Assessment of Canopy Porosity in Avocado Trees as a Surrogate for Restricted Transpiration Emanating from Phytophthora Root Rot

 ,
,  ,
,
Abstract

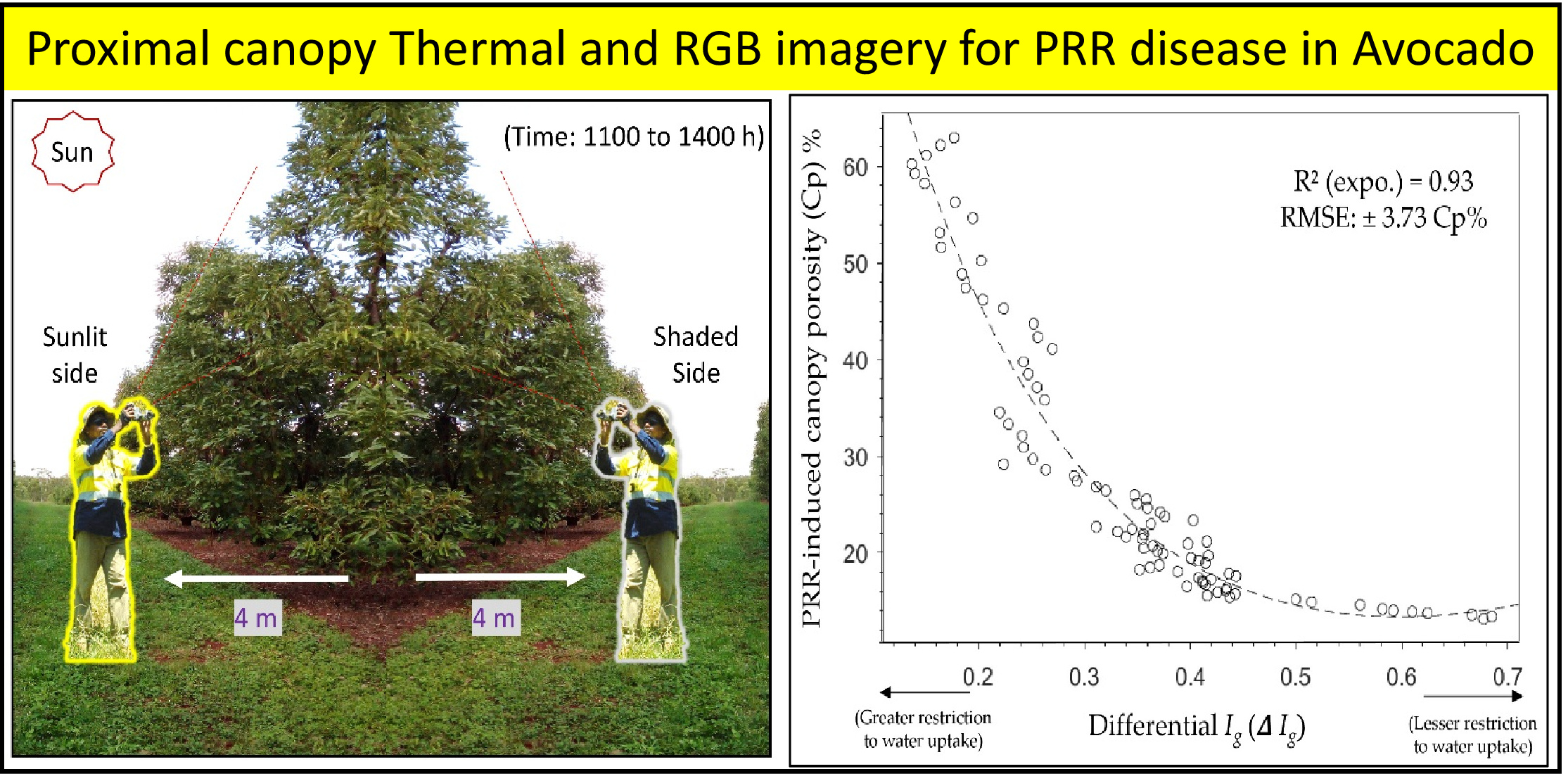{kind=link}
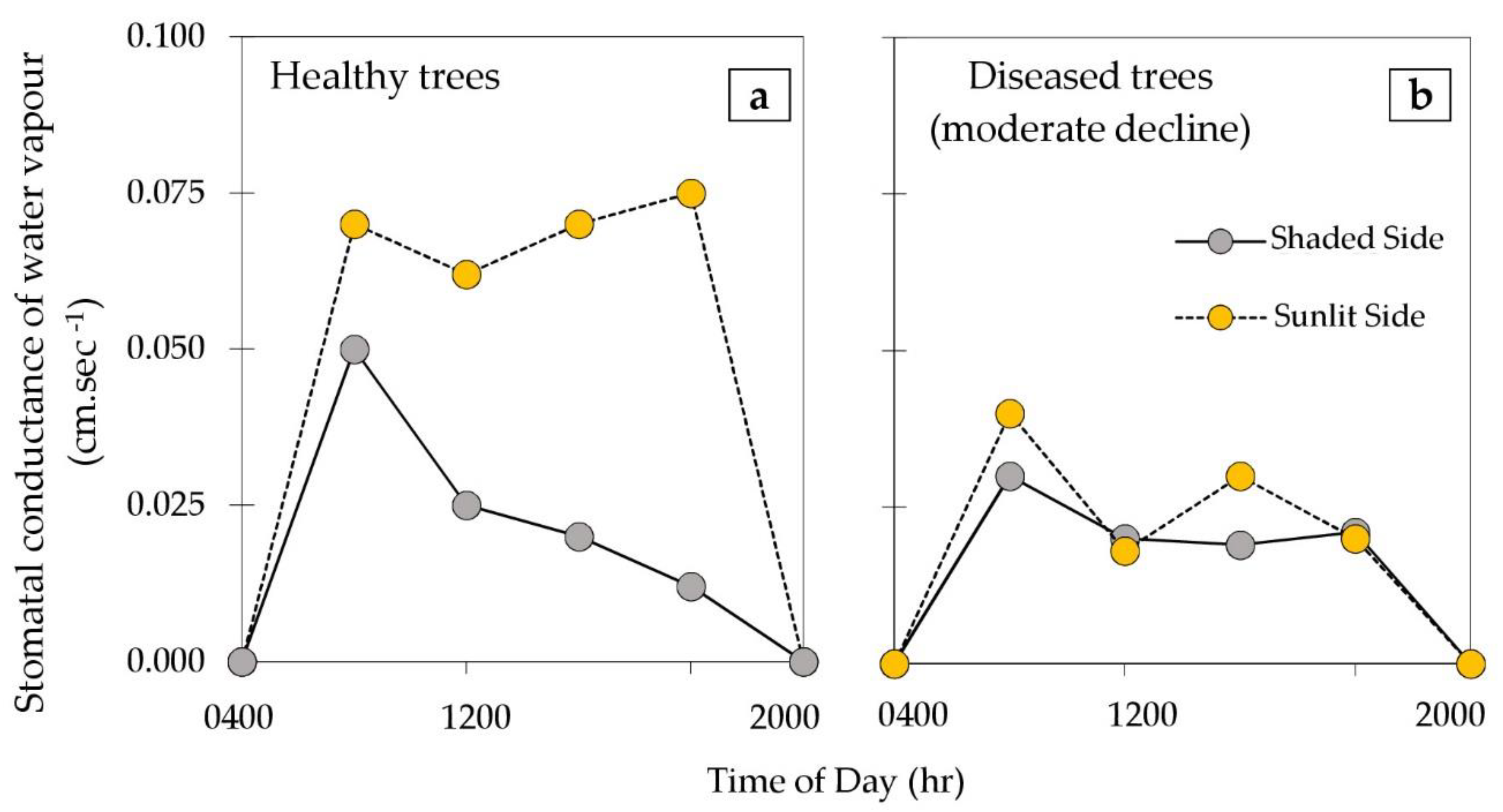{kind=link}
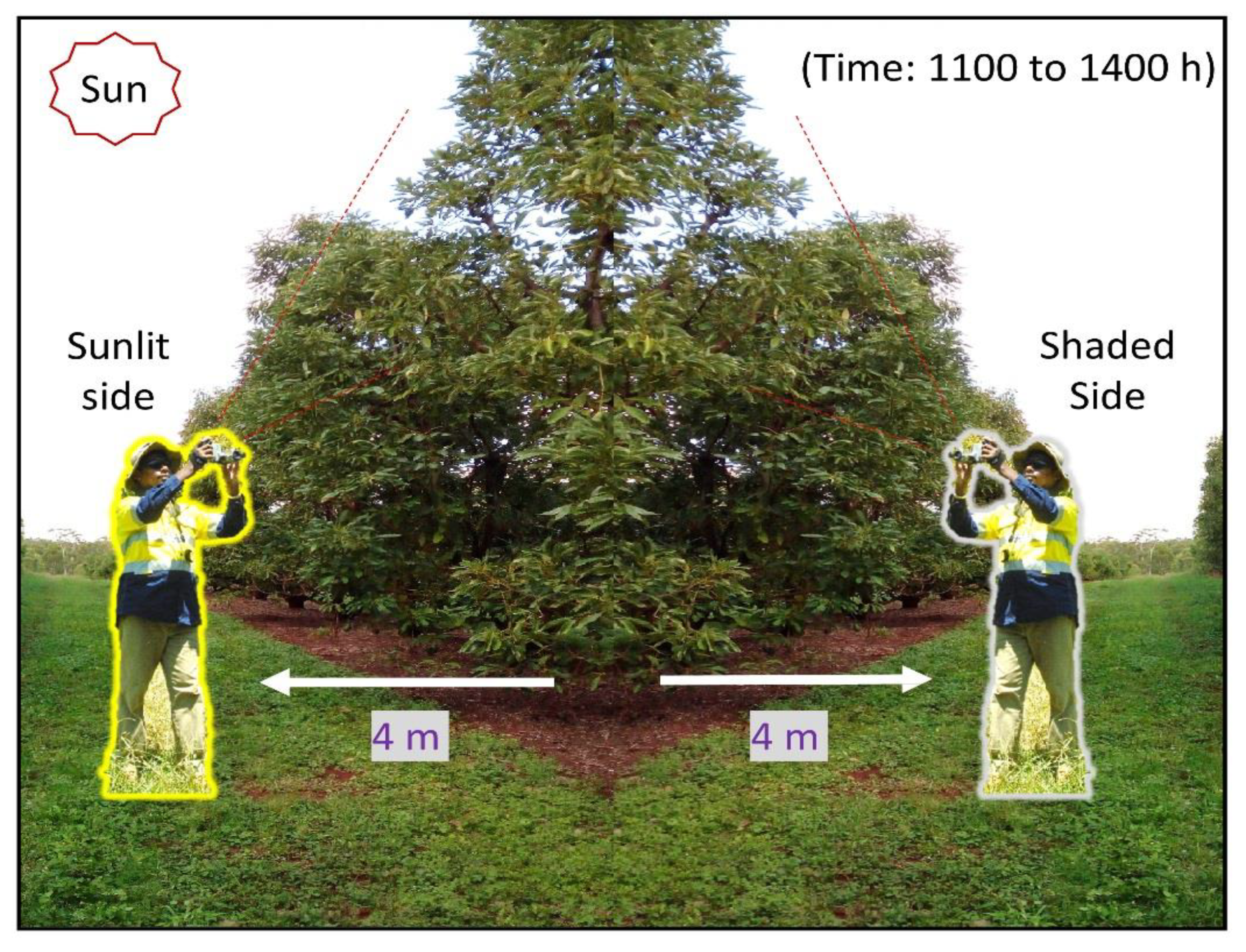{kind=link}
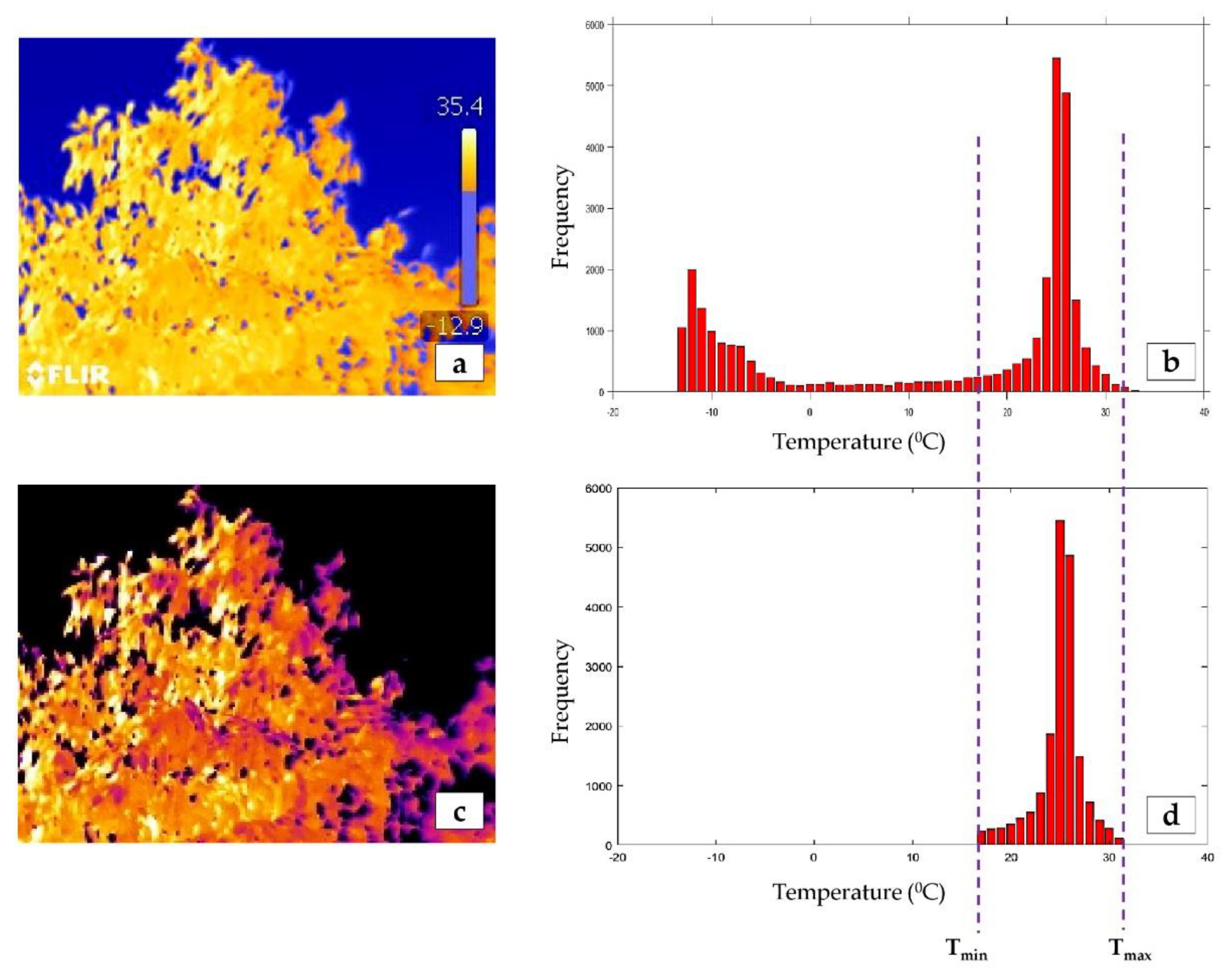{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Tree Sampling
2.2. Thermal Image Acquisition to Establish Lead Indicators of PRR-Induced Decline
2.3. Thermal Image Analysis
2.3.1. Generating Temperature Data Files
2.3.2. Thresholding Canopy Pixels
2.4. Deriving Thermal Indicators of PRR-Induced Canopy Decline
2.5. RGB Image Acquisition and Calculating Canopy Porosity
2.6. Statistical Analysis
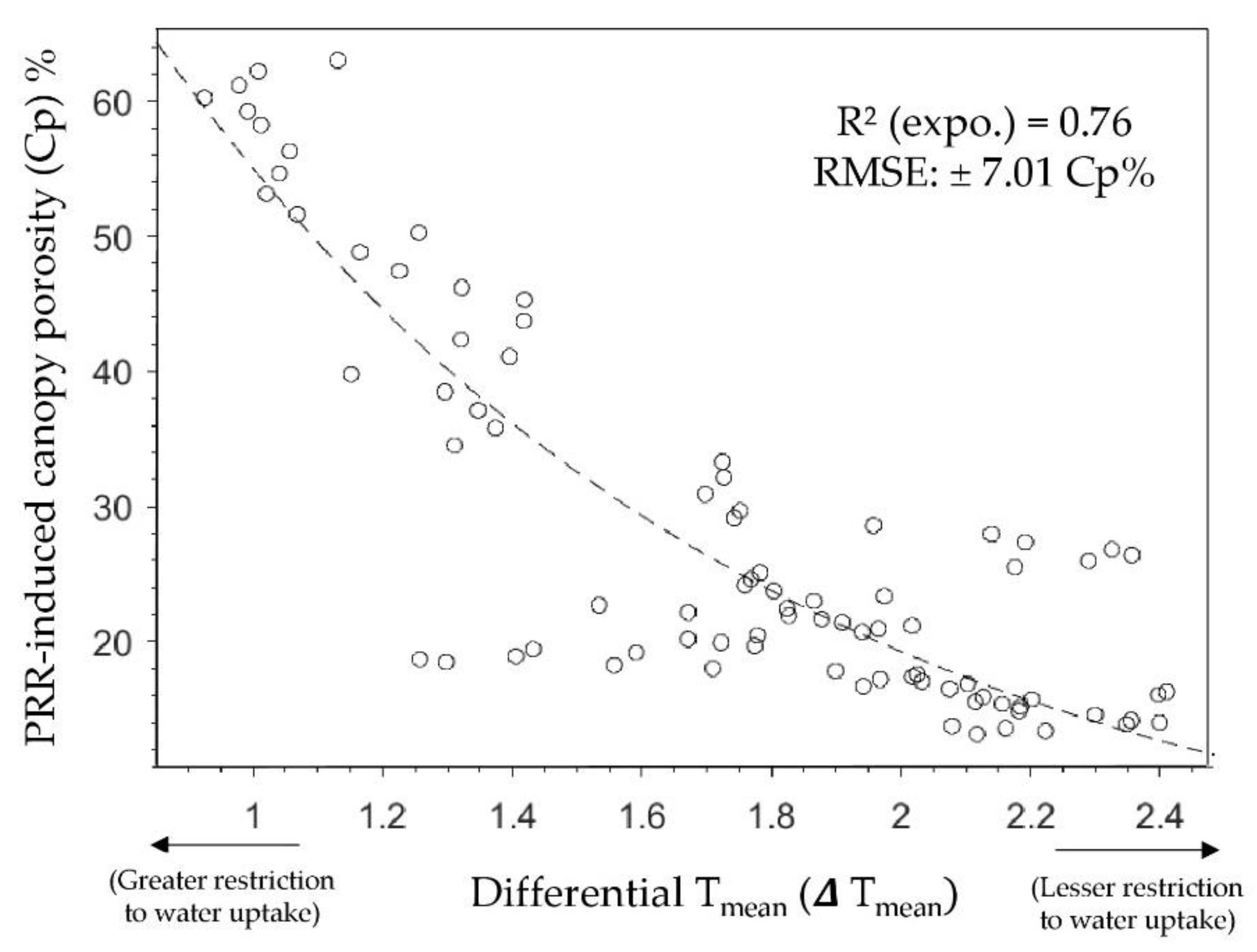
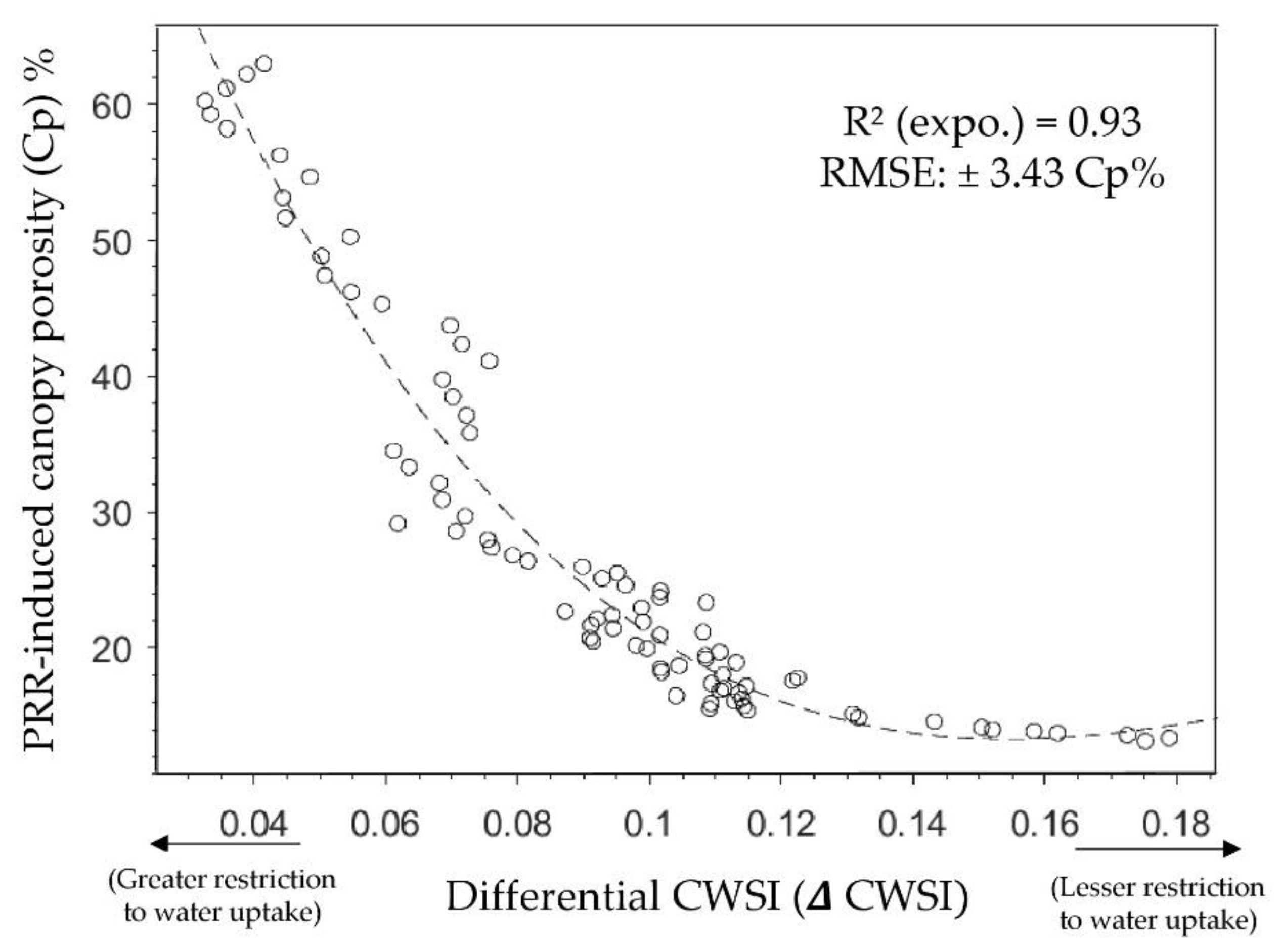
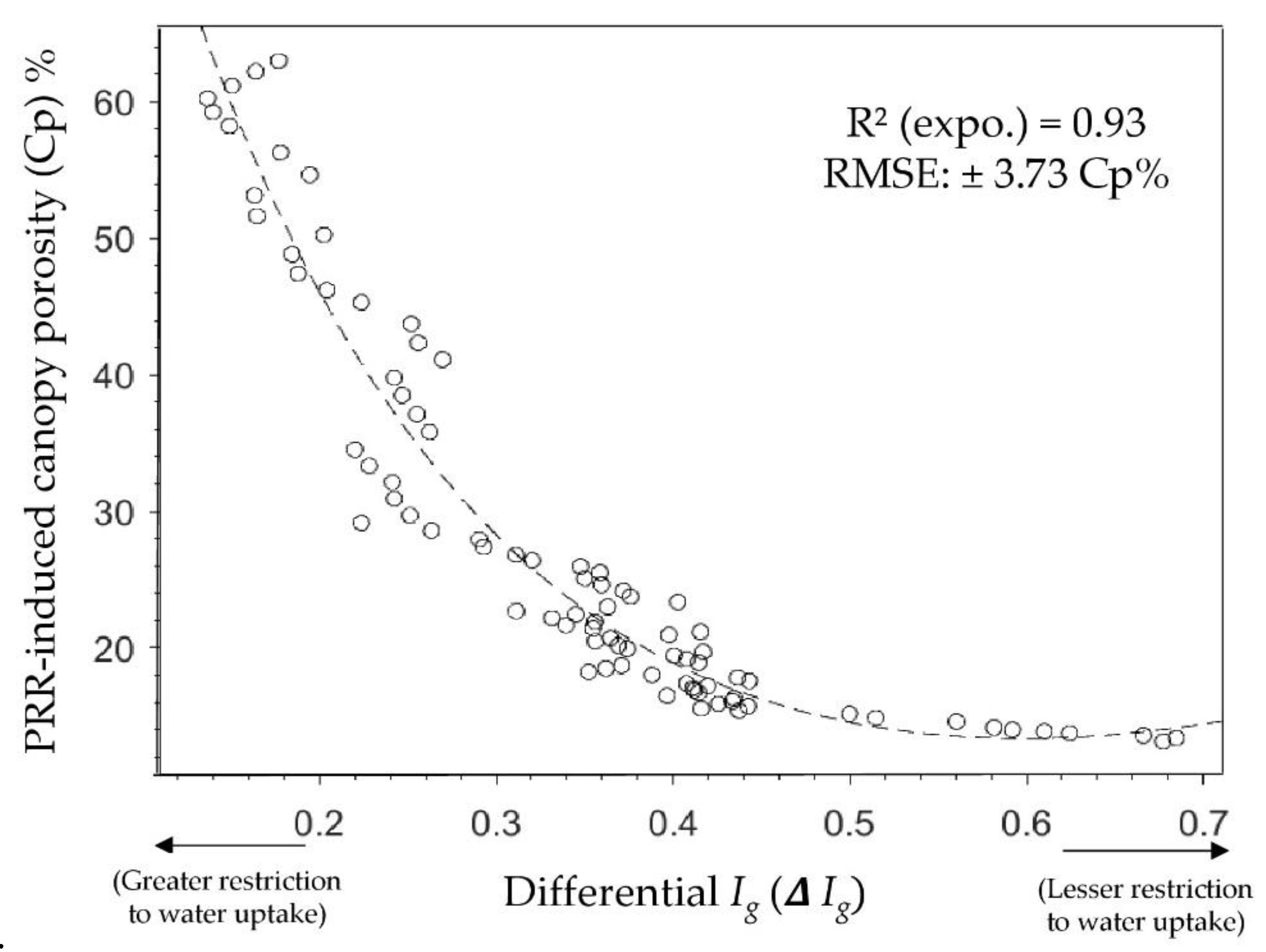
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Marais, L.J.; Menge, J.A.; Bender, G.S.; Faber, B. Phytophthora root rot. AvoResearch 2001, 2, 3–6. [Google Scholar]
- Whiley, A. The Australian avocado industry. In Proceedings of the First World Avocado Congress; South African Avocado Growers’ Association: Tzaneen, South Africa, 1987; pp. 28–30. [Google Scholar]
- Avocados Australia Australian Avocados-Facts at a Glance. 2018. Available online: https://www.avocado.org.au/wp-content/uploads/2018/10/2017-18_AAL-Facts-at-a-glance_FINAL.pdf (accessed on 04 March 2019).
- Howard, K.; Burgess, T.; Dunstan, B.; Hardy, G. Talking Avocado; Avocados Australia Limited: Rocklea, Australia, 2019; pp. 49–53. [Google Scholar]
- Dann, E.K.; Ploetz, R.C.; Coates, L.M.; Pegg, K.G. Foliar, fruit and soil-borne diseases. In The Avocado: Botany, Production and Uses; Schaffer, B., Whiley, A.W., Wolstenholme, B.N.C.N., Eds.; CABI: Cambridge, UK, 2012; pp. 397–398. ISBN 978-1-84593-701-0. [Google Scholar]
- Coffey, M.D. Phytophthora root rot of avocado: An integrated approach to control in California. Plant Dis. 1987, 71, 1046–1052. [Google Scholar]
- Darvas, J.M.; Toerien, J.C.; Milne, D.L. Control of avocado root rot by trunk injection with Phosethyl-Al. Plant Dis. 1984, 68, 691. [Google Scholar] [CrossRef]
- Mahlein, A.-K.; Oerke, E.-C.; Steiner, U.; Dehne, H.-W. Recent advances in sensing plant diseases for precision crop protection. Eur. J. Plant Pathol. 2012, 133, 197–209. [Google Scholar] [CrossRef]
- Barton, C.V.M. Advances in remote sensing of plant stress. Plant Soil 2012, 354, 41–44. [Google Scholar] [CrossRef]
- Diago, M.P.; Aquino, A.; Millan, B.; Palacios, F.; Tardaguila, J. On-the-go assessment of vineyard canopy porosity, bunch and leaf exposure by image analysis. Aust. J. Grape Wine Res. 2019, 25, 363–374. [Google Scholar] [CrossRef]
- Goodenough, A.E.; Goodenough, A.S. Development of a rapid and precise method of digital image analysis to quantify canopy density and structural complexity. ISRN Ecol. 2012, 2012, 1–11. [Google Scholar] [CrossRef]
- Calderón, R.; Navas-Cortés, J.; Zarco-Tejada, P. Early detection and quantification of verticillium wilt in olive using hyperspectral and thermal imagery over large areas. Remote Sens. 2015, 7, 5584–5610. [Google Scholar] [CrossRef]
- Granum, E.; Pérez-Bueno, M.L.; Calderón, C.E.; Ramos, C.; de Vicente, A.; Cazorla, F.M.; Barón, M. Metabolic responses of avocado plants to stress induced by Rosellinia necatrix analysed by fluorescence and thermal imaging. Eur. J. Plant Pathol. 2015, 142, 625–632. [Google Scholar] [CrossRef]
- Franke, J.; Menz, G. Multi-temporal wheat disease detection by multi-spectral remote sensing. Precis. Agric. 2007, 8, 161–172. [Google Scholar] [CrossRef]
- Moller, M.; Alchanatis, V.; Cohen, Y.; Meron, M.; Tsipris, J.; Naor, A.; Ostrovsky, V.; Sprintsin, M.; Cohen, S. Use of thermal and visible imagery for estimating crop water status of irrigated grapevine. J. Exp. Bot. 2006, 58, 827–838. [Google Scholar] [CrossRef] [PubMed]
- Robson, A. Remote sensing application for the determination of yeild, maturity and aflatoxin contamination in Peanut, Thesis submitted to Centre for Remote Sensing & Spatial Information Science School of Geography, Planning & Architecture. Ph.D. Thesis, University of Queensland, Brisbane, Australia, 2007. [Google Scholar]
- Abdullah, H.; Darvishzadeh, R.; Skidmore, A.; Heurich, M. Sensitivity of Landsat-8 OLI and TIRS Data to Foliar Properties of Early Stage Bark Beetle (Ips typographus, L.) Infestation. Remote Sens. 2019, 11, 398. [Google Scholar] [CrossRef]
- Pérez-Bueno, M.L.; Pineda, M.; Vida, C.; Fernández-Ortuño, D.; Torés, J.A.; de Vicente, A.; Cazorla, F.M.; Barón, M. Detection of white root rot in avocado trees by remote sensing. Plant Dis. 2019, 103, 1119–1125. [Google Scholar] [CrossRef] [PubMed]
- Salgadoe, S.; Robson, A.; Lamb, D.; Dann, E.; Searle, C. Quantifying the severity of phytophthora root rot disease in avocado trees using image analysis. Remote Sens. 2018, 10, 226. [Google Scholar] [CrossRef]
- Oerke, E.-C. Thermal imaging of cucumber leaves affected by downy mildew and environmental conditions. J. Exp. Bot. 2006, 57, 2121–2132. [Google Scholar] [CrossRef]
- Newton, A.C. Plant Pathology and Plant Pathogens, 3rd ed.; LUCAS, J.A., Ed.; Blackwell Science: Oxford, UK, 1998; p. 274. ISBN 0 632 03046 1. [Google Scholar]
- Lindenthal, M.; Steiner, U.; Dehne, H.-W.; Oerke, E.-C. Effect of downy mildew development on transpiration of cucumber leaves visualized by digital infrared thermography. Phytopathology 2005, 95, 233–240. [Google Scholar] [CrossRef]
- Sterne, R.E.; Kaufmann, M.R.; Zentmyer, G.A. Effect of Phytophthora root rot on water relations of avocado: Interpretation with a water transport model. Phytopathology 1978, 68, 595–602. [Google Scholar] [CrossRef]
- van den Honert, T.H. Water transport in plants as a catenary process. Discuss. Faraday Soc. 1948, 3, 146. [Google Scholar] [CrossRef]
- Clemenz, C.; Fleischmann, F.; Haberle, K.-H.; Matyssek, R.; Osswald, W. Photosynthetic and leaf water potential responses of Alnus glutinosa saplings to stem-base inoculaton with Phytophthora alni subsp. alni. Tree Physiol. 2008, 28, 1703–1711. [Google Scholar] [CrossRef]
- Guerfel, M.; Baccouri, O.; Boujnah, D.; Chaïbi, W.; Zarrouk, M. Impacts of water stress on gas exchange, water relations, chlorophyll content and leaf structure in the two main Tunisian olive (Olea europaea L.) cultivars. Sci. Hortic. 2009, 119, 257–263. [Google Scholar] [CrossRef]
- Barradas, V.L.; Nicolás, E.; Torrecillas, A.; Alarcón, J.J. Transpiration and canopy conductance in young apricot (Prunus armenica L.) trees subjected to different PAR levels and water stress. Agric. Water Manag. 2005, 77, 323–333. [Google Scholar] [CrossRef]
- Gómez-Bellot, M.J.; Nortes, P.A.; Sánchez-Blanco, M.J.; Ortuño, M.F. Sensitivity of thermal imaging and infrared thermometry to detect water status changes in Euonymus japonica plants irrigated with saline reclaimed water. Biosyst. Eng. 2015, 133, 21–32. [Google Scholar] [CrossRef]
- Chaerle, L.; Van Der Straeten, D. Imaging techniques and the early detection of plant stress. Trends Plant Sci. 2000, 5, 495–501. [Google Scholar] [CrossRef]
- Glenn, D.M. Infrared and chlorophyll fluorescence imaging methods for stress evaluation. HortScience 2012, 47, 697–698. [Google Scholar] [CrossRef]
- Costa, J.M.; Grant, O.M.; Chaves, M.M. Thermography to explore plant–environment interactions. J. Exp. Bot. 2013, 64, 3937–3949. [Google Scholar] [CrossRef]
- Jones, H.G.; Serraj, R.; Loveys, B.R.; Xiong, L.; Wheaton, A.; Price, A.H. Thermal infrared imaging of crop canopies for the remote diagnosis and quantification of plant responses to water stress in the field. Funct. Plant Biol. 2009, 36, 978. [Google Scholar] [CrossRef]
- Mahlein, A.-K. Plant disease detection by imaging sensors–parallels and specific demands for precision agriculture and plant phenotyping. Plant Dis. 2016, 100, 241–251. [Google Scholar] [CrossRef]
- Stoll, M.; Schultz, H.R.; Baecker, G.; Berkelmann-Loehnertz, B. Early pathogen detection under different water status and the assessment of spray application in vineyards through the use of thermal imagery. Precis. Agric. 2008, 9, 407–417. [Google Scholar] [CrossRef]
- Wang, X.; Yang, W.; Wheaton, A.; Cooley, N.; Moran, B. Automated canopy temperature estimation via infrared thermography: A first step towards automated plant water stress monitoring. Comput. Electron. Agric. 2010, 73, 74–83. [Google Scholar] [CrossRef]
- Fuentes, S.; De Bei, R.; Pech, J.; Tyerman, S. Computational water stress indices obtained from thermal image analysis of grapevine canopies. Irrig. Sci. 2012, 30, 523–536. [Google Scholar] [CrossRef]
- García-Tejero, I.F.; Hernández, A.; Padilla-Díaz, C.M.; Diaz-Espejo, A.; Fernández, J. Assessing plant water status in a hedgerow olive orchard from thermography at plant level. Agric. Water Manag. 2017, 188, 50–60. [Google Scholar] [CrossRef]
- García-Tejero, I.; Ortega-Arévalo, C.; Iglesias-Contreras, M.; Moreno, J.; Souza, L.; Tavira, S.; Durán-Zuazo, V. Assessing the Crop-Water Status in Almond (Prunus dulcis Mill.) Trees via Thermal Imaging Camera Connected to Smartphone. Sensors 2018, 18, 1050. [Google Scholar] [CrossRef] [PubMed]
- Sepúlveda-Reyes, D.; Ingram, B.; Bardeen, M.; Zúñiga, M.; Ortega-Farías, S.; Poblete-Echeverría, C. Selecting canopy zones and thresholding approaches to assess grapevine water status by using aerial and ground-based thermal imaging. Remote Sens. 2016, 8, 822. [Google Scholar] [CrossRef]
- Sullivan, D.G.; Fulton, J.P.; Shaw, J.N.; Bland, G. Evaluating the sensitivity of an unmanned thermal infrared aerial system to detect water stress in a cotton canopy. Trans. ASABE 2007, 50, 1963–1969. [Google Scholar] [CrossRef]
- Pour, T.; Miřijovský, J.; Purket, T. Airborne thermal remote sensing: The case of the city of Olomouc, Czech Republic. Eur. J. Remote Sens. 2019, 52, 209–218. [Google Scholar] [CrossRef]
- Irani Rahaghi, A.; Lemmin, U.; Sage, D.; Barry, D.A. Achieving high-resolution thermal imagery in low-contrast lake surface waters by aerial remote sensing and image registration. Remote Sens. Environ. 2019, 221, 773–783. [Google Scholar] [CrossRef]
- Huryna, H.; Cohen, Y.; Karnieli, A.; Panov, N.; Kustas, W.P.; Agam, N. Evaluation of TsHARP utility for thermal sharpening of Sentinel-3 satellite images using Sentinel-2 visual imagery. Remote Sens. 2019, 11, 2304. [Google Scholar] [CrossRef]
- Erwin, D.C.; Ribeiro, O.K. Phytophthora cinnamomi Rands (1922) var. cinnamomi. In Phytophthora Diseases Worldwide; CN-SB741.P58 E79 1996; APS Press: Paul, MN, USA, 1996; pp. 269–279. ISBN 978-0-89054-212-5. [Google Scholar]
- Salgadoe, A.S.A. ThermalImage_Processing. Available online: https://gitlab.une.edu.au/asalgado/thermalimage_processing/tree/Master (accessed on 22 February 2019).
- Salgadoe, A.; Robson, A.; Lamb, D.; Schneider, D. A non-reference temperature histogram method for determining Tc from ground-based thermal imagery of orchard tree canopies. Remote Sens. 2019, 11, 714. [Google Scholar] [CrossRef]
- Jones, H.; Stoll, M.; Santos, T.; de Sousa, C.; Chaves, M.M.; Grant, O.M. Use of infrared thermography for monitoring stomatal closure in the field: Application to grapevine. J. Exp. Bot. 2002, 53, 2249–2260. [Google Scholar] [CrossRef]
- Leinonen, I.; Jones, H.G. Combining thermal and visible imagery for estimating canopy temperature and identifying plant stress. J. Exp. Bot. 2004, 55, 1423–1431. [Google Scholar] [CrossRef]
- Idso, S.B.; Jackson, R.D.; Pinter, P.J.; Reginato, R.J.; Hatfield, J.L. Normalizing the stress-degree-day parameter for environmental variability. Agric. Meteorol. 1981, 24, 45–55. [Google Scholar] [CrossRef]
- Jones, H.G. Use of infrared thermometry for estimation of stomatal conductance as a possible aid to irrigation scheduling. Agric. For. Meteorol. 1999, 95, 139–149. [Google Scholar] [CrossRef]
- Gutiérrez, S.; Diago, M.P.; Fernández-Novales, J.; Tardaguila, J. Vineyard water status assessment using on-the-go thermal imaging and machine learning. PLoS ONE 2018, 13, e0192037. [Google Scholar] [CrossRef] [PubMed]
- Salgadoe, A.S.A. CanopyPorosity_Processing. Available online: https://gitlab.une.edu.au/asalgado/canopyporosity (accessed on 10 October 2019).
- Zentmyer, G.A. Avocado diseases. Trop. Pest Manag. 1984, 30, 388–400. [Google Scholar] [CrossRef]
- Camino, C.; Zarco-Tejada, P.; Gonzalez-Dugo, V. Effects of heterogeneity within tree crowns on airborne-quantified SIF and the CWSI as indicators of water stress in the context of precision agriculture. Remote Sens. 2018, 10, 604. [Google Scholar] [CrossRef]
- Gonzalez-Dugo, V.; Zarco-Tejada, P.; Berni, J.A.J.; Suárez, L.; Goldhamer, D.; Fereres, E. Almond tree canopy temperature reveals intra-crown variability that is water stress-dependent. Agric. For. Meteorol. 2012, 154, 156–165. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salgadoe, A.S.A.; Robson, A.J.; Lamb, D.W.; Dann, E.K. Assessment of Canopy Porosity in Avocado Trees as a Surrogate for Restricted Transpiration Emanating from Phytophthora Root Rot. Remote Sens. 2019, 11, 2972. https://doi.org/10.3390/rs11242972
Salgadoe ASA, Robson AJ, Lamb DW, Dann EK. Assessment of Canopy Porosity in Avocado Trees as a Surrogate for Restricted Transpiration Emanating from Phytophthora Root Rot. Remote Sensing. 2019; 11(24):2972. https://doi.org/10.3390/rs11242972
Chicago/Turabian StyleSalgadoe, Arachchige Surantha Ashan, Andrew James Robson, David William Lamb, and Elizabeth Kathryn Dann. 2019. "Assessment of Canopy Porosity in Avocado Trees as a Surrogate for Restricted Transpiration Emanating from Phytophthora Root Rot" Remote Sensing 11, no. 24: 2972. https://doi.org/10.3390/rs11242972
APA StyleSalgadoe, A. S. A., Robson, A. J., Lamb, D. W., & Dann, E. K. (2019). Assessment of Canopy Porosity in Avocado Trees as a Surrogate for Restricted Transpiration Emanating from Phytophthora Root Rot. Remote Sensing, 11(24), 2972. https://doi.org/10.3390/rs11242972