
Estimation of Rangeland Production in the Arid Oriental Region (Morocco) Combining Remote Sensing Vegetation and Rainfall Indices: Challenges and Lessons Learned


Abstract

1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Field Data Collection
2.3. Remote Sensing Data Processing
2.4. Climatic Data Processing
2.5. Models and Statistical Analysis
3. Results
3.1. Characteristics of Vegetation and Climate during the Period of Interest
3.2. Global Model for Estimation of Rangeland Biomass
3.3. Linear Relation between Biomass and ARVI
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Middleton, N.; Thomas, D.S.G. World Atlas of Desertification, 2nd ed.; Arnold: London, UK; John Wiley: Hoboken, NJ, USA, 1997. [Google Scholar]
- Weber, K.T.; Horst, S. Desertification and Livestock Grazing: The Roles of Sedentarization, Mobility and Rest. Pastor Res Policy Pract. 2011, 1, 19. [Google Scholar] [CrossRef]
- Scholes, R.J. The Future of Semi-Arid Regions: A Weak Fabric Unravels. Climate 2020, 8, 43. [Google Scholar] [CrossRef]
- Safriel, U.; Adeel, Z.; Niemeijer, D.; Puigdefabregas, J.; White, R.; Lal, R.; Winslow, M.; Ziedler, J.; Prince, S.; Archer, E.; et al. Dryland Systems. In Global Assessment Reports Volume 1: Current State and Trends; Millennium Ecosystem Assessment; Island Press: Washington, DC, USA, 2005. [Google Scholar]
- Huang, J.; Ji, M.; Xie, Y.; Wang, S.; He, Y.; Ran, J. Global Semi-Arid Climate Change over Last 60 Years. Clim. Dyn. 2016, 46, 1131–1150. [Google Scholar] [CrossRef]
- Eisfelder, C.; Kuenzer, C.; Dech, S. Derivation of Biomass Information for Semi-Arid Areas Using Remote-Sensing Data. Int. J. Remote Sens. 2012, 33, 2937–2984. [Google Scholar] [CrossRef]
- Dong, S. Overview: Pastoralism in the World. In Building Resilience of Human-Natural Systems of Pastoralism in the Developing World; Dong, S., Kassam, K.-A.S., Tourrand, J.F., Boone, R.B., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 1–38. ISBN 978-3-319-30730-5. [Google Scholar]
- Rishmawi, K.; Prince, S.; Xue, Y. Vegetation Responses to Climate Variability in the Northern Arid to Sub-Humid Zones of Sub-Saharan Africa. Remote Sens. 2016, 8, 910. [Google Scholar] [CrossRef]
- Sloat, L.L.; Gerber, J.S.; Samberg, L.H.; Smith, W.K.; Herrero, M.; Ferreira, L.G.; Godde, C.M.; West, P.C. Increasing Importance of Precipitation Variability on Global Livestock Grazing Lands. Nat. Clim Chang. 2018, 8, 214–218. [Google Scholar] [CrossRef]
- Godde, C.; Dizyee, K.; Ash, A.; Thornton, P.; Sloat, L.; Roura, E.; Henderson, B.; Herrero, M. Climate Change and Variability Impacts on Grazing Herds: Insights from a System Dynamics Approach for Semi-arid Australian Rangelands. Glob Chang. Biol 2019, 25, 3091–3109. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2013: The Physical Science Basis; Stocker, T.F.D., Qin, G.-K., Plattner, M., Tignor, S.K., Allen, J., Boschung, A., Nauels, Y., Xia, V.B., Midgley, P.M., Eds.; Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013. [Google Scholar]
- Food and Agriculture Organization of the United Nations. Pastoralism in Africa’s Drylands: Reducing Risks, Addressing Vulnerability and Enhancing Resilience; FAO: Rome, Italy, 2018. [Google Scholar]
- Koocheki, A.; Gliessman, S.R. Pastoral Nomadism, a Sustainable System for Grazing Land Management in Arid Areas. J. Sustain. Agric. 2005, 25, 113–131. [Google Scholar] [CrossRef]
- Boutaleb, A.; Firmian, I. Community Governance Of Natural Resources And Rangelands: The Case Of The Eastern Highlands Of Morocco. In The Governance of Rangelands—Collective Action for Sustainable Pastoralism; Routledge: London, UK, 2014; pp. 94–107. ISBN 978-1-315-76801-4. [Google Scholar]
- Dong, S.; Liu, S.; Wen, L. Vulnerability and Resilience of Human-Natural Systems of Pastoralism Worldwide. In Building Resilience of Human-Natural Systems of Pastoralism in the Developing World; Dong, S., Kassam, K.-A.S., Tourrand, J.F., Boone, R.B., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 39–92. ISBN 978-3-319-30730-5. [Google Scholar]
- Mahyou, H.; Tychon, B.; Balaghi, R.; Louhaichi, M.; Mimouni, J. A Knowledge-Based Approach for Mapping Land Degradation in the Arid Rangelands of North Africa: Mapping Land Degradation in the Arid Rangelands. Land Degrad. Dev. 2016, 27, 1574–1585. [Google Scholar] [CrossRef]
- Reinermann, S.; Asam, S.; Kuenzer, C. Remote Sensing of Grassland Production and Management—A Review. Remote Sens. 2020, 12, 1949. [Google Scholar] [CrossRef]
- Tucker, C.J. Red and Photographic Infrared Linear. Combinations for Monitoring Vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the Radiometric and Biophysical Performance of the MODIS Vegetation Indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Huete, A.R. A Soil-Adjusted Vegetation Index (SAVI). Remote Sens. Environ. 1988, 25, 295–309. [Google Scholar] [CrossRef]
- Baret, F.; Guyot, G.; Major, D.J. TSAVI: A Vegetation Index Which Minimizes Soil Brightness Effects On LAI And APAR Estimation. In Proceedings of the 12th Canadian Symposium on Remote Sensing Geoscience and Remote Sensing Symposium, Vancouver, BC, Canada, 10–14 July 1989; Volume 3, pp. 1355–1358. [Google Scholar]
- Qi, J.; Chehbouni, A.; Huete, A.R.; Kerr, Y.H.; Sorooshian, S. A Modified Soil Adjusted Vegetation Index. Remote Sens. Environ. 1994, 48, 119–126. [Google Scholar] [CrossRef]
- Rondeaux, G.; Steven, M.; Baret, F. Optimization of Soil-Adjusted Vegetation Indices. Remote Sens. Environ. 1996, 55, 95–107. [Google Scholar] [CrossRef]
- Hatfield, J.L.; Prueger, J.H.; Sauer, T.J.; Dold, C.; O’Brien, P.; Wacha, K. Applications of Vegetative Indices from Remote Sensing to Agriculture: Past and Future. Inventions 2019, 4, 71. [Google Scholar] [CrossRef]
- Ali, I.; Cawkwell, F.; Dwyer, E.; Barrett, B.; Green, S. Satellite Remote Sensing of Grasslands: From Observation to Management. J. Plant Ecol. 2016, 9, 649–671. [Google Scholar] [CrossRef]
- Baret, F.; Guyot, G. Potentials and Limits of Vegetation Indices for LAI and APAR Assessment. Remote Sens. Environ. 1991, 35, 161–173. [Google Scholar] [CrossRef]
- Diallo, O.; Diouf, A.; Hanan, N.P.; Ndiaye, A.; Prévost, Y. AVHRR Monitoring of Savanna Primary Production in Senegal, West Africa: 1987–1988. Int. J. Remote Sens. 1991, 12, 1259–1279. [Google Scholar] [CrossRef]
- Diouf, A.; Lambin, E.F. Monitoring Land-Cover Changes in Semi-Arid Regions: Remote Sensing Data and Field Observations in the Ferlo, Senegal. J. Arid Environ. 2001, 48, 129–148. [Google Scholar] [CrossRef]
- Schucknecht, A.; Meroni, M.; Kayitakire, F.; Boureima, A. Phenology-Based Biomass Estimation to Support Rangeland Management in Semi-Arid Environments. Remote Sens. 2017, 9, 463. [Google Scholar] [CrossRef]
- Mahyou, H.; Tychon, B.; Lang, M.; Balaghi, R. Phytomass Estimation Using EMODIS NDVI and Ground Data in Arid Rangelands of Morocco. Afr. J. Range Forage Sci. 2018, 35, 1–12. [Google Scholar] [CrossRef]
- Mahyou, H. Estimation de la production fourragère des terres de parcours des hauts plateaux de l’oriental (Maroc) par les indices de télédétection. Afr. Mediterr. Agric. Res. J. Al-Awamia 2020, 19, 128. [Google Scholar]
- Benseghir, L.; Bachari, N.E.I. Shortwave Infrared Vegetation Index-Based Modelling for Aboveground Vegetation Biomass Assessment in the Arid Steppes of Algeria. Afr. J. Range Forage Sci. 2021, 1–10. [Google Scholar] [CrossRef]
- Chen, F.; Weber, K.T.; Gokhale, B. Herbaceous Biomass Estimation from SPOT 5 Imagery in Semiarid Rangelands of Idaho. Giscience Remote Sens. 2011, 48, 195–209. [Google Scholar] [CrossRef]
- Numata, I.; Roberts, D.A.; Chadwick, O.A.; Schimel, J.; Sampaio, F.R.; Leonidas, F.C.; Soares, J.V. Characterization of Pasture Biophysical Properties and the Impact of Grazing Intensity Using Remotely Sensed Data. Remote Sens. Environ. 2007, 109, 314–327. [Google Scholar] [CrossRef]
- Mundava, C.; Helmholz, P.; Schut, A.G.T.; Corner, R.; McAtee, B.; Lamb, D.W. Evaluation of Vegetation Indices for Rangeland Biomass Estimation in the Kimberley Area of Western Australia. ISPRS Ann. Photogramm. Remote Sens. Spat. Inf. Sci. 2014, II–7, 47–53. [Google Scholar] [CrossRef]
- Pordel, F.; Ebrahimi, A.; Azizi, Z. Canopy Cover or Remotely Sensed Vegetation Index, Explanatory Variables of above-Ground Biomass in an Arid Rangeland, Iran. J. Arid Land 2018, 10, 767–780. [Google Scholar] [CrossRef]
- Hadian, F.; Jafari, R.; Bashari, H.; Tarkesh, M.; Clarke, K.D. Effects of Drought on Plant Parameters of Different Rangeland Types in Khansar Region, Iran. Arab. J. Geosci 2019, 12, 93. [Google Scholar] [CrossRef]
- Gholami Baghi, N.; Oldeland, J. Do Soil-Adjusted or Standard Vegetation Indices Better Predict above Ground Biomass of Semi-Arid, Saline Rangelands in North-East Iran? Int. J. Remote Sens. 2019, 40, 8223–8235. [Google Scholar] [CrossRef]
- Xie, Y.; Sha, Z.; Yu, M.; Bai, Y.; Zhang, L. A Comparison of Two Models with Landsat Data for Estimating above Ground Grassland Biomass in Inner Mongolia, China. Ecol. Model. 2009, 220, 1810–1818. [Google Scholar] [CrossRef]
- John, R.; Chen, J.; Giannico, V.; Park, H.; Xiao, J.; Shirkey, G.; Ouyang, Z.; Shao, C.; Lafortezza, R.; Qi, J. Grassland Canopy Cover and Aboveground Biomass in Mongolia and Inner Mongolia: Spatiotemporal Estimates and Controlling Factors. Remote Sens. Environ. 2018, 213, 34–48. [Google Scholar] [CrossRef]
- Otgonbayar, M.; Atzberger, C.; Chambers, J.; Damdinsuren, A. Mapping Pasture Biomass in Mongolia Using Partial Least Squares, Random Forest Regression and Landsat 8 Imagery. Int. J. Remote Sens. 2019, 40, 3204–3226. [Google Scholar] [CrossRef]
- Kaufman, Y.J.; Tanre, D. Atmospherically Resistant Vegetation Index (ARVI) for EOS-MODIS. IEEE Trans. Geosci. Remote Sens. 1992, 30, 261–270. [Google Scholar] [CrossRef]
- Huete, A.; Justice, C.; Liu, H. Development of Vegetation and Soil Indices for MODIS-EOS. Remote Sens. Environ. 1994, 49, 224–234. [Google Scholar] [CrossRef]
- Penuelas, J.; Baret, F.; Filella, I. Semi-Empirical Indices to Assess Carotenoids/Chlorophyll a Ratio from Leaf Spectral Reflectance. Photosynthetica. 1995, 31, 221–230. [Google Scholar]
- Marticorena, B.; Bergametti, G. Modeling the Atmospheric Dust Cycle: 1. Design of a Soil-derived Dust Emission Scheme. J. Geophys. Res. 1995, 100, 16415–16430. [Google Scholar] [CrossRef]
- Tegen, I.; Schepanski, K. The Global Distribution of Mineral Dust. Iop Conf. Ser. Earth Environ. Sci. 2009, 7, 012001. [Google Scholar] [CrossRef]
- Gao, B. NDWI—A Normalized Difference Water Index for Remote Sensing of Vegetation Liquid Water from Space. Remote Sens. Environ. 1996, 58, 257–266. [Google Scholar] [CrossRef]
- Lotsch, A.; Friedl, M.A.; Anderson, B.T.; Tucker, C.J. Coupled Vegetation-Precipitation Variability Observed from Satellite and Climate Records: Vegetation-Precipitation Dynamics. Geophys. Res. Lett. 2003, 30. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M. Evaluating the Impact of Drought Using Remote Sensing in a Mediterranean, Semi-Arid Region. Nat. Hazards 2007, 40, 173–208. [Google Scholar] [CrossRef]
- Sierra-Soler, A.; Adamowski, J.; Malard, J.; Qi, Z.; Saadat, H.; Pingale, S. Assessing Agricultural Drought at a Regional Scale Using LULC Classification, SPI, and Vegetation Indices: Case Study in a Rainfed Agro-Ecosystem in Central Mexico. Geomat. Nat. Hazards Risk 2016, 7, 1460–1488. [Google Scholar] [CrossRef]
- Hua, L.; Wang, H.; Sui, H.; Wardlow, B.; Hayes, M.J.; Wang, J. Mapping the Spatial-Temporal Dynamics of Vegetation Response Lag to Drought in a Semi-Arid Region. Remote Sens. 2019, 11, 1873. [Google Scholar] [CrossRef]
- Reddy, G.P.O.; Kumar, N.; Sahu, N.; Srivastava, R.; Singh, S.K.; Naidu, L.G.K.; Chary, G.R.; Biradar, C.M.; Gumma, M.K.; Reddy, B.S.; et al. Assessment of Spatio-Temporal Vegetation Dynamics in Tropical Arid Ecosystem of India Using MODIS Time-Series Vegetation Indices. Arab. J. Geosci. 2020, 13, 704. [Google Scholar] [CrossRef]
- Nandintsetseg, B.; Shinoda, M. Assessment of Drought Frequency, Duration, and Severity and Its Impact on Pasture Production in Mongolia. Nat. Hazards 2013, 14, 995–1008. [Google Scholar] [CrossRef]
- Diouf, A.; Hiernaux, P.; Brandt, M.; Faye, G.; Djaby, B.; Diop, M.; Ndione, J.; Tychon, B. Do Agrometeorological Data Improve Optical Satellite-Based Estimations of the Herbaceous Yield in Sahelian Semi-Arid Ecosystems? Remote Sens. 2016, 8, 668. [Google Scholar] [CrossRef]
- McKee, T.B.; Doesken, N.J.; Kleist, J. The Relationship Of Drought Frequency And Duration To Time Scales. In Proceedings of the Eighth Conference on Applied Climatology, Anaheim, CA, USA, 17–22 January 1993. [Google Scholar]
- Edwards, D.C. Characteristics of 20th Century Drought in the United States at Multiple Time Scales. Master’s Thesis, Colorado State University, Fort Collins, CO, USA, 1997. [Google Scholar]
- Horion, S.; Carrão, H.; Singleton, A.; Barbosa, P.; Vogt, J. JRC Experience on the Development of Drought Information Systems: Europe, Africa and Latin America; Publications Office of the European Union: Luxembourg, 2012. [Google Scholar] [CrossRef]
- World Meteorological Organization. Standardized Precipitation Index User Guide; World Meteorological Organization: Geneva, Switzerland, 2012; ISBN 978-92-63-11091-6. [Google Scholar]
- Liu, L.; Zhang, Y.; Wu, S.; Li, S.; Qin, D. Water Memory Effects and Their Impacts on Global Vegetation Productivity and Resilience. Sci. Rep. 2018, 8, 2962. [Google Scholar] [CrossRef]
- Guerschman, J.P.; Hill, M.J.; Leys, J.; Heidenreich, S. Vegetation Cover Dependence on Accumulated Antecedent Precipitation in Australia: Relationships with Photosynthetic and Non-Photosynthetic Vegetation Fractions. Remote Sens. Environ. 2020, 240, 111670. [Google Scholar] [CrossRef]
- Verrelst, J.; Camps-Valls, G.; Muñoz-Marí, J.; Rivera, J.P.; Veroustraete, F.; Clevers, J.G.P.W.; Moreno, J. Optical Remote Sensing and the Retrieval of Terrestrial Vegetation Bio-Geophysical Properties—A Review. ISPRS J. Photogramm. Remote Sens. 2015, 108, 273–290. [Google Scholar] [CrossRef]
- Ali, I.; Greifeneder, F.; Stamenkovic, J.; Neumann, M.; Notarnicola, C. Review of Machine Learning Approaches for Biomass and Soil Moisture Retrievals from Remote Sensing Data. Remote Sens. 2015, 7, 16398–16421. [Google Scholar] [CrossRef]
- Weiss, M.; Jacob, F.; Duveiller, G. Remote Sensing for Agricultural Applications: A Meta-Review. Remote Sens. Environ. 2020, 236, 111402. [Google Scholar] [CrossRef]
- Zhu, X.X.; Tuia, D.; Mou, L.; Xia, G.-S.; Zhang, L.; Xu, F.; Fraundorfer, F. Deep Learning in Remote Sensing: A Comprehensive Review and List of Resources. IEEE Geosci. Remote Sens. Mag. 2017, 5, 8–36. [Google Scholar] [CrossRef]
- Ma, L.; Liu, Y.; Zhang, X.; Ye, Y.; Yin, G.; Johnson, B.A. Deep Learning in Remote Sensing Applications: A Meta-Analysis and Review. ISPRS J. Photogramm. Remote Sens. 2019, 152, 166–177. [Google Scholar] [CrossRef]
- Ball, J.E.; Anderson, D.T.; Chan, C.S. Comprehensive Survey of Deep Learning in Remote Sensing: Theories, Tools, and Challenges for the Community. J. Appl. Remote Sens. 2017, 11, 1. [Google Scholar] [CrossRef]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Hothorn, T. Survival Ensembles. Biostatistics 2005, 7, 355–373. [Google Scholar] [CrossRef]
- Strobl, C.; Boulesteix, A.-L.; Zeileis, A.; Hothorn, T. Bias in Random Forest Variable Importance Measures: Illustrations, Sources and a Solution. BMC Bioinform. 2007, 8, 25. [Google Scholar] [CrossRef]
- Strobl, C.; Boulesteix, A.-L.; Kneib, T.; Augustin, T.; Zeileis, A. Conditional Variable Importance for Random Forests. BMC Bioinform. 2008, 9, 307. [Google Scholar] [CrossRef]
- Bjorn, S. Thematic Mapping API. TM_WORLD_BORDERS-0.3. Available online: https://thematicmapping.org/downloads/world_borders.php (accessed on 12 May 2021).
- OpenAfrica. Morocco—Road Network. Available online: https://open.africa/fr/dataset/morocco-maps/resource/ee9a2c6e-95d8-4de0-b525-6725f439b8a5 (accessed on 12 February 2021).
- Jarvis, A.; Reuter, H.I.; Nelson, A.; Guevara, E. Hole-Filled SRTM for the Globe Version 4, available from the CGIAR-CSI SRTM 90m Database. 2008. Available online: http://srtm.csi.cgiar.org (accessed on 22 April 2021).
- Bechchari, A.; Aich, A.E.; Mahyou, H.; Baghdad, B.; Bendaou, M. Etude de la dégradation des pâturages steppiques dans les communes de Maâtarka et Béni Mathar (Maroc oriental). J. Mater. Environ. Sci. 2014, 5, 2572–2583. [Google Scholar]
- Acherkouk, M.; El Houmaizi, M.A. Évaluation de l’impact des aménagements pastoraux sur la dynamique de la production des pâturages dégradés au Maroc oriental. Ecol. Mediterr. 2013, 39, 69–84. [Google Scholar] [CrossRef]
- Kirmse, R.D.; Norton, B.E. Comparison of the Reference Unit Method and Dimensional Analysis Methods for Two Large Shrubby Species in the Caatinga Woodlands. J. Range Manag. 1985, 38, 425. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 22 April 2021).
- United States Geological Survey, Department of the Interior. USGS EROS Archive—Landsat Archives—Landsat 7 ETM+ Level-2 Data Products—Surface Reflectance. Available online: https://www.usgs.gov/centers/eros/science/usgs-eros-archive-landsat-archives-landsat-7-etm-level-2-data-products-surface?qt-science_center_objects=0#qt-science_center_objects (accessed on 5 July 2019).
- United States Geological Survey, Department of the Interior Landsat. 4-7 Collection 1 Surface Reflectance Code LEDAPS Product Guide. Available online: https://www.usgs.gov/media/files/landsat-4-7-collection-1-surface-reflectance-code-ledaps-product-guide (accessed on 22 April 2021).
- United States Geological Survey, Department of the Interior. Landsat—Earth Observation Satellites. Fact Sheet 2015-3081; U.S. Geological Survey. 2016. Available online: https://doi.org/10.3133/fs20153081 (accessed on 22 April 2021).
- Xu, D.; Guo, X. A Study of Soil Line Simulation from Landsat Images in Mixed Grassland. Remote Sens. 2013, 5, 4533–4550. [Google Scholar] [CrossRef]
- Ahmadian, N.; Demattê, J.; Xu, D.; Borg, E.; Zölitz, R. A New Concept of Soil Line Retrieval from Landsat 8 Images for Estimating Plant Biophysical Parameters. Remote Sens. 2016, 8, 738. [Google Scholar] [CrossRef]
- Koenker, R. Quantreg: Quantile Regression; R Package Version 5.73. Available online: https://CRAN.R-project.org/package=quantreg (accessed on 22 April 2021).
- Funk, C.; Peterson, P.; Landsfeld, M.; Pedreros, D.; Verdin, J.; Shukla, S.; Husak, G.; Rowland, J.; Harrison, L.; Hoell, A.; et al. The Climate Hazards Infrared Precipitation with Stations—A New Environmental Record for Monitoring Extremes. Sci Data 2015, 2, 150066. [Google Scholar] [CrossRef] [PubMed]
- Povoa, L.V.; Nery, J.T. Precintcon: Precipitation Intensity, Concentration and Anomaly Analysis. R Package Version 4.0.2. Available online: https://cran.r-project.org/web/packages/precintcon/index.html (accessed on 11 May 2020).
- Khun, M. Caret: Classification and Regression Training; 2020; R Package Version 6.0-86. Available online: https://CRAN.R-project.org/package=caret (accessed on 22 April 2021).
- Strobl, C.; Malley, J.; Tutz, G. An Introduction to Recursive Partitioning: Rationale, Application and Characteristics of Classification and Regression Trees, Bagging and Random Forests. Psychol. Methods 2010, 14, 323. [Google Scholar] [CrossRef]
- Jacques, D.C.; Kergoat, L.; Hiernaux, P.; Mougin, E.; Defourny, P. Monitoring Dry Vegetation Masses in Semi-Arid Areas with MODIS SWIR Bands. Remote Sens. Environ. 2014, 153, 40–49. [Google Scholar] [CrossRef]
- Louhaichi, M.; Gamoun, M. Stipa Tenacissima: Nurse Species to Initiate the Process of Ecosystem Restoration. International Center for Agricultural Research in the Dry Areas (ICARDA), Beirut, Lebanon. 2017. Available online: https://hdl.handle.net/20.500.11766/8567 (accessed on 22 April 2021).
- Chen, Y.; Gillieson, D. Evaluation of Landsat TM Vegetation Indices for Estimating Vegetation Cover on Semi-Arid Rangelands: A Case Study from Australia. Can. J. Remote Sens. 2009, 35, 12. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Name | Formula | Ref. |
|---|---|---|---|
| NIR/R | - | ||
| NIR/SWIR1 | - | ||
| NIR/SWIR2 | - | ||
| SWIR1/SWIR2 | - | ||
| NDVI | Normalized Difference Vegetation Index | [18] | |
| SAVI | Soil-Adjusted Vegetation Index | With L = 1 | [20] |
| TSAVI | Transformed Soil-Adjusted Vegetation Index | With: - s = 1.14 (slope of the soil line) - i = −0.002 (intercept of the soil line) - Χ = 0.08 | [21,26] |
| MSAVI | Modified Soil-Adjusted Vegetation Index | With: - - s = 1.14 (slope of the soil line) | [22] |
| OSAVI | Optimized Soil-Adjusted Vegetation Index | [23] | |
| EVI | Enhanced Vegetation Index | With: - G = 2.5 (gain factor) - L = 1 (canopy background adjustment factor) - C1 = 6 and C2 = 7.5 (coefficients of the aerosol resistance term) | [19] |
| ARVI | Atmospherically Resistant Vegetation Index | With: - - | [42,43] |
| SIPI | Structure Insensitive Pigment Index | [44] | |
| NDWI1 | Normalized Difference Water Index | [47] | |
| NDWI2 | Normalized difference water index | [47] |
| Variable | Pearson | Spearman |
|---|---|---|
| ARVI | 0.77 (***) | 0.72 (***) |
| NIR/R | 0.73 (***) | 0.65 (***) |
| NDVI | 0.72 (***) | 0.65 (***) |
| EVI | 0.70 (***) | 0.69 (***) |
| TSAVI | 0.70 (***) | 0.65 (***) |
| OSAVI | 0.69 (***) | 0.65 (***) |
| SIPI | −0.68 (***) | −0.71 (***) |
| BAND_7 | −0.64 (***) | −0.61 (***) |
| BAND_3 | −0.64 (***) | −0.56 (***) |
| NIR/SWIR2 | 0.63 (***) | 0.67 (***) |
| NDWI2 | 0.63 (***) | 0.67 (***) |
| SAVI | 0.63 (***) | 0.63 (***) |
| MSAVI | 0.63 (***) | 0.63 (***) |
| BAND_5 | −0.63 (***) | −0.59 (***) |
| SWIR1/SWIR2 | 0.58 (***) | 0.57 (***) |
| NIR/SWIR1 | 0.54 (***) | 0.58 (***) |
| NDWI1 | 0.54 (***) | 0.58 (***) |
| BAND_2 | −0.50 (***) | −0.45 (***) |
| BAND_1 | −0.37 (***) | −0.40 (***) |
| BAND_4 | −0.36 (***) | −0.24 (***) |
| SPI_6 | 0.35 (***) | 0.41 (***) |
| SPI_9 | 0.21 (**) | 0.29 (***) |
| SPI_12 | 0.20 (**) | 0.27 (***) |
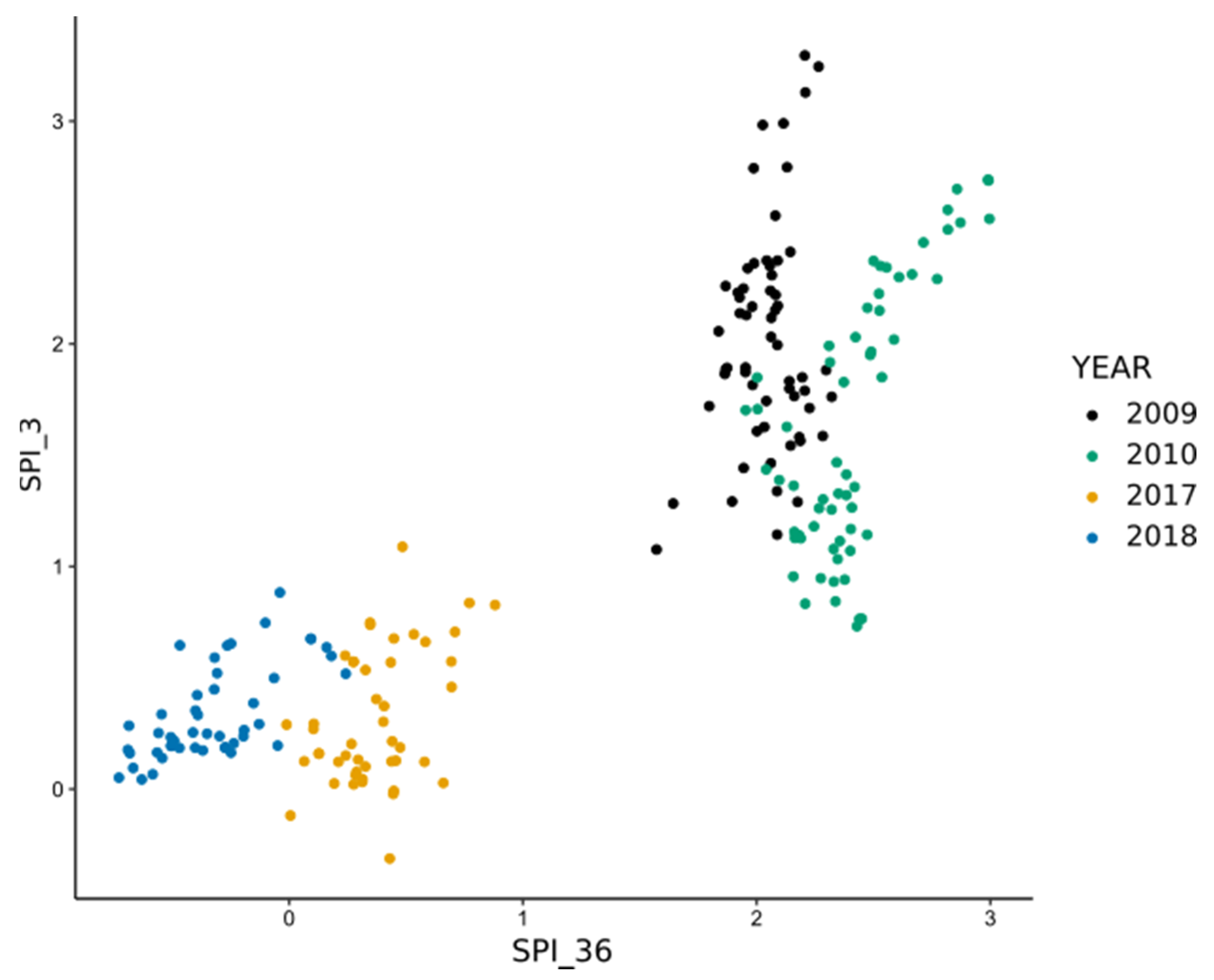
| SPI_3 | 0.11 | 0.21 (**) |
| SPI_24 | 0.09 | 0.05 |
| SPI_2 | −0.09 | −0.01 |
| SPI_18 | 0.05 | −0.04 |
| SPI_36 | 0.04 | −0.06 |
| SPI_48 | 0.00 | −0.05 |
| SPI_1 | 0.00 | 0.02 |
| R2 | RMSE (t/ha) | MAE (t/ha) | Mean Biomass (t/ha) | |
|---|---|---|---|---|
| Training (calibration) | 0.56 | 0.46 | 0.32 | 0.61 |
| Testing (validation) | 0.63 | 0.53 | 0.33 | 0.67 |
| Variable | Conditional Permutation Importance | |
|---|---|---|
| Absolute Value | Scaled (100 to 0) | |
| ARVI | 2.39 × 10−2 | 100 |
| NDVI | 1.20 × 10−2 | 51 |
| EVI | 1.01 × 10−2 | 43 |
| TSAVI | × 10−2 | 43 |
| NIR/R | 9.46 × 10−3 | 41 |
| SIPI | 9.17 × 10−3 | 39 |
| OSAVI | 6.74 × 10−3 | 29 |
| BAND_7 | 3.29 × 10−3 | 15 |
| SPI_2 | 3.06 × 10−3 | 14 |
| BAND_3 | 2.53 × 10−3 | 12 |
| SPI_48 | 2.28 × 10−3 | 11 |
| BAND_5 | 2.05 × 10−3 | 10 |
| NDWI2 | 1.98 × 10−3 | 10 |
| SWIR1/SWIR2 | 1.74 × 10−3 | 9 |
| MSAVI | 1.69 × 10−3 | 9 |
| SPI_3 | 1.68 × 10−3 | 9 |
| NIR/SWIR1 | 1.20 × 10−3 | 7 |
| SPI_18 | 1.18 × 10−3 | 6 |
| SPI_24 | 9.45 × 10−4 | 6 |
| SPI_36 | 9.41 × 10−4 | 5 |
| SAVI | 8.99 × 10−4 | 5 |
| NDWI1 | 8.87 × 10−4 | 5 |
| SPI_1 | 8.45 × 10−4 | 5 |
| NIR/SWIR2 | 8.06 × 10−4 | 5 |
| BAND_2 | 5.69 × 10−4 | 4 |
| SPI_12 | 5.65 × 10−4 | 4 |
| BAND_4 | 4.02 × 10−4 | 3 |
| SPI_9 | 2.51 × 10−4 | 3 |
| SPI_6 | −3.45 × 10−4 | 0 |
| BAND_1 | −3.90 × 10−4 | 0 |
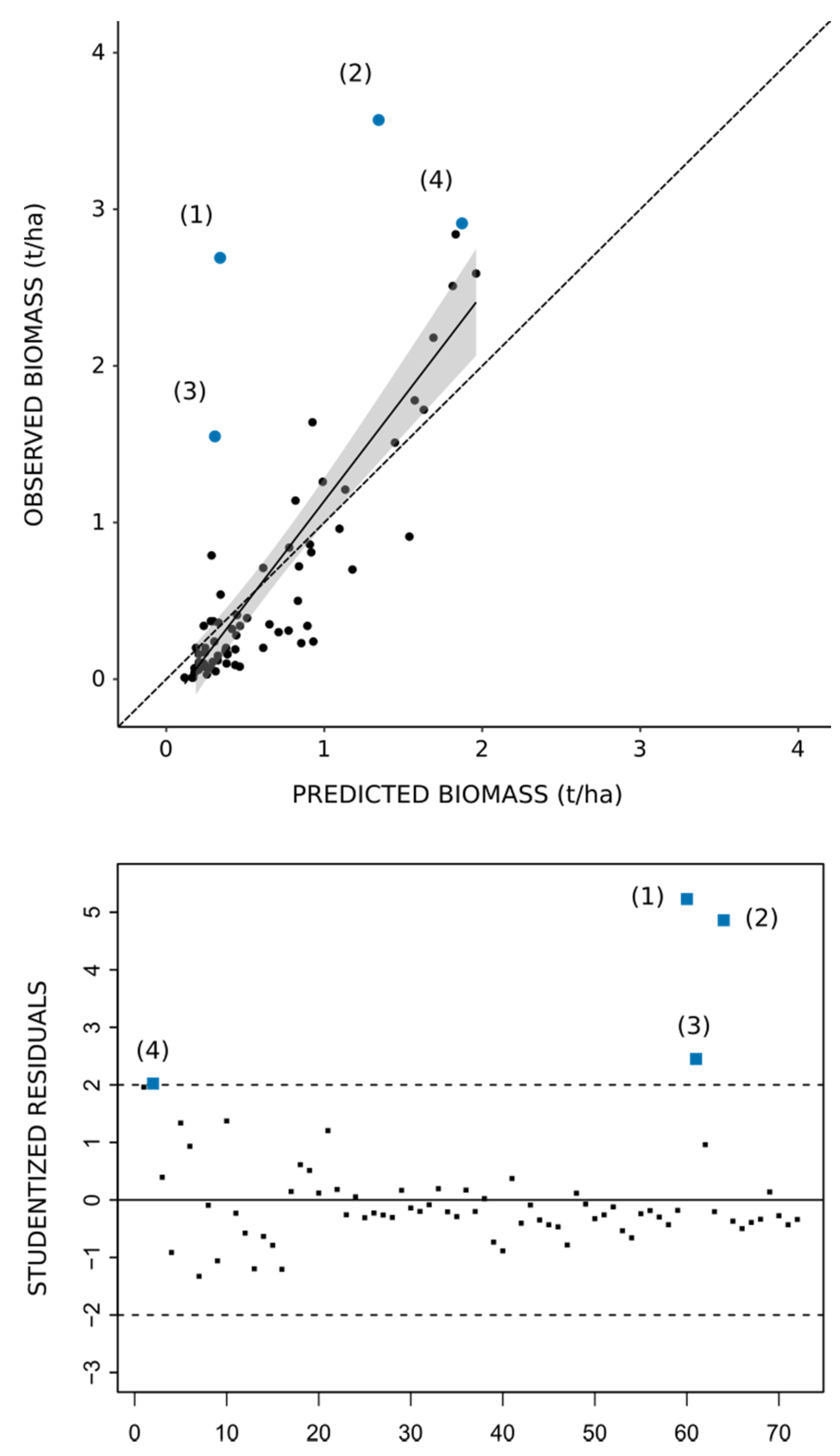
| ID | Year | Vegetation Formation | Observed Biomass (t/ha) | Predicted Biomass (t/ha) | Residuals (t/ha) | Studentized Residuals |
|---|---|---|---|---|---|---|
| 1 | 2018 | Alfa steppes | 2.69 | 0.34 | 2.35 | 5.23 |
| 2 | 2018 | Alfa steppes | 3.57 | 1.34 | 2.23 | 4.86 |
| 3 | 2018 | Mixed steppes | 1.55 | 0.31 | 1.24 | 2.45 |
| 4 | 2009 | Alfa steppes | 2.91 | 1.87 | 1.04 | 2.02 |
| Vegetation Formation | n obs. | R2 | Adj. R2 | RMSE (t/ha) | MAE (t/ha) | Mean Biomass (t/ha) |
|---|---|---|---|---|---|---|
| All | 214 | 0.60 | 0.59 | 0.46 | 0.31 | 0.65 |
| Alfa | 60 | 0.43 | 0.42 | 0.68 | 0.51 | 1.09 |
| Other | 154 | 0.68 | 0.67 | 0.32 | 0.23 | 0.45 |
| Year | Vegetation Formation | Observed Biomass (t/ha) | Predicted Biomass (t/ha) | Residuals (t/ha) | Studentized Residuals |
|---|---|---|---|---|---|
| 2018 | Alfa steppes | 3.57 | 1.25 | 2.32 | 5.34 |
| 2018 | Alfa steppes | 2.69 | 0.54 | 2.15 | 4.91 |
| 2018 | Alfa steppes | 2.71 | 0.84 | 1.87 | 4.21 |
| 2018 | Alfa steppes | 3.19 | 1.49 | 1.70 | 3.80 |
| 2018 | Alfa steppes | 2.54 | 1.02 | 1.52 | 3.38 |
| 2018 | Mixed steppes | 1.59 | 0.19 | 1.40 | 3.09 |
| 2018 | Mixed steppes | 1.55 | 0.42 | 1.13 | 2.47 |
| Year | n obs. | Intercept Estimate (Std. Error) | Slope Estimate (Std. Error) | Model F-Statistic (df) | p-Value | Sign. Level |
|---|---|---|---|---|---|---|
| 2009 | 18 | 1.16 (0.14) | 12.77 (2.56) | 24.83 (16) | 1.354 × 10−4 | *** |
| 2010 | 19 | 0.77 (0.04) | 7.39 (0.56) | 176.7 (17) | 2.075 × 10−10 | *** |
| 2017 | 10 | 1.74 (0.29) | 15.01 (4.2) | 12.8 (8) | 7.217 × 10−3 | ** |
| 2018 | 13 | 2.72 (0.31) | 19.86 (3.99) | 24.72 (11) | 4.210 × 10−4 | *** |
| Year | R2 | Adj. R2 | RMSE (t/ha) | MAE (t/ha) | Mean Biomass (t/ha) |
|---|---|---|---|---|---|
| 2009 | 0.61 | 0.58 | 0.47 | 0.41 | 1.53 |
| 2010 | 0.91 | 0.91 | 0.14 | 0.12 | 0.51 |
| 2017 | 0.62 | 0.57 | 0.34 | 0.27 | 0.79 |
| 2018 | 0.69 | 0.66 | 0.63 | 0.49 | 1.53 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lang, M.; Mahyou, H.; Tychon, B. Estimation of Rangeland Production in the Arid Oriental Region (Morocco) Combining Remote Sensing Vegetation and Rainfall Indices: Challenges and Lessons Learned. Remote Sens. 2021, 13, 2093. https://doi.org/10.3390/rs13112093
Lang M, Mahyou H, Tychon B. Estimation of Rangeland Production in the Arid Oriental Region (Morocco) Combining Remote Sensing Vegetation and Rainfall Indices: Challenges and Lessons Learned. Remote Sensing. 2021; 13(11):2093. https://doi.org/10.3390/rs13112093
Chicago/Turabian StyleLang, Marie, Hamid Mahyou, and Bernard Tychon. 2021. "Estimation of Rangeland Production in the Arid Oriental Region (Morocco) Combining Remote Sensing Vegetation and Rainfall Indices: Challenges and Lessons Learned" Remote Sensing 13, no. 11: 2093. https://doi.org/10.3390/rs13112093
APA StyleLang, M., Mahyou, H., & Tychon, B. (2021). Estimation of Rangeland Production in the Arid Oriental Region (Morocco) Combining Remote Sensing Vegetation and Rainfall Indices: Challenges and Lessons Learned. Remote Sensing, 13(11), 2093. https://doi.org/10.3390/rs13112093