
Modeling Artificial Light Exposure after Vegetation Trimming at a Marine Turtle Nesting Beach


Abstract
:1. Introduction
2. Materials and Methods
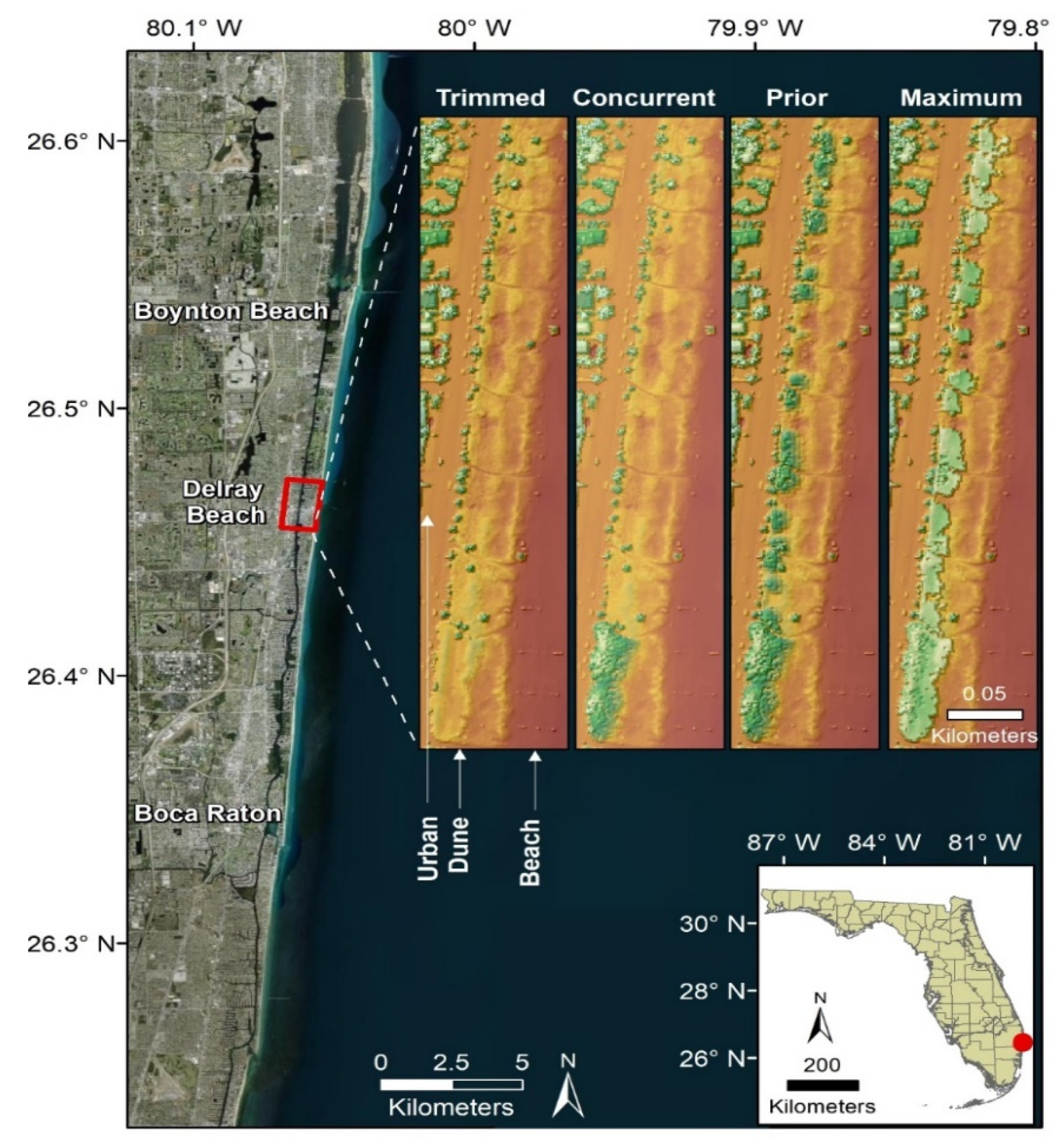
2.1. Study Site
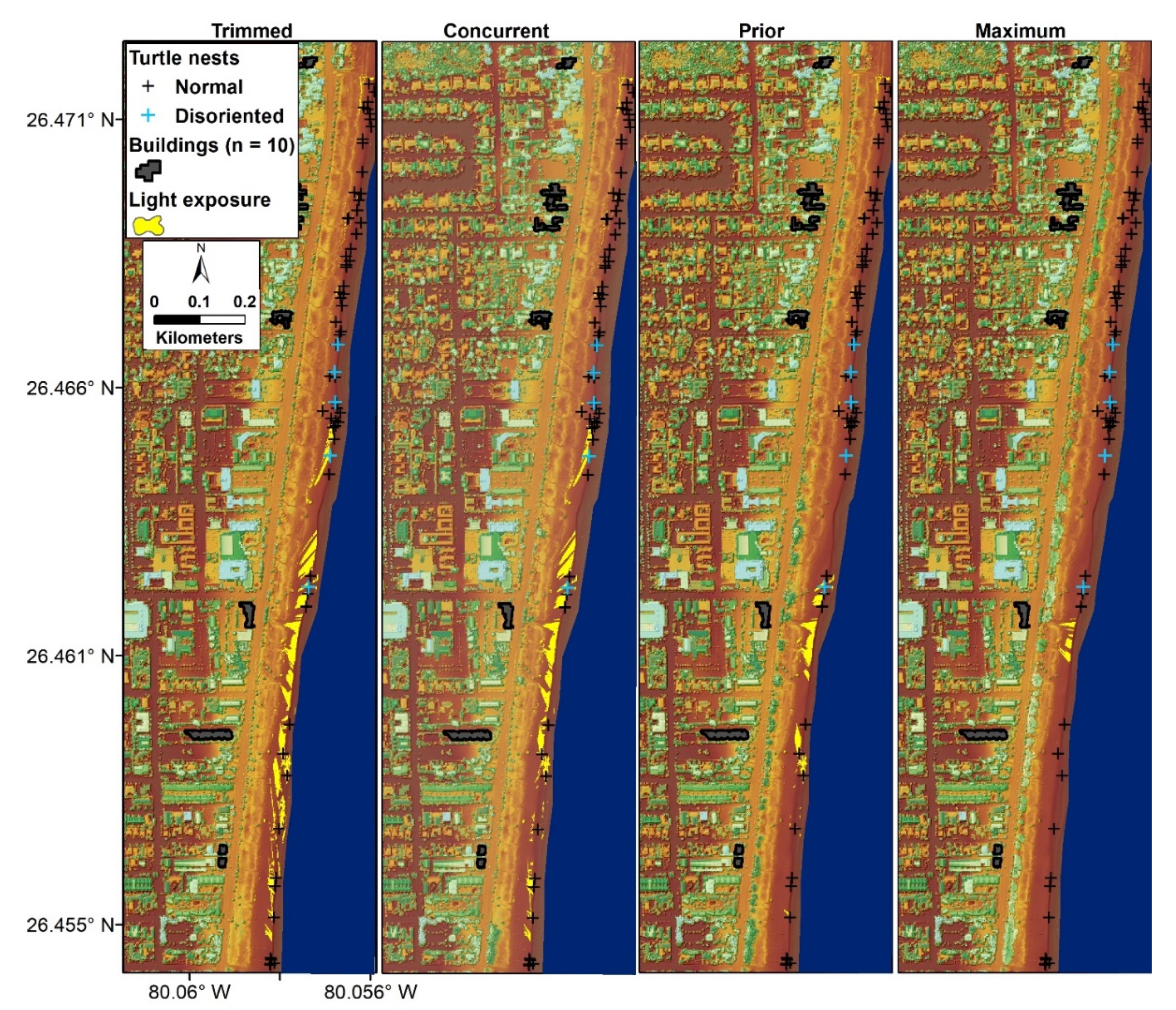
2.2. Data Collection
2.3. Surface Models, Buildings, and Vegetation Profiles
2.4. Viewshed Models
2.4.1. Cumulative Viewshed
2.4.2. Randomly Iterated Viewsheds
2.4.3. Nonrandomly Selected Viewsheds
3. Results
3.1. Surface Model Summaries
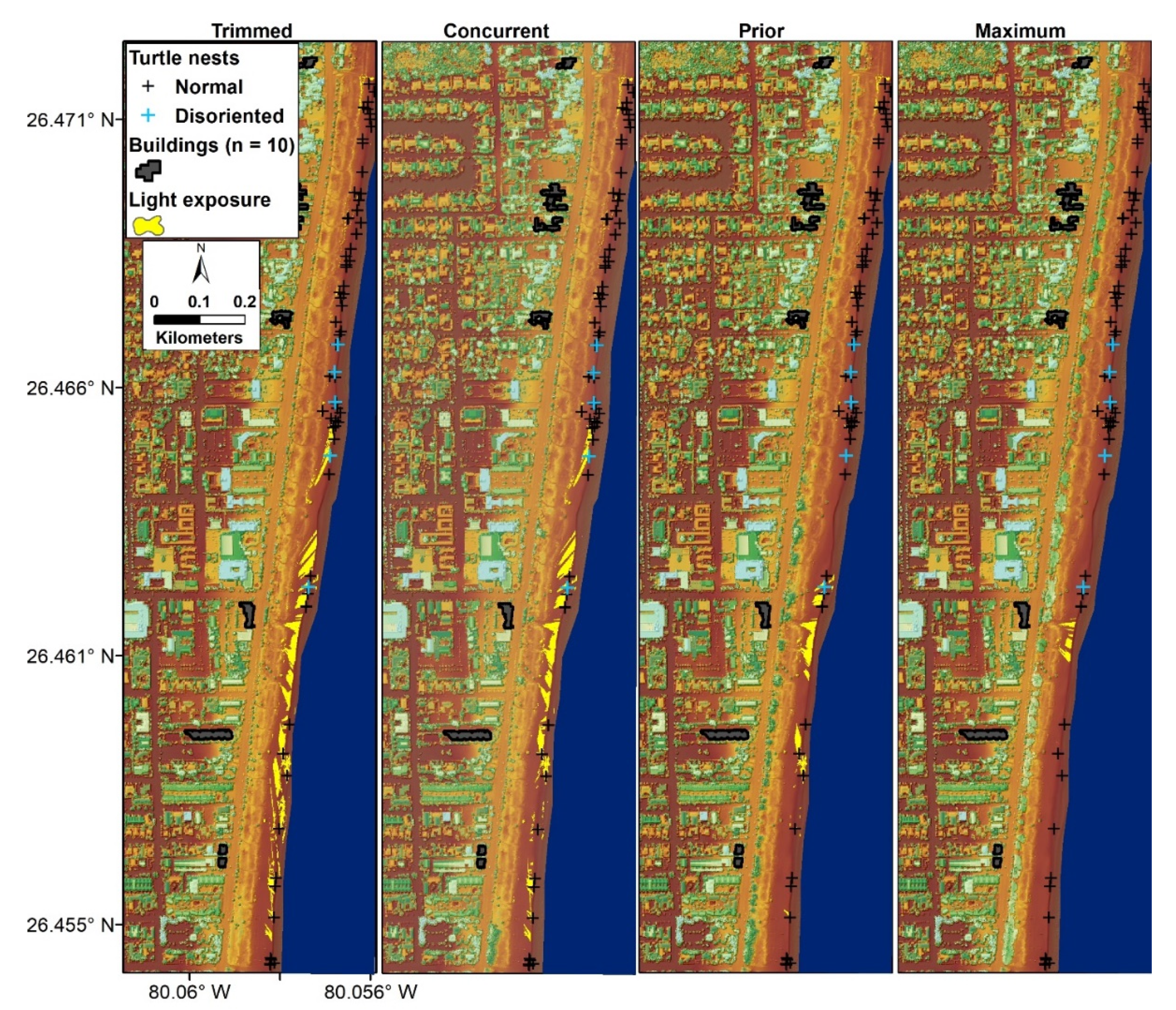
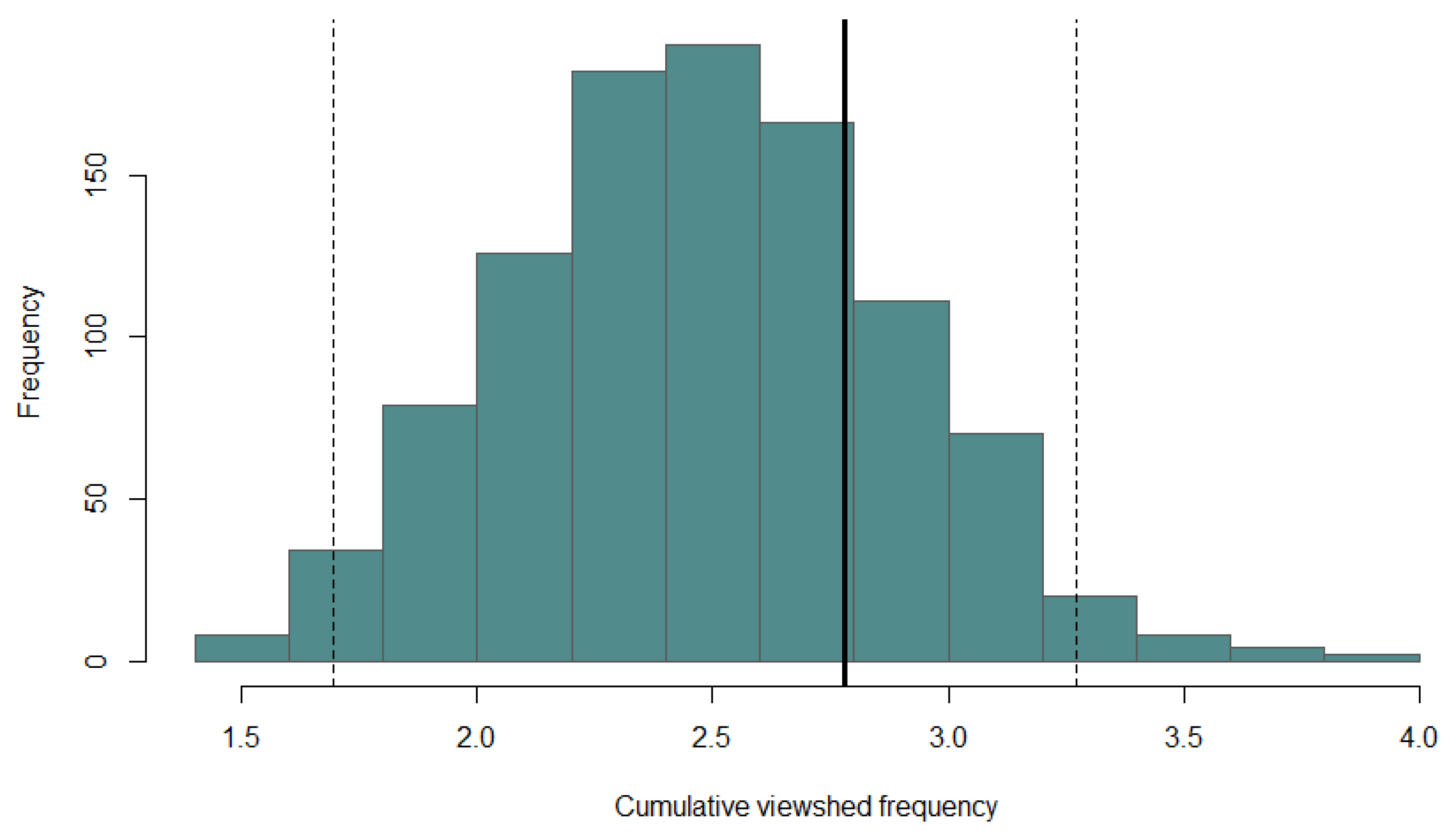
3.2. Cumulative Viewshed
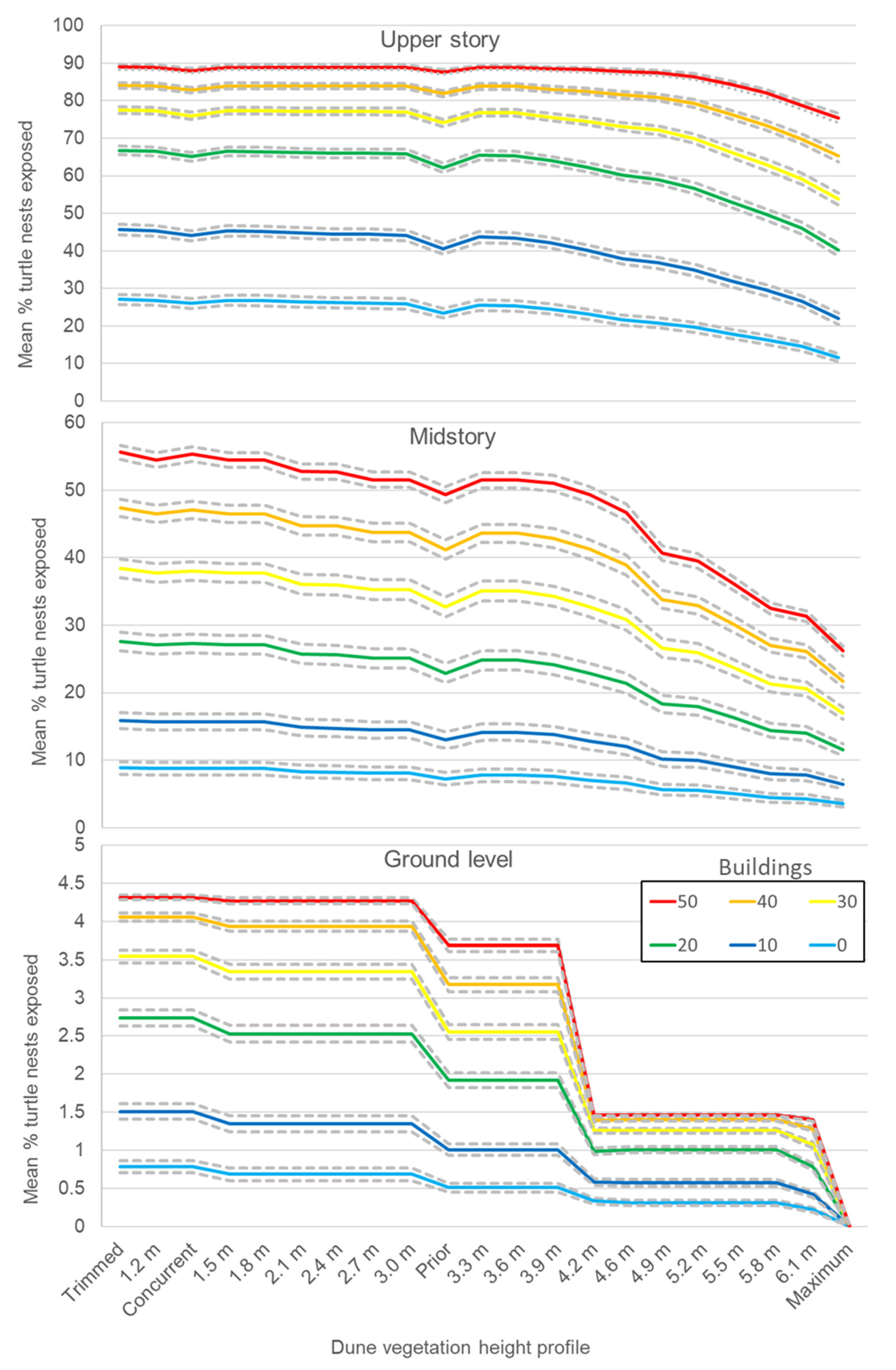
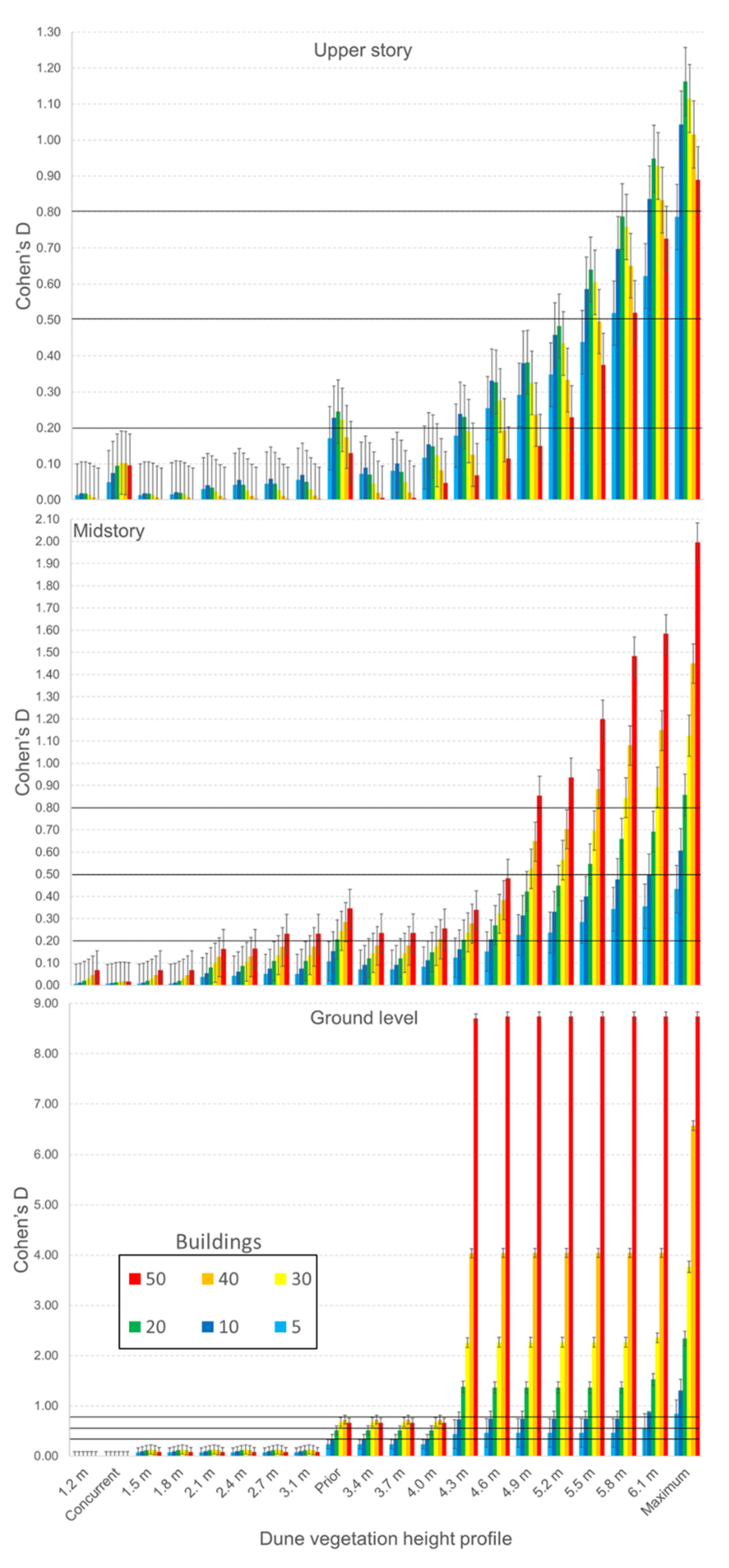
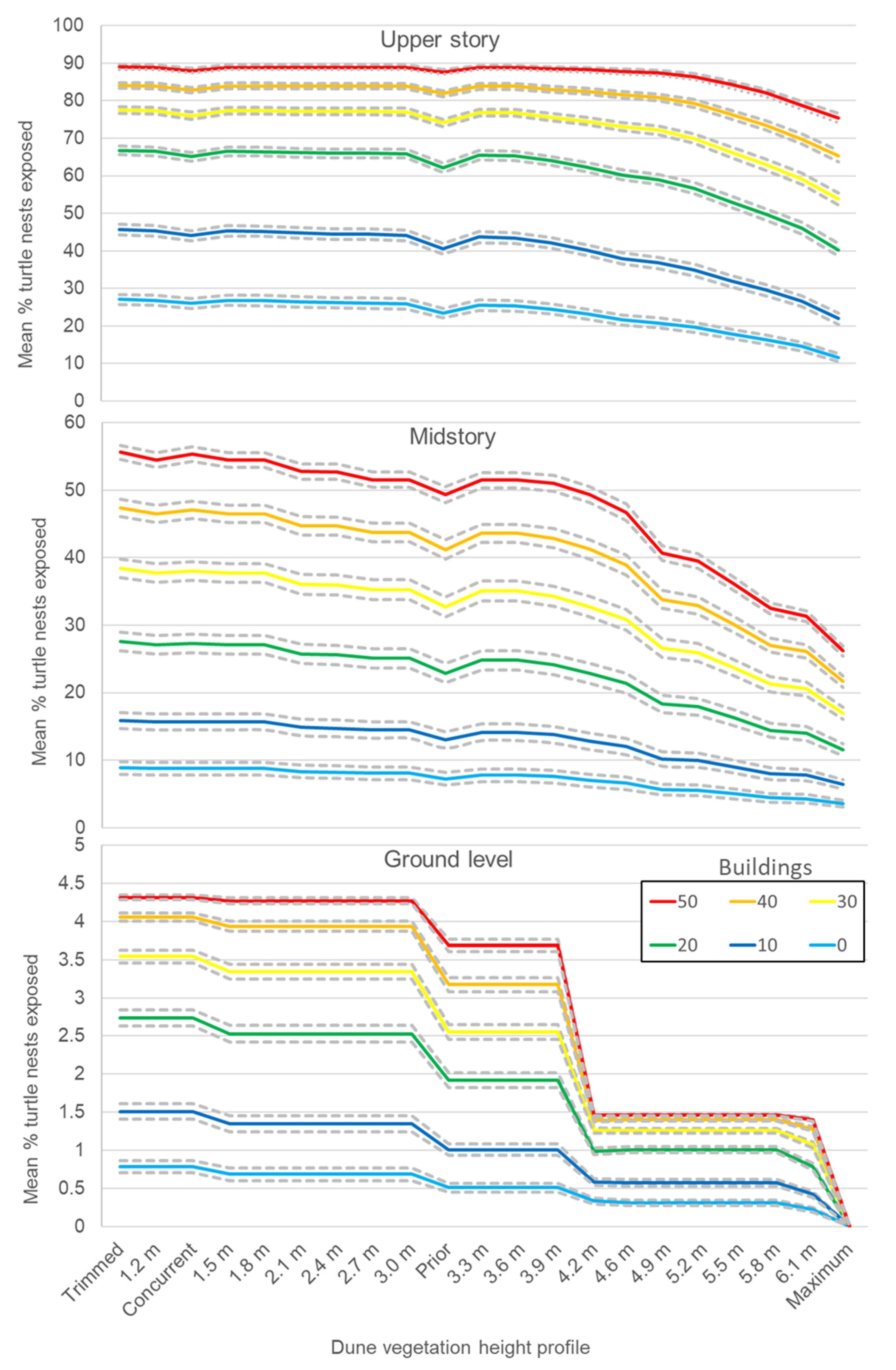
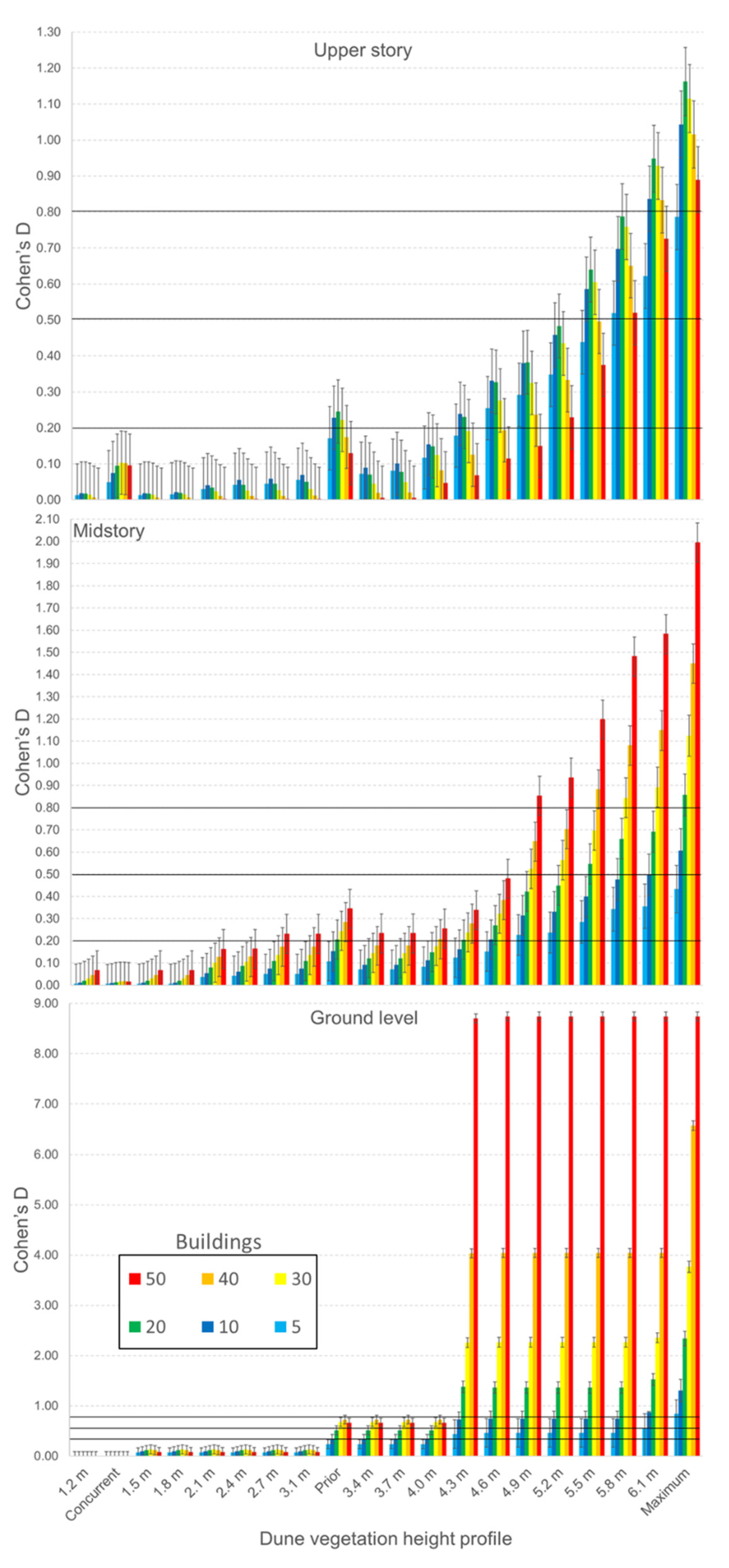
3.3. Randomly Iterated Viewsheds
3.4. Nonrandomly Selected Viewsheds
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bennie, J.; Duffy, J.P.; Davies, T.W.; Correa-Cano, M.E.; Gaston, K.J. Global trends in exposure to light pollution in natural terrestrial ecosystems. Remote Sens. 2015, 7, 2715–2730. [Google Scholar] [CrossRef] [Green Version]
- Hölker, F.; Moss, T.; Griefahn, B.; Kloas, W.; Voigt, C.C.; Henckel, D.; Hänel, A.; Kappeler, P.M.; Völker, S.; Schwope, A.; et al. The dark side of light: A transdisciplinary research agenda for light pollution policy. Ecol. Soc. 2010, 15, 13. [Google Scholar] [CrossRef]
- Gaston, K.J.; Bennie, J.; Davies, T.W.; Hopkins, J. The ecological impacts of nighttime light pollution: A mechanistic appraisal. Biol. Rev. 2013, 88, 912–927. [Google Scholar] [CrossRef] [PubMed]
- Longcore, T.; Rich, C. Ecological light pollution. Front. Ecol. Environ. 2004, 2, 191–198. [Google Scholar] [CrossRef]
- Davies, T.W.; Bennie, J.; Inger, R.; de Ibarra, N.H.; Gaston, K.J. Artificial light pollution: Are shifting spectral signatures changing the balance of species interactions? Glob. Chang. Biol. 2013, 19, 1417–1423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karnad, D.; Isvaran, K.; Kar, C.S.; Shanker, K. Lighting the way: Towards reducing misorientation of olive ridley hatchlings due to artificial lighting at Rushikulya, India. Biol. Conserv. 2009, 142, 2083–2088. [Google Scholar] [CrossRef]
- Kamrowski, R.L.; Limpus, C.; Moloney, J.; Hamann, M. Coastal light pollution and marine turtles: Assessing the magnitude of the problem. Endanger. Species Res. 2012, 19, 85–98. [Google Scholar] [CrossRef] [Green Version]
- Kamrowski, R.L.; Limpus, C.; Pendoley, K.; Hamann, M. Influence of industrial light pollution on the sea-finding behaviour of flatback turtle hatchlings. Wildl. Res. 2014, 41, 421–434. [Google Scholar] [CrossRef]
- Verutes, G.M.; Huang, C.; Estrella, R.R.; Loyd, K. Exploring scenarios of light pollution from coastal development reaching sea turtle nesting beaches near Cabo Pulmo, Mexico. Glob. Ecol. Conserv. 2014, 2, 170–180. [Google Scholar] [CrossRef] [Green Version]
- Weishampel, Z.A.; Cheng, W.-H.H.; Weishampel, J.F. Sea turtle nesting patterns in Florida vis-à-vis satellite-derived measures of artificial lighting. Remote Sens. Ecol. Conserv. 2016, 2, 59–72. [Google Scholar] [CrossRef]
- Kelly, I.; Leon, J.X.; Gilby, B.L.; Olds, A.D.; Schlacher, T.A. Marine turtles are not fussy nesters: A novel test of small-scale nest site selection using structure from motion beach terrain information. PeerJ 2017, 5, e2770. [Google Scholar] [CrossRef] [PubMed]
- Bolten, A.B.; Crowder, L.B.; Dodd, M.G.; Macpherson, S.L.; Musick, J.A.; Schroeder, B.A.; Witherington, B.E.; Long, K.J.; Snover, M.L. Quantifying multiple threats to endangered species: An example from loggerhead sea turtles. Front. Ecol. Environ. 2011, 9, 295–301. [Google Scholar] [CrossRef] [Green Version]
- Witherington, B.E.; Bjorndal, K.A. Influences of artificial lighting on the seaward orientation of hatchling loggerhead turtles Caretta caretta. Biol. Conserv. 1991, 55, 139–149. [Google Scholar] [CrossRef]
- Witherington, B.E.; Martin, R.E. Understanding, Assessing, and Resolving Light-Pollution Problems on Sea Turtle Nesting Beaches, 3rd ed.; Technical Report TR-2; Florida Marine Research Institute: St. Petersburg, FL, USA, 2000; pp. 1–73. [Google Scholar]
- Witherington, B.E.; Bjorndal, K.A. Influences of wavelength and intensity on hatchling sea turtle phototaxis: Implications for sea-finding behavior. Copeia 1991, 4, 1060–1069. [Google Scholar] [CrossRef]
- Limpus, C.J.; Kamrowski, R.L. Ocean-finding in marine turtles: The importance of low horizon elevation as an orientation cue. Behaviour 2013, 150, 863–893. [Google Scholar] [CrossRef]
- Salmon, M.; Wyneken, J.; Fritz, E.; Lucas, M. Seafinding by hatchling sea turtles: Role of brightness, silhouette and beach slope as orientation cues. Behavior 1992, 122, 56–77. [Google Scholar]
- Pankaew, K.; Milton, S.L. The effects of extended crawling on the physiology and swim performance of loggerhead and green sea turtle hatchlings. J. Exp. Biol. 2018, 221, jeb165225. [Google Scholar] [CrossRef] [Green Version]
- Erb, V.; Wyneken, J. Nest-to-surf mortality of loggerhead sea turtle (Caretta caretta) hatchlings on Florida’s east coast. Front. Mar. Sci. 2019, 6, 271. [Google Scholar] [CrossRef]
- Nelson Sella, K.A.; Sicius, L.; Fuentes, M.M.P.B. Using expert elicitation to determine the relative impact of coastal modifications on marine turtle nesting grounds. Coast. Manag. 2019, 47, 492–506. [Google Scholar] [CrossRef]
- Dimitriadis, C.; Fournari-Konstantinidou, I.; Sourbès, L.; Koutsoubas, D.; Mazaris, A.D. Reduction of sea turtle population recruitment caused by nightlight: Evidence from the Mediterranean region. Ocean Coast. Manag. 2018, 153, 108–115. [Google Scholar] [CrossRef]
- Bertolotti, L.; Salmon, M. Do embedded roadway lights protect sea turtles? Environ. Manage. 2005, 36, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Gaston, K.J.; Davies, T.W.; Bennie, J.; Hopkins, J. Reducing the ecological consequences of night-time light pollution: Options and developments. J. Appl. Ecol. 2012, 49, 1256–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salmon, M.; Reiners, R.; Lavin, C.; Wyneken, J. Behavior of loggerhead sea turtles on an urban beach. II. Correlates of nest placement. J. Herpetol. 1995, 29, 560–567. [Google Scholar] [CrossRef]
- Small, C.; Nicholls, R.J. A global analysis of human settlement in coastal zones. J. Coast. Res. 2003, 19, 584–599. [Google Scholar]
- Sigren, J.M.; Figlus, J.; Highfield, W.; Feagin, R.A.; Armitage, A.R. The effects of coastal dune volume and vegetation on storm-induced property damage: Analysis from Hurricane Ike. J. Coast. Res. 2018, 34, 164–173. [Google Scholar] [CrossRef]
- Nordstrom, K.F.; Lampe, R.; Vandemark, L.M. Reestablishing naturally functioning dunes on developed coasts. Environ. Manag. 2000, 25, 37–51. [Google Scholar] [CrossRef]
- Hamilton, S.E.; Morgan, A. Integrating LiDAR, GIS and hedonic price modeling to measure amenity values in urban beach residential property markets. Comput. Environ. Urban Syst. 2010, 34, 133–141. [Google Scholar] [CrossRef] [Green Version]
- Hindsley, P.; Hamilton, S.E.; Morgan, O.A. Gulf views: Toward a better understanding of viewshed scope in hedonic property models. J. Real Estate Financ. Econ. 2013, 47, 489–505. [Google Scholar] [CrossRef] [Green Version]
- Bird, B.L.; Branch, L.C.; Miller, D.L. Effects of coastal lighting on foraging behavior of beach mice. Conserv. Biol. 2004, 18, 1435–1439. [Google Scholar] [CrossRef]
- Aben, J.; Pellikka, P.; Travis, J.M.J. A call for viewshed ecology: Advancing our understanding of the ecology of information through viewshed analysis. Methods Ecol. Evol. 2018, 9, 624–633. [Google Scholar] [CrossRef]
- Lefsky, M.A.; Cohen, W.B.; Parker, G.G.; Harding, D.J. Lidar remote sensing for ecosystem studies. Bioscience 2002, 52, 19–30. [Google Scholar] [CrossRef]
- Lecigne, B.; Eitel, J.U.H.; Rachlow, J.L. viewshed3d: An R package for quantifying 3D visibility using terrestrial LiDAR data. Methods Ecol. Evol. 2020, 11, 733–738. [Google Scholar] [CrossRef] [Green Version]
- Troy, J.R.; Holmes, N.D.; Green, M.C. Modeling artificial light viewed by fledgling seabirds. Ecosphere 2011, 2, art109. [Google Scholar] [CrossRef]
- Florida Department of Environmental Protection (FDEP). Coastal Range Monument Location; FDEP Bureau of Beaches and Coastal Systems: Tallahassee, FL, USA, 2017. Available online: https://floridadep.gov/sites/default/files/FDEP_Range_Monuments_FAQ_04-2020.pdf (accessed on 5 May 2021).
- Meylan, A.B.; Schroeder, B.A.; Mosier, A. Sea Turtle Nesting Activity in the State of Florida 1979–1992; Florida Marine Research Institute: St. Petersburg, FL, USA; Florida Department of Environmental Protection: Tallahassee, FL, USA, 1995; pp. 1–51. [Google Scholar]
- Gilman, E.F.; Watson, D.G.; Klein, R.W.; Koeser, A.K.; Hilbert, D.R.; McLean, D.C. Coccoloba uvifera: Seagrape; Series No. ENH334; Environmental Horticulture Department of University of Florida/Institute of Food and Agricultural Sciences Extension: Gainesville, FL, USA, 2018. [Google Scholar]
- Aptim Environmental and Infrastructure LLC; Coastal Protection Engineering LLC. City of Delray Beach Sea Grape Trimming Plan; Aptim Environmental and Infrastructure LLC and Coastal Protection Engineering LLC: Boca Raton, FL, USA, 2020. Available online: https://www.delraybeachfl.gov/home/showpublisheddocument?id=8160 (accessed on 5 May 2021).
- Florida Fish and Wildlife Conservation Commission (FWC). Florida Fish and Wildlife Conservation Commission Marine Turtle Conservation Handbook; Florida Fish and Wildlife Conservation Commission (FWC): Tallahassee, FL, USA, 2016. [Google Scholar]
- Florida Fish and Wildlife Conservation Commission (FWC). Post-Construction Monitoring Program Database; Florida Fish and Wildlife Conservation Commission (FWC) Imperiled Species Management: Tallahassee, FL, USA, 2020. [Google Scholar]
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: http://www.R-project.org/ (accessed on 5 May 2021).
- Wickham, H.R.; François, L.H.; Müller, K. A Grammar of Data Manipulation, R Package Version 1.0.6. 7 February 2021, pp. 1–78. Available online: https://cran.r-project.org/web/packages/dplyr/dplyr.pdf (accessed on 5 May 2021).
- Bates, D.; Maechler, M. Sparse and Dense Matrix Classes and Methods, R Package Version 1.3-3. 23 March 2021, pp. 1–226. Available online: https://cran.r-project.org/web/packages/Matrix/Matrix.pdf (accessed on 5 May 2021).
- Torchiano, M. Efficient Effect Size Computation, R Package Version 0.8.1. 5 October 2020, pp. 1–11. Available online: https://cran.r-project.org/web/packages/effsize/effsize.pdf (accessed on 5 May 2021).
- Cohen, J. A power primer. Psychol. Bull. 1992, 112, 155–159. [Google Scholar] [CrossRef]
- Long, T.M.; Florida Fish and Wildlife Conservation Commission, Tallahassee, FL, USA. Personal Communication, 2022.
- National Marine Fisheries Service (NMFS); U.S. Fish and Wildlife Service. Recovery Plan for the Northwest Atlantic Population of the Loggerhead Sea Turtle (Caretta caretta), 2nd ed.; National Marine Fisheries Service (NMFS): Silver Spring, MD, USA, 2008. [Google Scholar]
- Jensen, M.P.; Allen, C.D.; Eguchi, T.; Bell, I.P.; LaCasella, E.L.; Hilton, W.A.; Hof, C.A.M.; Dutton, P.H. Environmental warming and feminization of one of the largest sea turtle populations in the world. Curr. Biol. 2018, 28, 154–159. [Google Scholar] [CrossRef] [Green Version]
- Santidrián Tomillo, P.; Oro, D.; Paladino, F.V.; Piedra, R.; Sieg, A.E.; Spotila, J.R. High beach temperatures increased female-biased primary sex ratios but reduced output of female hatchlings in the leatherback turtle. Biol. Conserv. 2014, 176, 71–79. [Google Scholar] [CrossRef]
- McNeill, J.B.; Avens, L.; Hall, A.G.; Goshe, L.R.; Harms, C.A.; Owens, D.W. Female-bias in a long-term study of a species with temperature-dependent sex determination: Monitoring sex ratios for climate change research. PLoS ONE 2016, 11, e0160911. [Google Scholar] [CrossRef]
- Reboul, I.; Booth, D.; Rusli, U. Artificial and natural shade: Implications for green turtle (Chelonia mydas) rookery management. Ocean Coast. Manag. 2021, 204, 105521. [Google Scholar] [CrossRef]
- Kamel, S.J.; Mrosovsky, N. Deforestation: Risk of sex ratio distortion in hawksbill sea turtles. Ecol. Appl. 2006, 16, 923–931. [Google Scholar] [CrossRef] [Green Version]
- Esteban, N.; Laloë, J.-O.; Mortimer, J.A.; Guzman, A.N.; Hays, G.C. Male hatchling production in sea turtles from one of the world’s largest marine protected areas, the Chagos Archipelago. Sci. Rep. 2016, 6, 20339. [Google Scholar] [CrossRef] [Green Version]
- Esteban, N.; Laloë, J.O.; Kiggen, F.S.P.L.; Ubels, S.M.; Becking, L.E.; Meesters, E.H.; Berkel, J.; Hays, G.C.; Christianen, M.J.A. Optimism for mitigation of climate warming impacts for sea turtles through nest shading and relocation. Sci. Rep. 2018, 8, 17625. [Google Scholar] [CrossRef] [PubMed]
- Patrício, A.R.; Marques, A.; Barbosa, C.; Broderick, A.C.; Godley, B.J.; Hawkes, L.A.; Rebelo, R.; Regalla, A.; Catry, P. Balanced primary sex ratios and resilience to climate change in a major sea turtle population. Mar. Ecol. Prog. Ser. 2017, 577, 189–203. [Google Scholar] [CrossRef]
- Feagin, R.A.; Furman, M.; Salgado, K.; Martinez, M.L.; Innocenti, R.A.; Eubanks, K.; Figlus, J.; Huff, T.P.; Sigren, J.; Silva, R. The role of beach and sand dune vegetation in mediating wave run up erosion. Estuar. Coast. Shelf Sci. 2019, 219, 97–106. [Google Scholar] [CrossRef]
- Miller, T.E.; Gornish, E.S.; Buckley, H.L. Climate and coastal dune vegetation: Disturbance, recovery, and succession. Plant Ecol. 2009, 206, 97–104. [Google Scholar] [CrossRef]
- Witherington, B.E.; Bjorndal, K.A.; Bolten, A.B. An Evaluation of the Use of Dune Structure to Reduce Effects of Artificial Lighting on Hatchling Sea Turtle Sea-Finding and Offshore Orientation; Report Submitted to US Air Force; Cape Canaveral Air Force Station: Cape Canaveral, FL, USA; Florida Cooperative Fish and Wildlife Research Unit: Gainesville, FL, USA, 1994. [Google Scholar]
- Long, T.M.; Eldridge, J.; Hancock, J.; Hirama, S.; Koperski, M.; Trindell, R.N.; Long, T.M.; Eldridge, J.; Hancock, J.; Hirama, S.; et al. Balancing human and sea turtle safety: Evaluating long-wavelength streetlights as a coastal roadway management tool. Coast. Manag. 2022, 50, 184–196. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Height Profile | Mean (m) | SD | Description |
|---|---|---|---|
| Maximum | 6.23 | 2.04 | Random height from 6.1 m to 8.0 m |
| 6.1 | 5.22 | 1.71 | Maximum height leveled to 6.1 m |
| 5.8 | 5.03 | 1.61 | 6.1 m − 0.3 m |
| 5.5 | 4.80 | 1.51 | 6.1 m − 0.3 m × 2 |
| 5.2 | 4.61 | 1.43 | 6.1 m − 0.3 m × 3 |
| 4.9 | 4.37 | 1.34 | 6.1 m − 0.3 m × 4 |
| 4.6 | 4.14 | 1.27 | 6.1 m − 0.3 m × 5 |
| 4.3 | 3.95 | 1.22 | 6.1 m − 0.3 m × 6 |
| 4.0 | 3.72 | 1.17 | 6.1 m − 0.3 m × 7 |
| 3.7 | 3.53 | 1.14 | 6.1 m − 0.3 m × 8 |
| 3.4 | 3.30 | 1.13 | 6.1 m − 0.3 m × 9 |
| Prior | 3.11 | 1.90 | Height based on 2004 DSM |
| 3.1 | 3.10 | 1.13 | 6.1 m − 0.3 m × 10 |
| 2.7 | 2.87 | 1.15 | 6.1 m − 0.3 m × 11 |
| 2.4 | 2.64 | 1.19 | 6.1 m − 0.3 m × 12 |
| 2.1 | 2.45 | 1.24 | 6.1 m − 0.3 m × 13 |
| 1.8 | 2.22 | 1.31 | 6.1 m − 0.3 m × 14 |
| 1.5 | 2.02 | 1.38 | 6.1 m − 0.3 m × 15 |
| Concurrent | 1.81 | 2.06 | Height based on 2018 DSM |
| 1.2 | 1.79 | 1.47 | 6.1 m − 0.3 m × 16 |
| Trimmed | 1.53 | 1.61 | Height limited to 0.9 m or 1.1 m |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barrett, M.A.; Nelson Sella, K. Modeling Artificial Light Exposure after Vegetation Trimming at a Marine Turtle Nesting Beach. Remote Sens. 2022, 14, 2702. https://doi.org/10.3390/rs14112702
Barrett MA, Nelson Sella K. Modeling Artificial Light Exposure after Vegetation Trimming at a Marine Turtle Nesting Beach. Remote Sensing. 2022; 14(11):2702. https://doi.org/10.3390/rs14112702
Chicago/Turabian StyleBarrett, Mark A., and Kristen Nelson Sella. 2022. "Modeling Artificial Light Exposure after Vegetation Trimming at a Marine Turtle Nesting Beach" Remote Sensing 14, no. 11: 2702. https://doi.org/10.3390/rs14112702
APA StyleBarrett, M. A., & Nelson Sella, K. (2022). Modeling Artificial Light Exposure after Vegetation Trimming at a Marine Turtle Nesting Beach. Remote Sensing, 14(11), 2702. https://doi.org/10.3390/rs14112702