Abstract
Urban green spaces provide a host of ecosystem services, the quantity and structure of which play an important role in human well-being. Rapid urbanization may modify urban green spaces, having various effects on plant diversity. Tropical coastal cities have urbanized rapidly in recent decades, but few studies have been conducted with a focus on their green spaces. We studied the responses of cultivated and spontaneous plants, both key components of urban flora, to the landscape structure of urban green spaces and possible social drivers. We analyzed existing relationships between plant diversity indices, urban green space landscape metrics (using Systeme Probatoire d’Observation de la Terre (SPOT) data,), and social factors, including the type, population density, construction age, and GPS coordinates of each Urban Functional Unit, or UFU. We found that UFUs with more green space patches had higher cultivated and spontaneous species richness than those with fewer green space patches. Spontaneous species richness decreased when green space patches became fragmented, and it increased when green space patches were more connected (e.g., via land bridges). Conversely, cultivated species richness increased with green space patch fragmentation. The phylogenetic diversity of both cultivated and spontaneous plants were weakly associated with green space structure, which was strongly driven by land use. Old UFUs and those with larger populations had more green space patches overall, although they tended to be small and fragmented. Green space patch density was found to increase as the UFU age increased. From the viewpoint of knowledge transfer, understanding the effects and drivers of landscape patterns of urban green spaces could inform the development of improved policies and management of urban green space areas.
1. Introduction
Cities often differ greatly in terms of development history, rate and mode of urbanization, and cultural and social conditions [1,2]. The resulting urban fabric and associated landscape pattern could mold the distribution, composition, and diversity of urban flora. Landscape pattern refers to a spatial arrangement of various landscape elements of different sizes and shapes [3]. Recent studies have reported that landscape pattern plays an important role in influencing local plant community assemblage and diversity [4]. Spatial relationships among landscape elements present one of the key drivers of local biodiversity [5,6]. Landscape patterns have been found to explain much of the variance in species richness observed within forest ecosystems [7]. Several studies have shown that landscape pattern is an important determinant of species diversity through patch fragmentation and habitat modification [8]. Furthermore, landscape pattern influences many ecological processes, such as the distribution of materials and nutrients, and the persistence and movement of organisms [9,10]. Previous studies have even suggested that landscape patterns could be enlisted as an effective biodiversity indicator [6,11]. In urban areas, the biodiversity of cultural zones depends notably on land use [9,12], and especially on green space availability. The overall biodiversity in a city is contingent upon the size, quantity, and quality of urban green spaces [13], which are crucial for nurturing and protecting plant diversity [14].
Urban green spaces include natural, semi-natural, and artificial ecological systems within and around a city [15], such as the formal green infrastructure of public parks and gardens, and other green patches, such as remnant natural vegetation, urban waste-lands and vacant lots, and private gardens and yards [14]. Urban green spaces are key components of urban ecosystems, playing an important role in complex ecological processes [16] and providing significant ecosystem services, such as purifying air, reducing noise [17], mitigating urban heat island effects [18], and furnishing habitats for wildlife [19]. The diverse green spaces found within cities represent a gradient of economic and management inputs [12,20]. In modern times, the intensive anthropogenic impacts in urban regions necessitate an understanding of the key socioeconomic factors, such as land use pattern and structure [9], as well as other factors, such as population density, demographic profile, building age, and property price. The high land value in cities often restricts the provision of green space, especially in densely developed precincts. If a landowner can benefit notably from the use of urban land for commercial, industrial, and residential purposes, then that land is unlikely to be assigned to green infrastructure [21]. Moreover, the maintenance of urban green space is costly, and funding can become limited especially in times of economic stringency [22]. To ensure a sufficient supply of urban green spaces in conjunction with new developments and redevelopments, and to provide adequate maintenance to the existing stock, it is important to find persuasive justifications in their provision of multiple ecosystem services. Identifying the ecological role and conservation value of urban green space is therefore of critical importance to promote their protection, maintenance, and development [20].
Urban vegetation can be divided into two major categories: (1) cultivated vegetation in managed areas, including parks and street trees, and (2) spontaneous vegetation with growth and reproduction regulated by natural ecosystem processes independent of human inputs [23,24]. The management strategy of cultivated and spontaneous plants is markedly different; therefore, the response of these two types of plant diversity to urban landscape patterns may differ. Few studies have investigated the impact of different urban landscape patterns on the diversity of cultivated and spontaneous plant species. In addition, most studies on urban plant diversity were conducted in developed countries outside of the tropics. Tropical cities characterized by high inherent and modified biological diversity and in a densely-packed compact development mode have been neglected [25]. A recent study in Haikou, a tropical city in China, found that plant diversity was affected by land use and land cover, but it did not assess the effects of landscape patterns and different plant types [26]. It is necessary to fill this knowledge gap by conducting landscape ecology assessments of urban green space in tropical compact cities [16].
This study was carried out in the tropical coastal city Zhanjiang, located in Guangdong Province, south China. This city has undergone rapid urbanization in recent decades, bringing drastic landscape changes from natural to artificial. The changes in landscape patterns could be evaluated as transformations in the shape, area, quality, arrangement of green space patches, and distribution of individual landscape components [27]. The aims of this study were: (1) to identify the response of cultivated and spontaneous plants to the landscape pattern of urban green spaces; and (2) to determine the importance of social factors in relation to the landscape pattern of urban green spaces and in molding urban plant species diversity.
2. Materials and Methods
2.1. Study Area
Zhanjiang city (109°40′–110°58′E, 20°13′–21°57′N) is located within the Leizhou Peninsula at the western end of Guangdong Province on the south China coast (Figure 1). It covers a total area of 2217 km2, an urban area of 226 km2, and has a coastline of 1243.7 km—the third longest coastline of Chinese cities [28]. Average annual precipitation is between 1396 mm and 1723 mm, and the mean annual temperature is 23.4 °C. In 2018, the total population of Zhanjiang was 7.33 million, and in 2019, the gross domestic product (GDP) was about 306.47 billion Yuan (RMB) according to Zhanjiang’s government report. With competitive industries, Zhanjiang has become an important economic zone of the Beibuwan Region. With both rapid economic development and population growth, large tracts of agricultural land have been converted to nonagricultural and urban uses to accommodate recent urban expansion. Our study area within Zhanjiang covers 109.85 km2, with a total population of approximately 0.68 million as of 2018.
Figure 1.
A map of China showing the locations of (a) Zhanjiang city, (b) the study area outlined in red, and (c) the field survey units within the study area indicated by black dots.
2.2. Floristic Field Survey
A survey of the vascular plant diversity of the study area was conducted from July to August of 2017. The spatial sampling design was hierarchical, with the first step based on a grid of square cells measuring 0.65 km × 0.65 km, with one Urban Functional Unit (UFU) randomly selected within each cell. There was a total of 273 grid cells, although only 260 cells were surveyed as 13 cells were excluded due to a lack of green areas (e.g., areas containing plant life) or inaccessibility. In each selected UFU, three 20 m × 20 m plots were demarcated for tree diversity surveys (“tree plots”); within the four corners and center of each tree plot, five 5 m × 5 m subplots were set up for shrub diversity surveys (“shrub plots”); and within each shrub plot, a 1 m × 1 m subplot was set up for herb diversity surveys (“herb plots”). All species of trees, shrubs, and herbs were recorded in their respective sampling plots. Botanical nomenclature and taxonomy followed the standards of the Flora of China [29] and the Plant List (http://www.theplantlist.org/).
2.3. Species Richness and Phylogenetic Diversity Calculation
Within each sampling unit, species richness was determined according to the numbers of both cultivated and spontaneous plant species. For each UFU, three measures of phylogenetic diversity were calculated: Faith’s phylogenetic diversity (PD), which measures the sum of all phylogenetic distances; phylogenetic Mean Pairwise Distance (MPD), which measures the differences across the entire community and the mean distance between all species; and phylogenetic Mean Nearest Taxon Distance (MNTD), which identifies the similarities between co-occurring species and measures the mean distance between each species’ closest relative in a community [30,31]. We constructed a phylogenetic tree including all our vascular species using “Phylomatic” (http://phylodiversity.net/phylomatic/) and the software “Fig tree”. As a backbone, we used the PhytoPhylo tree of Qian & Jin [32]. Phylogenies generated by Scenario 3 are particularly robust in studies analyzing patterns of phylogenetic properties [32]. The observed PD, MPD, and MNTD values were compared to their expected values under a null model using respective standardized effect sizes (ses; PD.ses, MPD.ses, and MNTD.ses) to obtain phylogenetic diversity metrics that are independent of species richness [33]. We used the taxa.labels null model to simulate null assemblages (n = 999 randomizations) in the R package “picante”, which randomizes taxon labels on the phylogeny for the species included in the sampling pool [34]. MNTD.ses was found to be strongly correlated with PD.ses (cultivated plants Pearson’s r > 0.81, spontaneous plants Pearson’s r > 0.92) in this study; therefore, MNTD.ses was removed in further analysis to avoid multicollinearity (Table 1).

Table 1.
The correlation coefficients between eight pairs of biodiversity indices. The Mean Nearest Taxon Distance standardized effect size (MNTD.ses) of cultivated and spontaneous plants were removed because their correlation coefficients were more than |0.7|.
If the values of ses are negative, the phylogenetic diversity is lower than random, and if the values of ses are positive, the phylogenetic diversity is higher than random. Values that are significantly more negative than random suggest a phylogenetically clustered tendency, whereas those that are significantly more positive than random indicate an over-dispersed structure [35].
2.4. Landscape Analysis
For this analysis, we used the cloud free Systeme Probatoire d’Observation de la Terre (SPOT) remote sensing image with a spatial resolution of 15 m taken on 27 November 2016. Supervised classification by maximum likelihood classifier was adopted for image interpretation. The interpretation was completed in ArcGIS 10.2, which was developed by Esri (Esri, Environmental Systems Research Institute, Inc., Redlands, CA, USA) in the USA. The land uses were defined as: green space, road, building area, or water area. The overall classification accuracy was up to 85%. The interpreted land use data was then used to calculate landscape pattern metrics in FRAGSTATS 4.2 [36]. The diversity, density, splitting, cohesion, and splitting metrics of the landscape, based on the landscape level, were calculated with FRAGSTATS 4.2 [36] (Appendix A). At last, seven landscape metrics were used for further analyses based on correlation analysis (Table 2). The variable pairs with a correlation coefficient greater than 0.6 were removed according to correlation analysis (Table 3).
Table 2.
Descriptions of landscape pattern metrics used in this study.
Table 3.
The correlation coefficients between seven pairs of landscape metrics. Seven landscape metrics were kept because their correlation coefficients were less than |0.6|.
2.5. Socioeconomic Variables in Each Urban Functional Unit (UFU)
We chose to investigate UFU type, construction age, housing price, human population, and GPS coordinates as the key characteristics of each UFU because (1) accurate data of these five factors could be obtained; (2) UFU types refer to schools, factories, hospitals and so on, which are indicative of different land uses (Table 4); (3) construction age was previously linked to urbanization history; (4) housing price and GPS together could serve as a proxy for urbanization level in Zhanjiang, as the property prices at the city center and along coastal beaches were much higher than other areas; and (5) a large human population may contribute to more intensive disturbance of urban vegetation. We classified each UFU as one of the following types: Commercial (C), Institutional (I), Multi-family residential (M), Residential (R), Transportation (T), Utility (U), Factory (F), or Park (P) (Table 4). Construction age, or the length of time over which a UFU has been developed, was calculated as 2017 minus the year of establishment; for example, if established in 2010, a UFU would have a construction age of seven years. The date of establishment for each UFU was determined by one or more of the following methods: (1) checking the UFU’s official website, (2) interviewing the manager(s) of the UFU, and (3) analyzing past aerial photos compiled from Google Earth. Housing price data were acquired from the Taofang Property Website (www.taofang.com.cn), with query time from August to September of 2018; if the UFU was not a residential area, the housing price of the nearest residential area was assigned. The human population density of each UFU was calculated by dividing the total human population by the total area of the UFU (people/km2). We determined the total population of each UFU by (1) interviewing the UFU manager(s), (2) consulting the UFU’s official website, and (3) estimating the population in residential areas. In our population estimations, we assumed that if each apartment had an average of four occupants [38], then the total population of each apartment building would be equal to the number of domestic units multiplied by four occupants. The GPS coordinates of each UFU were measured using a GPS locator.
Table 4.
The number of Urban Functional Unit (UFU) types and their categories.
2.6. Data Analysis
A correlation analysis was performed among landscape metrics and plant diversity indices to exclude pairs that were too closely related. This screening permitted the use of seven landscape metrics and six diversity indices for subsequent data analysis (Table 5). All predictor variables were log transformed and standardized to achieve normalization. Because the UFU types are categorical variables, we assigned a numerical value to each UFU type: Commercial area (C) = 1, Factory (F) = 2, Institutional area (I) = 3, Multi-family residential (M) = 4, Park (P) = 5, Residential area (R) = 6, Transportation area (T) = 7, Utility (U) = 8. In this way, we could introduce UFU types into our model. We adopted Simple Linear Regression Models (LMs) to test the effect of single predictor variables on response variables. The predictor and response variables for plant diversity analyses were listed in Table 5. We then used p-values to select significant predictor variables for further analysis. We used General Linear Models (GLMs) to analyze how different social factors influence landscape patterns, and how different landscape patterns influence urban plant diversity. To find the subset of models that could best explain these landscape and plant diversity patterns, we employed a model selection procedure based on Akaike information criterion (AICc). Accordingly, we performed all the model combinations and chose models with the smallest AICc value.
Table 5.
Summary of response and predictor variables used in linear regression models (LM) to analyze plant diversity and landscape pattern; “spon.” and “cul.” refer to the spontaneous and cultivated species pools, respectively.
3. Results
3.1. Plant Species Richness and Phylogenetic Diversity
The predictor and response variables for plant diversity analysis were listed in Table 5. According to GLM results, plant diversity was influenced by Patch number (NP), Patch richness (PR), Shannon’s evenness index (SHEI), Splitting index (SPLIT), and Connectance. NP had a significant positive influence on the species richness of both cultivated and spontaneous species; for example, the more fragmented the landscape, the higher the cultivated and spontaneous species richness (cultivated species p = 0.0043, spontaneous species p = 0.0003). SPLIT had a significant positive influence on cultivated species richness, and a significant negative influence on spontaneous species richness (cultivated species p = 0.0137, spontaneous species p = 0.0639). Additionally, cultivated species richness was significantly negatively influenced by PR (p = 0.0516), and spontaneous species richness was significantly positively influenced by CONNECT (p = 0.0081) (Table 6).
Table 6.
Top models predicting species richness of spontaneous and cultivated species, and landscape pattern in Zhanjiang, Guangdong province, China. For each model, we reported the R-squared value (R2), and for each predictor its estimated coefficient.
As shown by the linear regression analysis (Figure 2), the relationships between the phylogenetic diversity of cultivated and spontaneous plants and the landscape pattern of green space were not significant. Only PD.ses of spontaneous plants were significantly affected by PR (R2 = 0.025; p = 0.0496); the PD.ses of cultivated plants was not affected by any of the landscape metrics. The MPD.ses of both cultivated and spontaneous plants were weakly affected by COHESION (cultivated plants R2 = 0.022, p= 0.0619; spontaneous plants R2 = 0.0231, p = 0.058093).
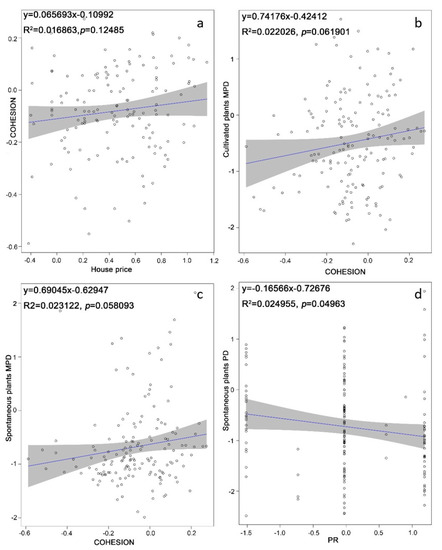
Figure 2.
The results of linear regression modeling (LMs): (a) House price weakly affected patch cohesion; (b,c) Cohesion significantly affected the Mean Pairwise Distance standardized effect size (MPD.ses) of both cultivated and spontaneous plants; (d) Patch richness (PR) significantly affected the phylogenetic diversity standardized effect size (PD.ses) of spontaneous plants. The shaded belt shows the 95% confidence interval. The trend line illustrates the trend depicted by the regression modeling.
3.2. Landscape Pattern and UFU Characteristics
The landscape metrics, except for COHESION, were significantly affected by all UFU characteristics: UFU type, construction age, and population density. Most of the landscape metrics were significantly affected by UFU type. Nearly all landscape metrics were positively affected by construction age and population density. NP and SPLIT were significantly positively affected by construction age and population density (construction age p = 0.0203; population density p = 0.006 for NP; construction age p = 0.0247; population density p = 0.001 for SPLIT). Additionally, DP was significantly positively affected by construction age, and PR was significantly positively affected by population density.
The patch SHEI, splitting index, density differences, and connectance among eight types of UFUs were significant. SHEI was the highest in institutional areas (0.94 ± 0.03) and the lowest in parks (0.78 ± 0.12). The splitting index was the highest in residential areas (11.88 ± 3.77) and the lowest in transport areas (4.25 ± 1.19). The highest green space patch density was in factories (6931.95 ± 1615.52) and the lowest green space patch density was in parks (2056.62 ± 936.69) (Figure 3). Connectance was the highest in transportation areas (77.58 ± 10.76) and the lowest in residential areas (8.64 ± 5.24).
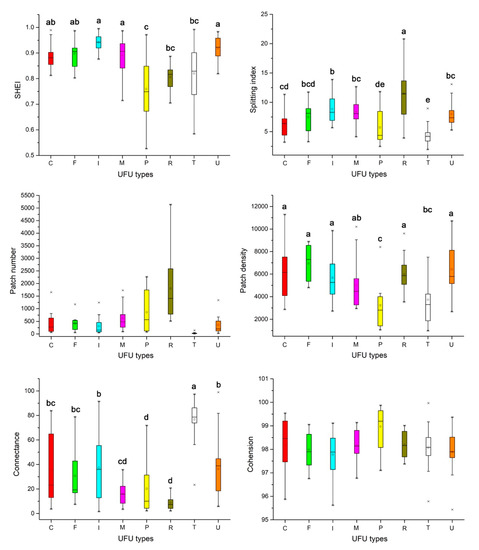
Figure 3.
The distribution of the landscape metrics of green space among eight UFU types. Shannon’s evenness index (SHEI), splitting index, patch density, and connectance differed significantly among these eight UFU types. a, b, c, d, e represent significant differences determined by Fisher’s least significant difference (LSD) tests (p < 0.05) on the same landscape metric for different UFU types.
4. Discussion
4.1. The Effect of Green Space Landscape Metrics on Cultivated and Spontaneous Plant Diversity
The size, shape, and distribution of urban green space play a decisive role in defining their ecological and landscape functions, such as biodiversity [39]. The ability of green space to support high biodiversity can also be affected by the urban structure [20]. The importance of the landscape context in determining species richness has been demonstrated for multiple taxa of a variety of plant life forms [40]. Garden management creates diverse vegetation cover, including perennially vegetated habitats, such as lawns, or annually vegetated habitats, such as vegetable beds [41]. These diverse microhabitats support urban biodiversity and have the ability to furnish a range of nature’s contributions to people [42].
Similar to Huston’s research [43], the species richness of both cultivated and spontaneous plants has been observed to increase with the number of green space patches available. Urbanization has induced landscape fragmentation and thus increased patch numbers, which has in turn led to increasingly smaller yet more numerous habitats for urban plants. In urban areas, man-made vegetation communities are usually constrained in small patches—it had been found that these small patches are important for urban biodiversity [44]. Different man-made vegetation communities are commonly composed of diverse collections of cultivated plant species, bringing high aggregate cultivated plant diversity as these green space patches increase in number. People tend to plant different cultivated species within different patches to bring high spatial variation in the aesthetic landscape; thus, the wide spatial spread of cultivated plant species mostly depends on human activities within urban areas. Ultimately, connectance among green space patches had no effect on the species richness of cultivated plant species due to the high intensity of human disturbance (Table 6). COHESION among green space patches had a weak effect on the phylogenetic diversity of cultivated plant species, such that the higher the cohesion, the greater the phylogenetic diversity (Figure 2b). This means that an increase in green space patch cohesion would not affect the species richness, but does have a weak effect on phylogenetic diversity. This relationship is likely due to the large variety of species that humans chose to plant in urban green spaces, as it was proven that cultivated non-native species promoted phylogenetic differentiation [45]. In other words, the cohesion of green space patches had nearly no effect on the diversity of cultivated plants. Overall, the green space landscape pattern has little affection on cultivated plant diversity.
Unlike cultivated plants, the species richness of spontaneous plants was significantly positively related to the patch cohesion index (Table 6). The phylogenetic diversity of spontaneous plants was weakly positively related to the patch cohesion index (Figure 2c). This means that for spontaneous plants, the patch cohesion effected species richness more than phylogenetic diversity. Thus, species richness can be inconsistent with phylogenetic diversity. This is because spontaneous plants in urban areas are more phylogenetically related, and this phylogenetic relatedness results in more similar functional traits and environmental adaptations than those exhibited by distant relatives [46]. The patch cohesion index could directly reflect the natural connectivity of the patches, with a high value indicating a low degree of fragmentation [47]. The inherent patchiness of land use in cities has generated urban green spaces that are often small and isolated [48]. The effect of fragmentation on species depends on their dispersal capacity and other functional attributes [49]. As many spontaneous plants are dispersed by animals, such species may be more influenced by landscape patterns, because their seeds need to be transported between habitat patches which may be separated by intervening hostile land use [50]. Some small annual species may be able to maintain populations in the smallest green space fragments [51]. Connectivity between patches is especially important for spontaneous plants, as the establishment of annual spontaneous plant assemblages has been attributed to connectivity among urban green space fragments at fine spatial scales [51]. Proponents of preserving and adding small green spaces to the urban landscape had expounded their contributions to the maintenance of stepping-stone connectivity, which benefits otherwise isolated plant populations [52]. Landscape fragmentation can impede gene flow, disrupt long-standing spatial genetic patterns, and impose strong structuring of the genetic diversity of populations and species, including highly mobile ones [53]. Overall, the magnitude of the contribution of small fragments to connectivity will depend on the characteristics of individual patches and focal biota [20]. Some research has even shown that the present landscape connectivity did not influence on plant species richness, while the past connectivity had a positive effect [44]. Spontaneous species were effectively filtered by urban environmental conditions, thus their existence depended more on long-lasting disturbances and changing environmental conditions. On the other hand, the dispersal and reproduction of cultivated plants are more influenced by inherent habitat traits, because their life cycles and survival are more dependent on human assistance and associated maintenance practices [54].
4.2. The Relationship between Social Factors and Green Space Landscape Metrics
The modern city is a key example of a human-dominated social-ecological system [55]. In this context, a green area can be understood as a social-ecological system or subsystem regulated by social and biophysical factors [56]. The type and magnitude of the human disturbance regimes constitute the key determinants of urban green space landscape characteristics [16]. In Zhanjiang, UFU type, construction age, and population density were three main social factors that affected all green space landscape metrics, with UFU type having a particularly significant effect (Table 6, Figure 3). The SHEI, SPLIT, DP, and Connectance were significantly different among different UFU types (Figure 3). Cities consist of a mosaic of individual UFUs that vary spatially by size, shape, and structure in conjunction with their respective socioeconomic functions. In a given city block, the main traits of UFUs are usually specific to their spatial constitution and arrangement [57]. Different UFU types have different functions which can influence differential land use, in turn modifying the composition and configuration of landscape patterns, [58], whereas the same land use tends to engender similar landscape patterns [59]. Thus, land use could regulate the amount and ecological quality of urban green spaces [16].
Population density positively affected patch number (NP), patch richness (PR), and splitting index (SPLIT) in the study area. SPLIT, associated with NP and PR, offers an appropriate measure to compare the fragmentation of regions of different sizes [60]. Higher values of NP and PR can result in higher SPLIT. Population density is another aspect of human activity which impacts urban greening: green space coverage declined in proportion to increasing human population density in European cities, excluding the UK [61]. Research in tropical Southeast Asia and Europe has reported that cities with higher population densities had less green space overall [59,61].
The construction age of UFUs reflected the effect of urban-development history on urban green space. Older UFUs within Zhanjiang registered higher values of PR, DP, and SPLIT, and tended to have more green space patches, which were typically small and highly fragmented. In newer UFUs, green spaces were installed in recent years and were thus reflective of greater attention to modern high-quality design standards, including provision of important ecosystem services. Liberal adoption of enhanced planning policies and management of urban green spaces would contribute to this enlightened trend [62]. A similar phenomenon was also observed in other tropical Chinese cities, such as Hong Kong and Shenzhen [16,62], where urban green spaces in old districts were found to be more fragmented than new ones. The fragmentation of urban green spaces in relation to urban-development age could thus be attributed to relevant urban-ecological knowledge and its application to public policies.
5. Conclusions
As some tropical coastal cities are anticipated to develop rapidly in the future, we could expect notable changes in their urban green space landscape patterns and associated urban plant diversity. It is necessary to acquire and apply relevant knowledge with the goal of improving spatial patterns of urban green spaces in fast-expanding metropolises. Uncovering the social drivers of urban green space landscape patterns and assessing their potential impacts on both cultivated and spontaneous plants is critical for optimizing the planning, design, and management of urban green spaces. Developing cities should aim to protect and ameliorate the quantity and quality of urban green space with a focus on enhancing their ecosystem services. The findings of this study are useful to inform the development of comprehensive, ecologically oriented, and socially relevant green space plans. For example, the landscape pattern of urban green space was significantly associated with UFU type—this means that UFU function plays an important role in deciding the landscape pattern of urban green spaces. Species richness was significantly associated with the number of green space patches; therefore, the more fragmentation of urban green space the greater the cultivated species richness, but the lower the spontaneous species richness. This phenomenon may depend on certain taxa which don’t need large habitat, such as cultivated plants and small spontaneous plants. This detailed empirical research was focused on one city, and it could be surmised that the drivers and effects of changes in green space patterns may differ in other cities. Further research in other cities within different climate zones, countries, and continents are therefore needed to validate or corroborate our findings. Although we were able to detect the above relationships in our data, it should be noted that the R2 values in our models are low. This indicates that there are some additional, important factors at play which may also affect plant diversity and green space landscape patterns. These factors have yet to be uncovered. Therefore, future work will also need to address this issue by focusing on some other factors at the city scale, such as policy, culture, greening investment, and so on.
Author Contributions
Conceptualization, H.F.W.; methodology, X.-L.C.; software, X.-L.C.; validation, H.-F.W.; formal analysis, X.-L.C. and M.M.N.; investigation, X.-L.C.; resources, X.-L.C. and H.F.W.; data curation, H.-F.W.; writing—original draft preparation, X.-L.C. and M.M.N.; writing—review and editing, X.-L.C., C.Y.J., S.Q., H.F.W., Z.-X.Z., and K.B.; visualization, H.-F.W. and L.-J.D.; supervision, H.-F.W.; project administration, H.-F.W.; funding acquisition, H.-F.W. and S.Q. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the Shanghai Key lab for urban Ecological processes and Eco-Restoration (SHUES2020A07) and a start-up fund from Hainan University [kyqd1633 and kyqd1840 (ZR)].
Acknowledgments
We thank Guoyin Cai for his help with the data analyses.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
The list of landscape metrics and plants diversity indices of each Urban Functional Unit in Zhanjiang, China.
References
- Gómez-Baggethun, E.; Barton, D.N. Classifying and valuing ecosystem services for urban planning. Ecol. Econ. 2013, 86, 235–245. [Google Scholar] [CrossRef]
- Fischer, L.K.; Rodorff, V.; von der Lippe, M.; Kowarik, I. Drivers of biodiversity patterns in parks of a growing South American megacity. Urban Ecosyst. 2016, 19, 1231–1249. [Google Scholar] [CrossRef]
- Liu, J.; Kuang, W.; Zhang, Z.; Xu, X.; Qin, Y.; Ning, J.; Zhou, W.; Zhang, S.; Li, R.; Yan, C.; et al. Spatiotemporal characteristics, patterns, and causes of land-use changes in China since the late 1980s. J. Geogr. Sci. 2014, 24, 195–210. [Google Scholar] [CrossRef]
- Torras, O.; Gil-Tena, A.; Saura, S. How does forest landscape structure explain tree species richness in a Mediterranean context? Biodivers. Conserv. 2007, 17, 1227–1240. [Google Scholar] [CrossRef]
- Wiens, J.A. Spatial Scaling in Ecology. Funct. Ecol. 1989, 3, 385. [Google Scholar] [CrossRef]
- Dauber, J.; Hirsch, M.; Simmering, D.; Waldhardt, R.; Otte, A.; Wolters, V. Landscape structure as an indicator of biodiversity: Matrix effects on species richness. Agric. Ecosyst. Environ. 2003, 98, 321–329. [Google Scholar] [CrossRef]
- Amici, V.; Rocchini, D.; Filibeck, G.; Bacaro, G.; Santi, E.; Geri, F.; Landi, S.; Scoppola, A.; Chiarucci, A. Landscape structure effects on forest plant diversity at local scale: Exploring the role of spatial extent. Ecol. Complex. 2015, 21, 44–52. [Google Scholar] [CrossRef]
- López-Martínez, J.O.; Hernández-Stefanoni, J.L.; Dupuy, J.M.; Meave, J.A. Partitioning the variation of woody plant β-diversity in a landscape of secondary tropical dry forests across spatial scales. J. Veg. Sci. 2012, 24, 33–45. [Google Scholar] [CrossRef]
- Walz, U. Landscape Structure, Landscape Metrics and Biodiversity. Living Rev. Landsc. Res. 2011, 5, 1–35. [Google Scholar] [CrossRef]
- Cheng, X.-L.; Yuan, L.-X.; Nizamani, M.M.; Zhu, Z.-X.; Friedman, C.R.; Wang, H.-F. Taxonomic and phylogenetic diversity of vascular plants at Ma’anling volcano urban park in tropical Haikou, China: Reponses to soil properties. PLoS ONE 2018, 6, e0198517. [Google Scholar] [CrossRef]
- Wang, H.-F.; Cheng, X.-L.; Nizamani, M.M.; Balfour, K.; Da, L.-J.; Zhu, Z.X.; Qureshi, S. An Integrated approach to study spatial patterns and drivers of land cover within urban functional units: A multi-city comparative study in China. Remote Sens. 2020, 7, 2201. [Google Scholar] [CrossRef]
- Sven, T.; Verhofstad, M.J.J.M.; Bakker, E.S. Managing Successional Stage Heterogeneity to Maximize Landscape-Wide Biodiversity of Aquatic Vegetation in Ditch Networks. Front. Plant Sci. 2018, 9, 1013. [Google Scholar] [CrossRef]
- Beninde, J.; Veith, M.; Hochkirch, A. Biodiversity in cities needs space: A meta-analysis of factors determining intra-urban biodiversity variation. Ecol. Lett. 2015, 18, 581–592. [Google Scholar] [CrossRef] [PubMed]
- Aronson, M.F.; Lepczyk, C.A.; Evans, K.L.; Goddard, M.A.; Lerman, S.B.; MacIvor, J.S.; Nilon, C.H.; Vargo, T. Biodiversity in the city: Key challenges for urban green space management. Front. Ecol. Environ. 2017, 15, 189–196. [Google Scholar] [CrossRef]
- Cilliers, S.; Cilliers, J.; Lubbe, R.; Siebert, S. Ecosystem services of urban green spaces in African countries—perspectives and challenges. Urban Ecosyst. 2012, 16, 681–702. [Google Scholar] [CrossRef]
- Tian, Y.; Jim, C.Y.; Wang, H. Assessing the landscape and ecological quality of urban green spaces in a compact city. Landsc. Urban Plan. 2014, 121, 97–108. [Google Scholar] [CrossRef]
- Cohen, P.; Potchter, O.; Schnell, I. The impact of an urban park on air pollution and noise levels in the Mediterranean city of Tel-Aviv, Israel. Environ. Pollut. 2014, 195, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Tan, Z.; Lau, K.K.-L.; Ng, E. Urban tree design approaches for mitigating daytime urban heat island effects in a high-density urban environment. Energy Build. 2016, 114, 265–274. [Google Scholar] [CrossRef]
- Chiquet, C.; Dover, J.W.; Mitchell, P. Birds and the urban environment: The value of green walls. Urban Ecosyst. 2012, 16, 453–462. [Google Scholar] [CrossRef]
- Lepczyk, C.A.; Aronson, M.F.J.; Evans, K.L.; Goddard, M.A.; Lerman, S.B.; MacIvor, J.S. Biodiversity in the City: Fundamental Questions for Understanding the Ecology of Urban Green Spaces for Biodiversity Conservation. BioScience 2017, 67, 799–807. [Google Scholar] [CrossRef]
- Kruize, H.; van der Vliet, N.; Staatsen, B.; Bell, R.; Chiabai, A.; Muiños, G.; Higgins, S.; Quiroga, S.; Martinez-Juarez, P.; Yngwe, M.A.; et al. Urban Green Space: Creating a Triple Win for Environmental Sustainability, Health, and Health Equity through Behavior Change. IJERPH 2019, 16, 4403. [Google Scholar] [CrossRef] [PubMed]
- Tempesta, T. Benefits and costs of urban parks: A review. Aestimum 2015, 127, 127–143. [Google Scholar]
- Cubino, J.P.; Cavender-Bares, J.; Hobbie, S.E.; Pataki, D.E.; Avolio, M.L.; Darling, L.E.; Larson, K.L.; Hall, S.J.; Groffman, P.M.; Trammell, T.L.E.; et al. Drivers of plant species richness and phylogenetic composition in urban yards at the continental scale. Landsc. Ecol. 2018, 34, 63–77. [Google Scholar] [CrossRef]
- Iuliana, P.P.; Adelina, D.; Valentin, S.; Doina, C.; Gergel, M. Ecological and aesthetic role of spontaneous flora in urban sustainable landscapes development. J. Plant Dev. 2011, 18, 169–177. [Google Scholar]
- Maffi, L. Biocultural Diversity Conservation; Routledge: London, UK, 2012. [Google Scholar]
- Zhu, Z.-X.; Pei, H.-Q.; Schamp, B.S.; Qiu, J.-X.; Cai, G.-Y.; Cheng, X.-L.; Wang, H.-F. Land cover and plant diversity in tropical coastal urban Haikou, China. Urban Urban Green. 2019, 44, 126395. [Google Scholar] [CrossRef]
- Zhou, N.Q.; Zhao, S. Urbanization process and induced environmental geological hazards in China. Nat. Hazards 2013, 67, 797–810. [Google Scholar] [CrossRef]
- Yu, J.; Chen, B.G.; Huang, Z.R.; Chen, Z.Z. Changes in the coastline of three typical bays in Guangdong during recent 10 years revealed by satellite image. Trans. Oceanol. Limnol. 2014, 3, 91–96. [Google Scholar]
- Chinese Academy of Sciences Editorial Committee of Flora Reipublicae Popularis Sinicae. Flora Reipublicae Popularis Sinicae; Science Press: Beijing, China, 1959–2004. [Google Scholar]
- Faith, D.P. Conservation evaluation and phylogenetic diversity. Biol. Conserv. 1992, 61, 1–10. [Google Scholar] [CrossRef]
- Tucker, C.M.; Cadotte, M.W.; Carvalho, S.B.; Davies, T.J.; Ferrier, S.; Fritz, S.A.; Grenyer, R.; Helmus, M.R.; Jin, L.S.; Mooers, A.O.; et al. A guide to phylogenetic metrics for conservation, community ecology and macroecology. Biol. Rev. 2016, 92, 698–715. [Google Scholar] [CrossRef]
- Qian, H.; Jin, Y. An updated megaphylogeny of plants, a tool for generating plant phylogenies and an analysis of phylogenetic community structure. JPECOL 2015, 9, 233–239. [Google Scholar] [CrossRef]
- Pavoine, S.; Bonsall, M.B. Measuring biodiversity to explain community assembly: A unified approach. Biol. Rev. 2010, 86, 792–812. [Google Scholar] [CrossRef]
- Kembel, S.W.; Cowan, P.D.; Helmus, M.R.; Cornwell, W.K.; Morlon, H.; Ackerly, D.D.; Blomberg, S.P.; Webb, C.O. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 2010, 26, 1463–1464. [Google Scholar] [CrossRef] [PubMed]
- Webb, C.O.; Ackerly, D.D.; Kembel, S.W. Phylocom: Software for the analysis of phylogenetic community structure and trait evolution. Bioinformatics 2008, 24, 2098–2100. [Google Scholar] [CrossRef]
- McGarigal, K.; Cushman, S.A.; Ene, E. FRAGSTATS v4: Spatial Pattern Analysis Program for Categorical and Continuous Maps. Computer Software Program Produced by the Authors at the University of Massachusetts, Amherst. 2012. Available online: http://www.umass.edu/landeco/research/fragstats/fragstats.html (accessed on 29 April 2020).
- Zhang, Z.Y. Families with Children and Household Financial Assets Allocation in Urban: Empirical Research; Xiangtan University: Xiangtan, China, 2016. [Google Scholar]
- Kong, F.; Yin, H.; Nakagoshi, N. Using GIS and landscape metrics in the hedonic price modeling of the amenity value of urban green space: A case study in Jinan City, China. Landsc. Urban Plan. 2007, 79, 240–252. [Google Scholar] [CrossRef]
- Zhang, N. Landscape Ecology; Science Press: Beijing, China, 2017. [Google Scholar]
- Prevedello, J.A.; Vieira, M.V. Does the type of matrix matter? A quantitative review of the evidence. Biodivers. Conserv. 2009, 19, 1205–1223. [Google Scholar] [CrossRef]
- Loram, A.; Warren, P.H.; Gaston, K.J. Urban Domestic Gardens (XIV): The Characteristics of Gardens in Five Cities. Environ. Manag. 2008, 42, 361–376. [Google Scholar] [CrossRef]
- Goddard, M.A.; Dougill, A.J.; Benton, T.G. Scaling up from gardens: Biodiversity conservation in urban environments. Trends Ecol. Evol. 2010, 25, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Huston, M.A. Local Processes and Regional Patterns: Appropriate Scales for Understanding Variation in the Diversity of Plants and Animals. Oikos 1999, 86, 393. [Google Scholar] [CrossRef]
- Wintle, B.A.; Kujala, H.; Whitehead, A.; Cameron, A.; Veloz, S.; Kukkala, A.; Moilanen, A.; Gordon, A.; Lentini, P.E.; Cadenhead, N.C.R.; et al. Global synthesis of conservation studies reveals the importance of small habitat patches for biodiversity. Proc. Natl. Acad. Sci. USA 2018, 116, 909–914. [Google Scholar] [CrossRef]
- Padullés, C.J.; Cavender-Bares, J.; Hobbie, S.E.; Hall, S.J.; Trammell, T.L.E.; Neill, C.; Avolio, M.L.; Darling, L.E.; Groffman, P.M. Contribution of non-native plants to the phylogenetic homogenization of U.S. yard floras. Ecosphere 2019, 10, e02638. [Google Scholar] [CrossRef]
- Macivor, J.S.; Cadotte, M.W.; Livingstone, S.W.; Lundholm, J.T.; Yasui, S.L.E. Phylogenetic ecology and the greening of cities. J. Appl. Ecol. 2016, 53, 1470–1476. [Google Scholar] [CrossRef]
- You, Q.; Fang, N.; Liu, L.; Yang, W.; Zhang, L.; Wang, Y. Effects of land use, topography, climate and socioeconomic factors on geographical variation pattern of inland surface water quality in China. PLoS ONE 2019, 14, e0217840. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Xu, C.; Pauleit, S.; Kindler, A.; Banzhaf, E. Spatial patterns of urban green infrastructure for equity: A novel exploration. J. Clean. Prod. 2019, 238, 117858. [Google Scholar] [CrossRef]
- Santamaría, S.; Sánchez, A.M.; López-Angulo, J.; Ornosa, C.; Mola, I.; Escudero, A. Landscape effects on pollination networks in Mediterranean gypsum islands. Plant Biol. J. 2017, 20, 184–194. [Google Scholar] [CrossRef] [PubMed]
- Damschen, E.I.; Baker, D.V.; Bohrer, G.; Nathan, R.; Orrock, J.L.; Turner, J.R.; Brudvig, L.A.; Haddad, N.M.; Levey, D.J.; Tewksbury, J.J. How fragmentation and corridors affect wind dynamics and seed dispersal in open habitats. Proc. Natl. Acad. Sci. USA 2014, 111, 3484–3489. [Google Scholar] [CrossRef]
- Luzuriaga, A.L.; Sánchez, A.M.; López-Angulo, J.; Escudero, A. Habitat fragmentation determines diversity of annual plant communities at landscape and fine spatial scales. Basic Appl. Ecol. 2018, 29, 12–19. [Google Scholar] [CrossRef]
- Rudd, H.; Vala, J.; Schaefer, V. Importance of Backyard Habitat in a Comprehensive Biodiversity Conservation Strategy: A Connectivity Analysis of Urban Green Spaces. Restor. Ecol. 2002, 10, 368–375. [Google Scholar] [CrossRef]
- Haag, T.; Santos, A.S.; Sana, D.A.; Morato, R.G.; Cullen, L., Jr.; Crawshaw, P.G., Jr.; De Angelo, C.; Di Bitetti, M.S.; Salzano, F.M.; Eizirik, E. The effect of habitat fragmentation on the genetic structure of a top predator: Loss of diversity and high differentiation among remnant populations of Atlantic Forest jaguars (Panthera onca). Mol. Ecol. 2010, 19, 4906–4921. [Google Scholar] [CrossRef]
- Song, H.; Xu, Y.; Hao, J.; Zhao, B.; Guo, D.; Shao, H. Investigating distribution pattern of species in a warm-temperate conifer-broadleaved-mixed forest in China for sustainably utilizing forest and soils. Sci. Total Environ. 2017, 578, 81–89. [Google Scholar] [CrossRef]
- Alberti, M.; Marzluff, J.M.; Shulenberger, E.; Bradley, G.; Ryan, C.; Zumbrunnen, C. Integrating humans into ecology: Opportunities and challenges for studying urban ecosystems. Urban Ecol. 2008, 53, 143–158. [Google Scholar]
- Silvis, H. The Economics of Ecosystems and Biodiversity in National and International Policymaking. Eur. Rev. Agric. 2011, 39, 186–188. [Google Scholar] [CrossRef]
- Yoshida, H.; Omae, M. An approach for analysis of urban morphology: Methods to derive morphological properties of city blocks by using an urban landscape model and their interpretations. Comput. Environ. Urban Syst. 2005, 29, 223–247. [Google Scholar] [CrossRef]
- Richards, D.R.; Passy, P.; Oh, R.R.Y. Impacts of Population Density and Wealth on the Quantity and Structure of Urban Green Space in Tropical Southeast Asia. Landsc. Urban Plan. 2017, 157, 553–560. [Google Scholar] [CrossRef]
- Zhang, Y.; Qin, K.; Bi, Q.; Cui, W.; Li, G. Landscape Patterns and Building Functions for Urban Land-Use Classification from Remote Sensing Images at the Block Level: A Case Study of Wuchang District, Wuhan, China. Remote Sens. 2020, 12, 1831. [Google Scholar] [CrossRef]
- Jaeger, J.A.G. Landscape Division, Splitting Index, and Effective Mesh Size: New Measures of Landscape Fragmentation. Landsc. Ecol. 2000, 15, 115–130. [Google Scholar] [CrossRef]
- Fuller, R.A.; Gaston, K.J. The scaling of green space coverage in European cities. Biol. Lett. 2009, 5, 352–355. [Google Scholar] [CrossRef]
- Qian, Y.; Li, Z.; Zhou, W.; Chen, Y. Quantifying Spatial Pattern of Urban Greenspace from a Gradient Perspective of Built-up Age. Phys. Chem. Earth 2019, 111, 78–85. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).