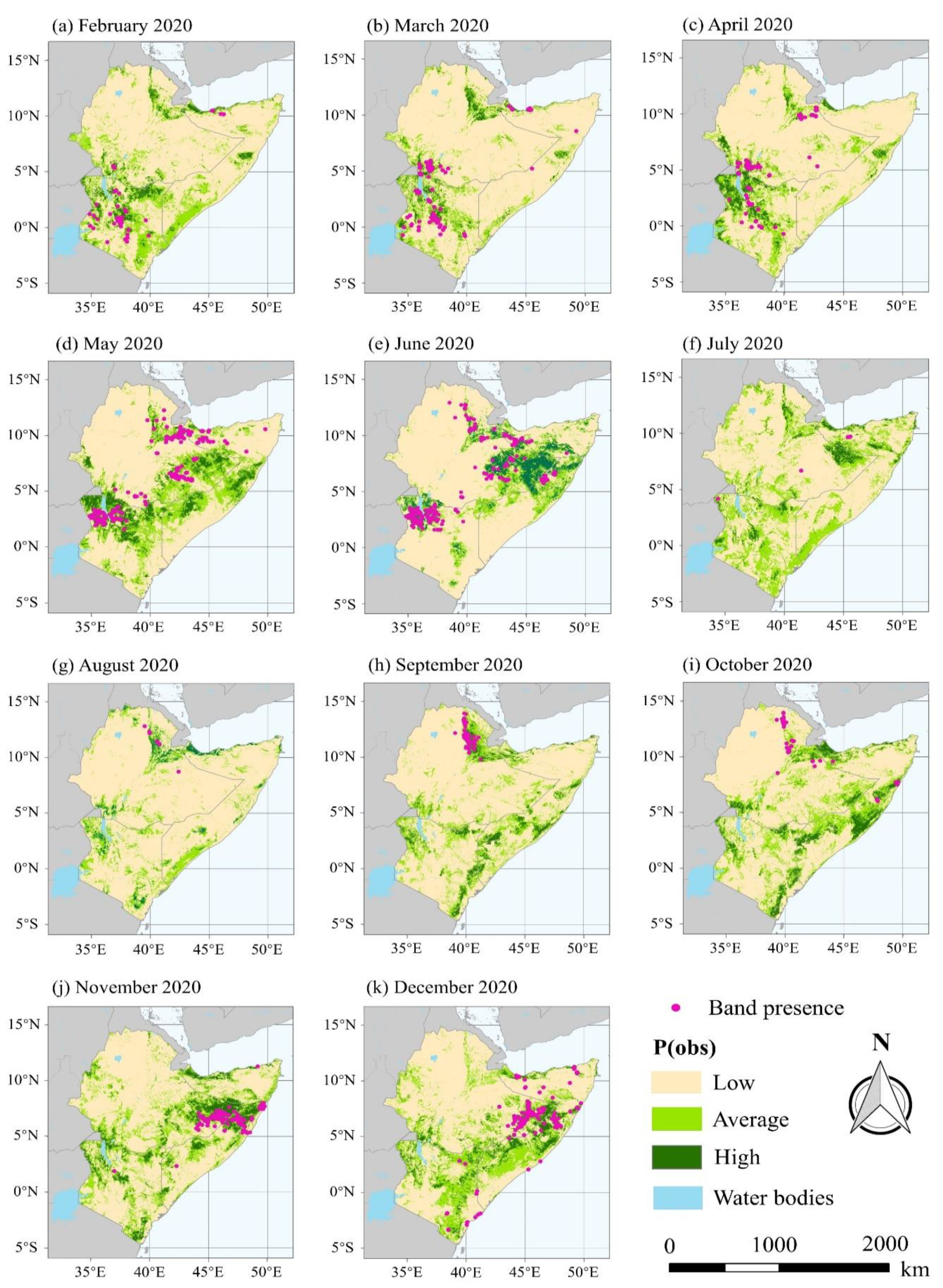
3.2. Dynamic Forecast of Hopper Band Presence in SEK
4. Discussion
4.1. Lag Effect of Environmental Drivers in Desert Locust Ecology
4.2. Strengths and Weaknesses of Environmental Drivers for Forecast Framework
4.3. Feasibility and Robustness of Forecast Model for Early Warning
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, W.; Dong, Y.; Zhao, L.; Geng, Y.; Ruan, C.; Zhang, B.; Sun, Z.; Zhang, H.; Ye, H.; Wang, K. Review of locust remote sensing monitoring and early warning (Chinese). J. Remote Sens. 2020, 24, 1270–1279. [Google Scholar]
- Salih, A.A.M.; Baraibar, M.; Mwangi, K.K.; Artan, G. Climate change and locust outbreak in East Africa. Nat. Clim. Change 2020, 10, 584–585. [Google Scholar] [CrossRef]
- Hemming, C.F.; Popov, G.B.; Roffey, J.; Waloff, Z. Characteristics of desert locust plague upsurges. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1979, 287, 375–386. [Google Scholar]
- Hassanali, A.; Bashir, M.O.; Njagi, P.G.N.; Ely, S.O. Desert locust gregarization: A conceptual kinetic model. J. Orthoptera Res. 2005, 14, 223–226. [Google Scholar] [CrossRef]
- Latchininsky, A.V. Locusts and remote sensing: A review. J. Appl. Remote Sens. 2013, 7, 75099. [Google Scholar] [CrossRef] [Green Version]
- Michel, L. Recent progress in Desert and Migratory Locust management in Africa. Are preventative actions possible ? J. Orthoptera Res. 2001, 10, 277–291. [Google Scholar]
- Lecoq, M. Desert locust threat to agricultural development and food security and FAO/international role in its control. In Proceedings of the Eighth Arab Congress of Plant Protection, El-Beida, Libya, 12–16 October 2003; Arab Society for Plant Protection: Beyrouth, Lebanon, 2003. [Google Scholar]
- Magor, J.I.; Lecoq, M.; Hunter, D.M. Preventive control and Desert Locust plagues. Crop. Prot. 2008, 27, 1527–1533. [Google Scholar] [CrossRef]
- Sword, G.A.; Lecoq, M.; Simpson, S.J. Phase polyphenism and preventative locust management. J. Insect Physiol. 2010, 56, 949–957. [Google Scholar] [CrossRef]
- Cressman, K. Role of remote sensing in desert locust early warning. J. Appl. Remote Sens. 2013, 7, 75098. [Google Scholar] [CrossRef]
- Zhang, L.; Lecoq, M.; Latchininsky, A.; Hunter, D. Locust and grasshopper management. Annu. Rev. Entomol. 2019, 64, 15–34. [Google Scholar] [CrossRef]
- Symmons, P.M.; Cressman, K. Desert Locust Guidelines: Biology and Behaviour; FAO: Rome, Italy, 2001. [Google Scholar]
- Dobson, H.M. Desert Locust Guidelines: Control; FAO: Rome, Italy, 2001. [Google Scholar]
- Symmons, P.M. A computer simulation of Desert Locust band target block detection and demarcation. Crop. Prot. 2004, 23, 183–190. [Google Scholar] [CrossRef]
- van Huis, A.; Cressman, K.; Magor, J.I. Preventing desert locust plagues: Optimizing management interventions. Entomol. Exp. Appl. 2007, 122, 191–214. [Google Scholar] [CrossRef]
- Gay, P.E.; Lecoq, M.; Piou, C. The limitations of locust preventive management faced with spatial uncertainty: Exploration with a multi-agent model. Pest. Manag. Sci. 2019, 76, 1094–1102. [Google Scholar] [CrossRef]
- Cressman, K. The use of new technologies in desert locust early warning. Outlooks Pest Manag. 2008, 19, 55–59. [Google Scholar] [CrossRef]
- Roffey, J.; Magor, J.I. Desert Locust Technical Series—Desert Locust Popualation Dynamics Parameters; Desert Locust Field Reserch Stations (FAO): Rome, Italy, 2003; p. 35. [Google Scholar]
- Wang, Y.; Wu, M.; Lin, P.; Wang, Y.; Chen, A.; Jiang, Y.; Zhai, B.; Chapman, J.W.; Hu, G. Plagues of desert locusts: Very low invasion risk to china. Insects 2020, 11, 628. [Google Scholar] [CrossRef] [PubMed]
- Amarjyothi, K.; Kumar, D.P.; Saikrishnan, K.C. Identification and tracking of locust swarms by indian doppler weather radar. IEEE Geosci. Remote Sens. Lett. 2022, 19, 1–4. [Google Scholar] [CrossRef]
- Ceccato, P.; Cressman, K.; Giannini, A.; Trzaska, S. The desert locust upsurge in West Africa (2003–2005): Information on the desert locust early warning system and the prospects for seasonal climate forecasting. Int. J. Pest Manag. 2007, 53, 7–13. [Google Scholar] [CrossRef]
- Kimathi, E.; Tonnang, H.E.Z.; Subramanian, S.; Cressman, K.; Abdel-Rahman, E.M.; Tesfayohannes, M.; Niassy, S.; Torto, B.; Dubois, T.; Tanga, C.M.; et al. Prediction of breeding regions for the desert locust Schistocerca gregaria in East Africa. Sci. Rep. UK 2020, 10, 11937. [Google Scholar] [CrossRef]
- Cressman, K.; Stefanski, R. Weather and Desert Locusts; World Meteorological Organization and Food and Agriculture Organization of the United Nations: Rome, Italy, 2016. [Google Scholar]
- Despland, E.; Collett, M.; Simpson, S.J. Small-scale processes in desert locust swarm formation: How vegetation patterns influence gregarization. Oikos 2000, 88, 652–662. [Google Scholar] [CrossRef]
- Culmsee, H. The habitat functions of vegetation in relation to the behaviour of the desert locust Schistocerca gregaria (Forskal) (Acrididae: Orthoptera)—A study in Mauritania (West Africa). Phytocoenologia 2002, 32, 645–664. [Google Scholar] [CrossRef]
- Gómez, D.; Salvador, P.; Sanz, J.; Casanova, C.; Taratiel, D.; Casanova, J.L. Desert locust detection using Earth observation satellite data in Mauritania. J. Arid Environ. 2019, 164, 29–37. [Google Scholar] [CrossRef]
- Escorihuela, M.J.; Merlin, O.; Stefan, V.; Moyano, G.; Eweys, O.A.; Zribi, M.; Kamara, S.; Benahi, A.S.; Ebbe, M.A.B.; Chihrane, J.; et al. SMOS based high resolution soil moisture estimates for desert locust preventive management. Remote Sens. Appl. Soc. Environ. 2018, 11, 140–150. [Google Scholar]
- Ellenburg, W.L.; Mishra, V.; Roberts, J.B.; Limaye, A.S.; Case, J.L.; Blankenship, C.B.; Cressman, K. Detecting desert locust breeding grounds: A Satellite-Assisted modeling approach. Remote Sens. 2021, 13, 1276. [Google Scholar] [CrossRef]
- Hielkema, J.U.; Roffey, J.; Tucker, C.J. Assessment of ecological conditions associated with the 1980/81 desert locust plague upsurge in West Africa using environmental satellite data. Int. J. Remote Sens. 1986, 7, 1609–1622. [Google Scholar] [CrossRef]
- Ceccato, P. Operational early warning system using SPOT-vegetation and Terra-MODIS to predict Desert Locust outbreaks. In Proceedings of the 2nd International Vegetation User Conference: 1998–2004: 6 Years of Operational Activities, European Commission, Antwerp, Belgium, 24–26 March 2004. [Google Scholar]
- Renier, C.; Waldner, F.; Jacques, D.; Babah Ebbe, M.; Cressman, K.; Defourny, P. A dynamic vegetation senescence indicator for Near-Real-Time desert locust habitat monitoring with MODIS. Remote Sens. 2015, 7, 7545–7570. [Google Scholar] [CrossRef] [Green Version]
- Tucker, C.J.; Hielkema, J.U.; Roffey, J. The potential of satellite remote sensing of ecological conditions for survey and forecasting desert-locust activity. Int. J. Remote Sens. 1985, 6, 127–138. [Google Scholar] [CrossRef]
- Pekel, J.; Ceccato, P.; Vancutsem, C.; Cressman, K.; Vanbogaert, E.; Defourny, P. Development and application of Multi-Temporal colorimetric transformation to monitor vegetation in the desert locust habitat. IEEE J. Stars 2011, 4, 318–326. [Google Scholar] [CrossRef]
- Waldner, F.; Ebbe, M.; Cressman, K.; Defourny, P. Operational monitoring of the desert locust habitat with earth observation: An assessment. ISPRS Int. J. Geo-Inf. 2015, 4, 2379–2400. [Google Scholar] [CrossRef] [Green Version]
- Piou, C.; Lebourgeois, V.; Benahi, A.S.; Bonnal, V.; Jaavar, M.E.H.; Lecoq, M.; Vassal, J. Coupling historical prospection data and a remotely-sensed vegetation index for the preventative control of Desert locusts. Basic Appl. Ecol. 2013, 14, 593–604. [Google Scholar] [CrossRef]
- Piou, C.; Gay, P.E.; Benahi, A.S.; Babah Ebbe, M.A.O.; Chihrane, J.; Ghaout, S.; Cisse, S.; Diakite, F.; Lazar, M.; Cressman, K.; et al. Soil moisture from remote sensing to forecast desert locust presence. J. Appl. Ecol. 2019, 56, 966–975. [Google Scholar] [CrossRef]
- Gómez, D.; Salvador, P.; Sanz, J.; Casanova, C.; Taratiel, D.; Casanova, J.L. Machine learning approach to locate desert locust breeding areas based on ESA CCI soil moisture. J. Appl. Remote Sens. 2018, 12, 036011. [Google Scholar] [CrossRef] [Green Version]
- Gómez, D.; Salvador, P.; Sanz, J.; Casanova, J.L. Modelling desert locust presences using 32-year soil moisture data on a large-scale. Ecol. Indic. 2020, 117, 106655. [Google Scholar] [CrossRef]
- Cisse, S.; Ghaout, S.; Mazih, A.; Babah Ebbe, M.A.O.; Benahi, A.S.; Piou, C. Effect of vegetation on density thresholds of adult desert locust gregarization from survey data in Mauritania. Entomol. Exp. Appl. 2013, 149, 159–165. [Google Scholar] [CrossRef]
- Lazar, M.; Piou, C.; Doumandji-Mitiche, B.; Lecoq, M. Importance of solitarious desert locust population dynamics: Lessons from historical survey data in Algeria. Entomol. Exp. Appl. 2016, 161, 168–180. [Google Scholar] [CrossRef] [Green Version]
- Huang, J. Remote sensing of locust and grasshopper plague in China: A review. In Proceedings of the 5th International Conference on Agro-Geoinformatics, Tianjin, China, 18–20 July 2016. [Google Scholar]
- Amichay, G.; Ariel, G.; Ayali, A. The effect of changing topography on the coordinated marching of locust nymphs. Peer J. 2016, 4, e2742. [Google Scholar] [CrossRef] [Green Version]
- Steedman, A. Locust Handbook, 2nd ed.; Overseas Development Natural Resource Institute: London, UK, 1988. [Google Scholar]
- Sivanpillai, R.; Latchininsky, A.V.; Driese, K.L.; Kambulin, V.E. Mapping locust habitats in River Ili Delta, Kazakhstan, using LANDSAT imagery. Agric. Ecosyst. Environ. 2006, 117, 128–134. [Google Scholar] [CrossRef]
- Zhao, L.; Huang, W.; Chen, J.; Dong, Y.; Ren, B.; Geng, Y. Land use/cover changes in the Oriental migratory locust area of China: Implications for ecological control and monitoring of locust area. Agric. Ecosyst. Environ. 2020, 303, 107110. [Google Scholar] [CrossRef]
- Shi, Y.; Huang, W.; Dong, Y.; Peng, D.; Zheng, Q.; Yang, P. The influence of landscape’s dynamics on the Oriental Migratory Locust habitat change based on the time-series satellite data. J. Environ. Manag. 2018, 218, 280–290. [Google Scholar] [CrossRef]
- Geng, Y.; Zhao, L.; Dong, Y.; Huang, W.; Shi, Y.; Ren, Y.; Ren, B. Migratory locust habitat analysis with PB-AHP model using Time-Series satellite images. IEEE Access 2020, 8, 166813–166823. [Google Scholar] [CrossRef]
- Tierney, J.E.; Ummenhofer, C.C.; DeMenocal, P.B. Past and future rainfall in the Horn of Africa. Sci. Adv. 2015, 1, e1500682. [Google Scholar] [CrossRef] [Green Version]
- FAO-DLIS; WCS. Seasonal precipitation predictions in Desert Locust spring/summer/winter breeding areas (June–November 2021); FAO: Rome, Italy, 2021. [Google Scholar]
- Kalakkal, J.; Singh, A. Desert Locusts’ Upsurges: A Harbinger of Emerging Climate Change-Induced Crises? The United Nations Environment Programme (UNEP): Nairobi, Kenya, 2021. [Google Scholar]
- Stone, M. A Plague of Locusts has Descended on East Africa. Climate Change May be to Blame; National Geographic: Washington, DC, USA, 2020; Volume 2021. [Google Scholar]
- Aiello-Lammens, M.E.; Boria, R.A.; Radosavljevic, A.; Vilela, B.; Anderson, R.P. SpThin: An R package for spatial thinning of species occurrence records for use in ecological niche models. Ecography 2015, 38, 541–545. [Google Scholar] [CrossRef]
- Bedia, J.; Iturbide, M.; Gutiérrez, J. Tackling Uncertainties of Species Distribution Model Projections with Package mopa. R J. 2018, 10, 122–135. [Google Scholar]
- Rew, J.; Cho, Y.; Hwang, E. A robust prediction model for species distribution using bagging ensembles with deep neural networks. Remote Sens. 2021, 13, 1495. [Google Scholar] [CrossRef]
- Chefaoui, R.M.; Lobo, J.M. Assessing the effects of pseudo-absences on predictive distribution model performance. Ecol. Model. 2008, 210, 478–486. [Google Scholar] [CrossRef]
- Graham, C.H.; Hijmans, R.J. A comparison of methods for mapping species ranges and species richness. Glob. Ecol. Biogeogr. 2006, 15, 578–587. [Google Scholar] [CrossRef]
- Iturbide, M.; Bedia, J.; Herrera, S.; Del Hierro, O.; Pinto, M.; Gutiérrez, J.M. A framework for species distribution modelling with improved pseudo-absence generation. Ecol. Model. 2015, 312, 166–174. [Google Scholar] [CrossRef] [Green Version]
- Dinku, T.; Ceccato, P.; Cressman, K.; Connor, S.J. Evaluating detection skills of satellite rainfall estimates over desert locust recession regions. J. Appl. Meteorol. Clim. 2010, 49, 1322–1332. [Google Scholar] [CrossRef]
- Dinku, T.; Funk, C.; Peterson, P.; Maidment, R.; Tadesse, T.; Gadain, H.; Ceccato, P. Validation of the CHIRPS satellite rainfall estimates over eastern Africa. Q. J. R. Meteorol. Soc. 2018, 144, 292–312. [Google Scholar] [CrossRef] [Green Version]
- Funk, C.; Peterson, P.; Landsfeld, M.; Pedreros, D.; Verdin, J.; Shukla, S.; Husak, G.; Rowland, J.; Harrison, L.; Hoell, A.; et al. The climate hazards infrared precipitation with stations—A new environmental record for monitoring extremes. Sci. Data 2015, 2, 150066. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.V.; Peters-Lidard, C.D.; Tian, Y.; Houser, P.R.; Geiger, J.; Olden, S.; Lighty, L.; Eastman, J.L.; Doty, B.; Dirmeyer, P. Land information system: An interoperable framework for high resolution land surface modeling. Environ. Model. Softw. 2006, 21, 1402–1415. [Google Scholar] [CrossRef]
- McNally, A.; Arsenault, K.; Kumar, S.; Shukla, S.; Peterson, P.; Wang, S.; Funk, C.; Peters-Lidard, C.D.; Verdin, J.P. A land data assimilation system for sub-Saharan Africa food and water security applications. Sci. Data 2017, 4, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Case, J.L.; Mungai, J.; Sakwa, V.; Zavodsky, B.T.; Srikishen, J.; Limaye, A.S.; Blankenship, C.B. Transitioning Enhanced Land Surface Initialization and Model Verification Capabilities to the Kenya Meteorological Service; American Meteorological Society Fall Meeting: New Orleans, LA, USA, 2016. [Google Scholar]
- Hengl, T.; Mendes De Jesus, J.; Heuvelink, G.B.M.; Ruiperez Gonzalez, M.; Kilibarda, M.; Blagotić, A.; Shangguan, W.; Wright, M.N.; Geng, X.; Bauer-Marschallinger, B.; et al. SoilGrids250m: Global gridded soil information based on machine learning. PLoS ONE 2017, 12, e169748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchhorn, M.; Lesiv, M.; Tsendbazar, N.; Herold, M.; Bertels, L.; Smets, B. Copernicus global land cover layers—Collection 2. Remote Sens. 2020, 12, 1044. [Google Scholar] [CrossRef] [Green Version]
- NASA JPL. NASADEM Merged DEM Global 1 arc second V001. 2020, distributed by NASA EOSDIS Land Processes DAAC. Available online: https://doi.org/10.5067/MEaSUREs/NASADEM/NASADEM_HGT.001 (accessed on 20 December 2021).
- Evans, J.S.; Murphy, M.A.; Holden, Z.A.; Cushman, S.A. Modeling Species Distribution and Change Using Random Forest; Springer: New York, NY, USA, 2011; pp. 139–159. [Google Scholar]
- Pearce, J.; Ferrier, S. Evaluating the predictive performance of habitat models developed using logistic regression. Ecol. Model. 2000, 133, 225–245. [Google Scholar] [CrossRef] [Green Version]
- Keating, K.A.; Cherry, S. Use and interpretation of logistic regression in habitat-selection studies. J. Wildl. Manag. 2004, 68, 774–789. [Google Scholar] [CrossRef]
- Azen, R.; Traxel, N. Using dominance analysis to determine predictor importance in logistic regression. J. Educ. Behav. Stat. 2009, 34, 319–347. [Google Scholar] [CrossRef]
- Luo, W.; Azen, R. Determining predictor importance in hierarchical linear models using dominance analysis. J. Educ. Behav. Stat. 2013, 38, 3–31. [Google Scholar] [CrossRef]
- Soares, F.M.C.D. Modelling the Distribution of São Tomé Bird Species: Ecological Determinants and Conservation Prioritization. Doctoral dissertation, University of Lisbon, Lisbon, Portugal, 2017. [Google Scholar]
- Han, H.; Guo, X.; Yu, H. Variable selection using mean decrease accuracy and mean decrease gini based on random forest. In Proceedings of the 2016 7th IEEE International Conference on Software Engineering and Service Science (ICSESS), Beijing, China, 26–28 August 2016. [Google Scholar]
- Zhang, J.; Li, S. A Review of Machine Learning Based Species’ Distribution Modelling. In Proceedings of the 2017 International Conference on Industrial Informatics-Computing Technology, Intelligent Technology, Industrial Information Integration (ICIICII). Wuhan, China, 2–3 December 2017. [Google Scholar]
- Chang, T.; Hansen, A.J.; Piekielek, N. Patterns and variability of projected bioclimatic habitat for pinus albicaulis in the greater yellowstone area. PLoS ONE 2014, 9, e111669. [Google Scholar] [CrossRef] [Green Version]
- Hong, C.S.; Oh, T.G. TPR-TNR plot for confusion matrix. Commun. Stat. Appl. Methods 2021, 28, 161–169. [Google Scholar] [CrossRef]
- Sterr, U. Analysis of 2019 Kaggle ML & DS Survey. Available online: https://uwesterr.github.io/KaggleMlSurvey2019/ (accessed on 20 December 2021).
- Magor, J.I. Rainfall as a factor in the geographical distribution of the desert locust breeding areas, with particular reference to the summer breeding area of India and Pakistan. Doctoral dissertation, University of Edinburgh, Edinburgh, UK, 1963. [Google Scholar]
- Roffey, J.; Popov, G.; Hemming, C.F. Outbreaks and recession populations of the desert locust Schistocerca gregaria (Forsk.). Bull. Entomol. Res. 1970, 59, 675–680. [Google Scholar] [CrossRef]
- Bennett, L.V. The development and termination of the 1968 plague of the Desert locust, Schistocerca gregaria (Forskål) (Orthoptera, Acrididae). Bull. Entomol. Res. 1976, 66, 511–552. [Google Scholar] [CrossRef]
- Dempster, J.P. The population dynamics of grasshoppers and locusts. Biol. Rev. 1963, 38, 490–529. [Google Scholar] [CrossRef]
- Latchininsky, A.V.; Sivanpillai, R. Locust Habitat Monitoring and Risk Assessment Using Remote Sensing and GIS Technologies; Springer: Dordrecht, The Netherlands, 2010; pp. 163–188. [Google Scholar]
- Tratalos, J.A.; Cheke, R.A.; Healey, R.G.; Stenseth, N.C. Desert Locust Populations, Rainfall and Climate Change: Insights from Phenomenological Models Using Gridded Monthly data. Clim. Res. 2010, 43, 229–239. [Google Scholar] [CrossRef]
- Hunter, J.P. Egg Development in the Desert Locust (Schistocerca gregaria Forsk.) in Relation to the Availability of Water; Wiley Online Library: Hoboken, NJ, USA, 1964; pp. 25–33. [Google Scholar]
- Popov, G.B. Ecological Studies on Oviposition by Swarms of the Desert Locust (Schistocerca gregaria Forskal) in Eastern Africa. Anti-Locust Research Centre: London, UK, 1958. [Google Scholar]
- Symmons, P.M. A model to predict the incubation and nymphal development periods of the desert locust, Schistocerca gregaria (Orthoptera: Acrididae) 1. Bull. Entomol. Res. 1992, 82, 517–520. [Google Scholar]
- Wardhaugh, K.; Ashour, Y.; Ibrahim, A.O.; Khan, A.M.; Bassonbol, M. Experiments on the incubation and hopper development periods of the desert locust (Schistocerca gregaria Forskál) in Saudi Arabia. Lond. Anti locust Res. Cent Anti locust Bull. 1969, 46, 35–38. [Google Scholar]
- Nishide, Y.; Suzuki, T.; Tanaka, S. Synchrony in the hatching of eggs in the desert locust Schistocerca gregaria (Orthoptera: Acrididae): Egg condition influences hatching time in the laboratory and under simulated field temperatures. Appl. Entomol. Zool. 2017, 52, 599–604. [Google Scholar] [CrossRef]
- Mohanty, B.P.; Cosh, M.H.; Lakshmi, V.; Montzka, C. Soil moisture remote sensing: State-of-the-science. Vadose Zone J. 2017, 16, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Deveson, E.D. Satellite normalized difference vegetation index data used in managing Australian plague locusts. J. Appl. Remote Sens. 2013, 7, 75096. [Google Scholar] [CrossRef] [Green Version]
- Gómez, D.; Salvador, P.; Sanz, J.; Rodrigo, J.F.; Gil, J.; Casanova, J.L. Prediction of desert locust breeding areas using machine learning methods and SMOS (MIR_SMNRT2) Near Real Time product. J. Arid Environ. 2021, 194, 104599. [Google Scholar] [CrossRef]
- Roffey, J.; Popov, G. Environmental and behavioural processes in a desert locust outbreak. Nature 1968, 219, 446–450. [Google Scholar] [CrossRef]
- Veran, S.; Simpson, S.J.; Sword, G.A.; Deveson, E.; Piry, S.; Hines, J.E.; Berthier, K. Modeling spatiotemporal dynamics of outbreaking species: Influence of environment and migration in a locust. Ecology 2015, 96, 737–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO. Desert Locust Bulletin—General Situation during August 2020 and Forecast until Mid-October 2020; FAO: Rome, Italy, 2020. [Google Scholar]
- FAO. Desert Locust Bulletin—General Situation during September 2020 and Forecast until Mid-November 2020; FAO: Rome, Italy, 2020. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Indicators | Data Source | Spatial Resolution | Temporal Resolution | Time Range | Description | |
|---|---|---|---|---|---|---|
| Dynamic indicators | PREC | CHIRPS 1 | 0.05° (~5 km) | daily | 1981–2021 | Precipitation (mm/day) |
| SM | ERA5-Land 2 | 0.1° (~10 km) | hourly | 1981–2021 | Volumetric soil water layer 2 (7–28 cm below the surface) (m3/m3) | |
| SERVIR 3 | 0.03° (~3 km) | daily | 2020–2021 | Volumetric soil moisture (0–10 cm below the surface) (%) | ||
| NDVI | MOD09GA | 1 km | daily | 2000–2021 | Red and NIR band normalization | |
| LST | MOD11A1 | 1 km | daily | 2000–2021 | Units converted from K to °C | |
| Static indicators | SND | ISRIC 4 SoilGrids [64] | 250 m | 3-year | 2014, 2017, 2021 | Sand in depth of 5–15 cm (g/kg) |
| CLY | Clay content in depth of 5–15 cm (g/kg) | |||||
| SLT | Silt in depth of 5–15 cm (g/kg) | |||||
| CRF | Coarse fragments in depth of 5–15 cm (cm3/dm3) | |||||
| LULC | CGLS-LC100 5 [65] | 100 m | 5-year | 2015–2019 | Discrete classification | |
| DEM | NASADEM [66] | 30 m | - | 2020 | Elevation (m) | |
| Date | Evaluation Metrics | ||||
|---|---|---|---|---|---|
| Accuracy (%) | Sensitivity | Specificity | ROC-AUC | F-Score | |
| February 2020 | 74.44 | 0.6047 | 0.8759 | 0.7403 | 0.7792 |
| March 2020 | 80.15 | 0.6934 | 0.9329 | 0.8131 | 0.7930 |
| April 2020 | 82.59 | 0.7264 | 0.9002 | 0.8133 | 0.7811 |
| May 2020 | 88.68 | 0.8886 | 0.8814 | 0.8850 | 0.9218 |
| June 2020 | 85.31 | 0.8971 | 0.6667 | 0.7819 | 0.9081 |
| July 2020 | 70.00 | 0.6167 | 0.7714 | 0.6940 | 0.6549 |
| August 2020 | 76.99 | 0.6238 | 0.9412 | 0.7825 | 0.7453 |
| September 2020 | 79.81 | 0.6314 | 0.9203 | 0.7759 | 0.7258 |
| October 2020 | 66.77 | 0.5988 | 0.7419 | 0.6704 | 0.6515 |
| November 2020 | 73.41 | 0.7500 | 0.7116 | 0.7308 | 0.7673 |
| December 2020 | 73.95 | 0.7087 | 0.7816 | 0.7451 | 0.7586 |
| Average | 77.46 | 0.7036 | 0.8296 | 0.7666 | 0.7715 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, R.; Huang, W.; Dong, Y.; Zhao, L.; Zhang, B.; Ma, H.; Geng, Y.; Ruan, C.; Xing, N.; Chen, X.; et al. Dynamic Forecast of Desert Locust Presence Using Machine Learning with a Multivariate Time Lag Sliding Window Technique. Remote Sens. 2022, 14, 747. https://doi.org/10.3390/rs14030747
Sun R, Huang W, Dong Y, Zhao L, Zhang B, Ma H, Geng Y, Ruan C, Xing N, Chen X, et al. Dynamic Forecast of Desert Locust Presence Using Machine Learning with a Multivariate Time Lag Sliding Window Technique. Remote Sensing. 2022; 14(3):747. https://doi.org/10.3390/rs14030747
Chicago/Turabian StyleSun, Ruiqi, Wenjiang Huang, Yingying Dong, Longlong Zhao, Biyao Zhang, Huiqin Ma, Yun Geng, Chao Ruan, Naichen Xing, Xidong Chen, and et al. 2022. "Dynamic Forecast of Desert Locust Presence Using Machine Learning with a Multivariate Time Lag Sliding Window Technique" Remote Sensing 14, no. 3: 747. https://doi.org/10.3390/rs14030747
APA StyleSun, R., Huang, W., Dong, Y., Zhao, L., Zhang, B., Ma, H., Geng, Y., Ruan, C., Xing, N., Chen, X., & Li, X. (2022). Dynamic Forecast of Desert Locust Presence Using Machine Learning with a Multivariate Time Lag Sliding Window Technique. Remote Sensing, 14(3), 747. https://doi.org/10.3390/rs14030747