A Review of Remote Sensing Challenges for Food Security with Respect to Salinity and Drought Threats


Abstract
1. Introduction
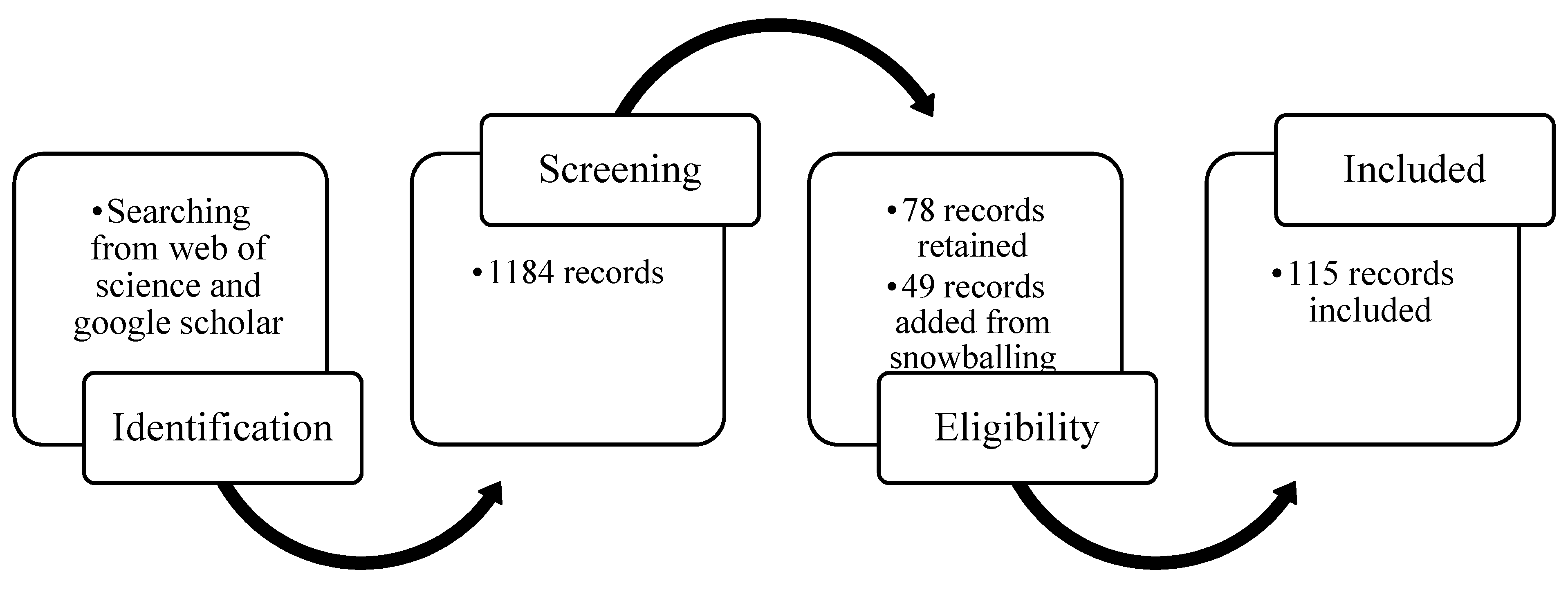
2. Methodology
2.1. Creating Representative Database though a Systematic Review
2.2. Extraction of Drought/Salinity Stress Information
2.3. Classification of Plant Traits and Vegetation Indices
2.4. Analyses of Vegetation Responses
3. Results
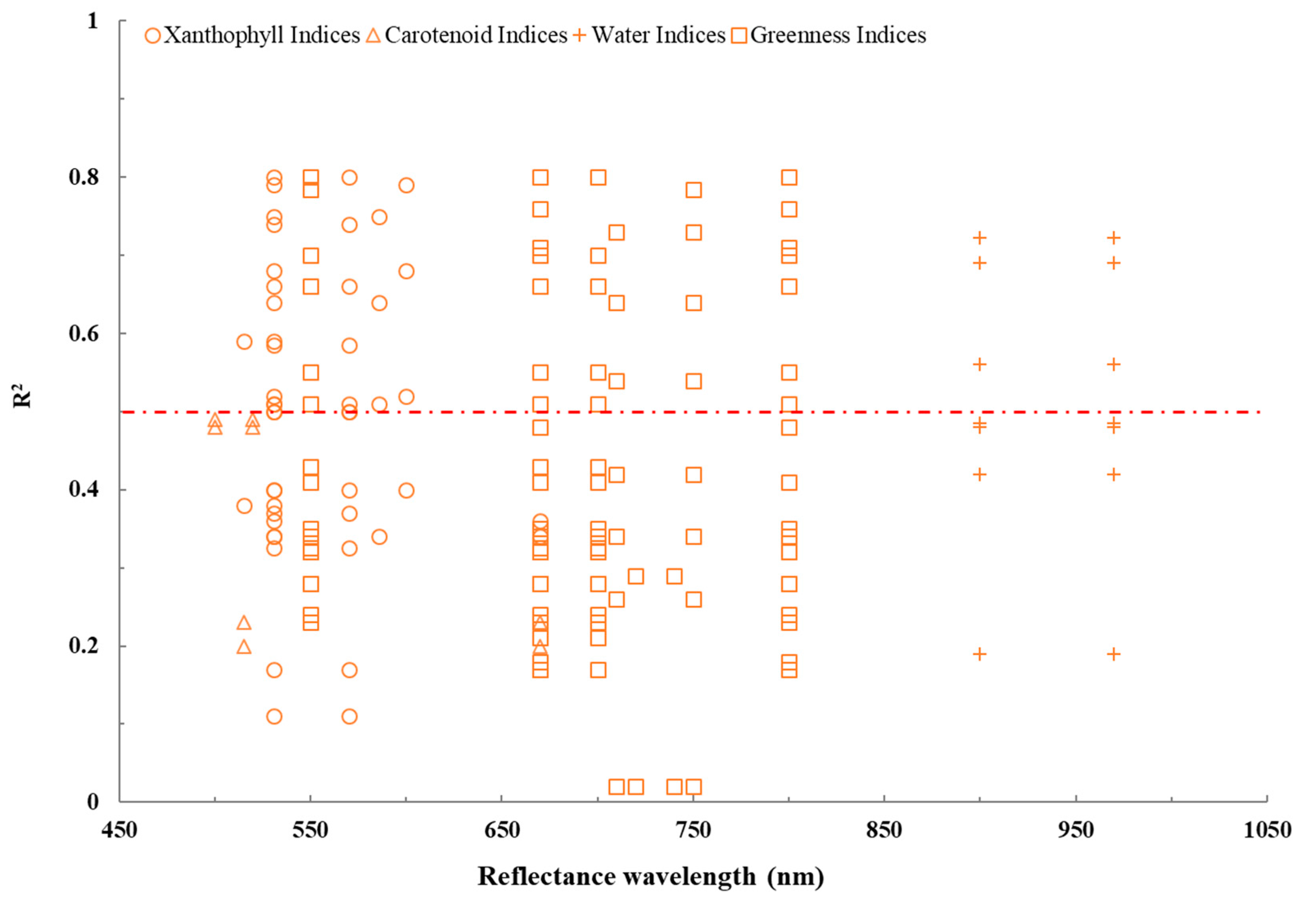
3.1. Spectral Signatures of VIs under Drought Stress
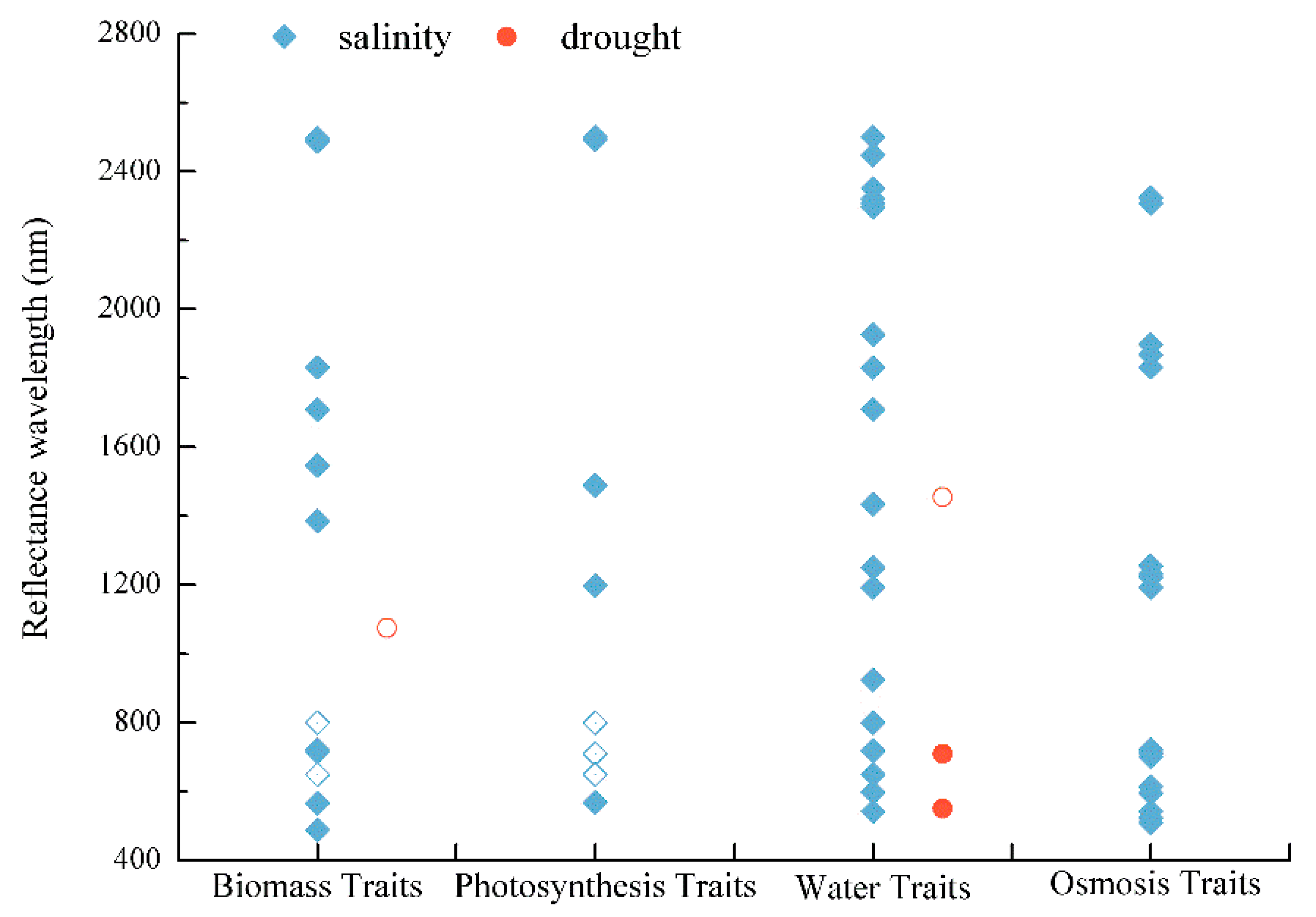
3.2. Spectral Signatures of Plant Traits under Drought and Salinity Stress
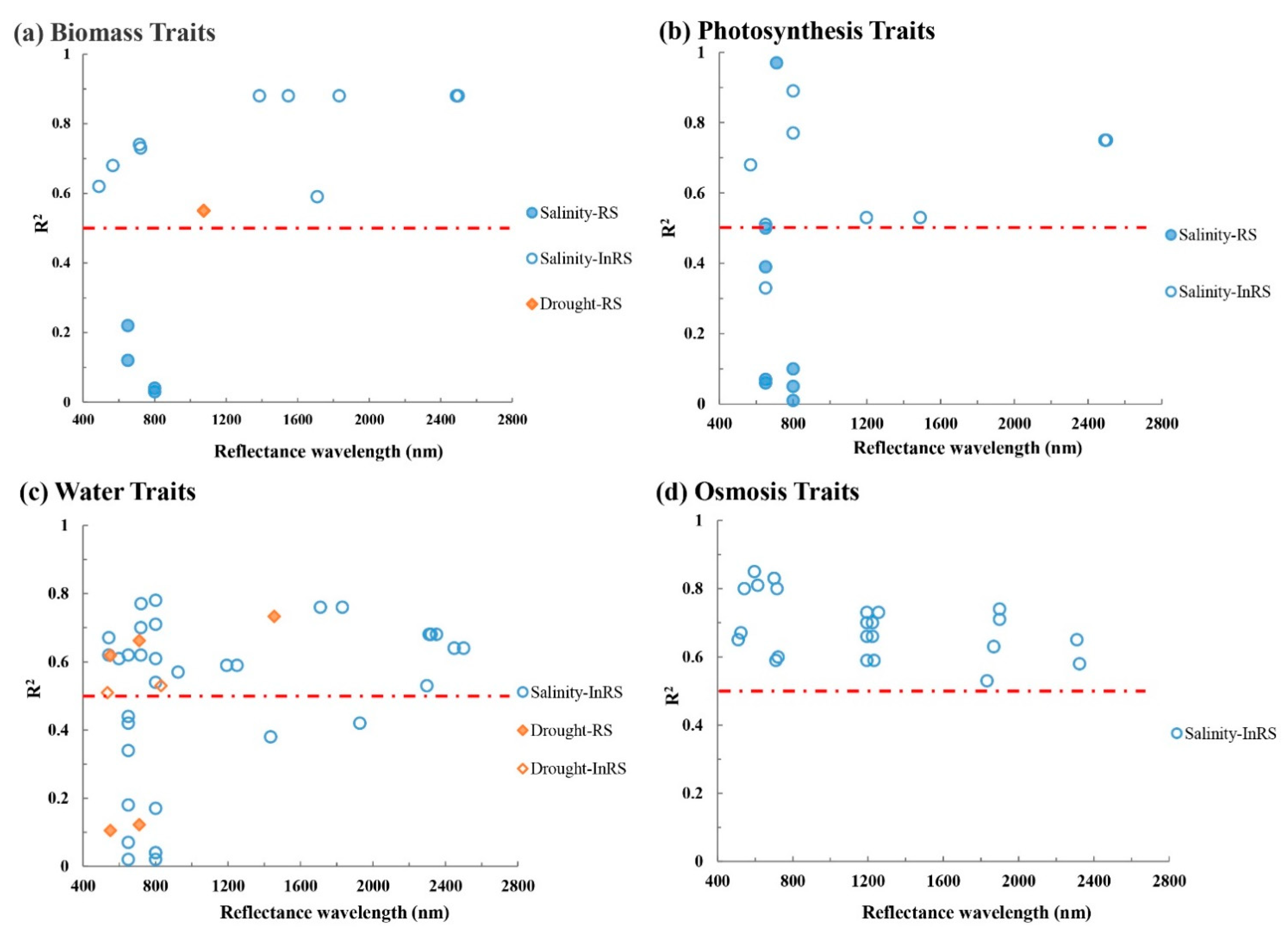
3.3. The Relationship between VIs and Plant Traits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- FAO. The State of the World’s Land and Water Resources for Food and Agriculture-Managing Systems at Risk; Earthscan: Abingdon, UK, 2011. [Google Scholar]
- FAO; UNICEF; WFP; WHO. The State of Food Security and Nutrition in the World 2020; FAO: Rome, Italy, 2020. [Google Scholar]
- Anami, B.S.; Malvade, N.N.; Palaiah, S. Classification of yield affecting biotic and abiotic paddy crop stresses using field images. Inf. Process. Agric. 2020, 7, 272–285. [Google Scholar] [CrossRef]
- Dresselhaus, T.; Hückelhoven, R. Biotic and Abiotic Stress Responses in Crop Plants. Agronomy 2018, 8, 267. [Google Scholar] [CrossRef]
- Dunn, R.J.H., Stanitski, D.M., Gobron, N., Willett, K.M., Eds.; Global climate state of the climate in 2019. Bull. Amer. Meteor. 2020, 101, S9–S127. [Google Scholar]
- Smith, A.B. U.S. Billion-Dollar Weather and Climate Disasters, 1980–Present; NCEI Accession 0209268; NOAA National Centers for Environmental Information: Asheville, NC, USA, 2020. [CrossRef]
- Su, B.; Huang, J.; Fischer, T.; Wang, Y.; Kundzewicz, Z.W.; Zhai, J.; Sun, H.; Wang, A.; Zeng, X.; Wang, G.; et al. Drought losses in China might double between the 1.5 °C and 2.0 °C warming. Proc. Natl. Acad. Sci. USA 2018, 115, 10600–10605. [Google Scholar] [CrossRef]
- FAO; IIASA; ISRIC; ISSCAS; JRC. Harmonized World Soil Database (Version 1.2); FAO: Rome, Italy; Laxenburg, Austria, 2012. [Google Scholar]
- Koohafkan, P. Water and Cereals in Drylands; Routledge: London, UK, 2012. [Google Scholar]
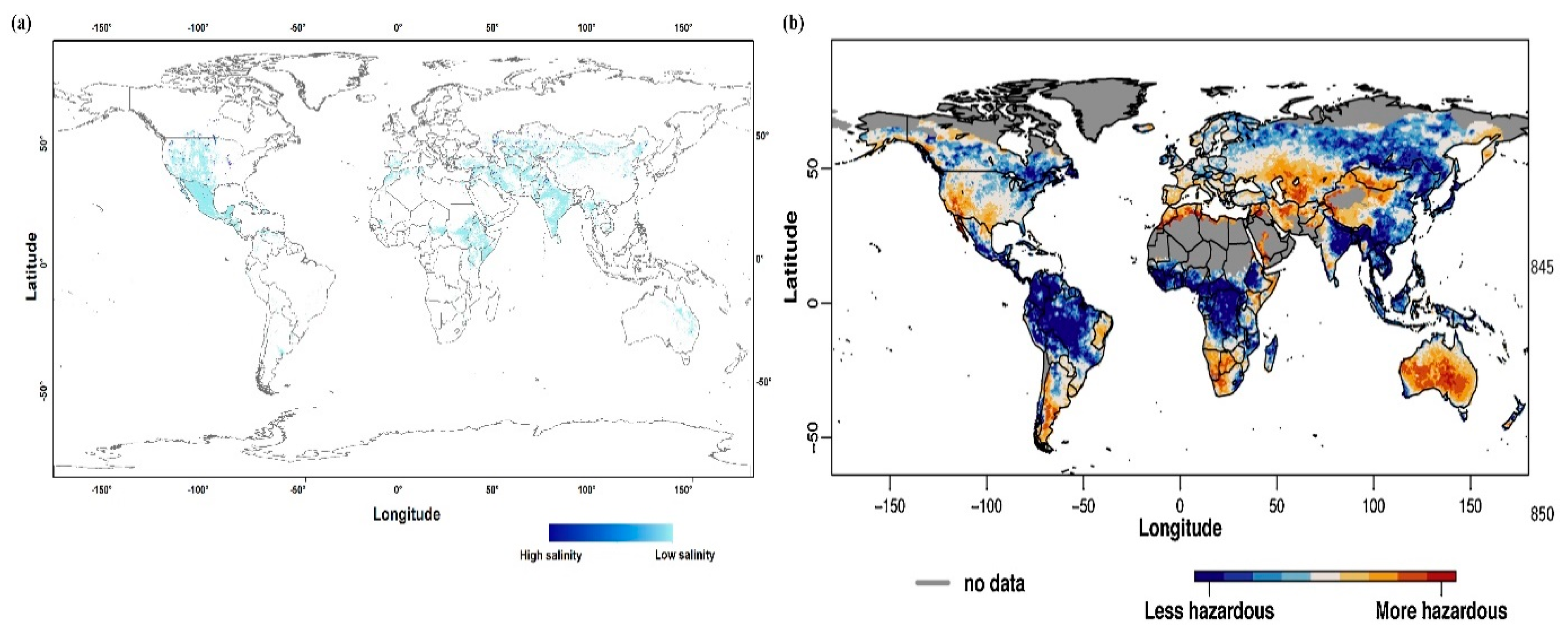
- Ivushkin, K.; Bartholomeus, H.; Bregt, A.K.; Pulatov, A.; Kempen, B.; de Sousa, L. Global mapping of soil salinity change. Remote Sens. Environ. 2019, 231, 111260. [Google Scholar] [CrossRef]
- Cook, B.I.; Ault, T.R.; Smerdon, J.E. Unprecedented 21st century drought risk in the American Southwest and Central Plains. Sci. Adv. 2015, 1, e1400082. [Google Scholar] [CrossRef]
- Trenberth, K.E.; Dai, A.; van der Schrier, G.; Jones, P.D.; Barichivich, J.; Briffa, K.R.; Sheffield, J. Global warming and changes in drought. Nat. Clim. Chang. 2014, 4, 17–22. [Google Scholar] [CrossRef]
- Schwalm, C.R.; Anderegg, W.R.L.; Michalak, A.M.; Fisher, J.B.; Biondi, F.; Koch, G.; Litvak, M.; Ogle, K.; Shaw, J.D.; Wolf, A.; et al. Global patterns of drought recovery. Nature 2017, 548, 202–205. [Google Scholar] [CrossRef]
- Mosley, L.M. Drought impacts on the water quality of freshwater systems: Review and integration. Earth Sci. Rev. 2015, 140, 203–214. [Google Scholar] [CrossRef]
- Mimi, Z.A.; Jamous, S.A. Climate change and agricultural water demand: Impacts and adaptations. Afr. J. Environ. Sci. Technol. 2010, 4, 183–191. [Google Scholar] [CrossRef]
- Butcher, K.; Wick, A.F.; DeSutter, T.; Chatterjee, A.; Harmon, J. Soil Salinity: A Threat to Global Food Security. Agron. J. 2016, 108, 2189–2200. [Google Scholar] [CrossRef]
- Rozema, J.; Flowers, T. Ecology: Crops for a salinized world. Science 2008, 322, 1478–1480. [Google Scholar] [CrossRef] [PubMed]
- Cramer, G.R.; Ergul, A.; Grimplet, J.; Tillett, R.L.; Tattersall, E.A.; Bohlman, M.C.; Vincent, D.; Sonderegger, J.; Evans, J.; Osborne, C.; et al. Water and salinity stress in grapevines: Early and late changes in transcript and metabolite profiles. Funct. Integr. Genom. 2007, 7, 111–134. [Google Scholar] [CrossRef] [PubMed]
- Oki, T.; Kanae, S. Global hydrological cycles and world water resources. Science 2006, 313, 1068–1072. [Google Scholar] [CrossRef] [PubMed]
- Hrdinka, T.; Novický, O.; Hanslík, E.; Rieder, M. Possible impacts of floods and droughts on water quality. J. Hydro-Environ. Res. 2012, 6, 145–150. [Google Scholar] [CrossRef]
- van Vliet, M.T.H.; Zwolsman, J.J.G. Impact of summer droughts on the water quality of the Meuse river. J. Hydrol. 2008, 353, 1–17. [Google Scholar] [CrossRef]
- Jones, E.; van Vliet, M.T.H. Drought impacts on river salinity in the southern US: Implications for water scarcity. Sci. Total Environ. 2018, 644, 844–853. [Google Scholar] [CrossRef]
- Wang, J.; Huang, X.; Zhong, T.; Chen, Z. Climate change impacts and adaptation for saline agriculture in north Jiangsu Province, China. Environ. Sci. Policy 2013, 25, 83–93. [Google Scholar] [CrossRef]
- Katschnig, D.; Broekman, R.; Rozema, J. Salt tolerance in the halophyte Salicornia dolichostachya Moss: Growth, morphology and physiology. Environ. Exp. Bot. 2013, 92, 32–42. [Google Scholar] [CrossRef]
- Pankova, Y.I.; Konyushkova, M.V. Effect of global warming on soil salinity of the arid regions. Russ. Agric. Sci. 2014, 39, 464–467. [Google Scholar] [CrossRef]
- Corwin, D.L. Climate change impacts on soil salinity in agricultural areas. Eur. J. Soil Sci. 2020. [Google Scholar] [CrossRef]
- Gopalakrishnan, T.; Hasan, M.; Haque, A.; Jayasinghe, S.; Kumar, L. Sustainability of Coastal Agriculture under Climate Change. Sustainability 2019, 11, 7200. [Google Scholar] [CrossRef]
- Carrão, H.; Naumann, G.; Barbosa, P. Mapping global patterns of drought risk: An empirical framework based on sub-national estimates of hazard, exposure and vulnerability. Glob. Environ. Chang. 2016, 39, 108–124. [Google Scholar] [CrossRef]
- Wu, B.; Gommes, R.; Zhang, M.; Zeng, H.; Yan, N.; Zou, W.; Zheng, Y.; Zhang, N.; Chang, S.; Xing, Q.; et al. Global Crop Monitoring: A Satellite-Based Hierarchical Approach. Remote Sens. 2015, 7, 3907–3933. [Google Scholar] [CrossRef]
- Tucker, C.J. Red and Photographic Infrared Linear Combinations for Monitoring Vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef]
- Rondeaux, G.; Steven, M.; Baret, F. Optimization of soil-adjusted vegetation indices. Remote Sens. Environ. 1996, 55, 95–107. [Google Scholar] [CrossRef]
- Huete, A.R. A Soil-Adjusted Vegetation Index (Savi). Remote Sens. Environ. 1988, 25, 295–309. [Google Scholar] [CrossRef]
- Kogan, F.N. Droughts of the Late 1980′s in the United States as Derived from Noaa Polar-Orbiting Satellite Data. Bull. Am. Meteorol. Soc. 1995, 76, 655–668. [Google Scholar] [CrossRef]
- Kogan, F.N. Global drought watch from space. Bull. Am. Meteorol. Soc. 1997, 78, 621–636. [Google Scholar] [CrossRef]
- Gao, B.C. NDWI—A normalized difference water index for remote sensing of vegetation liquid water from space. Remote Sens. Environ. 1996, 58, 257–266. [Google Scholar] [CrossRef]
- Liu, X.; Zhu, X.; Pan, Y.; Li, S.; Liu, Y.; Ma, Y. Agricultural drought monitoring: Progress, challenges, and prospects. J. Geogr. Sci. 2016, 26, 750–767. [Google Scholar] [CrossRef]
- Allbed, A.; Kumar, L. Soil Salinity Mapping and Monitoring in Arid and Semi-Arid Regions Using Remote Sensing Technology: A Review. Adv. Remote Sens. 2013, 2, 373–385. [Google Scholar] [CrossRef]
- Metternicht, G.I.; Zinck, J.A. Remote sensing of soil salinity: Potentials and constraints. Remote Sens. Environ. 2003, 85, 1–20. [Google Scholar] [CrossRef]
- Serbin, S.P.; Singh, A.; Desai, A.R.; Dubois, S.G.; Jablonski, A.D.; Kingdon, C.C.; Kruger, E.L.; Townsend, P.A. Remotely estimating photosynthetic capacity, and its response to temperature, in vegetation canopies using imaging spectroscopy. Remote Sens. Environ. 2015, 167, 78–87. [Google Scholar] [CrossRef]
- Serbin, S.P.; Singh, A.; McNeil, B.E.; Kingdon, C.C.; Townsend, P.A. Spectroscopic determination of leaf morphological and biochemical traits for northern temperate and boreal tree species. Ecol. Appl. 2014, 24, 1651–1669. [Google Scholar] [CrossRef]
- Gerhards, M.; Schlerf, M.; Mallick, K.; Udelhoven, T. Challenges and Future Perspectives of Multi-/Hyperspectral Thermal Infrared Remote Sensing for Crop Water-Stress Detection: A Review. Remote Sens. 2019, 11, 1240. [Google Scholar] [CrossRef]
- Vereecken, H.; Weihermüller, L.; Jonard, F.; Montzka, C. Characterization of Crop Canopies and Water Stress Related Phenomena using Microwave Remote Sensing Methods: A Review. Vadose Zone J. 2012, 11, vzj2011.0138ra. [Google Scholar] [CrossRef]
- Homolová, L.; Malenovský, Z.; Clevers, J.G.P.W.; García-Santos, G.; Schaepman, M.E. Review of optical-based remote sensing for plant trait mapping. Ecol. Complex. 2013, 15, 1–16. [Google Scholar] [CrossRef]
- Weiss, M.; Jacob, F.; Duveiller, G. Remote sensing for agricultural applications: A meta-review. Remote Sens. Environ. 2020, 236, 111402. [Google Scholar] [CrossRef]
- Berger, K.; Atzberger, C.; Danner, M.; D’Urso, G.; Mauser, W.; Vuolo, F.; Hank, T. Evaluation of the PROSAIL Model Capabilities for Future Hyperspectral Model Environments: A Review Study. Remote Sens. 2018, 10, 85. [Google Scholar] [CrossRef]
- Pérez-Harguindeguy, N.; Díaz, S.; Garnier, E.; Lavorel, S.; Poorter, H.; Jaureguiberry, P.; Bret-Harte, M.S.; Cornwell, W.K.; Craine, J.M.; Gurvich, D.E.; et al. New handbook for standardised measurement of plant functional traits worldwide. Aust. J. Bot. 2013, 61, 167–234. [Google Scholar] [CrossRef]
- Niinemets, U. Is there a species spectrum within the world-wide leaf economics spectrum? Major variations in leaf functional traits in the Mediterranean sclerophyll Quercus ilex. N. Phytol. 2015, 205, 79–96. [Google Scholar] [CrossRef] [PubMed]
- Gamon, J.A.; Penuelas, J.; Field, C.B. A Narrow-Waveband Spectral Index That Tracks Diurnal Changes in Photosynthetic Efficiency. Remote Sens. Environ. 1992, 41, 35–44. [Google Scholar] [CrossRef]
- Hernández-Clemente, R.; Navarro-Cerrillo, R.M.; Suárez, L.; Morales, F.; Zarco-Tejada, P.J. Assessing structural effects on PRI for stress detection in conifer forests. Remote Sens. Environ. 2011, 115, 2360–2375. [Google Scholar] [CrossRef]
- Panigada, C.; Rossini, M.; Meroni, M.; Cilia, C.; Busetto, L.; Amaducci, S.; Boschetti, M.; Cogliati, S.; Picchi, V.; Pinto, F.; et al. Fluorescence, PRI and canopy temperature for water stress detection in cereal crops. Int. J. Appl. Earth Obs. Geoinf. 2014, 30, 167–178. [Google Scholar] [CrossRef]
- Peñuelas, J.; Filella, I.; Biel, C.; Serrano, L.; Savé, R. The reflectance at the 950–970 nm region as an indicator of plant water status. Int. J. Remote Sens. 1993, 14, 1887–1905. [Google Scholar] [CrossRef]
- Idso, S.B.; Jackson, R.D.; Pinter, P.J.; Reginato, R.J.; Hatfield, J.L. Normalizing the Stress-Degree-Day Parameter for Environmental Variability. Agric. Meteorol. 1981, 24, 45–55. [Google Scholar] [CrossRef]
- Zarco-Tejada, P.J.; González-Dugo, V.; Berni, J.A.J. Fluorescence, temperature and narrow-band indices acquired from a UAV platform for water stress detection using a micro-hyperspectral imager and a thermal camera. Remote Sens. Environ. 2012, 117, 322–337. [Google Scholar] [CrossRef]
- Haboudane, D.; Miller, J.R.; Tremblay, N.; Zarco-Tejada, P.J.; Dextraze, L. Integrated narrow-band vegetation indices for prediction of crop chlorophyll content for application to precision agriculture. Remote Sens. Environ. 2002, 81, 416–426. [Google Scholar] [CrossRef]
- Gitelson, A.A. Remote estimation of canopy chlorophyll content in crops. Geophys. Res. Lett. 2005, 32, L08403. [Google Scholar] [CrossRef]
- Asrar, G.; Kanemasu, E.T.; Jackson, R.D.; Pinter, P.J. Estimation of Total above-Ground Phytomass Production Using Remotely Sensed Data. Remote Sens. Environ. 1985, 17, 211–220. [Google Scholar] [CrossRef]
- Zarco-Tejada, P.J.; González-Dugo, V.; Williams, L.E.; Suárez, L.; Berni, J.A.J.; Goldhamer, D.; Fereres, E. A PRI-based water stress index combining structural and chlorophyll effects: Assessment using diurnal narrow-band airborne imagery and the CWSI thermal index. Remote Sens. Environ. 2013, 138, 38–50. [Google Scholar] [CrossRef]
- Vogelmann, J.E.; Rock, B.N.; Moss, D.M. Red edge spectral measurements from sugar maple leaves. Int. J. Remote Sens. 1993, 14, 1563–1575. [Google Scholar] [CrossRef]
- Zarco-Tejada, P.J.; Miller, J.R.; Noland, T.L.; Mohammed, G.H.; Sampson, P.H. Scaling-up and model inversion methods with narrowband optical indices for chlorophyll content estimation in closed forest canopies with hyperspectral data. IEEE Trans. Geosci. Remote Sens. 2001, 39, 1491–1507. [Google Scholar] [CrossRef]
- Rahimzadeh-Bajgiran, P.; Darvishsefat, A.A.; Khalili, A.; Makhdoum, M.F. Using AVHRR-based vegetation indices for drought monitoring in the Northwest of Iran. J. Arid Environ. 2008, 72, 1086–1096. [Google Scholar] [CrossRef]
- Rahimzadeh-Bajgiran, P.; Omasa, K.; Shimizu, Y. Comparative evaluation of the Vegetation Dryness Index (VDI), the Temperature Vegetation Dryness Index (TVDI) and the improved TVDI (iTVDI) for water stress detection in semi-arid regions of Iran. ISPRS J. Photogramm. Remote Sens. 2012, 68, 1–12. [Google Scholar] [CrossRef]
- Zargar, A.; Sadiq, R.; Naser, B.; Khan, F.I. A review of drought indices. Environ. Rev. 2011, 19, 333–349. [Google Scholar] [CrossRef]
- Moreno-Martínez, Á.; Camps-Valls, G.; Kattge, J.; Robinson, N.; Reichstein, M.; van Bodegom, P.; Kramer, K.; Cornelissen, J.H.C.; Reich, P.; Bahn, M.; et al. A methodology to derive global maps of leaf traits using remote sensing and climate data. Remote Sens. Environ. 2018, 218, 69–88. [Google Scholar] [CrossRef]
- Mahajan, S.; Tuteja, N. Cold, salinity and drought stresses: An overview. Arch. Biochem. Biophys. 2005, 444, 139–158. [Google Scholar] [CrossRef]
- Yeo, A. Molecular biology of salt tolerance in the context of whole-plant physiology. J. Exp. Bot. 1998, 49, 915–929. [Google Scholar] [CrossRef]
- Niu, X.; Bressan, R.A.; Hasegawa, P.M.; Pardo, J.M. Ion Homeostasis in NaCl Stress Environments. Plant Physiol. 1995, 109, 735–742. [Google Scholar] [CrossRef] [PubMed]
- Bartels, D.; Sunkar, R. Drought and Salt Tolerance in Plants. Crit. Rev. Plant Sci. 2005, 24, 23–58. [Google Scholar] [CrossRef]
- El-Hendawy, S.E.; Al-Suhaibani, N.A.; Hassan, W.M.; Dewir, Y.H.; Elsayed, S.; Al-Ashkar, I.; Abdella, K.A.; Schmidhalter, U. Evaluation of wavelengths and spectral reflectance indices for high-throughput assessment of growth, water relations and ion contents of wheat irrigated with saline water. Agric. Water Manag. 2019, 212, 358–377. [Google Scholar] [CrossRef]
- Zhang, L.; Zhou, Z.; Zhang, G.; Meng, Y.; Chen, B.; Wang, Y. Monitoring cotton (Gossypium hirsutum L.) leaf ion content and leaf water content in saline soil with hyperspectral reflectance. Eur. J. Remote Sens. 2014, 47, 593–610. [Google Scholar] [CrossRef]
- Bayat, B.; van der Tol, C.; Verhoef, W. Remote Sensing of Grass Response to Drought Stress Using Spectroscopic Techniques and Canopy Reflectance Model Inversion. Remote Sens. 2016, 8, 557. [Google Scholar] [CrossRef]
- Botha, E.J.; Zebarth, B.J.; Leblon, B. Non-destructive estimation of potato leaf chlorophyll and protein contents from hyperspectral measurements using the PROSPECT radiative transfer model. Can. J. Plant Sci. 2006, 86, 279–291. [Google Scholar] [CrossRef]
- Jacquemoud, S.; Bacour, C.; Poilve, H.; Frangi, J.P. Comparison of Four Radiative Transfer Modelsto Simulate Plant Canopies Reflectance: Direct and Inverse Mode. Remote Sens. Environ. 2000, 74, 471–481. [Google Scholar] [CrossRef]
- Lu, B.; Dao, P.D.; Liu, J.; He, Y.; Shang, J. Recent Advances of Hyperspectral Imaging Technology and Applications in Agriculture. Remote Sens. 2020, 12, 2659. [Google Scholar] [CrossRef]
- Timmermans, J.; Verhoef, W.; van der Tol, C.; Su, Z. Retrieval of canopy component temperatures through Bayesian inversion of directional thermal measurements. Hydrol. Earth Syst. Sci. 2009, 13, 1249–1260. [Google Scholar] [CrossRef]
- Berni, J.; Zarco-Tejada, P.J.; Suarez, L.; Fereres, E. Thermal and Narrowband Multispectral Remote Sensing for Vegetation Monitoring from an Unmanned Aerial Vehicle. IEEE Trans. Geosci. Remote Sens. 2009, 47, 722–738. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | RS Methods | Traits | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Biomass traits | RS | LMA | LAI | -- | -- | -- | -- | -- | -- |
| In-RS | FS | SDW | BDW | BFW | -- | -- | -- | -- | |
| Photosynthesis traits | RS | Chl | Chla/Chlb | -- | -- | -- | -- | -- | -- |
| In-RS | A | Pn | ΔF/Fm | Chl × ΔF/Fm | -- | -- | -- | -- | |
| Water traits | RS | LCT | CWC | RWC | EWT | CWM | -- | ||
| In-RS | Gs | LOP | Ψp | LWP | Ψs | E | Tl − Tair | -- | |
| Osmosis traits | RS | -- | -- | -- | -- | -- | -- | -- | -- |
| In-RS | Na+ | Cl− | K+ | Ca2+ | K+/Na+ | TSS | TA | TSS/TA | |
| VIs | Meaning | Equation | Reference |
|---|---|---|---|
| Xanthophyll Indices | |||
| PRI570 | Photochemical reflectance index | (R531 − R570)/(R531 + R570) | [48] |
| PRI515 | Photochemical reflectance index | (R531 − R515)/(R531 + R515) | [49] |
| PRI586 | Photochemical reflectance index | (R531 − R586)/(R531 + R586) | [50] |
| PRI600 | Photochemical reflectance index | (R531 − R602)/(R531 + R602) | [49] |
| PRI670 | Photochemical reflectance index | (R531 − R668)/(R531 + R668) | [49] |
| Water Content Indices | |||
| WI | Water index | R900/R970 | [51] |
| CWSI | Crop Water Stress Index | CWSI = (Tleaf − Twet)/(Tdry − Twet) | [52] |
| Carotenoid Indices | |||
| R520/R500 | Carotenoid concentration | [53] | |
| R515/R570 | Carotenoid concentration | [53] | |
| Greenness Indices | |||
| OSAVI | Optimized Soil-Adjusted Vegetation Index | (R800 − R670)/(R800 + R670 + 0.16) | [31] |
| TCARI | The Transformed Chlorophyll Absorption in Reflectance Index | TCARI = 3 [(R700 − R670) − 0.2 (R700 − R550) (R700/R670)] | [54] |
| TCARI/OSAVI | Normalized by OSAVI to obtain | TCARI/OSAVI = [3 [(R700 − R670) − 0.2 (R700 − R550) (R700/R670)]]/[(1 + 0.16) (R800 − R670)/(R800 + R670 + 0.16)] | [54] |
| CIgreen | Green chlorophyll index | (R750/R550) − 1 | [55] |
| CIred edge | Red edge chlorophyll index | (R750/R710) − 1 | [55] |
| SR | Simple ratio | R800/R670 | [56] |
| Red edge ratio index | R700/R670 | [57] | |
| VOG1 | The chlorophyll a + b index | R740/R720 | [58] |
| ZM | The chlorophyll a + b index | R750/R710 | [59] |
| Vis | Biomass Traits | Photosynthesis Traits | Water Traits | Osmosis Traits | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| LAI * | FS | Chl * | ΔF/Fm | Chl × ΔF/Fm | Tl − Tair | CWC * | RWC * | EWT * | EWTcanopy * | LWP | Gs | TSS | TA | TSS/TA | |
| Xanthophyll Indices | |||||||||||||||
| PRI570 | 0.66 | 0.11 | -- | -- | 0.40 | 0.74 | -- | 0.51 | -- | -- | 0.37 | 0.59 | 0.17 | 0.50 | 0.50 |
| PRI515 | -- | -- | -- | -- | -- | -- | -- | -- | -- | -- | 0.38 | 0.59 | -- | -- | -- |
| PRI586 | 0.64 | -- | -- | 0.51 | 0.34 | -- | -- | -- | -- | 0.75 | -- | -- | -- | -- | -- |
| PRI600 | 0.40 | -- | -- | 0.68 | -- | 0.79 | -- | 0.52 | -- | -- | -- | -- | -- | -- | -- |
| PRI670 | -- | -- | -- | 0.34 | 0.36 | -- | -- | -- | -- | -- | -- | -- | -- | -- | -- |
| Carotenoid Indices | |||||||||||||||
| R520/R500 | -- | -- | -- | -- | -- | -- | -- | -- | -- | -- | 0.48 | 0.49 | -- | -- | -- |
| R515/R670 | -- | -- | -- | -- | -- | -- | -- | -- | -- | -- | 0.20 | 0.23 | -- | -- | -- |
| Water Content Indices | |||||||||||||||
| WI | 0.49 | -- | -- | 0.48 | 0.19 | 0.69 | 0.72 | 0.42 | -- | 0.56 | -- | -- | -- | -- | -- |
| CWSI | -- | -- | -- | -- | -- | -- | -- | -- | -- | -- | 0.78 | 0.77 | -- | -- | -- |
| Greenness Indices | |||||||||||||||
| OSAVI | 0.71 | -- | -- | 0.48 | 0.32 | -- | -- | -- | -- | 0.76 | -- | -- | -- | -- | -- |
| TCARI | -- | -- | 0.43 | -- | -- | -- | -- | -- | -- | -- | 0.325 | 0.32 | -- | -- | -- |
| TCARI/OSAVI | 0.34 | 0.32 | 0.66 | 0.70 | 0.51 | 0.80 | -- | 0.41 | 0.55 | -- | 0.28 | 0.23 | 0.24 | 0.35 | 0.28 |
| CIgreen | -- | -- | -- | -- | -- | -- | 0.78 | -- | -- | -- | -- | -- | -- | -- | -- |
| CIred edge | 0.64 | -- | -- | 0.42 | -- | 0.54 | 0.73 | 0.34 | -- | -- | -- | -- | -- | -- | -- |
| SR | -- | 0.18 | -- | -- | -- | -- | -- | -- | -- | -- | -- | -- | 0.28 | 0.34 | 0.17 |
| Red edge ratio index | -- | -- | -- | -- | -- | -- | -- | -- | -- | -- | -- | 0.21 | -- | -- | -- |
| VOG1 | -- | -- | -- | -- | -- | -- | -- | -- | -- | -- | 0.02 | 0.29 | -- | -- | -- |
| ZM | -- | -- | -- | -- | -- | -- | -- | -- | -- | -- | 0.02 | 0.26 | -- | -- | -- |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, W.; Timmermans, J.; Chen, Q.; van Bodegom, P.M. A Review of Remote Sensing Challenges for Food Security with Respect to Salinity and Drought Threats. Remote Sens. 2021, 13, 6. https://doi.org/10.3390/rs13010006
Wen W, Timmermans J, Chen Q, van Bodegom PM. A Review of Remote Sensing Challenges for Food Security with Respect to Salinity and Drought Threats. Remote Sensing. 2021; 13(1):6. https://doi.org/10.3390/rs13010006
Chicago/Turabian StyleWen, Wen, Joris Timmermans, Qi Chen, and Peter M. van Bodegom. 2021. "A Review of Remote Sensing Challenges for Food Security with Respect to Salinity and Drought Threats" Remote Sensing 13, no. 1: 6. https://doi.org/10.3390/rs13010006
APA StyleWen, W., Timmermans, J., Chen, Q., & van Bodegom, P. M. (2021). A Review of Remote Sensing Challenges for Food Security with Respect to Salinity and Drought Threats. Remote Sensing, 13(1), 6. https://doi.org/10.3390/rs13010006