Capturing the Impact of the 2018 European Drought and Heat across Different Vegetation Types Using OCO-2 Solar-Induced Fluorescence

 ,
,  , , ,
, , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Description of Datasets
2.2.1. SIF Data
2.2.2. MODIS Data
2.2.3. Corine Land Cover Data
2.3. Data Analysis
3. Results
3.1. Overall Spring–Summer SIF Variation and Anomaly
3.2. Intraseasonal SIF Variation and Anomalies for Different Vegetation Types
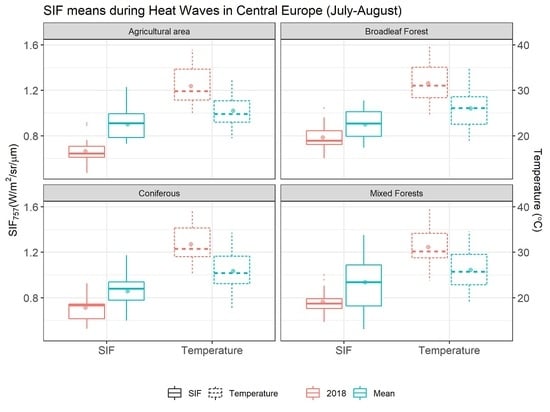
3.3. SIF Variation during the Heatwave
4. Discussion
4.1. Drought Impact on SIF
4.2. SIF Response during Drought Stress
4.3. OCO-2 SIF for Studying Drought Impact
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- IPCC. Summary for Policymakers. In Climate Change 2013: Contribution of Working Group I to the Fifth Assessment Report of the Intergouvernemental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2018. [Google Scholar]
- Dosio, A.; Mentaschi, L.; Fischer, E.M.; Wyser, K. Extreme Heat Waves Under 1.5 °C and 2 °C Global Warming. Environ. Res. Lett. 2018, 13, 054006. [Google Scholar] [CrossRef]
- Lhotka, O.; Kysely, J.; Farda, A. Climate Change Scenarios of Heat Waves in Central Europe and Their Uncertainties. Theor. Appl. Clim. 2017, 131, 1043–1054. [Google Scholar] [CrossRef]
- Van Loon, A.F.; Gleeson, T.; Clark, J.; Van Dijk, A.I.J.M.; Stahl, K.; Hannaford, J.; Di Baldassarre, G.; Teuling, A.J.; Tallaksen, L.M.; Uijlenhoet, R.; et al. Drought in the Anthropocene. Nat. Geosci. 2016, 9, 89–91. [Google Scholar] [CrossRef]
- Allen, C.D.; Breshears, D.D.; McDowell, N.G. On Underestimation of Global Vulnerability to Tree Mortality and Forest Die-off from Hotter Drought in the Anthropocene. Ecosphere 2015, 6, art129. [Google Scholar] [CrossRef]
- Buras, A.; Rammig, A.; Zang, C. Quantifying Impacts of the 2018 Drought on European Ecosystems in Comparison to 2003. Biogeosciences 2020, 17, 1655–1672. [Google Scholar] [CrossRef]
- Shekhar, A.; Bhattacharjee, S.; Chen, J.; Rammig, A. Spring-Summer Variation Analysis in OCO-2’s Solar Induced Fluorescence During the European Heatwave in 2018. In Geophysical Research Abstracts; Copernicus Publications: Gottingen, Germany, 2019. [Google Scholar]
- Heatwave in Northern Europe, Summer 2018. Available online: https://www.worldweatherattribution.org/attribution-of-the-2018-heat-in-northern-europe/ (accessed on 10 August 2020).
- Ciais, P.; Reichstein, M.; Viovy, N.; Granier, A.; Ogée, J.; Allard, V.; Aubinet, M.; Buchmann, N.; Bernhofer, C.; Carrara, A.; et al. Europe-Wide Reduction in Primary Productivity Caused by the Heat and Drought in 2003. Nature 2005, 437, 529–533. [Google Scholar] [CrossRef]
- Phillips, O.L.; Aragão, L.E.O.C.; Lewis, S.L.; Fisher, J.B.; Lloyd, J.; Lopez-Gonzalez, G.; Malhi, Y.; Monteagudo, A.; Peacock, J.; Quesada, C.A.; et al. Drought Sensitivity of the Amazon Rainforest. Science 2009, 323, 1344–1347. [Google Scholar] [CrossRef]
- Beer, C.; Reichstein, M.; Tomelleri, E.; Ciais, P.; Jung, M.; Carvalhais, N.; Rödenbeck, C.; Arain, M.A.; Baldocchi, D.; Bonan, G.B.; et al. Terrestrial Gross Carbon Dioxide Uptake: Global Distribution and Covariation with Climate. Science 2010, 329, 834–838. [Google Scholar] [CrossRef]
- Lawrence, D.M.; Oleson, K.W.; Flanner, M.G.; Thornton, P.E.; Swenson, S.; Lawrence, P.J.; Zeng, X.; Yang, Z.-L.; Levis, S.; Sakaguchi, K.; et al. Parameterization Improvements and Functional and Structural Advances in Version 4 of the Community Land Model. J. Adv. Model. Earth Syst. 2011, 3. [Google Scholar] [CrossRef]
- Li, X.; Xiao, J.; He, B. Chlorophyll Fluorescence Observed by OCO-2 Is Strongly Related to Gross Primary Productivity Estimated From Flux Towers in Temperate Forests. Remote Sens. Environ. 2018, 204, 659–671. [Google Scholar] [CrossRef]
- Smith, W.K.; Biederman, J.A.; Scott, R.L.; Moore, D.J.P.; He, M.; Kimball, J.S.; Yan, D.; Hudson, A.; Barnes, M.L.; MacBean, N.; et al. Chlorophyll Fluorescence Better Captures Seasonal and Interannual Gross Primary Productivity Dynamics Across Dryland Ecosystems of Southwestern North America. Geophys. Res. Lett. 2018, 45, 748–757. [Google Scholar] [CrossRef]
- Sun, Y.; Frankenberg, C.; Wood, J.; Schimel, D.S.; Jung, M.; Guanter, L.; Drewry, D.; Verma, M.; Porcar-Castell, A.; Griffis, T.; et al. OCO-2 Advances Photosynthesis Observation From Space via Solar-Induced Chlorophyll Fluorescence. Science 2017, 358, eaam5747. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Li, X.; He, B.; Arain, M.A.; Beringer, J.; Desai, A.R.; Emmel, C.; Hollinger, D.Y.; Krasnova, A.; Mammarella, I.; et al. Solar-Induced Chlorophyll Fluorescence Exhibits a Universal Relationship with Gross Primary Productivity Across a Wide Variety of Biomes. Glob. Chang. Boil. 2019, 25, 4. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Tang, J.; Mustard, J.F.; Lee, J.-E.; Rossini, M.; Joiner, J.; Munger, J.W.; Kornfeld, A.; Richardson, A.D. Solar-Induced Chlorophyll Fluorescence That Correlates With Canopy Photosynthesis on Diurnal and Seasonal Scales in a Temperate Deciduous Forest. Geophys. Res. Lett. 2015, 42, 2977–2987. [Google Scholar] [CrossRef]
- Baker, N.R. Chlorophyll Fluorescence: A Probe of Photosynthesis In Vivo. Annu. Rev. Plant. Boil. 2008, 59, 89–113. [Google Scholar] [CrossRef]
- Meroni, M.; Rossini, M.; Guanter, L.; Alonso, L.; Rascher, U.; Colombo, R.; Moreno, J. Remote Sensing of Solar-Induced Chlorophyll Fluorescence: Review of Methods and Applications. Remote Sens. Environ. 2009, 113, 2037–2051. [Google Scholar] [CrossRef]
- Zarco-Tejada, P.; Morales, A.; Testi, L.; Villalobos, F.J. Spatio-Temporal Patterns of Chlorophyll Fluorescence and Physiological and Structural Indices Acquired From Hyperspectral Imagery as Compared With Carbon Fluxes Measured With Eddy Covariance. Remote Sens. Environ. 2013, 133, 102–115. [Google Scholar] [CrossRef]
- Frankenberg, C.; Fisher, J.B.; Worden, J.; Badgley, G.; Saatchi, S.S.; Lee, J.-E.; Toon, G.C.; Butz, A.; Jung, M.; Kuze, A.; et al. New Global Observations of the Terrestrial Carbon Cycle From GOSAT: Patterns of Plant Fluorescence With Gross Primary Productivity. Geophys. Res. Lett. 2011, 38, 1–6. [Google Scholar] [CrossRef]
- Guanter, L.; Zhang, Y.; Jung, M.; Joiner, J.; Voigt, M.; Berry, J.A.; Frankenberg, C.; Huete, A.R.; Zarco-Tejada, P.; Lee, J.-E.; et al. Global and Time-Resolved Monitoring of Crop Photosynthesis With Chlorophyll Fluorescence. Proc. Natl. Acad. Sci. USA 2014, 111, E1327–E1333. [Google Scholar] [CrossRef]
- Joiner, J.; Vasilkov, A.P.; Yoshida, Y.; Corp, L.A.; Middleton, E.M. First Observations of Global and Seasonal Terrestrial Chlorophyll Fluorescence From Space. Biogeosciences 2011, 8, 637–651. [Google Scholar] [CrossRef]
- Parazoo, N.C.; Bowman, K.; Fisher, J.B.; Frankenberg, C.; Jones, D.B.A.; Cescatti, A.; Priego, Ó.P.; Wohlfahrt, G.; Montagnani, L. Terrestrial Gross Primary Production Inferred From Satellite Fluorescence and Vegetation Models. Glob. Chang. Boil. 2014, 20, 3103–3121. [Google Scholar] [CrossRef] [PubMed]
- Sanders, A.F.J.; Verstraeten, W.W.; Kooreman, M.L.; Van Leth, T.C.; Beringer, J.; Joiner, J. Spaceborne Sun-Induced Vegetation Fluorescence Time Series from 2007 to 2015 Evaluated with Australian Flux Tower Measurements. Remote Sens. 2016, 8, 895. [Google Scholar] [CrossRef]
- Verma, M.; Schimel, D.; Evans, B.; Frankenberg, C.; Beringer, J.; Drewry, D.T.; Magney, T.S.; Marang, I.; Hutley, L.B.; Moore, C.; et al. Effect of Environmental Conditions on the Relationship Between Solar-Induced Fluorescence and Gross Primary Productivity at an Ozflux Grassland Site. J. Geophys. Res. Biogeosciences 2017, 122, 716–733. [Google Scholar] [CrossRef]
- Wood, J.; Griffis, T.; Baker, J.; Frankenberg, C.; Verma, M.; Yuen, K. Multiscale Analyses of Solar-Induced Florescence and Gross Primary Production. Geophys. Res. Lett. 2017, 44, 533–541. [Google Scholar] [CrossRef]
- Zhang, Y.; Guanter, L.; Berry, J.A.; Joiner, J.; Van Der Tol, C.; Huete, A.R.; Gitelson, A.; Voigt, M.; Köhler, P. Estimation of Vegetation Photosynthetic Capacity From Space-Based Measurements of Chlorophyll Fluorescence for Terrestrial Biosphere Models. Glob. Chang. Boil. 2014, 20, 3727–3742. [Google Scholar] [CrossRef] [PubMed]
- Frankenberg, C.; O’Dell, C.; Berry, J.; Guanter, L.; Joiner, J.; Köhler, P.; Pollock, R.; Taylor, T. Prospects for Chlorophyll Fluorescence Remote Sensing from the Orbiting Carbon Observatory-2. Remote Sens. Environ. 2014, 147, 1–12. [Google Scholar] [CrossRef]
- Lee, J.-E.; Frankenberg, C.; Van Der Tol, C.; Berry, J.A.; Guanter, L.; Boyce, C.K.; Fisher, J.B.; Morrow, E.; Worden, J.R.; Asefi, S.; et al. Forest Productivity and Water Stress in Amazonia: Observations from Gosat Chlorophyll Fluorescence. Proc. R. Soc. B: Boil. Sci. 2013, 280, 20130171. [Google Scholar] [CrossRef]
- Yoshida, Y.; Joiner, J.; Tucker, C.; Berry, J.; Lee, J.-E.; Walker, G.; Reichle, R.; Koster, R.; Lyapustin, A.; Wang, Y. The 2010 Russian Drought Impact on Satellite Measurements of Solar-Induced Chlorophyll Fluorescence: Insights From Modeling and Comparisons with Parameters Derived From Satellite Reflectances. Remote Sens. Environ. 2015, 166, 163–177. [Google Scholar] [CrossRef]
- Sun, Y.; Fu, R.; Dickinson, R.; Joiner, J.; Frankenberg, C.; Gu, L.; Xia, Y.; Fernando, D.N. Drought Onset Mechanisms Revealed by Satellite Solar-Induced Chlorophyll Fluorescence: Insights From Two Contrasting Extreme Events. J. Geophys. Res. Biogeosciences 2015, 120, 2427–2440. [Google Scholar] [CrossRef]
- Koren, G.; Van Schaik, E.; Araújo, A.C.; Boersma, K.F.; Gärtner, A.; Killaars, L.; Kooreman, M.L.; Kruijt, B.; Luijkx, I.T.; Von Randow, C.; et al. Widespread Reduction in Sun-Induced Fluorescence From the Amazon during the 2015/2016 El Niño. Philos. Trans. R. Soc. B: Boil. Sci. 2018, 373, 20170408. [Google Scholar] [CrossRef]
- Zhang, Y.; Joiner, J.; Alemohammad, S.H.; Zhou, S.; Gentine, P. A Global Spatially Contiguous Solar-Induced Fluorescence (CSIF) Dataset Using Neural Networks. Biogeosciences 2018, 15, 5779–5800. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I. A Multiscalar Drought Index Sensitive to Global Warming: The Standardized Precipitation Evapotranspiration Index. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef]
- NOAA National Centers for Environmental Information State of the Climate. Global Climate Report for August 2018; NCEI: Asheville, NC, USA, 2018. [Google Scholar]
- NOAA National Centers for Environmental Information State of the Climate. Global Climate Report for July 2018; NCEI: Asheville, NC, USA, 2018. [Google Scholar]
- Damm, A.; Elbers, J.; Erler, A.; Gioli, B.; Hamdi, K.; Hutjes, R.; Košvancová, M.; Meroni, M.; Miglietta, F.; Moersch, A.; et al. Remote Sensing of Sun-Induced Fluorescence to Improve Modeling of Diurnal Courses of Gross Primary Production (GPP). Glob. Chang. Boil. 2010, 16, 171–186. [Google Scholar] [CrossRef]
- Damm, A.; Guanter, L.; Paul-Limoges, E.; Van Der Tol, C.; Hueni, A.; Buchmann, N.; Eugster, W.; Ammann, C.; Schaepman, M.E. Far-Red Sun-Induced Chlorophyll Fluorescence Shows Ecosystem-Specific Relationships to Gross Primary Production: An Assessment Based on Observational and Modeling Approaches. Remote Sens. Environ. 2015, 166, 91–105. [Google Scholar] [CrossRef]
- Frankenberg, C.; Berry, J. Solar Induced Chlorophyll Fluorescence: Origins, Relation to Photosynthesis and Retrieval; Elsevier: Amsterdam, The Netherlands, 2018; pp. 143–162. [Google Scholar]
- Frankenberg, C.; Butz, A.; Toon, G.C. Disentangling Chlorophyll Fluorescence From Atmospheric Scattering Effects in O2 A Band Spectra of Reflected Sun-Light. Geophys. Res. Lett. 2011, 38. [Google Scholar] [CrossRef]
- Sun, Y.; Frankenberg, C.; Jung, M.; Joiner, J.; Guanter, L.; Köhler, P.; Magney, T.S. Overview of Solar-Induced Chlorophyll Fluorescence (SIF) from the Orbiting Carbon Observatory-2: Retrieval, Cross-Mission Comparison, and Global Monitoring for GPP. Remote Sens. Environ. 2018, 209, 808–823. [Google Scholar] [CrossRef]
- Shekhar, A.; Chen, J.; Paetzold, J.C.; Dietrich, F.; Zhao, X.; Bhattacharjee, S.; Ruisinger, V.; Wofsy, S.C. Anthropogenic CO2 Emissions Assessment of Nile Delta Using XCO2 and Sif Data From OCO-2 Satellite. Environ. Res. Lett. 2020, 15, 095010. [Google Scholar] [CrossRef]
- Frankenberg, C. Solar Induced Chlorophyll Fluorescence: OCO-2 Lite Files (B7000) User Guide; California Institute of Technology: Pasadena, CA, USA, 2015. [Google Scholar]
- Zhang, Z.; Zhang, Y.; Joiner, J.; Migliavacca, M. Angle Matters: Bidirectional Effects Impact the Slope of Relationship Between Gross Primary Productivity and Sun-Induced Chlorophyll Fluorescence From Orbiting Carbon Observatory-2 Across Biomes. Glob. Chang. Boil. 2018, 24, 5017–5020. [Google Scholar] [CrossRef]
- Goulas, Y.; Daumard, F.; Ounis, A.; Rhoul, C.; Lopez, M.L.; Moya, I. Monitoring the Diurnal Time Course of Vegetation Dynamics with Geostationary Observations: The Gflex Project. In Proceedings of the 6th Workshop on Hyperspectral Image and Signal Processing: Evolution in Remote Sensing (WHISPERS), Laussane, Switzerland, 24–27 June 2014; pp. 1–4. [Google Scholar] [CrossRef]
- Yan, K.; Park, T.; Yan, G.; Chen, C.; Yang, B.; Liu, Z.; Nemani, R.R.; Knyazikhin, Y.; Myneni, R.B. Evaluation of MODIS LAI/FPAR Product Collection 6. Part 1: Consistency and Improvements. Remote Sens. 2016, 8, 359. [Google Scholar] [CrossRef]
- Yan, K.; Park, T.; Yan, G.; Liu, Z.; Yang, B.; Chen, C.; Nemani, R.R.; Knyazikhin, Y.; Myneni, R.B. Evaluation of MODIS LAI/FPAR Product Collection 6. Part 2: Validation and Intercomparison. Remote Sens. 2016, 8, 460. [Google Scholar] [CrossRef]
- Knyazikhin, Y.; Myneni, R.B.; Privette, J.L.; Running, S.W.; Nemani, R.; Zhang, Y.; Tian, Y.; Wang, Y.; Morissette, J.T.; Glassy, J.; et al. MODIS Leaf Area Index (LAI) and Fraction of Photosynthetically Active Radiation Absorbed by Vegetation (FPAR) Product (MOD15) Algorithm Theoretical Basis Document; Boston University: Boston, MA, USA, 1999. [Google Scholar]
- Solano, R.; Didan, K.; Jacobson, A.; Huete, A. MODIS Vegetation Index User’s Guide, MOD13 Series; University of Arizona: Tucson, AZ, USA, 2010. [Google Scholar]
- Büttner, G.; Kosztra, B. CLC2018 Technical Guidelines; EEA: Wien, Austria, 2017; pp. 1–60. [Google Scholar]
- Wable, P.S.; Jha, M.K.; Shekhar, A. Comparison of Drought Indices in a Semi-Arid River Basin of India. Water Resour. Manag. 2018, 33, 75–102. [Google Scholar] [CrossRef]
- Nam, W.-H.; Hayes, M.J.; Svoboda, M.D.; Tadesse, T.; Wilhite, D.A. Drought Hazard Assessment in the Context of Climate Change For South Korea. Agric. Water Manag. 2015, 160, 106–117. [Google Scholar] [CrossRef]
- Zang, C.; Buras, A.; Esquivel-Muelbert, A.; Jump, A.S.; Rigling, A.; Rammig, A. Standardized Drought Indices in Ecological Research: Why One Size Does Not Fit All. Glob. Chang. Boil. 2019, 26, 322–324. [Google Scholar] [CrossRef] [PubMed]
- Ionita, M.; Tallaksen, L.M.; Kingston, D.G.; Stagge, J.H.; Laaha, G.; Van Lanen, H.; Scholz, P.; Chelcea, S.M.; Haslinger, K. The European 2015 Drought From a Climatological Perspective. Hydrol. Earth Syst. Sci. 2017, 21, 1397–1419. [Google Scholar] [CrossRef]
- Dong, B.; Sutton, R.T.; Shaffrey, L.; Wilcox, L.J. The 2015 European Heat Wave. Bull. Am. Meteorol. Soc. 2016, 97, S57–S62. [Google Scholar] [CrossRef]
- McCornack, R.L. Extended Tables of the Wilcoxon Matched Pair Signed Rank Statistic. J. Am. Stat. Assoc. 1965, 60, 864–871. [Google Scholar] [CrossRef]
- KMI. Klimatologisch Maandoverzicht Juli 2018 (PDF); KMI: Brussels, Belgium. (In Dutch)
- KNMI—Hittegolf. 2019. Available online: www.knmi.nl (accessed on 10 August 2020).
- Danmarks Meteorologiske Institut. Juni bliver den varmeste og solrigeste i 26 år.; DMI: Copenhague, Denmark, 2018. (In Danish) [Google Scholar]
- DWD. Deutscher Wetterdienst; DWD: Offenbach, Germany, 2018. [Google Scholar]
- R Development Core Team. A Language and Environment for Statistical Computing. R. Found. Stat. Comput. 2011, 1, 409. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Wickham, H.; Francois, R. dplyr: A Grammar of Data Manipulation, R Packag. version 0.4.2; R Studio: Boston, MA, USA, 2015. [Google Scholar]
- Shekhar, A.; Shapiro, C.A. What Do Meteorological Indices Tell Us About a Long-Term Tillage Study? Soil Tillage Res. 2019, 193, 161–170. [Google Scholar] [CrossRef]
- Paul-Limoges, E.; Damm, A.; Hueni, A.; Liebisch, F.; Eugster, W.; Schaepman, M.E.; Buchmann, N. Effect of Environmental Conditions on Sun-Induced Fluorescence in a Mixed Forest and a Cropland. Remote Sens. Environ. 2018, 219, 310–323. [Google Scholar] [CrossRef]
- Miralles, D.G.; Teuling, A.J.; Van Heerwaarden, C.C.; De Arellano, J.V.-G. Mega-Heatwave Temperatures Due to Combined Soil Desiccation and Atmospheric Heat Accumulation. Nat. Geosci. 2014, 7, 345–349. [Google Scholar] [CrossRef]
- Sippel, S.; Forkel, M.; Rammig, A.; Thonicke, K.; Flach, M.; Heimann, M.; Otto, F.E.L.; Reichstein, M.; Mahecha, M.D. Contrasting and Interacting Changes in Simulated Spring and Summer Carbon Cycle Extremes in European Ecosystems. Environ. Res. Lett. 2017, 12, 075006. [Google Scholar] [CrossRef]
- Porcar-Castell, A.; Tyystjärvi, E.; Atherton, J.; Van Der Tol, C.; Flexas, J.; Pfündel, E.E.; Moreno, J.; Frankenberg, C.; Berry, J.A. Linking Chlorophyll a Fluorescence to Photosynthesis for Remote Sensing Applications: Mechanisms and Challenges. J. Exp. Bot. 2014, 65, 4065–4095. [Google Scholar] [CrossRef] [PubMed]
- Crausbay, S.D.; Ramirez, A.R.; Carter, S.L.; Cross, M.S.; Hall, K.R.; Bathke, D.; Betancourt, J.L.; Colt, S.; Cravens, A.E.; Dalton, M.S.; et al. Defining Ecological Drought for the Twenty-First Century. Bull. Am. Meteorol. Soc. 2017, 98, 2543–2550. [Google Scholar] [CrossRef]
- Sippel, S.; Zscheischler, J.; Reichstein, M. Ecosystem Impacts of Climate Extremes Crucially Depend on the Timing. Proc. Natl. Acad. Sci. USA 2016, 113, 5768–5770. [Google Scholar] [CrossRef]
- Chen, J.; Franklin, J.F.; Spies, T.A. Contrasting Microclimates Among Clearcut, Edge, and Interior of Old-Growth Douglas-Fir Forest. Agric. For. Meteorol. 1993, 63, 219–237. [Google Scholar] [CrossRef]
- Chen, J.; Saunders, S.C.; Crow, T.R.; Naiman, R.J.; Brosofske, K.D.; Mroz, G.D.; Brookshire, B.L.; Franklin, J.F. Microclimate in Forest Ecosystem and Landscape Ecology. Bioscience 1999, 49, 288–297. [Google Scholar] [CrossRef]
- Young, A.; Mitchell, N. Microclimate and Vegetation Edge Effects in a Fragmented Podocarp-Broadleaf Forest in New Zealand. Boil. Conserv. 1994, 67, 63–72. [Google Scholar] [CrossRef]
- Krishnan, P.; Black, T.A.; Grant, N.J.; Barr, A.G.; Hogg, E.; Jassal, R.S.; Morgenstern, K. Impact of Changing Soil Moisture Distribution on Net Ecosystem Productivity of a Boreal Aspen Forest During and Following Drought. Agric. For. Meteorol. 2006, 139, 208–223. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Schwalm, C.; Biondi, F.; Camarero, J.J.; Koch, G.; Litvak, M.; Ogle, K.; Shaw, J.D.; Shevliakova, E.; Williams, A.P.; et al. Pervasive Drought Legacies in Forest Ecosystems and Their Implications for Carbon Cycle Models. Science 2015, 349, 528–532. [Google Scholar] [CrossRef]
- Buras, A.; Schunk, C.; Zeiträg, C.; Herrmann, C.; Kaiser, L.; Lemme, H.; Straub, C.; Taeger, S.; Gößwein, S.; Klemmt, H.-J.; et al. Are Scots Pine Forest Edges Particularly Prone to Drought-Induced Mortality? Environ. Res. Lett. 2018, 13, 025001. [Google Scholar] [CrossRef]
- Forstmaier, A.; Shekhar, A.; Chen, J. Mapping of Eucalyptus in Natura 2000 areas using Sentinel 2 imagery and artificial neural networks. Remote Sens. 2020, 12, 2176. [Google Scholar] [CrossRef]
- Dobrowski, S.Z.; Pushnik, J.; Zarco-Tejada, P.; Ustin, S. Simple Reflectance Indices Track Heat and Water Stress-Induced Changes in Steady-State Chlorophyll Fluorescence at the Canopy Scale. Remote Sens. Environ. 2005, 97, 403–414. [Google Scholar] [CrossRef]
- Wang, S.; Huang, C.; Zhang, L.; Lin, Y.; Cen, Y.; Wu, T. Monitoring and Assessing the 2012 Drought in the Great Plains: Analyzing Satellite-Retrieved Solar-Induced Chlorophyll Fluorescence, Drought Indices, and Gross Primary Production. Remote Sens. 2016, 8, 61. [Google Scholar] [CrossRef]
- De Boeck, H.J.; Verbeeck, H. Drought-Associated Changes in Climate and Their Relevance for Ecosystem Experiments and Models. Biogeosciences 2011, 8, 1121–1130. [Google Scholar] [CrossRef]
- Fischer, E.M.; Seneviratne, S.I.; Vidale, P.L.; Luthi, D.; Schär, C. Soil Moisture–Atmosphere Interactions during the 2003 European Summer Heat Wave. J. Clim. 2007, 20, 5081–5099. [Google Scholar] [CrossRef]
- Chaves, M.M. Effects of Water Deficits on Carbon Assimilation. J. Exp. Bot. 1991, 42, 1–16. [Google Scholar] [CrossRef]
- Keenan, T.F.; Sabaté, S.; Gracia, C. The Importance of Mesophyll Conductance in Regulating Forest Ecosystem Productivity During Drought Periods. Glob. Chang. Boil. 2010, 16, 1019–1034. [Google Scholar] [CrossRef]
- Bréda, N.; Huc, R.; Granier, A.; Dreyer, E. Temperate Forest Trees and Stands Under Severe Drought: A Review of Ecophysiological Responses, Adaptation Processes and Long-Term Consequences. Ann. For. Sci. 2006, 63, 625–644. [Google Scholar] [CrossRef]
- Castro, A.O.; Chen, J.; Zang, C.; Shekhar, A.; Jiménez-Muñoz, J.C.; Bhattacharjee, S.; Kindu, M.; Morales, V.H.; Rammig, A. OCO-2 Solar-Induced Chlorophyll Fluorescence Variability across Ecoregions of the Amazon Basin and the Extreme Drought Effects of El Niño (2015–2016). Remote Sens. 2020, 12, 1202. [Google Scholar] [CrossRef]
- Kasahara, M.; Kagawa, T.; Oikawa, K.; Suetsugu, N.; Miyao, M.; Wada, M. Chloroplast Avoidance Movement Reduces Photodamage in Plants. Nature 2002, 420, 829–832. [Google Scholar] [CrossRef]
- Field, C.B. Ecological Scaling of Carbon Gain to Stress and Resource Availability. In Response of Plants to Multiple Stresses; Elsevier: Amsterdam, The Netherlands, 1991; pp. 35–65. [Google Scholar]
- Flexas, J.; Medrano, H. Drought-inhibition of Photosynthesis in C3 Plants: Stomatal and Non-stomatal Limitations Revisited. Ann. Bot. 2002, 89, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Peguero-Pina, J.J.; Morales, F.; Flexas, J.; Gil-Pelegrín, E.; Moya, I. Photochemistry, Remotely Sensed Physiological Reflectance Index and de-Epoxidation State of the Xanthophyll Cycle in Quercus Coccifera Under Intense Drought. Oecologia 2008, 156, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Van Der Tol, C. Linking Canopy Scattering of Far-Red Sun-Induced Chlorophyll Fluorescence with Reflectance. Remote Sens. Environ. 2018, 209, 456–467. [Google Scholar] [CrossRef]
- Zeng, Y.; Badgley, G.; DeChant, B.; Ryu, Y.; Chen, M.; Berry, J. A Practical Approach for Estimating the Escape Ratio of Near-Infrared Solar-Induced Chlorophyll Fluorescence. Remote Sens. Environ. 2019, 232, 111209. [Google Scholar] [CrossRef]
- Lee, J.-E.; Berry, J.; Van Der Tol, C.; Yang, X.; Guanter, L.; Damm, A.; Baker, I.; Frankenberg, C. Simulations of Chlorophyll Fluorescence Incorporated Into the Community Land Model Version 4. Glob. Chang. Boil. 2015, 21, 3469–3477. [Google Scholar] [CrossRef]
- Van Der Tol, C.; Berry, J.A.; Campbell, P.K.E.; Rascher, U. Models of Fluorescence and Photosynthesis for Interpreting Measurements of Solar-Induced Chlorophyll Fluorescence. J. Geophys. Res. Biogeosciences 2014, 119, 2312–2327. [Google Scholar] [CrossRef]
- Bhattacharjee, S.; Chen, J. Prediction of Satellite-Based Column CO2 Concentration by Combining Emission Inventory and LULC Information. IEEE Trans. Geosci. Remote Sens. 2020, 1–16. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
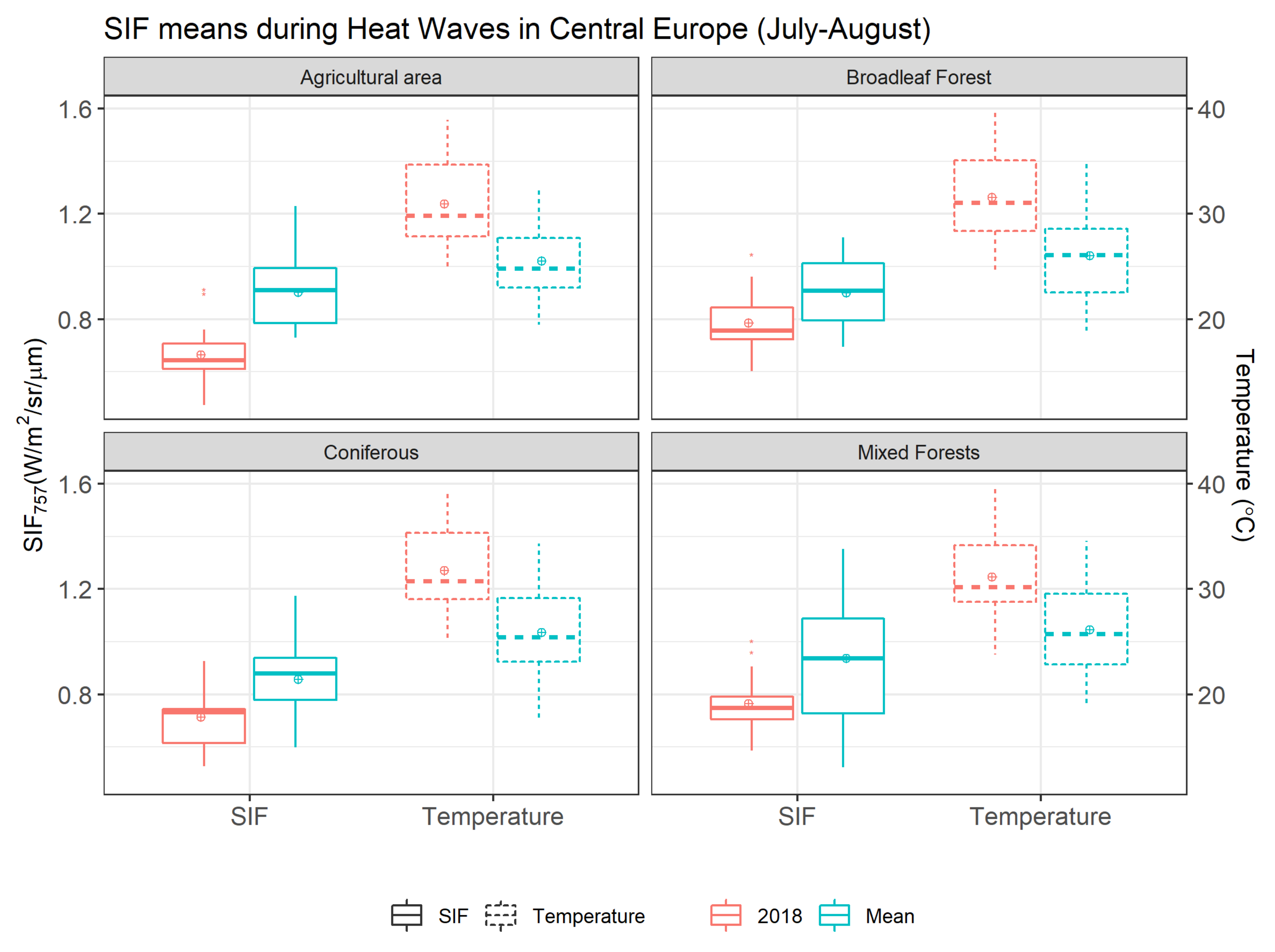{kind=link}
{kind=link}
| Area | (a) Combined Spring–Summer | (b) Spring | (c) Summer | ||||||
|---|---|---|---|---|---|---|---|---|---|
| SIF_2018 | SIF_Mean | SIF-Diff | SIF_2018 | SIF_Mean | SIF-Diff | SIF_2018 | SIF_Mean | SIF-Diff | |
| Europe | 0.642 | 0.670 | −0.028 | 0.523 | 0.534 | −0.011 | 0.766 | 0.820 | −0.054 * |
| Drought area | 0.558 | 0.596 | −0.038 | 0.407 | 0.388 | 0.019 | 0.711 | 0.814 | −0.103 * |
| Non-drought area | 0.684 | 0.660 | 0.024 | 0.601 | 0.560 | 0.041 | 0.766 | 0.767 | −0.001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shekhar, A.; Chen, J.; Bhattacharjee, S.; Buras, A.; Castro, A.O.; Zang, C.S.; Rammig, A. Capturing the Impact of the 2018 European Drought and Heat across Different Vegetation Types Using OCO-2 Solar-Induced Fluorescence. Remote Sens. 2020, 12, 3249. https://doi.org/10.3390/rs12193249
Shekhar A, Chen J, Bhattacharjee S, Buras A, Castro AO, Zang CS, Rammig A. Capturing the Impact of the 2018 European Drought and Heat across Different Vegetation Types Using OCO-2 Solar-Induced Fluorescence. Remote Sensing. 2020; 12(19):3249. https://doi.org/10.3390/rs12193249
Chicago/Turabian StyleShekhar, Ankit, Jia Chen, Shrutilipi Bhattacharjee, Allan Buras, Antony Oswaldo Castro, Christian S. Zang, and Anja Rammig. 2020. "Capturing the Impact of the 2018 European Drought and Heat across Different Vegetation Types Using OCO-2 Solar-Induced Fluorescence" Remote Sensing 12, no. 19: 3249. https://doi.org/10.3390/rs12193249
APA StyleShekhar, A., Chen, J., Bhattacharjee, S., Buras, A., Castro, A. O., Zang, C. S., & Rammig, A. (2020). Capturing the Impact of the 2018 European Drought and Heat across Different Vegetation Types Using OCO-2 Solar-Induced Fluorescence. Remote Sensing, 12(19), 3249. https://doi.org/10.3390/rs12193249