
A UAS and Machine Learning Classification Approach to Suitability Prediction of Expanding Natural Habitats for Endangered Flora Species





Abstract
1. Introduction
2. Materials and Methods
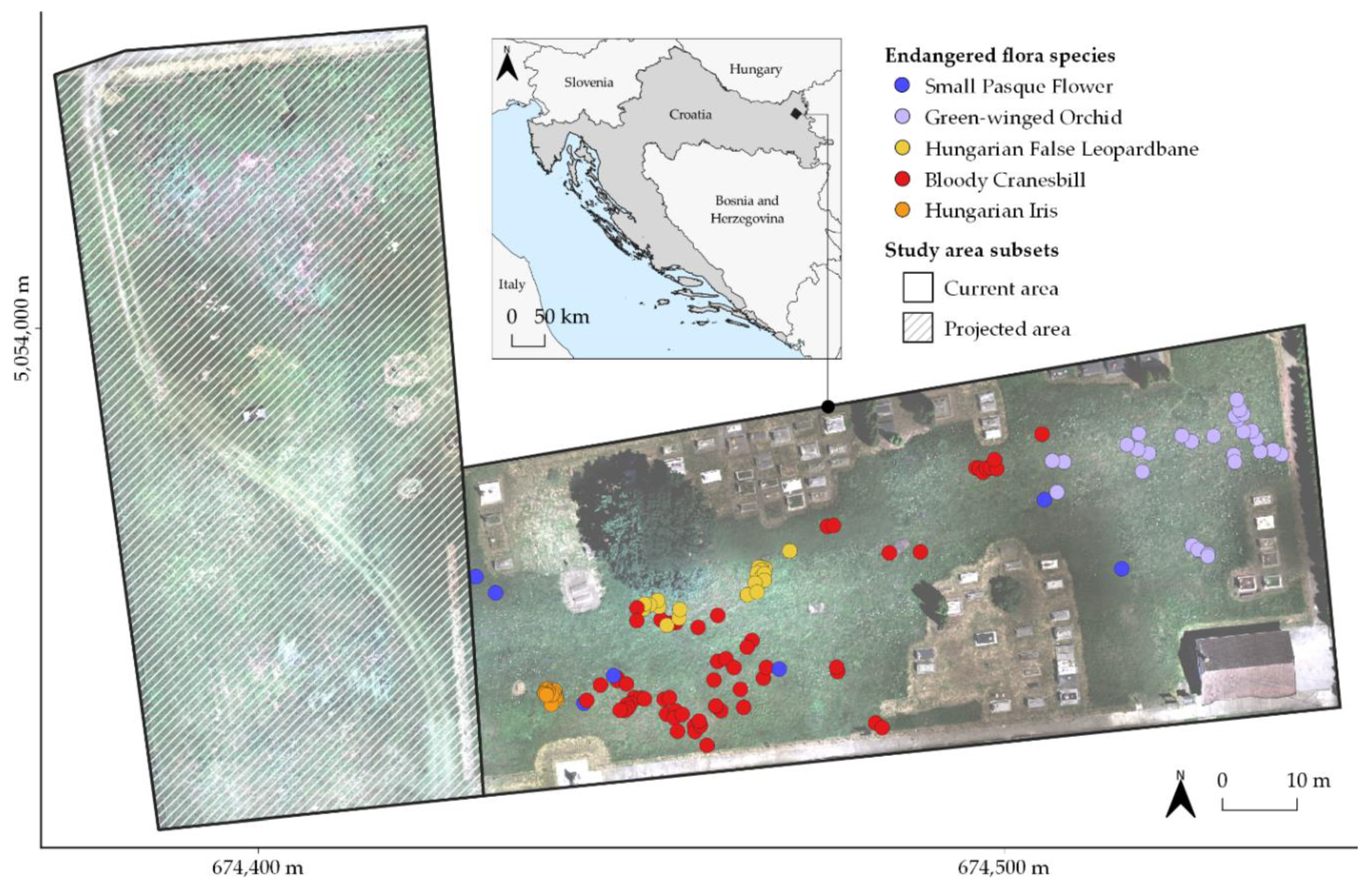
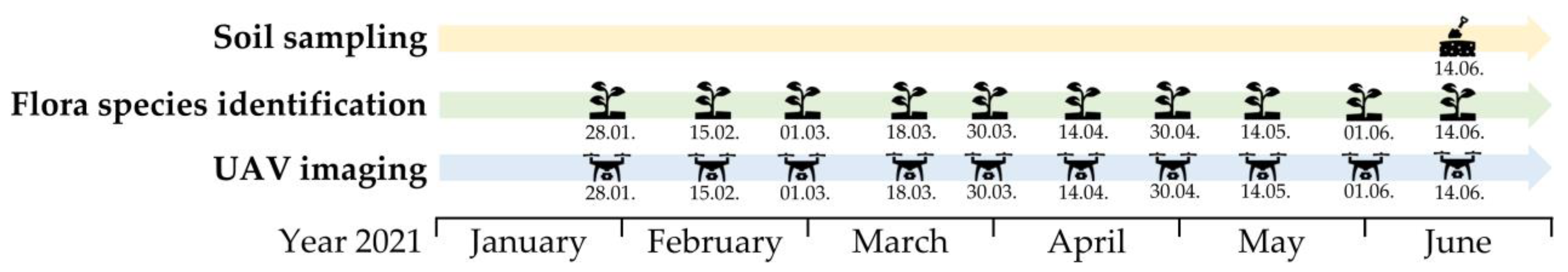
2.1. Study Area and Fieldwork
2.2. Spatial Modeling of Environmental Criteria
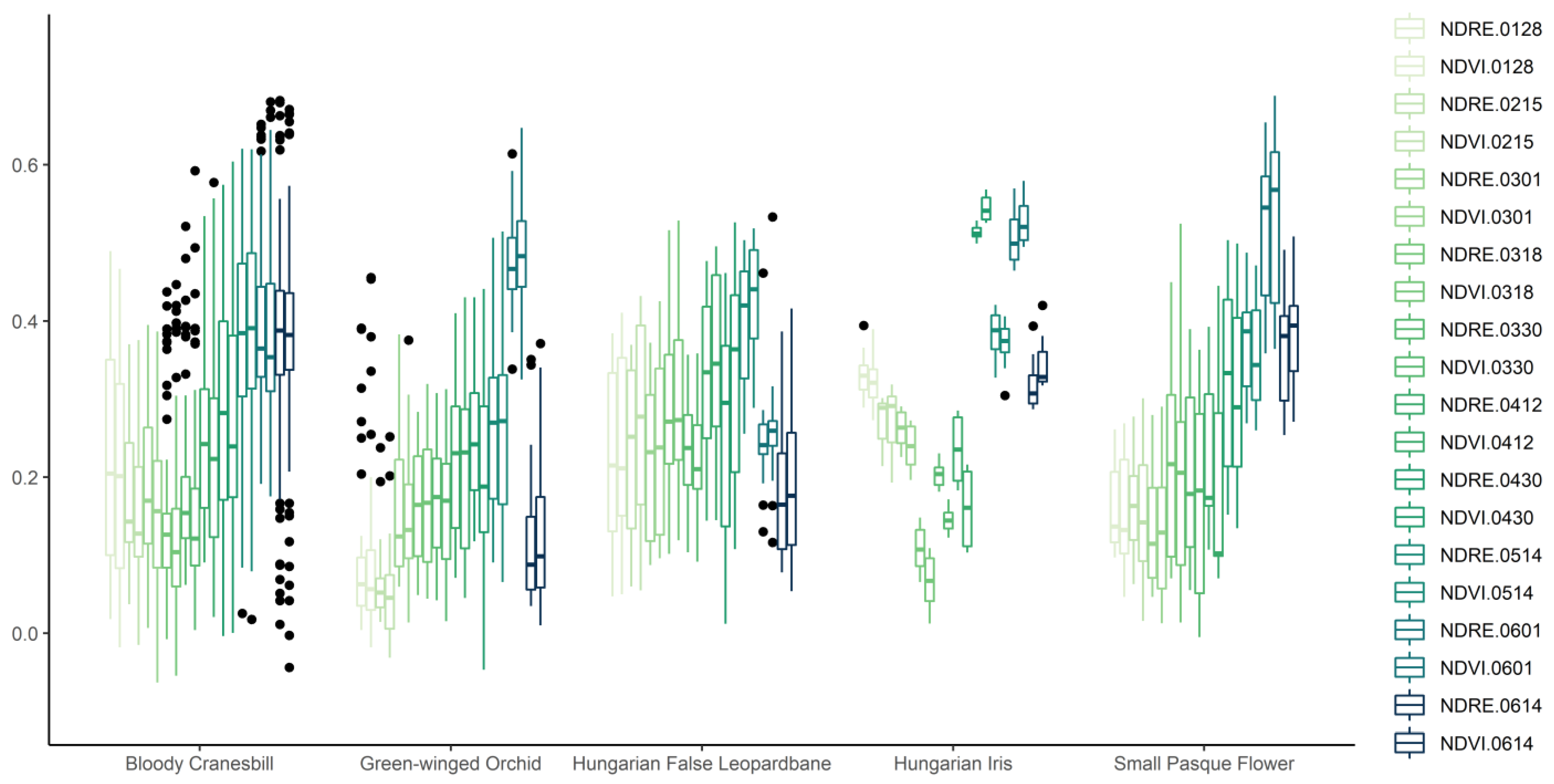
2.2.1. Vegetation Criteria
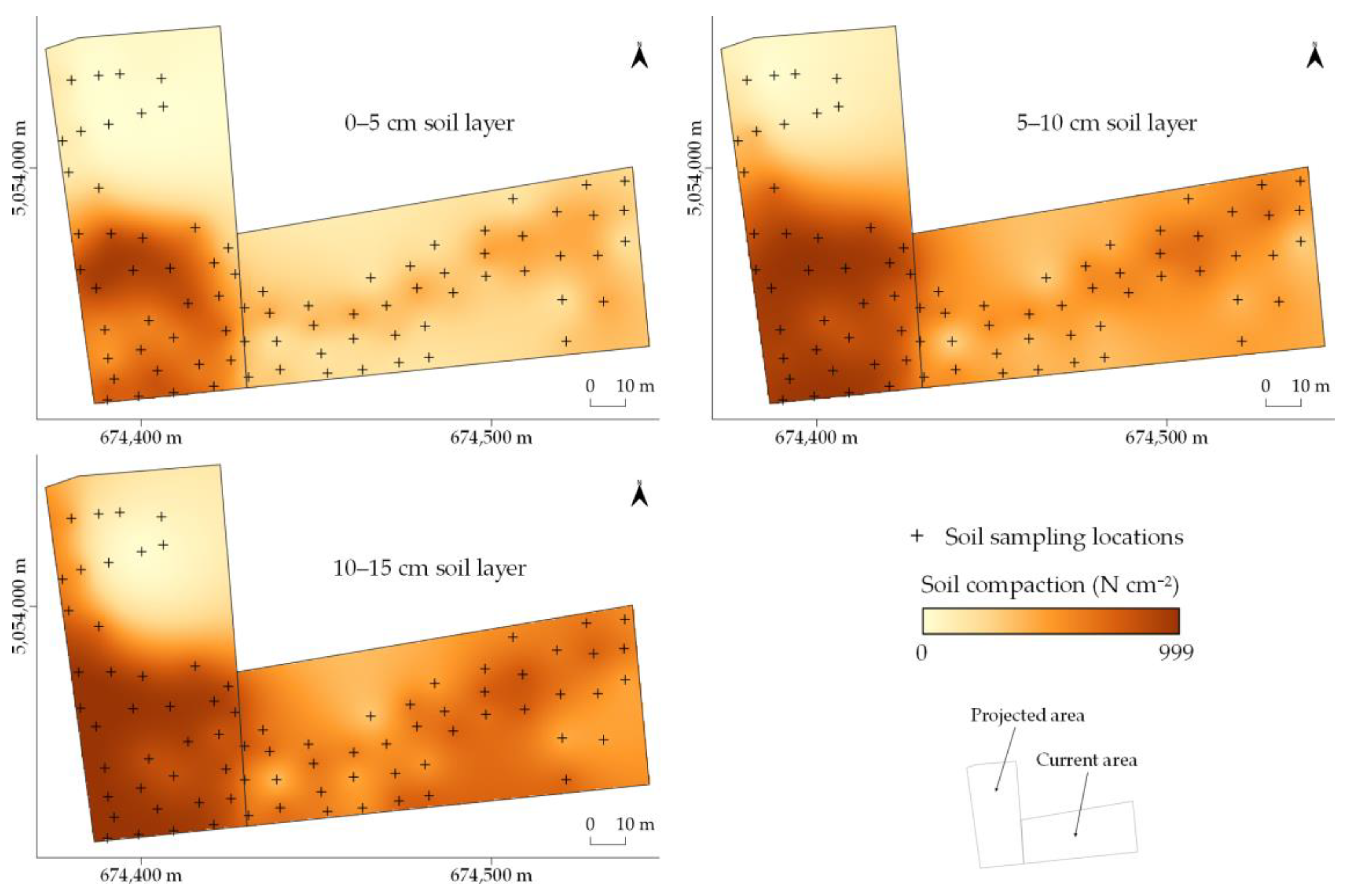
2.2.2. Soil Criteria
2.2.3. Topographic Criteria
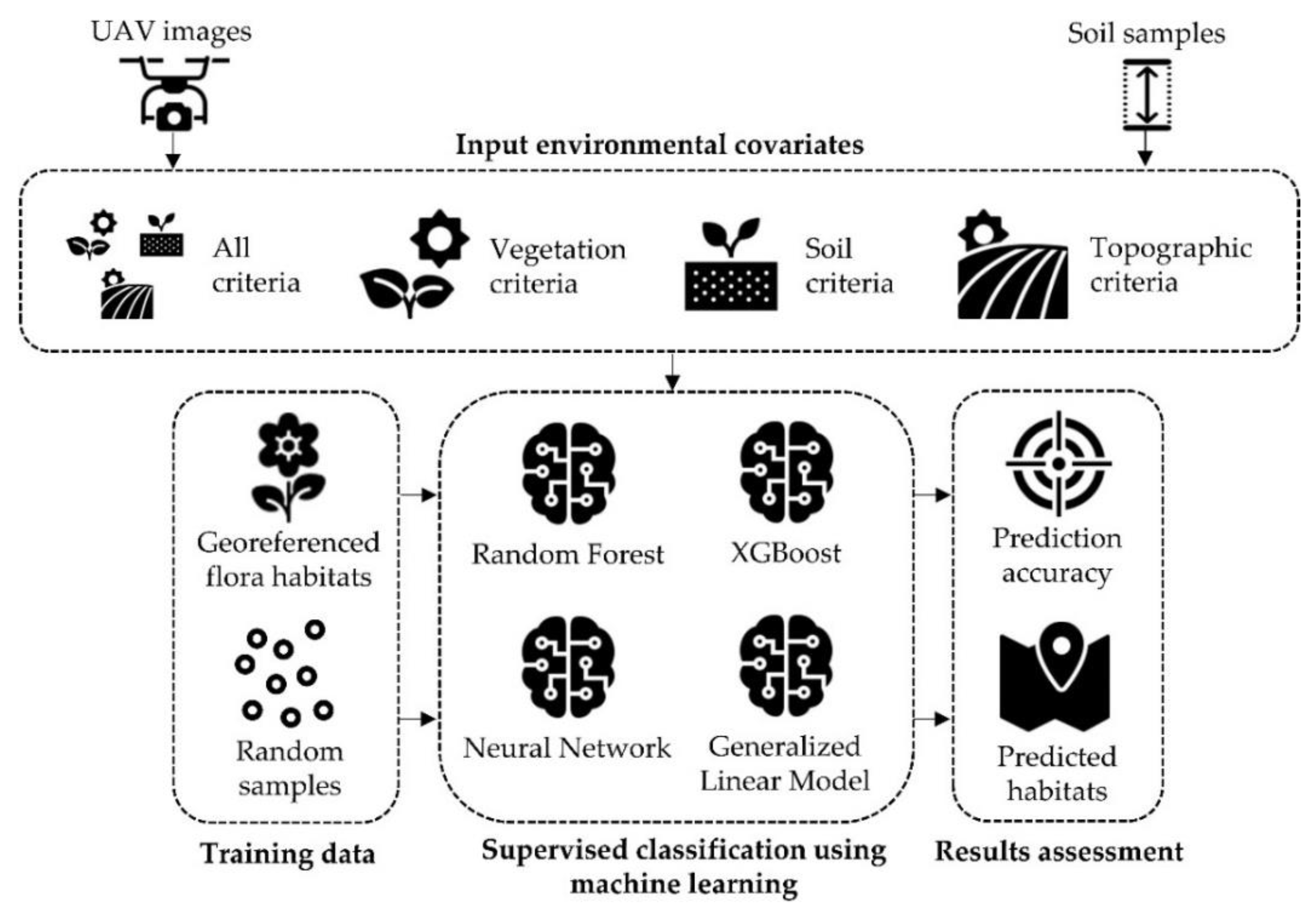
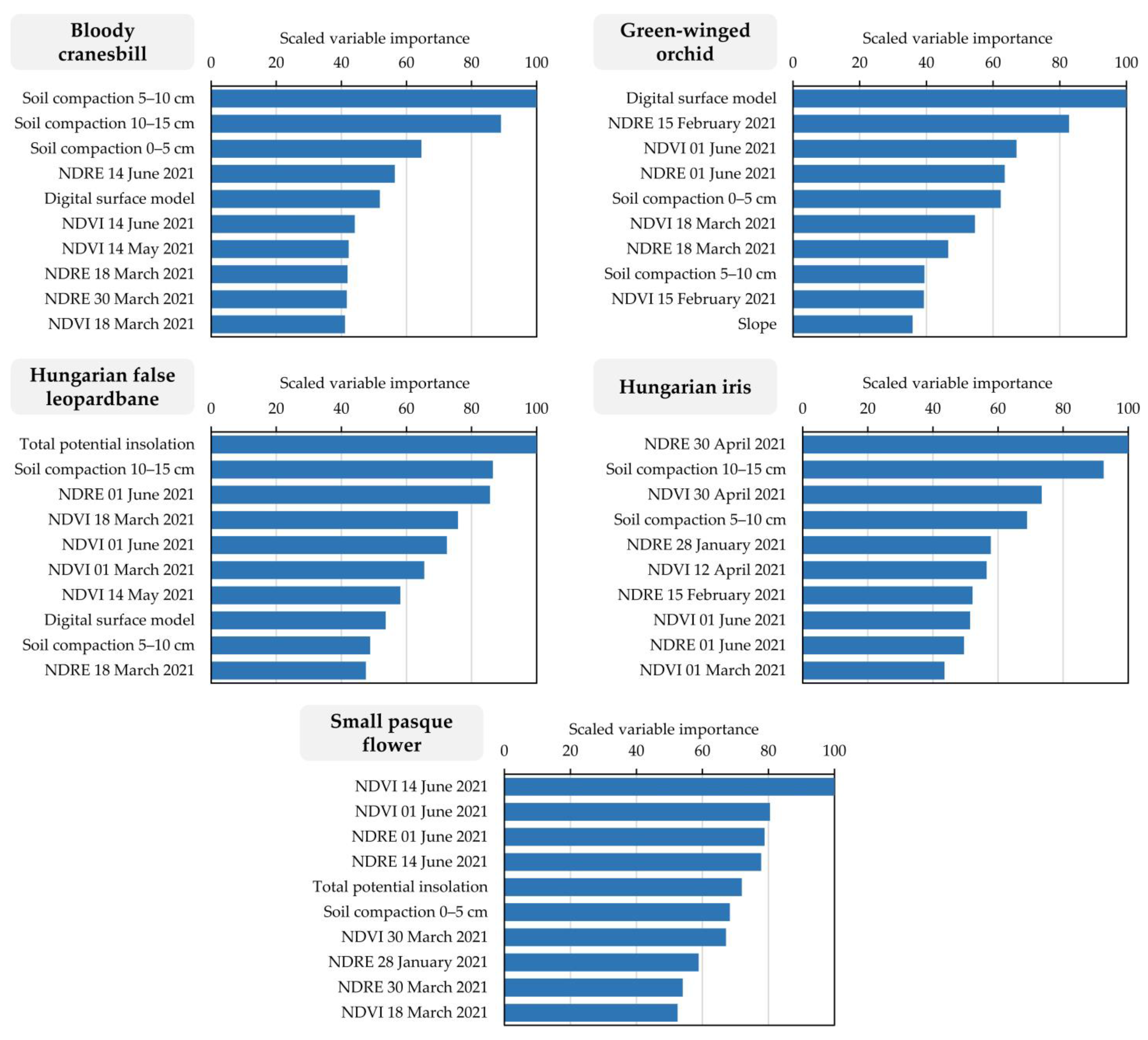
2.3. Habitat Suitability Prediction Using Machine Learning
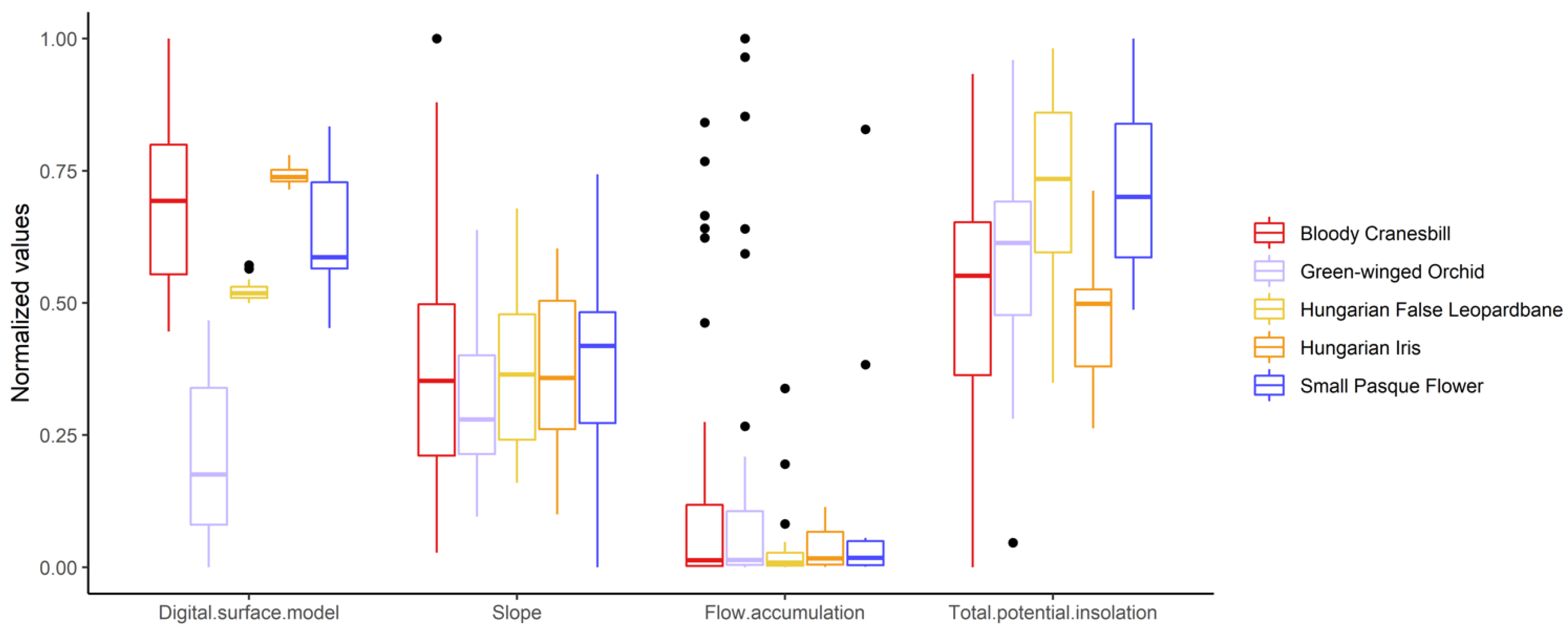
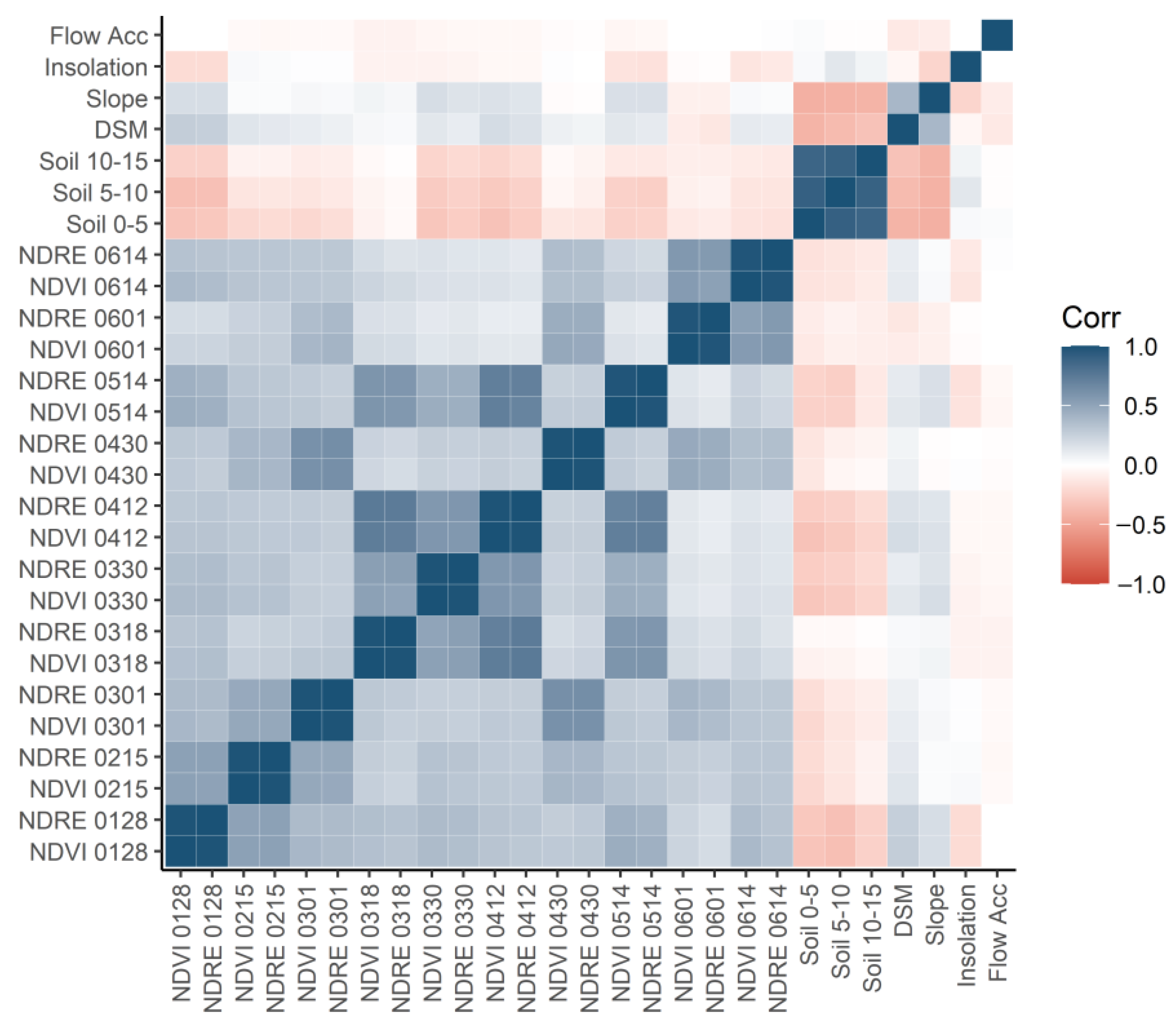
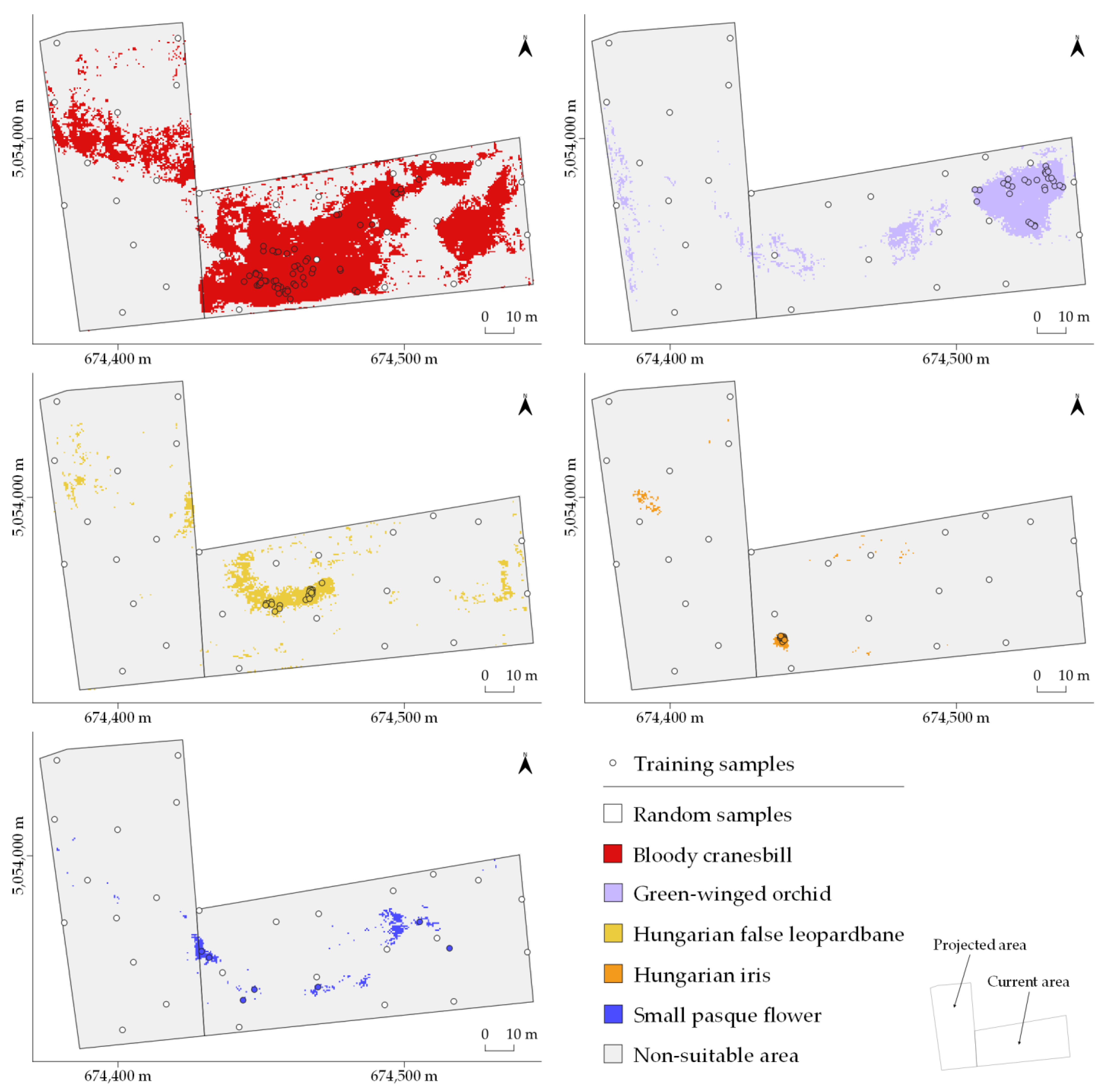
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
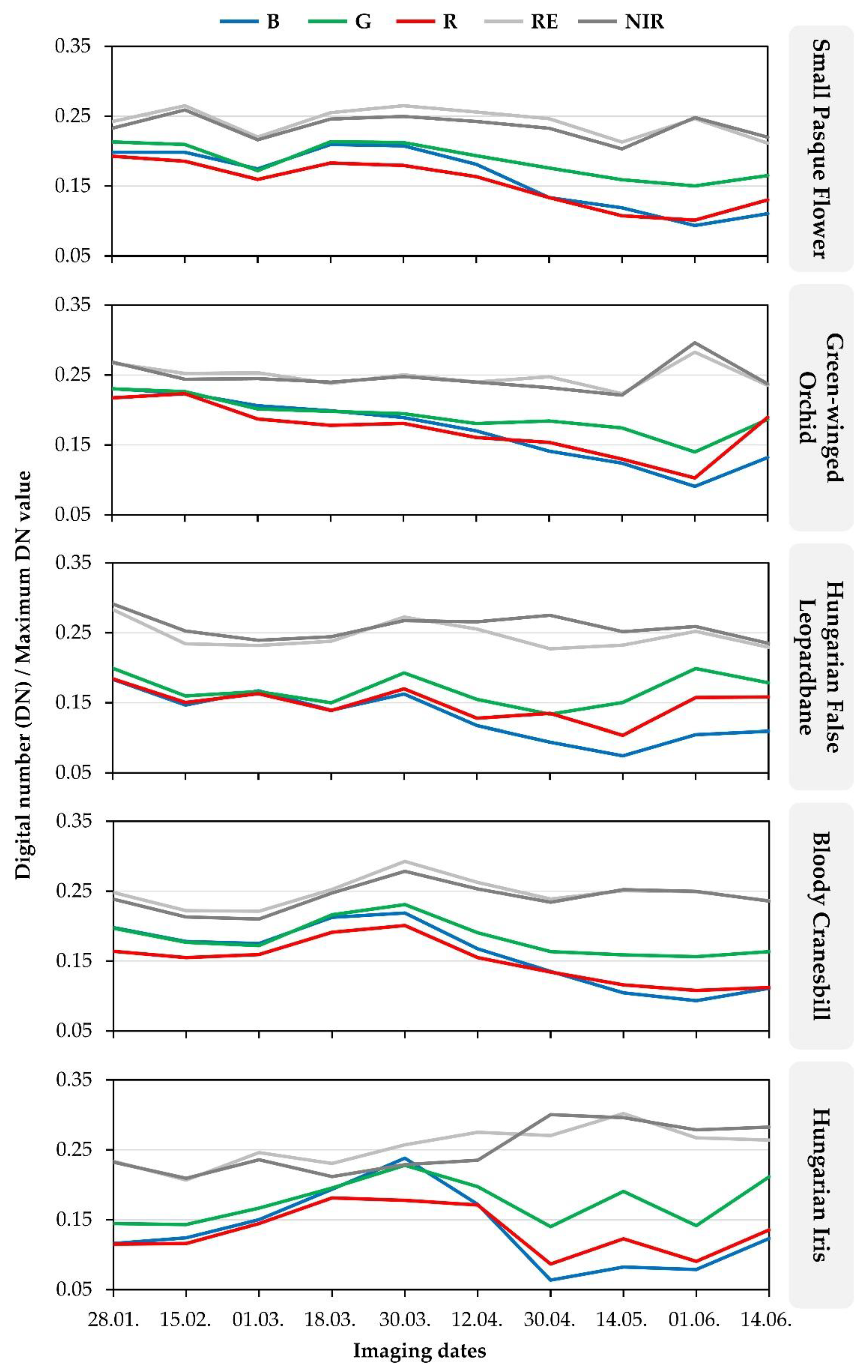
| Flora Species | Bands | Mean Digital Number (DN) Values Divided by a Maximum DN Value | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 28.01. | 15.02. | 01.03. | 18.03. | 30.03. | 12.04. | 30.04. | 14.05. | 01.06. | 14.06. | ||
| Small pasque flower | B | 0.198 | 0.198 | 0.175 | 0.210 | 0.208 | 0.181 | 0.134 | 0.119 | 0.094 | 0.111 |
| G | 0.213 | 0.209 | 0.172 | 0.214 | 0.212 | 0.193 | 0.176 | 0.159 | 0.150 | 0.165 | |
| R | 0.193 | 0.186 | 0.159 | 0.183 | 0.180 | 0.164 | 0.134 | 0.108 | 0.101 | 0.130 | |
| RE | 0.242 | 0.265 | 0.220 | 0.255 | 0.265 | 0.256 | 0.246 | 0.213 | 0.246 | 0.211 | |
| NIR | 0.233 | 0.259 | 0.216 | 0.246 | 0.250 | 0.243 | 0.233 | 0.203 | 0.248 | 0.220 | |
| Green-winged orchid | B | 0.231 | 0.224 | 0.206 | 0.199 | 0.189 | 0.170 | 0.141 | 0.124 | 0.091 | 0.132 |
| G | 0.230 | 0.226 | 0.201 | 0.198 | 0.195 | 0.181 | 0.184 | 0.174 | 0.140 | 0.187 | |
| R | 0.217 | 0.223 | 0.187 | 0.178 | 0.181 | 0.161 | 0.154 | 0.130 | 0.103 | 0.190 | |
| RE | 0.267 | 0.252 | 0.253 | 0.238 | 0.250 | 0.240 | 0.248 | 0.223 | 0.283 | 0.235 | |
| NIR | 0.268 | 0.244 | 0.245 | 0.240 | 0.248 | 0.240 | 0.232 | 0.221 | 0.296 | 0.237 | |
| Hungarian false leopardbane | B | 0.183 | 0.147 | 0.167 | 0.139 | 0.162 | 0.117 | 0.094 | 0.074 | 0.105 | 0.109 |
| G | 0.199 | 0.160 | 0.166 | 0.150 | 0.193 | 0.155 | 0.134 | 0.150 | 0.199 | 0.178 | |
| R | 0.184 | 0.150 | 0.163 | 0.139 | 0.170 | 0.128 | 0.135 | 0.104 | 0.157 | 0.158 | |
| RE | 0.283 | 0.234 | 0.232 | 0.238 | 0.273 | 0.255 | 0.227 | 0.232 | 0.252 | 0.229 | |
| NIR | 0.291 | 0.253 | 0.239 | 0.244 | 0.267 | 0.266 | 0.275 | 0.252 | 0.259 | 0.235 | |
| Bloody cranesbill | B | 0.198 | 0.178 | 0.175 | 0.213 | 0.219 | 0.167 | 0.135 | 0.105 | 0.093 | 0.111 |
| G | 0.197 | 0.177 | 0.172 | 0.216 | 0.231 | 0.190 | 0.164 | 0.159 | 0.156 | 0.163 | |
| R | 0.164 | 0.155 | 0.159 | 0.191 | 0.201 | 0.155 | 0.134 | 0.116 | 0.108 | 0.112 | |
| RE | 0.248 | 0.222 | 0.221 | 0.252 | 0.293 | 0.262 | 0.239 | 0.251 | 0.249 | 0.236 | |
| NIR | 0.239 | 0.213 | 0.210 | 0.247 | 0.279 | 0.253 | 0.234 | 0.252 | 0.250 | 0.236 | |
| Hungarian iris | B | 0.116 | 0.124 | 0.150 | 0.194 | 0.238 | 0.172 | 0.064 | 0.082 | 0.079 | 0.123 |
| G | 0.145 | 0.143 | 0.167 | 0.195 | 0.228 | 0.197 | 0.140 | 0.191 | 0.142 | 0.211 | |
| R | 0.115 | 0.116 | 0.145 | 0.181 | 0.178 | 0.171 | 0.087 | 0.123 | 0.091 | 0.136 | |
| RE | 0.234 | 0.207 | 0.246 | 0.231 | 0.257 | 0.275 | 0.271 | 0.302 | 0.267 | 0.264 | |
| NIR | 0.232 | 0.209 | 0.236 | 0.212 | 0.229 | 0.235 | 0.300 | 0.296 | 0.279 | 0.283 | |

References
- Biondi, E.; Casavecchia, S.; Pesaresi, S.; Zivkovic, L. Natura 2000 and the Pan-European Ecological Network: A New Methodology for Data Integration. Biodivers. Conserv. 2012, 21, 1741–1754. [Google Scholar] [CrossRef]
- Bhatti, A.Z.; Farooque, A.A.; Li, Q.; Abbas, F.; Acharya, B. Spatial Distribution and Sustainability Implications of the Canadian Groundwater Resources under Changing Climate. Sustainability 2021, 13, 9778. [Google Scholar] [CrossRef]
- Gregor, T.; Dressler, S.; Nierbauer, K.U.; Zizka, G. Loss of Plant Species Diversity in a Rural German Region-Assessment on Basis of a Historical Herbarium. Tuexenia 2016, 36, 191–204. [Google Scholar] [CrossRef]
- Hussein, E.A.; Abd El-Ghani, M.M.; Hamdy, R.S.; Shalabi, L.E. Do Anthropogenic Activities Affect Floristic Diversity and Vegetation Structure More than Natural Soil Properties in Hyper-Arid Desert Environments? Diversity 2021, 13, 157. [Google Scholar] [CrossRef]
- Jaric, I.; Roberts, D.L.; Gessner, J.; Solow, A.R.; Courchamp, F. Science Responses to IUCN Red Listing. PeerJ 2017, 5, e4025. [Google Scholar] [CrossRef]
- Hegde, R.; Niranjana, K.; Srinivas, S.; Danorkar, B.A.; Singh, S.K. Site-Specific Land Resource Inventory for Scientific Planning of Sujala Watersheds in Karnataka. Curr. Sci. 2018, 115, 644–652. [Google Scholar] [CrossRef]
- Pfeiffer, T.Z.; Maronic, D.S.; Zahirovic, V.; Stevic, F.; Zjalic, M.; Kajan, K.; Ozimec, S.; Mihaljevic, M. Early Spring Flora of the Sub-Pannonic Steppic Grassland (NATURA 2000 Site) in Bilje, Northeast Croatia. Acta Bot. Croat. 2016, 75, 157–163. [Google Scholar] [CrossRef][Green Version]
- Wright, J.W.; Davies, K.F.; Lau, J.A.; McCall, A.C.; McKay, J.K. Experimental Verification of Ecological Niche Modeling in a Heterogeneous Environment. Ecology 2006, 87, 2433–2439. [Google Scholar] [CrossRef]
- Radočaj, D.; Jurišić, M.; Gašparović, M.; Plaščak, I.; Antonić, O. Cropland Suitability Assessment Using Satellite-Based Biophysical Vegetation Properties and Machine Learning. Agronomy 2021, 11, 1620. [Google Scholar] [CrossRef]
- Nikolić, T.; Topić, J. Croatian Red Book of Vascular Flora; Ministry of Culture: Zagreb, Croatia, 2005.
- Flora Croatica Database, Vascular Plants Taxonomy and Bibliography of Croatian Flora. Available online: https://hirc.botanic.hr/fcd/ (accessed on 20 April 2022).
- Flora Croatica Database, Small Pasque Flower. Available online: https://hirc.botanic.hr/fcd/DetaljiFrame.aspx?IdVrste=8206&taxon=Pulsatilla+pratensis+(L.)+Miller+ssp.+nigricans+(St%C3%B6rck)+Zam (accessed on 19 April 2022).
- Flora Croatica Database, Green-Winged Orchid. Available online: https://hirc.botanic.hr/fcd/DetaljiFrame.aspx?IdVrste=6997&taxon=Orchis+morio+L (accessed on 19 April 2022).
- Flora Croatica Database, Hungarian False Leopardbane. Available online: https://hirc.botanic.hr/fcd/DetaljiFrame.aspx?IdVrste=27467&taxon=Doronicum+hungaricum+Rchb.+f (accessed on 19 April 2022).
- Flora Croatica Database, Bloody Cranesbill. Available online: https://hirc.botanic.hr/fcd/DetaljiFrame.aspx?IdVrste=4812&taxon=Geranium+sanguineum+L (accessed on 19 April 2022).
- Flora Croatica Database, Hungarian Iris. Available online: https://hirc.botanic.hr/fcd/DetaljiFrame.aspx?IdVrste=5437&taxon=Iris+variegata+L (accessed on 19 April 2022).
- Taghizadeh-Mehrjardi, R.; Nabiollahi, K.; Rasoli, L.; Kerry, R.; Scholten, T. Land Suitability Assessment and Agricultural Production Sustainability Using Machine Learning Models. Agronomy 2020, 10, 573. [Google Scholar] [CrossRef]
- Bako, G.; Molnar, Z.; Bakk, L.; Horvath, F.; Feher, L.; Abram, O.; Morvai, E.; Biro, C.; Papay, G.; Furesz, A.; et al. Toward a High Spatial Resolution Aerial Monitoring Network for Nature Conservation-How Can Remote Sensing Help Protect Natural Areas? Sustainability 2021, 13, 8807. [Google Scholar] [CrossRef]
- Zlinszky, A.; Heilmeier, H.; Balzter, H.; Czucz, B.; Pfeifer, N. Remote Sensing and GIS for Habitat Quality Monitoring: New Approaches and Future Research. Remote Sens. 2015, 7, 7987–7994. [Google Scholar] [CrossRef]
- Hallik, L.; Kuusk, A.; Lang, M.; Kuusk, J. Reflectance Properties of Hemiboreal Mixed Forest Canopies with Focus on Red Edge and Near Infrared Spectral Regions. Remote Sens. 2019, 11, 1717. [Google Scholar] [CrossRef]
- Rajah, P.; Odindi, J.; Mutanga, O. Evaluating the Potential of Freely Available Multispectral Remotely Sensed Imagery in Mapping American Bramble (Rubus cuneifolius). S. Afr. Geogr. J. 2018, 100, 291–307. [Google Scholar] [CrossRef]
- Zhou, H.; Zhu, J.; Li, J.; Xu, Y.; Li, Q.; Yan, E.; Zhao, S.; Xiong, Y.; Mo, D. Opening a New Era of Investigating Unreachable Cliff Flora Using Smart UAVs. Remote Sens. Ecol. Conserv. 2021, 7, 638–648. [Google Scholar] [CrossRef]
- Casalegno, S.; Amatulli, G.; Bastrup-Birk, A.; Durrant, T.H.; Pekkarinen, A. Modelling and Mapping the Suitability of European Forest Formations at 1-Km Resolution. Eur. J. For. Res. 2011, 130, 971–981. [Google Scholar] [CrossRef]
- Radočaj, D.; Jurišić, M.; Gašparović, M. A Wildfire Growth Prediction and Evaluation Approach Using Landsat and MODIS Data. J. Environ. Manag. 2022, 304, 114351. [Google Scholar] [CrossRef]
- Awais, M.; Aslam, B.; Maqsoom, A.; Khalil, U.; Ullah, F.; Azam, S.; Imran, M. Assessing Nitrate Contamination Risks in Groundwater: A Machine Learning Approach. Appl. Sci. 2021, 11, 10034. [Google Scholar] [CrossRef]
- Rodrigues, N.M.; Batista, J.E.; Mariano, P.; Fonseca, V.; Duarte, B.; Silva, S. Artificial Intelligence Meets Marine Ecotoxicology: Applying Deep Learning to Bio-Optical Data from Marine Diatoms Exposed to Legacy and Emerging Contaminants. Biology 2021, 10, 932. [Google Scholar] [CrossRef]
- Edalat, M.; Dastres, E.; Jahangiri, E.; Moayedi, G.; Zamani, A.; Pourghasemi, H.R.; Tiefenbacher, J.P. Spatial Mapping Zataria Multiflora Using Different Machine-Learning Algorithms. Catena 2022, 212, 106007. [Google Scholar] [CrossRef]
- Zheng, C.; Wen, Z.; Liu, Y.; Guo, Q.; Jiang, Y.; Ren, H.; Fan, Y.; Yang, Y. Integrating Habitat Suitability and the Near-Nature Restoration Priorities into Revegetation Plans Based on Potential Vegetation Distribution. Forests 2021, 12, 218. [Google Scholar] [CrossRef]
- Fois, M.; Cuena-Lombraña, A.; Fenu, G.; Bacchetta, G. Using Species Distribution Models at Local Scale to Guide the Search of Poorly Known Species: Review, Methodological Issues and Future Directions. Ecol. Model. 2018, 385, 124–132. [Google Scholar] [CrossRef]
- Republic of Croatia, State Geodetic Administration CROPOS Users’ Manual. Available online: https://www.cropos.hr/files/docs/cropos_users-manual.pdf (accessed on 22 November 2021).
- Hengl, T.; MacMillan, R.A. Predictive Soil Mapping with R; OpenGeoHub Foundation: Wageningen, The Netherlands, 2019; ISBN 978-0-359-30635-0. [Google Scholar]
- Kim, D.; Kupfer, J.A. Tri-Variate Relationships among Vegetation, Soil, and Topography along Gradients of Fluvial Biogeomorphic Succession. PLoS ONE 2016, 11, e0163223. [Google Scholar] [CrossRef] [PubMed]
- Begert, M.; Frei, C. Long-Term Area-Mean Temperature Series for SwitzerlandCombining Homogenized Station Data and High Resolution Grid Data. Int. J. Climatol. 2018, 38, 2792–2807. [Google Scholar] [CrossRef]
- Conrad, O.; Bechtel, B.; Bock, M.; Dietrich, H.; Fischer, E.; Gerlitz, L.; Wehberg, J.; Wichmann, V.; Boehner, J. System for Automated Geoscientific Analyses (SAGA) v. 2.1.4. Geosci. Model Dev. 2015, 8, 1991–2007. [Google Scholar] [CrossRef]
- Maresma, A.; Ariza, M.; Martinez, E.; Lloveras, J.; Martinez-Casasnovas, J.A. Analysis of Vegetation Indices to Determine Nitrogen Application and Yield Prediction in Maize (Zea mays L.) from a Standard UAV Service. Remote Sens. 2016, 8, 973. [Google Scholar] [CrossRef]
- Rouse, J.; Haas, R.H.; Schell, J.A.; Deering, D.W. Monitoring Vegetation Systems in the Great Plains with ERTS. NASA Spec. Publ. 1974, 351, 309. [Google Scholar]
- Barnes, E.M.; Clarke, T.R.; Richards, S.E.; Colaizzi, P.D.; Haberland, J.; Kostrzewski, M.; Waller, P.; Choi, C.; Riley, E.; Thompson, T.; et al. Coincident Detection of Crop Water Stress, Nitrogen Status and Canopy Density Using Ground-Based Multispectral Data. In Proceedings of the Fifth International Conference on Precision Agriculture, Bloomington, MN, USA, 16–19 July 2000; pp. 1–5. [Google Scholar]
- Easterday, K.; Kislik, C.; Dawson, T.E.; Hogan, S.; Kelly, M. Remotely Sensed Water Limitation in Vegetation: Insights from an Experiment with Unmanned Aerial Vehicles (UAVs). Remote Sens. 2019, 11, 1853. [Google Scholar] [CrossRef]
- Csajbok, J.; Buday-Bodi, E.; Nagy, A.; Feher, Z.Z.; Tamas, A.; Virag, I.C.; Bojtor, C.; Forgacs, F.; Vad, A.M.; Kutasy, E. Multispectral Analysis of Small Plots Based on Field and Remote Sensing Surveys-A Comparative Evaluation. Sustainability 2022, 14, 3339. [Google Scholar] [CrossRef]
- Blanco, V.; Jose Blaya-Ros, P.; Castillo, C.; Soto-Valles, F.; Torres-Sanchez, R.; Domingo, R. Potential of UAS-Based Remote Sensing for Estimating Tree Water Status and Yield in Sweet Cherry Trees. Remote Sens. 2020, 12, 2359. [Google Scholar] [CrossRef]
- Yue, L.; Wang, Y.; Wang, L.; Yao, S.; Cong, C.; Ren, L.; Zhang, B. Impacts of Soil Compaction and Historical Soybean Variety Growth on Soil Macropore Structure. Soil Tillage Res. 2021, 214, 105166. [Google Scholar] [CrossRef]
- Hengl, T.; Heuvelink, G.B.M.; Stein, A. A Generic Framework for Spatial Prediction of Soil Variables Based on Regression-Kriging. Geoderma 2004, 120, 75–93. [Google Scholar] [CrossRef]
- Radočaj, D.; Jug, I.; Vukadinović, V.; Jurišić, M.; Gašparović, M. The Effect of Soil Sampling Density and Spatial Autocorrelation on Interpolation Accuracy of Chemical Soil Properties in Arable Cropland. Agronomy 2021, 11, 2430. [Google Scholar] [CrossRef]
- Böhner, J.; Antonić, O. Chapter 8 Land-Surface Parameters Specific to Topo-Climatology. In Developments in Soil Science; Hengl, T., Reuter, H.I., Eds.; Geomorphometry; Elsevier: Amsterdam, The Netherlands, 2009; Volume 33, pp. 195–226. [Google Scholar]
- Fan, B.; Tao, W.; Qin, G.; Hopkins, I.; Zhang, Y.; Wang, Q.; Lin, H.; Guo, L. Soil Micro-Climate Variation in Relation to Slope Aspect, Position, and Curvature in a Forested Catchment. Agric. For. Meteorol. 2020, 290, 107999. [Google Scholar] [CrossRef]
- Kammer, P.M.; Schoeb, C.; Eberhard, G.; Gallina, R.; Meyer, R.; Tschanz, C. The Relationship between Soil Water Storage Capacity and Plant Species Diversity in High Alpine Vegetation. Plant Ecol. Divers. 2013, 6, 457–466. [Google Scholar] [CrossRef]
- Freeman, T.G. Calculating Catchment Area with Divergent Flow Based on a Regular Grid. Comput. Geosci. 1991, 17, 413–422. [Google Scholar] [CrossRef]
- Johnson, B.G.; Thackray, G.D.; Van Kirk, R. The Effect of Topography, Latitude, and Lithology on Rock Glacier Distribution in the Lemhi Range, Central Idaho, USA. Geomorphology 2007, 91, 38–50. [Google Scholar] [CrossRef]
- Kuhn, M. Building Predictive Models in R Using the Caret Package. J. Stat. Softw. 2008, 28, 1–26. [Google Scholar] [CrossRef]
- Ogola, H.J.O.; Selvarajan, R.; Tekere, M. Local Geomorphological Gradients and Land Use Patterns Play Key Role on the Soil Bacterial Community Diversity and Dynamics in the Highly Endemic Indigenous Afrotemperate Coastal Scarp Forest Biome. Front. Microbiol. 2021, 12, 592725. [Google Scholar] [CrossRef]
- Kurtz, B.C.; Souza, V.C.; Magalhaes, A.M.; de Paula-Souza, J.; Duarte, A.R.; Joaquim, G.O. The Vascular Flora and Vegetation of Queimada Grande Island, Sao Paulo State, Southeastern Brazil. Biota Neotrop. 2017, 17, e20170336. [Google Scholar] [CrossRef]
- Campos, V.E.; Gatica, G.; Bellis, L.M. Remote Sensing Variables as Predictors of Habitat Suitability of the Viscacha Rat (Octomys mimax), a Rock-Dwelling Mammal Living in a Desert Environment. Mammal. Res. 2015, 60, 117–126. [Google Scholar] [CrossRef]
- Fish, A. Crash Theory: Entrapments of Conservation Drones and Endangered Megafauna. Sci. Technol. Hum. Values 2021, 46, 425–451. [Google Scholar] [CrossRef]
- De Deyn, G.B.; Raaijmakers, C.E.; Zoomer, H.R.; Berg, M.P.; de Ruiter, P.C.; Verhoef, H.A.; Bezemer, T.M.; van der Putten, W.H. Soil Invertebrate Fauna Enhances Grassland Succession and Diversity. Nature 2003, 422, 711–713. [Google Scholar] [CrossRef] [PubMed]
- Luft, L.; Neumann, C.; Freude, M.; Blaum, N.; Jeltsch, F. Hyperspectral Modeling of Ecological Indicators—A New Approach for Monitoring Former Military Training Areas. Ecol. Indic. 2014, 46, 264–285. [Google Scholar] [CrossRef]
- Li, Q.; Wong, F.K.K.; Fung, T. Mapping Multi-Layered Mangroves from Multispectral, Hyperspectral, and LiDAR Data. Remote Sens. Environ. 2021, 258, 112403. [Google Scholar] [CrossRef]
- Saad, F.; Biswas, S.; Huang, Q.; Corte, A.P.D.; Coraiola, M.; Macey, S.; Carlucci, M.B.; Leimgruber, P. Detectability of the Critically Endangered Araucaria Angustifolia Tree Using Worldview-2 Images, Google Earth Engine and UAV-LiDAR. Land 2021, 10, 1316. [Google Scholar] [CrossRef]
- Rominger, K.; Meyer, S.E. Application of UAV-Based Methodology for Census of an Endangered Plant Species in a Fragile Habitat. Remote Sens. 2019, 11, 719. [Google Scholar] [CrossRef]
- Wylie, B.K.; Pastick, N.J.; Picotte, J.J.; Deering, C.A. Geospatial Data Mining for Digital Raster Mapping. GISci. Remote Sens. 2019, 56, 406–429. [Google Scholar] [CrossRef]
- Leduc, M.-B.; Knudby, A.J. Mapping Wild Leek through the Forest Canopy Using a UAV. Remote Sens. 2018, 10, 70. [Google Scholar] [CrossRef]
- Radočaj, D.; Obhođaš, J.; Jurišić, M.; Gašparović, M. Global Open Data Remote Sensing Satellite Missions for Land Monitoring and Conservation: A Review. Land 2020, 9, 402. [Google Scholar] [CrossRef]
- Eugenio Abrego, M.; Acuna-Perales, N.; Alfaro-Shigueto, J.; Azocar, J.; Barragan Rocha, A.R.; Baquero, A.; Cotto, A.; Darquea, J.; de Paz, N.; Donoso, M.; et al. Enhanced, Coordinated Conservation Efforts Required to Avoid Extinction of Critically Endangered Eastern Pacific Leatherback Turtles. Sci. Rep. 2020, 10, 4772. [Google Scholar] [CrossRef]
- Warner, K.; Zommers, Z.; Wreford, A.; Hurlbert, M.; Viner, D.; Scantlan, J.; Halsey, K.; Halsey, K.; Tamang, C. Characteristics of Transformational Adaptation in Climate-Land-Society Interactions. Sustainability 2019, 11, 356. [Google Scholar] [CrossRef]












| English Name | Latin Name | Family | IUCN Class | Units Detected |
|---|---|---|---|---|
| Small pasque flower | Pulsatilla pratensis (L.) Miller ssp. nigricans (Störck) Zämelis | Ranunculaceae | CR | 7 |
| Green-winged orchid | Orchis morio (L.) | Orchidaceae | NT | 29 |
| Hungarian false leopardbane | Doronicum hungaricum Rchb.f. | Asteraceae | CR | 20 |
| Bloody cranesbill | Geranium sanguineum (L.) | Geraniaceae | LC | 61 |
| Hungarian iris | Iris variegata (L.) | Iridaceae | NT | 12 |
| First Observation | Second Observation | NDVI | NDRE | ||
|---|---|---|---|---|---|
| t | p | t | p | ||
| 28 January 2021 | 15 February 2021 | 5.2524 | <0.0001 | 5.8283 | <0.0001 |
| 15 February 2021 | 1 March 2021 | 2.1964 | 0.0290 | 0.9416 | 0.3472 |
| 1 March 2021 | 18 March 2021 | −2.9588 | 0.0034 | −2.7005 | 0.0074 |
| 18 March 2021 | 30 March 2021 | −2.8197 | 0.0052 | −4.9542 | <0.0001 |
| 30 March 2021 | 12 April 2021 | −6.4170 | <0.0001 | −7.021 | <0.0001 |
| 12 April 2021 | 30 April 2021 | −1.6302 | 0.1043 | −0.5633 | 0.5737 |
| 30 April 2021 | 14 May 2021 | −5.8383 | <0.0001 | −6.2335 | <0.0001 |
| 14 May 2021 | 1 June 2021 | 0.6704 | 0.5032 | 0.9132 | 0.3620 |
| 1 June 2021 | 14 June 2021 | 9.0903 | <0.0001 | 9.9859 | <0.0001 |
| Flora Species | Metric | Random Forest | XGBoost | Neural Network | Generalized Linear Model |
|---|---|---|---|---|---|
| Bloody cranesbill | ROC | 0.964 | 0.956 | 0.914 | 0.694 |
| Sensitivity | 0.567 | 0.617 | 0.517 | 0.500 | |
| Specificity | 0.967 | 0.933 | 0.902 | 0.883 | |
| Green-winged orchid | ROC | 0.967 | 0.956 | 0.922 | 0.964 |
| Sensitivity | 0.900 | 0.817 | 0.683 | 0.783 | |
| Specificity | 0.933 | 0.933 | 0.867 | 0.967 | |
| Hungarian false leopardbane | ROC | 0.999 | 0.967 | 0.958 | 0.825 |
| Sensitivity | 0.967 | 0.933 | 0.733 | 0.633 | |
| Specificity | 0.950 | 0.900 | 0.950 | 0.900 | |
| Hungarian iris | ROC | 0.999 | 0.999 | 0.999 | 0.850 |
| Sensitivity | 0.999 | 0.950 | 0.867 | 0.650 | |
| Specificity | 0.999 | 0.999 | 0.999 | 0.999 | |
| Small pasque flower | ROC | 0.809 | 0.738 | 0.762 | 0.607 |
| Sensitivity | 0.933 | 0.867 | 0.633 | 0.700 | |
| Specificity | 0.429 | 0.143 | 0.857 | 0.714 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jurišić, M.; Radočaj, D.; Plaščak, I.; Rapčan, I. A UAS and Machine Learning Classification Approach to Suitability Prediction of Expanding Natural Habitats for Endangered Flora Species. Remote Sens. 2022, 14, 3054. https://doi.org/10.3390/rs14133054
Jurišić M, Radočaj D, Plaščak I, Rapčan I. A UAS and Machine Learning Classification Approach to Suitability Prediction of Expanding Natural Habitats for Endangered Flora Species. Remote Sensing. 2022; 14(13):3054. https://doi.org/10.3390/rs14133054
Chicago/Turabian StyleJurišić, Mladen, Dorijan Radočaj, Ivan Plaščak, and Irena Rapčan. 2022. "A UAS and Machine Learning Classification Approach to Suitability Prediction of Expanding Natural Habitats for Endangered Flora Species" Remote Sensing 14, no. 13: 3054. https://doi.org/10.3390/rs14133054
APA StyleJurišić, M., Radočaj, D., Plaščak, I., & Rapčan, I. (2022). A UAS and Machine Learning Classification Approach to Suitability Prediction of Expanding Natural Habitats for Endangered Flora Species. Remote Sensing, 14(13), 3054. https://doi.org/10.3390/rs14133054