
The Regenerative Potential of Managed Calluna Heathlands—Revealing Optical and Structural Traits for Predicting Recovery Dynamics

 ,
, 
Abstract

1. Introduction
- A.
- The potential resprouting of Calluna heath,
- B.
- The potential threat of grass invasion in heathland sites?
2. Materials and Methods
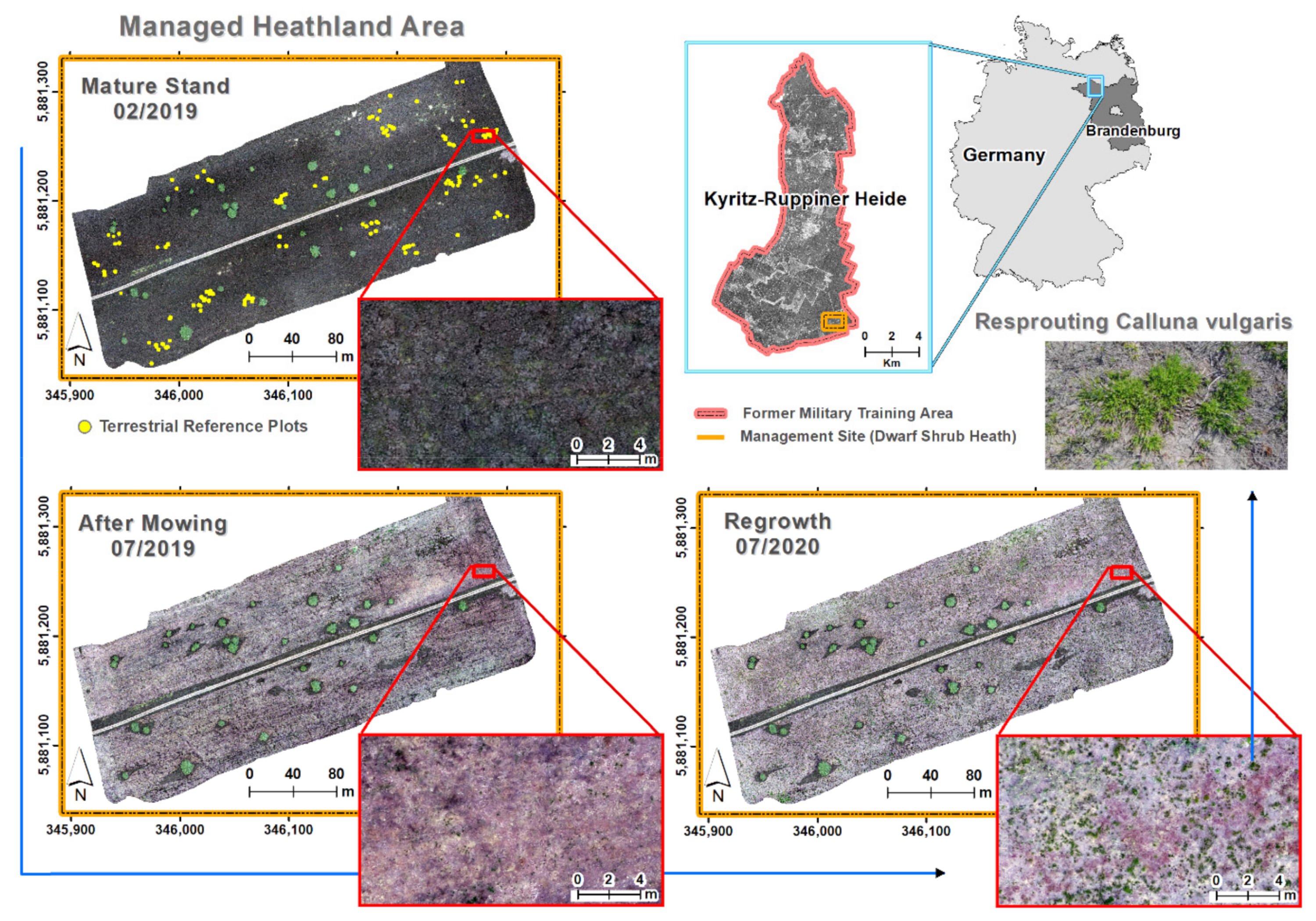
2.1. Study Area and Field Survey
2.2. UAV Image Processing
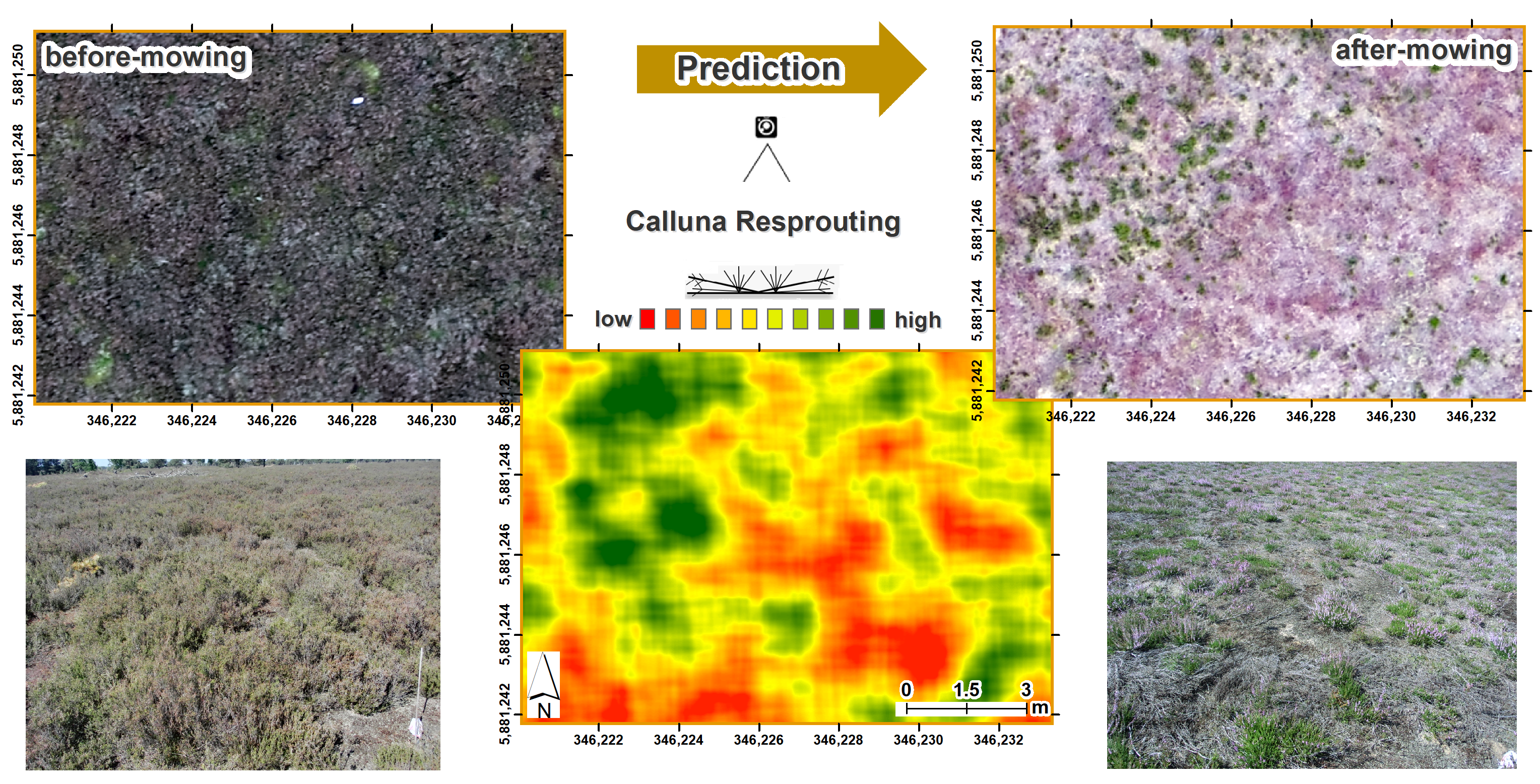
2.3. Spatial Detection of After-Mowing Dynamics
2.4. Statistical Modeling of Before-Mowing Predictors
3. Results
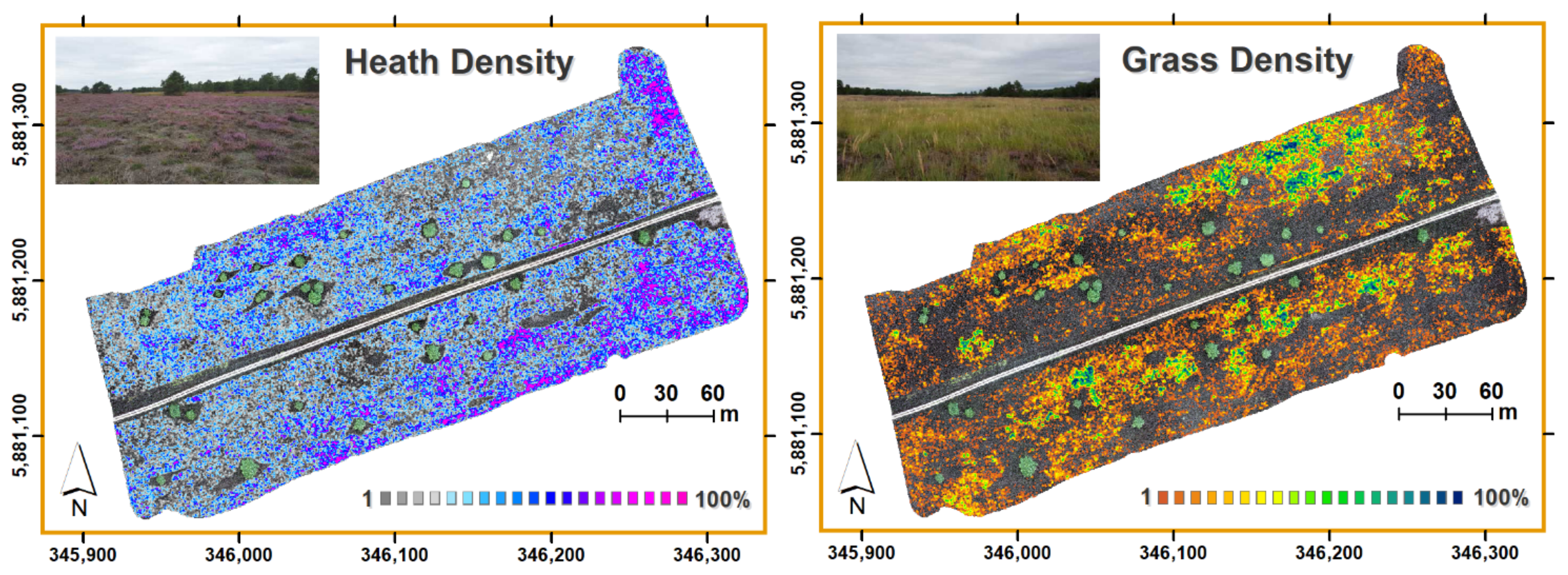
3.1. Spatial Patterns of Heathland Regeneration after Mowing
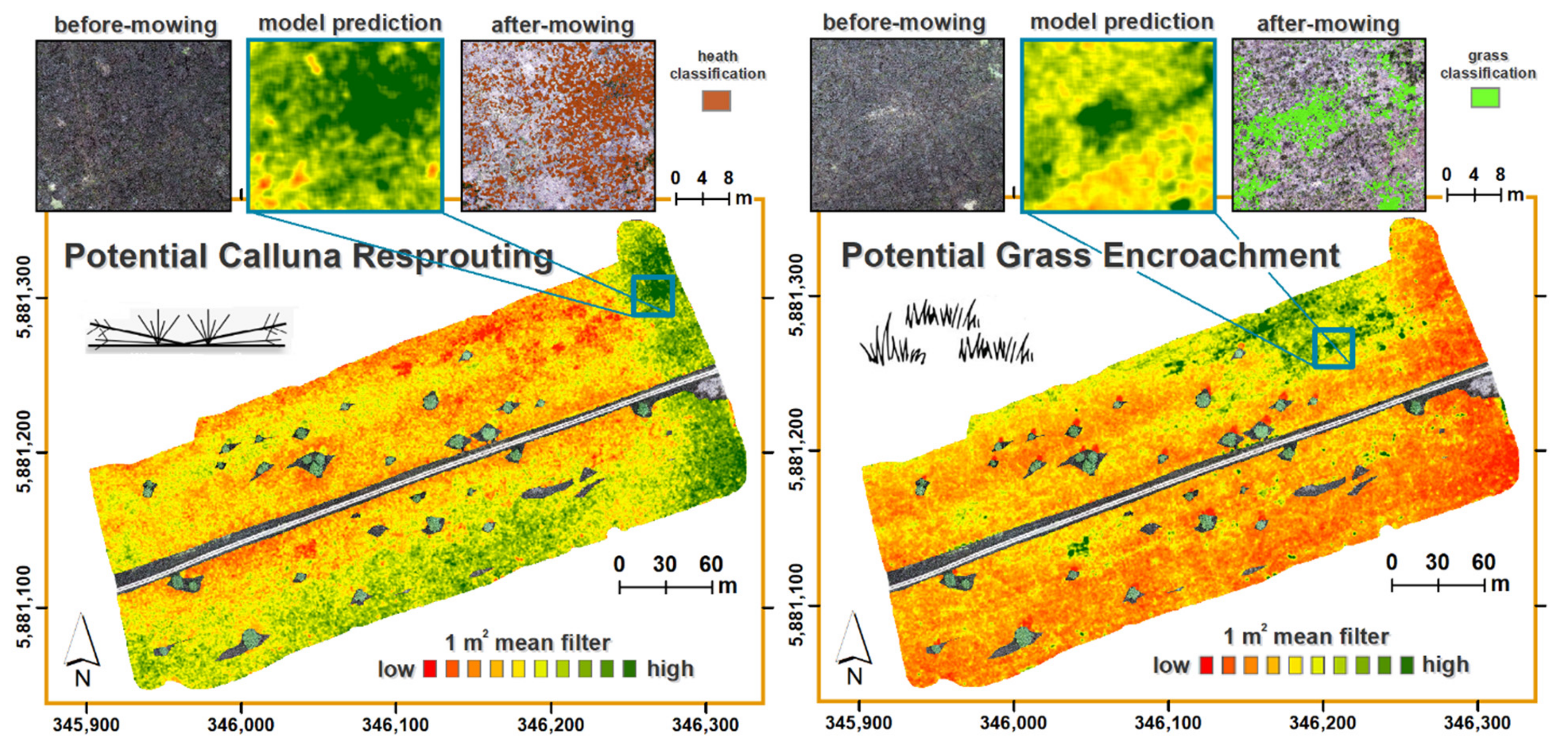
3.2. The Predictive Potential of Calluna Resprouting and Grass Encroachment
4. Discussion
4.1. The Predictive Values of Before-Mowing Stand Properties in UAV Imagery
4.2. Consequences for Nature Conservation Monitoring
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bond, W.J.; Midgley, J.J. Ecology of Sprouting in Woody Plants: The Persistence Niche. Trends Ecol. Evol. 2001, 16, 45–51. [Google Scholar] [CrossRef]
- Vesk, P.A. Plant Size and Resprouting Ability: Trading Tolerance and Avoidance of Damage? J. Ecol. 2006, 94, 1027–1034. [Google Scholar] [CrossRef]
- Pausas, J.G.; Pratt, R.B.; Keeley, J.E.; Jacobsen, A.L.; Ramirez, A.R.; Vilagrosa, A.; Paula, S.; Kaneakua-Pia, I.N.; Davis, S.D. Towards Understanding Resprouting at the Global Scale. New Phytol. 2016, 209, 945–954. [Google Scholar] [CrossRef] [PubMed]
- Pausas, J.G.; Keeley, J.E. Evolutionary Ecology of Resprouting and Seeding in Fire-Prone Ecosystems. New Phytol. 2014, 204, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Clarke, P.J.; Lawes, M.; Midgley, J.J.; Lamont, B.; Ojeda, F.; Burrows, G.; Enright, N.; Knox, K. Resprouting as a Key Functional Trait: How Buds, Protection and Resources Drive Persistence after Fire. New Phytol. 2013, 197, 19–35. [Google Scholar] [CrossRef] [PubMed]
- Zeppel, M.J.; Harrison, S.P.; Adams, H.D.; Kelley, D.I.; Li, G.; Tissue, D.T.; Dawson, T.E.; Fensham, R.; Medlyn, B.E.; Palmer, A.; et al. Drought and Resprouting Plants. New Phytol. 2015, 206, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Giljohann, K.M.; McCarthy, M.A.; Keith, D.A.; Kelly, L.T.; Tozer, M.G.; Regan, T.J. Interactions between Rainfall, Fire and Herbivory Drive Resprouter Vital Rates in a Semi-Arid Ecosystem. J. Ecol. 2017, 105, 1562–1570. [Google Scholar] [CrossRef]
- Klimešová, J.; Klimeš, L. Resprouting of Herbs in Disturbed Habitats: Is It Adequately Described by Bellingham-Sparrow’s Model? Oikos 2003, 103, 225–229. [Google Scholar] [CrossRef]
- Midgley, J. Why the World’s Vegetation Is Not Totally Dominated by Resprouting Plants; Because Resprouters Are Shorter than Reseeders. Ecography 1996, 19, 92–95. [Google Scholar] [CrossRef]
- Tälle, M.; Deák, B.; Poschlod, P.; Valkó, O.; Westerberg, L.; Milberg, P. Similar Effects of Different Mowing Frequencies on the Conservation Value of Semi-Natural Grasslands in Europe. Biodivers. Conserv. 2018, 27, 2451–2475. [Google Scholar] [CrossRef]
- Henning, K.; von Oheimb, G.; Härdtle, W.; Fichtner, A.; Tischew, S. The Reproductive Potential and Importance of Key Management Aspects for Successful Calluna Vulgaris Rejuvenation on Abandoned Continental Heaths. Ecol. Evol. 2017, 7, 2091–2100. [Google Scholar] [CrossRef]
- Bignal, E.M.; McCracken, D.I. The Nature Conservation Value of European Traditional Farming Systems. Environ. Rev. 2000, 8, 149–171. [Google Scholar] [CrossRef]
- Webb, N. The Traditional Management of European Heathlands. J. Appl. Ecol. 1998, 35, 987–990. [Google Scholar] [CrossRef]
- Fagúndez, J. Heathlands Confronting Global Change: Drivers of Biodiversity Loss from Past to Future Scenarios. Ann. Bot. 2012, 111, 151–172. [Google Scholar] [CrossRef]
- Cordingley, J.E.; Newton, A.C.; Rose, R.J.; Clarke, R.T.; Bullock, J.M. Can Landscape-Scale Approaches to Conservation Management Resolve Biodiversity–Ecosystem Service Trade-Offs? J. Appl. Ecol. 2016, 53, 96–105. [Google Scholar] [CrossRef]
- De Graaf, M.C.; Bobbink, R.; Smits, N.A.; Van Diggelen, R.; Roelofs, J.G. Biodiversity, Vegetation Gradients and Key Biogeochemical Processes in the Heathland Landscape. Biol. Conserv. 2009, 142, 2191–2201. [Google Scholar] [CrossRef]
- Schellenberg, J.; Bergmeier, E. Heathland Plant Species Composition and Vegetation Structures Reflect Soil-Related Paths of Development and Site History. Appl. Veg. Sci. 2020, 23, 386–405. [Google Scholar] [CrossRef]
- Schellenberg, J. Rethinking Relations between Age, Growth Phases and Vitality. In Proceedings of the 15th European Heathlands Network Workshop, Nijmegen, The Netherlands, 20–25 August 2017. [Google Scholar]
- Neumann, C.; Behling, R.; Schindhelm, A.; Itzerott, S.; Weiss, G.; Wichmann, M.; Müller, J. The Colors of Heath Flowering–Quantifying Spatial Patterns of Phenology in Calluna Life-Cycle Phases Using High-Resolution Drone Imagery. Remote Sens. Ecol. Conserv. 2020, 6, 35–51. [Google Scholar] [CrossRef]
- Mohamed, B.; Gimingham, C. The Morphology of Vegetative Regeneration in Calluna Vulgaris. New Phytol. 1970, 69, 743–750. [Google Scholar] [CrossRef]
- Matt Davies, G.; Adam Smith, A.; MacDonald, A.J.; Bakker, J.D.; Legg, C.J. Fire Intensity, Fire Severity and Ecosystem Response in Heathlands: Factors Affecting the Regeneration of Calluna Vulgaris. J. Appl. Ecol. 2010, 47, 356–365. [Google Scholar] [CrossRef]
- Berdowski, J.; Siepel, H. Vegetative Regeneration of Calluna Vulgaris at Different Ages and Fertilizer Levels. Biol. Conserv. 1988, 46, 85–93. [Google Scholar] [CrossRef]
- Gimingham, C.H. Ecology of Heathlands; Chapman and Hall: London, UK, 1972. [Google Scholar]
- Miller, G.; Miles, J. Regeneration of Heather (Calluna Vulgaris (L.) Hull) at Different Ages and Seasons in North-East Scotland. J. Appl. Ecol. 1970, 7, 51–60. [Google Scholar] [CrossRef]
- Sedláková, I.; Chytrỳ, M. Regeneration Patterns in a Central European Dry Heathland: Effects of Burning, Sod-Cutting and Cutting. Plant Ecol. 1999, 143, 77–87. [Google Scholar] [CrossRef]
- Anderson, K.; Gaston, K.J. Lightweight Unmanned Aerial Vehicles Will Revolutionize Spatial Ecology. Front. Ecol. Environ. 2013, 11, 138–146. [Google Scholar] [CrossRef]
- Müllerová, J.; Bartaloš, T.; Bruuna, J.; Dvořák, P.; Vítková, M. Unmanned Aircraft in Nature Conservation: An Example from Plant Invasions. Int. J. Remote Sens. 2017, 38, 2177–2198. [Google Scholar] [CrossRef]
- Díaz-Delgado, R.; Cazacu, C.; Adamescu, M. Rapid Assessment of Ecological Integrity for LTER Wetland Sites by Using UAV Multispectral Mapping. Drones 2019, 3, 3. [Google Scholar] [CrossRef]
- Baena, S.; Boyd, D.S.; Moat, J. UAVs in Pursuit of Plant Conservation-Real World Experiences. Ecol. Inform. 2018, 47, 2–9. [Google Scholar] [CrossRef]
- Klosterman, S.; Melaas, E.; Wang, J.; Martinez, A.; Frederick, S.; O’Keefe, J.; Orwig, D.A.; Wang, Z.; Sun, Q.; Schaaf, C.; et al. Fine-Scale Perspectives on Landscape Phenology from Unmanned Aerial Vehicle (UAV) Photography. Agric. For. Meteorol. 2018, 248, 397–407. [Google Scholar] [CrossRef]
- Jiménez López, J.; Mulero-Pázmány, M. Drones for Conservation in Protected Areas: Present and Future. Drones 2019, 3, 10. [Google Scholar] [CrossRef]
- EU Council Directive 92/43/EEC of 21 May 1992 on the Conservation of Natural Habitats and of Wild Fauna and Flora. J. Eur. Union 1992, 11, 7–50.
- Strecha, C.; Van Gool, L.; Fua, P. A Generative Model for True Orthorectification. In Proceedings of the ISPRS Congress, Beijing, China, 3–11 July 2008. [Google Scholar]
- Küng, O.; Strecha, C.; Beyeler, A.; Zufferey, J.-C.; Floreano, D.; Fua, P.; Gervaix, F. The Accuracy of Automatic Photogrammetric Techniques on Ultra-Light UAV Imagery. In Proceedings of the UAV-g 2011-Unmanned Aerial Vehicle in Geomatics, Zurich, Switzeland, 14–16 September 2011. [Google Scholar]
- Haralick, R.M.; Shanmugam, K.; Dinstein, I. Textural Features for Image Classification. IEEE Trans. Syst. Man Cybern. 1973, 3, 610–621. [Google Scholar] [CrossRef]
- Szantoi, Z.; Escobedo, F.; Abd-Elrahman, A.; Smith, S.; Pearlstine, L. Analyzing Fine-Scale Wetland Composition Using High Resolution Imagery and Texture Features. Int. J. Appl. Earth Obs. Geoinf. 2013, 23, 204–212. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Kaufman, Y.J.; Stark, R.; Rundquist, D. Novel Algorithms for Remote Estimation of Vegetation Fraction. Remote Sens. Environ. 2002, 80, 76–87. [Google Scholar] [CrossRef]
- Hunt, E.R., Jr.; Doraiswamy, P.C.; McMurtrey, J.E.; Daughtry, C.S.; Perry, E.M.; Akhmedov, B. A Visible Band Index for Remote Sensing Leaf Chlorophyll Content at the Canopy Scale. Int. J. Appl. Earth Obs. Geoinf. 2013, 21, 103–112. [Google Scholar] [CrossRef]
- Bendig, J.; Yu, K.; Aasen, H.; Bolten, A.; Bennertz, S.; Broscheit, J.; Gnyp, M.L.; Bareth, G. Combining UAV-Based Plant Height from Crop Surface Models, Visible, and near Infrared Vegetation Indices for Biomass Monitoring in Barley. Int. J. Appl. Earth Obs. Geoinf. 2015, 39, 79–87. [Google Scholar] [CrossRef]
- Louhaichi, M.; Borman, M.M.; Johnson, D.E. Spatially Located Platform and Aerial Photography for Documentation of Grazing Impacts on Wheat. Geocarto Int. 2001, 16, 65–70. [Google Scholar] [CrossRef]
- Woebbecke, D.M.; Meyer, G.E.; Von Bargen, K.; Mortensen, D.A. Color Indices for Weed Identification under Various Soil, Residue, and Lighting Conditions. Trans. ASAE 1995, 38, 259–269. [Google Scholar] [CrossRef]
- Hague, T.; Tillett, N.; Wheeler, H. Automated Crop and Weed Monitoring in Widely Spaced Cereals. Precis. Agric. 2006, 7, 21–32. [Google Scholar] [CrossRef]
- Marchant, J.A.; Onyango, C.M. Shadow-Invariant Classification for Scenes Illuminated by Daylight. JOSA A 2000, 17, 1952–1961. [Google Scholar] [CrossRef] [PubMed]
- Böhner, J.; Selige, T. Spatial Prediction of Soil Attributes Using Terrain Analysis and Climate Regionalisation. Göttinger Geogr. Abh. 2006, 115, 13–28. [Google Scholar]
- Moore, I.D.; Grayson, R.; Ladson, A. Digital Terrain Modelling: A Review of Hydrological, Geomorphological, and Biological Applications. Hydrol. Process. 1991, 5, 3–30. [Google Scholar] [CrossRef]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Liaw, A.; Wiener, M. Classification and Regression by RandomForest. R News 2002, 2, 18–22. [Google Scholar]
- Story, M.; Congalton, R.G. Accuracy Assessment: A User’s Perspective. Photogramm. Eng. Remote Sens. 1986, 52, 397–399. [Google Scholar]
- Cutler, D.R.; Edwards, T.C.; Beard, K.H.; Cutler, A.; Hess, K.T.; Gibson, J.; Lawler, J.J. Random Forests for Classification in Ecology. Ecology 2007, 88, 2783–2792. [Google Scholar] [CrossRef]
- Akaike, H. Information Theory and an Extension of the Maximum Likelihood Principle. In Selected Papers of Hirotugu Akaike; Springer: Berlin, Germany, 1973. [Google Scholar]
- Fox, J.; Monette, G. Generalized Collinearity Diagnostics. J. Am. Stat. Assoc. 1992, 87, 178–183. [Google Scholar] [CrossRef]
- Belkin, M.; Hsu, D.J.; Mitra, P. Overfitting or Perfect Fitting? Risk Bounds for Classification and Regression Rules That Interpolate. In Proceedings of the Advances in Neural Information Processing Systems, Montreal, QC, USA, 2–8 December 2018; pp. 2300–2311. [Google Scholar]
- Wyner, A.J.; Olson, M.; Bleich, J.; Mease, D. Explaining the Success of Adaboost and Random Forests as Interpolating Classifiers. J. Mach. Learn. Res. 2017, 18, 1558–1590. [Google Scholar]
- Watt, A.S. Bracken Versus Heather, A Study in Plant Sociology. J. Ecol. 1955, 43, 490. [Google Scholar] [CrossRef]
- Barclay-Estrup, P. The Description and Interpretation of Cyclical Processes in a Heath Community: II. Changes in Biomass and Shoot Production during the Calluna Cycle. J. Ecol. 1970, 58, 243–249. [Google Scholar] [CrossRef]
- Arroyo-Mora, J.P.; Kalacska, M.; Inamdar, D.; Soffer, R.; Lucanus, O.; Gorman, J.; Naprstek, T.; Schaaf, E.S.; Ifimov, G.; Elmer, K.; et al. Implementation of a UAV–Hyperspectral Pushbroom Imager for Ecological Monitoring. Drones 2019, 3, 12. [Google Scholar] [CrossRef]
- Mac Arthur, A.; Malthus, T. Calluna Vulgaris Foliar Pigments and Spectral Reflectance Modelling. Int. J. Remote Sens. 2012, 33, 5214–5239. [Google Scholar] [CrossRef]
- Heil, G.; Bobbink, R. Impact of atmospheric nitrogen deposition on dry heathlands. In Heathlands; Springer: Berlin, Germany, 1993; pp. 181–200. [Google Scholar]
- Neumann, C.; Itzerott, S.; Weiss, G.; Kleinschmit, B.; Schmidtlein, S. Mapping Multiple Plant Species Abundance Patterns-A Multiobjective Optimization Procedure for Combining Reflectance Spectroscopy and Species Ordination. Ecol. Inform. 2016, 36, 61–76. [Google Scholar] [CrossRef]
- Britton, A.; Carey, P.; Pakeman, R.; Marrs, R. A Comparison of Regeneration Dynamics Following Gap Creation at Two Geographically Contrasting Heathland Sites. J. Appl. Ecol. 2000, 37, 832–844. [Google Scholar] [CrossRef]
- Britton, A.J.; Pakeman, R.J.; Carey, P.D.; Marrs, R.H. Impacts of Climate, Management and Nitrogen Deposition on the Dynamics of Lowland Heathland. J. Veg. Sci. 2001, 12, 797–806. [Google Scholar] [CrossRef]
- Probo, M.; Ascoli, D.; Lonati, M.; Marzano, R.; Lombardi, G. Restoration Treatments to Control Molinia Arundinacea and Woody and Alien Species Encroachment in Calluna Vulgaris Heathlands at the Southern Edge of Their Distribution. Biol. Conserv. 2017, 211, 102–109. [Google Scholar] [CrossRef]
- Remke, E.; Brouwer, E.; Kooijman, A.; Blindow, I.; Esselink, H.; Roelofs, J.G. Even Low to Medium Nitrogen Deposition Impacts Vegetation of Dry, Coastal Dunes around the Baltic Sea. Environ. Pollut. 2009, 157, 792–800. [Google Scholar] [CrossRef] [PubMed]
- Britton, A.; Marrs, R.; Carey, P.; Pakeman, R. Comparison of Techniques to Increase Calluna Vulgaris Cover on Heathland Invaded by Grasses in Breckland, South East England. Biol. Conserv. 2000, 95, 227–232. [Google Scholar] [CrossRef]
- Buters, T.M.; Bateman, P.W.; Robinson, T.; Belton, D.; Dixon, K.W.; Cross, A.T. Methodological Ambiguity and Inconsistency Constrain Unmanned Aerial Vehicles as a Silver Bullet for Monitoring Ecological Restoration. Remote Sens. 2019, 11, 1180. [Google Scholar] [CrossRef]
- Carvajal-Ramírez, F.; Marques da Silva, J.R.; Agüera-Vega, F.; Martínez-Carricondo, P.; Serrano, J.; Moral, F.J. Evaluation of Fire Severity Indices Based on Pre-and Post-Fire Multispectral Imagery Sensed from UAV. Remote Sens. 2019, 11, 993. [Google Scholar] [CrossRef]
- Larrinaga, A.R.; Brotons, L. Greenness Indices from a Low-Cost UAV Imagery as Tools for Monitoring Post-Fire Forest Recovery. Drones 2019, 3, 6. [Google Scholar] [CrossRef]
- Buters, T.M.; Belton, D.; Cross, A.T. Multi-Sensor UAV Tracking of Individual Seedlings and Seedling Communities at Millimetre Accuracy. Drones 2019, 3, 81. [Google Scholar] [CrossRef]
- Gimingham, C. A Reappraisal of Cyclical Processes in Calluna Heath. Vegetatio 1988, 77, 61–64. [Google Scholar] [CrossRef]
- Schmidt, J.; Fassnacht, F.E.; Neff, C.; Lausch, A.; Kleinschmit, B.; Förster, M.; Schmidtlein, S. Adapting a Natura 2000 Field Guideline for a Remote Sensing-Based Assessment of Heathland Conservation Status. Int. J. Appl. Earth Obs. Geoinf. 2017, 60, 61–71. [Google Scholar] [CrossRef]
- Barker, C.G.; Power, S.A.; Bell, J.; Orme, C. Effects of Habitat Management on Heathland Response to Atmospheric Nitrogen Deposition. Biol. Conserv. 2004, 120, 41–52. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| After-Mowing (n-Layers = 15) | Before-Mowing (n-Layers = 48) | DEM (n-Layers = 2) | ||||
|---|---|---|---|---|---|---|
| image dates | 24/07/2020 | 08/09/2020 | 02/10/2020 | 06/04/2018 | 13/02/2019 | |
| optical traits | ||||||
| RGB grey-values | - | - | - | 3 | 3 | - |
| RGB-HSV transformed | 3 | 3 | 3 | 3 | 3 | - |
| RGB chromaticity | - | - | - | 3 | 3 | - |
| spectral indices | - | - | - | 8 | 8 | - |
| structural traits | - | |||||
| green texture | - | 6 | - | 6 | 6 | - |
| canopy height | - | - | - | 1 | 1 | - |
| micro-relief | - | |||||
| topographic wetness index | - | - | - | - | - | 1 |
| slope-length factor | - | - | - | - | - | 1 |
| Index | Name | Equation | Citation |
|---|---|---|---|
| VARI | Visible Atmospheric Resistant Index | (G - R) / (G + R -B) | [37] |
| TGI | Triangular Greeness Index | G - 0.39 · R - 0.61 · B | [38] |
| NGRDI | Normalized Green Red Difference Index | (G - R) / (G + R) | [37] |
| MGRVI | Modified Green Red Vegetation Index | (G2 - R2) / (G2 + R2) | [39] |
| RGBVI | Red Green Blue Vegetation Index | G2 - (R · B) / G2 + (R · B) | [39] |
| GLI | Green Leaf Index | (2 · G – R – B) / (2 · G + R + B) | [40] |
| EXG | Excess Green Index | 2 · G - B - R | [41] |
| DAVI | Daylight Adapted Vegetation Index | G/((R0.667) · (B(1 - 0.667))) | [42,43] |
| Class | Individuals | n-Pixels per Layer |
|---|---|---|
| Calluna | Calluna vulgaris | 5292 |
| Grass | Calamagrostis epigejos, Agrostis capillaris, Carex arenaria; Carex pilulifera, Nardus stricta, Deschampsia flexuosa, Corynephorus canescens | 3360 |
| Herb | Rumex acetosella, Hypericum perforatum, Hieracium pilosella | 1753 |
| Litter | Calluna vulgaris (dead, senescent branches) | 2090 |
| Cryptogams | Cladonia mitis, Cladonia coccifera, Cladonia furcata, Pleurozium schreberi, Hypnum jutlandicum, Polytrichum piliferum | 2271 |
| Shrub | Rubus spec., Populus tremula, Betula pendula, Pinus sylvestris | 3030 |
| Background | open soil substrate | 1368 |
| Classification | |||||||||
| Calluna | Grass | Herb | Litter | Crypto | Shrub | Backgr. | Producer’s Accuracy (%) | ||
| Reference | Calluna | 5244 | 11 | 2 | 8 | 18 | 9 | 0 | 99.09 |
| Grass | 26 | 3309 | 10 | 6 | 9 | 45 | 0 | 97.18 | |
| Herb | 10 | 10 | 1713 | 2 | 18 | 0 | 0 | 97.72 | |
| Litter | 16 | 20 | 0 | 2029 | 15 | 0 | 10 | 97.08 | |
| Crypto | 29 | 30 | 2 | 7 | 2201 | 0 | 2 | 96.92 | |
| Shrub | 52 | 12 | 2 | 0 | 0 | 2964 | 0 | 97.82 | |
| Backgr. | 0 | 8 | 0 | 12 | 2 | 0 | 1354 | 98.40 | |
| User’s accuracy (%) | 97.53 | 97.32 | 99.07 | 98.3 | 97.26 | 98.21 | 99.12 | ||
| R2 | RMSE (%) | Selected Predictors | |
|---|---|---|---|
| (a) Calluna resprouting | |||
| Multivariate | 0.15 | 21.6 | mean.green06/04/2018, hsv.green06/04/2018, DAVI06/04/2018 |
| Random Forest | 0.17 | 20.1 | mean.green06/04/2018, green06/04/2018, variance.green06/04/2018 |
| (b) Grass encroachment | |||
| Multivariate | 0.31 | 18.64 | dissimilarity13/02/2019, nDSM13/02/2019, hsv.red06/04/2018, dissimilarity06/04/2018, second.moment06/04/2018, TWI06/04/2018 |
| Random Forest | 0.38 | 17.00 | red13/02/2019, red06/04/2018, RGBVI06/04/2018, variance06/04/2018, nDSM13/02/2019, variance13/02/2019 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neumann, C.; Schindhelm, A.; Müller, J.; Weiss, G.; Liu, A.; Itzerott, S. The Regenerative Potential of Managed Calluna Heathlands—Revealing Optical and Structural Traits for Predicting Recovery Dynamics. Remote Sens. 2021, 13, 625. https://doi.org/10.3390/rs13040625
Neumann C, Schindhelm A, Müller J, Weiss G, Liu A, Itzerott S. The Regenerative Potential of Managed Calluna Heathlands—Revealing Optical and Structural Traits for Predicting Recovery Dynamics. Remote Sensing. 2021; 13(4):625. https://doi.org/10.3390/rs13040625
Chicago/Turabian StyleNeumann, Carsten, Anne Schindhelm, Jörg Müller, Gabriele Weiss, Anna Liu, and Sibylle Itzerott. 2021. "The Regenerative Potential of Managed Calluna Heathlands—Revealing Optical and Structural Traits for Predicting Recovery Dynamics" Remote Sensing 13, no. 4: 625. https://doi.org/10.3390/rs13040625
APA StyleNeumann, C., Schindhelm, A., Müller, J., Weiss, G., Liu, A., & Itzerott, S. (2021). The Regenerative Potential of Managed Calluna Heathlands—Revealing Optical and Structural Traits for Predicting Recovery Dynamics. Remote Sensing, 13(4), 625. https://doi.org/10.3390/rs13040625